
Abstract
Proteases are ubiquitous enzymes, having significant physiological roles in both synthesis and degradation. The use of microbial proteases in food fermentation is an age-old process, which is today being successfully employed in other industries with the advent of ‘omics’ era and innovations in genetic and protein engineering approaches. Proteases have found application in industries besides food, like leather, textiles, detergent, waste management, agriculture, animal husbandry, cosmetics, and pharmaceutics. With the rising demands and applications, researchers are exploring various approaches to discover, redesign, or artificially synthesize enzymes with better applicability in the industrial processes. These enzymes offer a sustainable and environmentally safer option, besides possessing economic and commercial value. Various bacterial and fungal proteases are already holding a commercially pivotal role in the industry. The current review summarizes the characteristics and types of proteases, microbial source, their current and prospective applications in various industries, and future challenges. Promoting these biocatalysts will prove significant in betterment of the modern world.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Proteases represent a broad group of enzymes which break down or hydrolyze proteins or peptides. The proteases act on the peptide bonds joining the adjacent amino acid residues in a protein molecule and cleave them leading to formation of shorter peptides and amino acids (Razzaq et al. 2019). These hydrolytic enzymes are ubiquitous in nature and have been found in all living forms encompassing the eukaryotes like plants, animals, fungi, protists as well as the prokaryotic domains of bacteria and archaea. Even several viruses are also known to encode their own proteases (Bernardo et al. 2018).
As per the Enzyme Commission classification, proteases are placed in the class 3(hydrolases), sub-class 4 with each proteolytic enzyme assigned unique number as EC 3.4.x.x (Contesini et al. 2017). These enzymes have been categorized on the basis of various parameters like the site of action, the type of substrate, active pH range, mechanism of action involving particular amino acid present in the active site (Guleria et al. 2016a, b). Depending on the site of action, broadly these enzymes can be classified as endopeptidase and exopeptidase. The former tend to hydrolyze non-terminal peptide bonds, leading to formation of shorter peptides, while the latter act on peptide bonds located at the termini of the substrate. Also, depending on the termini on which a particular exopeptidase acts preferentially, exopeptidases are further categorized into aminopeptidases or carboxypeptidases depending on whether they act on the N-terminal or C-terminal respectively (Naveed et al. 2021). The exopeptidases release dipeptides or tripeptides or amino acids and correspondingly shortened peptides.
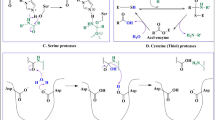
Another approach of protease categorization is based on the presence of specific amino acid residue(s) in the active site and mechanism of action. Based on this approach, the major classes of protease are serine proteases, cysteine protease, threonine proteases, glutamic proteases, asparagines proteases, aspartic proteases, mixed proteases, etc. (Contesini et al. 2017). The MEROPS database (https://www.ebi.ac.uk/merops/) is a resource providing comprehensive details about the different types of proteases. The enzymes are categorized on the primarily on the basis of their phylogenetic relationships and mechanism of action in this database (Rawlings et al. 2018). Similarly, the proteolytic enzymes have also been sorted into alkaline proteases, acidic proteases and neutral proteases on the basis of their optimal pH range (Tavano et al. 2018).
The proteases play a very significant role in the physiology and metabolism of all living organisms. Apart from the obvious role in the digestion of proteins and peptides, these enzymes also play significant role in the regulation of a vast array of physiological processes by controlling various stages involved in the protein synthesis, protein activation–inactivation, signaling, protein turnover as well as gene expression (Bond 2019). Protease and protease inhibitors account for more than 2% of the human genes. Not only do the proteases play important role in the normal functioning of the body and maintenance of homeostasis, but also these have significant role in infections, immunity, inflammation and disease development (Patel et al. 2018). For example, various common characteristic features of cancer like uncontrolled growth, angiogenesis, metastasis, immune evasion have been associated with various aberrant protease activities. On the other hand, proteases have also been found to play crucial role in tumor suppression (Dudani et al. 2018). Therefore, proteases are being utilized for prognostic, diagnostic as well as therapeutic purpose in the field of cancer management (Dudani et al. 2018). Owing to the crucial role of proteases in the life cycle of all the organisms including the infectious agents, the proteolytic enzymes offer lucrative avenue for drug discovery as is evident in case of protease inhibitors like ritonavir which is widely used for controlling Human Immunodeficiency Virus (HIV) infections (Tigabu et al. 2020). Recently, Jo et al. (2020) screened a flavinoid library for assessing the potential antiviral activities of the flavinoids which inhibited the 3C-like protease of coronaviruses and reported promising results in case of Herbacetin, rhoifolin and pectolinarin. Apart from this, there are many other applications of proteases in the field of medicine like control of cardiovascular diseases, digestive disorders, inflammatory diseases as well as promotion of tissue repair in cases of burns, fractures, accidental or surgical trauma, etc. (Kumar and Jain 2018; Bond 2019). Hence, both proteases and protease inhibitors play a crucial role in the field of medicine (Agbowuro et al. 2018).
The proteases also find a very significant role in various industries. The classical example of industrial application of proteases is for cleaning purposes like detergent additive, contact lens cleaning solution component, etc. (Salwan and Sharma 2019; Singh and Bajaj 2017; Lam et al. 2018). The textile industry makes use of proteases for a variety of purposes ranging from degumming of silk, biopolishing of wool in a very much sustainable and eco-friendly approach (Chatha et al. 2017; Mamo and Assefa 2018). Similarly, leather industry is also reducing the utilization of chemicals and laying more and more emphasis on the application of proteolytic enzymes for carrying out different steps of leather processing (Fang et al. 2017). A lot of keratinous wastes are generated by poultries, slaughter houses, etc., which are difficult to manage and cause a lot of problems like soil pollution, water pollution, aesthetic problems, clogging of drains, spreading of diseases, etc. (Kamarudin et al. 2017). Another proteinaceous waste of concern is collagen, which is generated by fish processing industries, sea-food processing industries as well as slaughter houses. Improper disposal of such wastes also pose a great pollution threat apart from the direct health risk to humans and animals (due to possible transmission of pathogenic microbes). Proteases, especially the keratinases and collagenases are also playing pivotal role in the arena of waste management and pollution control by breaking down these problematic components (Bhagwat and Dange 2018; Razzaq et al. 2019; Yusuf et al. 2019). The keratinases (proteases, which are capable of degrading keratin) have been used for degradation of keratin in the waste materials, thus hel** in the waste management. Also, the keratinase treated residues may be used as animal feeds and nitrogenous fertilizers (Kumawat et al. 2018). Collagenase can be used for extraction of collagen from the fish and animal carcasses, which in turn will not only lead to lower waste generation, but also help in reclaiming collagen (Pal and Suresh 2016). The collagen can be used by various food, pharmaceutical and cosmetic industries.
Apart from these, proteases possess a plethora of applications in case of the food industries like manufacture of protein supplements, infant food, debittering of protein hydrolysates, meat tenderization, production of various types of fermented foods and beverages like cheese, beer, removal of haze from beverages, etc. (Banerjee and Ray 2017; dos Santos Aguilar and Sato 2018). The proteolytic enzymes also act as important tools in carrying out various molecular biology and genetic engineering experiments. The applications of proteases in various fields are discussed in detail in the subsequent sections.
Proteases represent a wide array of enzymes capable of acting on a variety of proteinaceous substrates. Not only do these enzymes carry out degradative activity, but also these are also capable of synthetic activity. Proteases have been utilized for synthesis of peptides for application in diverse fields like food industry, medicine, agriculture, etc. Białkowska et al. (2017) have specifically reviewed the application of proteases from thermophilic, halophilic and psychrophlic organisms for peptide synthesis. Zanutto-Elgui et al. (2018) reported production of milk peptides with antimicrobial and antioxidant properties with the help of fungal proteases from Aspergillus oryzae and A. flavipes.
Scope of the review
This review aims to provide state of the art information related to the proteases with emphasis on microbial origin proteases. First part of this review highlights the various categories of proteases, second showed various strategies of production and purification of the enzyme. The subsequent section presents a comprehensive overview about some of the pertinent applications of proteases. It is then followed by an in depth analysis about the concept of protein engineering and its application in the improvement of properties of proteases. The review entails the compilation and analysis of up to date information about the microbial proteases.
Microbial proteases
Proteases are among one of the oldest enzymes to be discovered and studied, with several animal origin proteases like pepsin, trypsin and plant origin proteases like papain being discovered and characterized as early as the 1800s or the earlier decades of 1900s (Bond 2019). Subsequently, the microbial proteases also came into the focus and quickly became quite popular owing to several advantages associated with them (Guleria et al. 2014; Singh et al. 2016). Microorganisms, as source of proteases, offer several inherent advantages like rapid production rates, lower investment in terms of land and time requirements, not being subjected to climatic influences (Bhatia et al. 2021). Moreover, due to the diverse environments in which the microorganisms are found, it is quite easier to screen out the microbes producing enzymes with desirable characteristics (Putatunda et al. 2019). For example, if one is interested in thermostable proteases, it is highly probable to get such enzymes from thermophilic bacteria growing in hot springs (Sahay et al. 2017). Moreover, due to shorter generation time and relatively simpler genetic make-up, the genetic manipulation of microorganisms is much easier (Ali et al. 2016). This in turn, further opens up vistas for designing ‘tailor-made’ enzymes with the desired features. The ethical issues confronted by the workers in dealing with animal origin proteases are also absent in case of microbial origin proteases (da Silva et al. 2016a, b, 2017). The capability of microbes to produce extracellular proteases further makes the production process much easier by simplifying the downstreaming (Razzaq et al. 2019). Not only this, several microbes have been grown on cheaper substrates or waste materials, with the aim of further making the process more economical (Hamza 2017; Limkar et al. 2019).
A lot of work has been done on various proteases of microbial origin, some of the recent ones are presented in Table 1. From the commercial point of view, bacterial origin proteases and fungal origin proteases have received a lot of attention and that is why many researchers have explored these enzymes. As per Gurumallesh et al. (2019), the microbial proteases form two-third of the total proteases used in the industries. The major contributors are the bacterial alkaline proteases and fungal acidic proteases, although the viral proteases are also important. Protease producing microorganisms have been isolated from a variety of environments like rhizosphere soil, slaughter house soil or floor washing, sewage, food waste, etc. (Jadhav et al. 2020; Hakim et al. 2018; Ash et al. 2018; Prajapati et al. 2017). These have also been from saline environments like the marine environments, sea sediments, hypersaline lakes, salted food, soda lakes, etc. (Maruthiah et al. 2016; Boughachiche et al. 2016; Ibrahim et al. 2019). Among the bacteria, the genus Bacillus has a very prominent place in terms of the commercial production of proteases, while the fungal sources are dominated by the genera Aspergillus, Trichoderma, Penicillium, etc. (Gurumallesh et al. 2019). Apart from these, many researchers have also explored the endophytic fungi for production of proteases for various purposes (Meshram et al. 2016; Mandal and Banerjee 2019). Meshram and Saxena (2016) reported proteolytic and fibrinolytic activity in an endophytic fungal isolate Lasiodiplodia pseudotheobromae obtained from the stem of the Aegle marmelos, and proposed its potential use for dissolution of blood clots.
Many different proteases have been identified and characterized in different viruses; however, from the commercial point of view, these have not gained much importance and mainly serve as targets of various antiviral drugs. Some of the viral origin proteases include the cysteine proteases, identified in Adenovirus, Alphavirus; serine protease reported in Hepatitis C Virus; aspartic protease found in HIV, etc. (Sharma and Gupta 2017).
Alkaline proteases
The bacterial proteases used for various commercial purposes generally belong to the alkaline proteases and neutral proteases categories. The alkaline proteases, especially the extracellular proteases, from the members of the genus Bacillus, which are usually the serine proteases have been in use in since quite long time (Pathak and Rathod 2018). The enzymes from Bacillus have drawn a lot of attention and hence a lot of information is available with respect to their activity, size, structure, etc. (Contesini et al. 2017). Two of the most important types of alkaline proteases are subtilisin Carlsberg and subtilisin novo, which are again obtained from the genus Bacillus (Contesini et al. 2017). Moreover, the strains of B. subtilis and other related members of Bacillus, used for enzyme production for commercial purposed are regarded as GRAS (generally recognized as safe) (Zhang et al. 2019). So, even now more and more reports about alkaline protease from novel strains of Bacillus sp. as well as novel proteases from already discovered strains are being published from around the globe (Guleria et al. 2016a; Hakim et al. 2018; Dorra et al. 2018; Jadhav et al. 2020; Qamar et al. 2020; Far et al. 2020). Mothe and Sultanpuram (2016) carried out the purification and characterization of serine alkaline protease from a novel B. caseinilyticus. Hammami et al. (2020) have reported the characterization of a novel H2O2-tolerant alkaline protease from B. invictae AH1. Recently, Zhou et al. (2020) carried out genetic modification of B. licheniformis 2709 which lead to 62.19% enhancement in alkaline protease production along with reduction in the formation of mucopolysaccharides and other foaming agents.
Although Bacillus genus proteases are most widely characterized and used, there are reports of several other bacteria capable of producing alkaline proteases. Alkaline protease production by Streptomyces sp. isolated from saltpan of Algeria was reported by Boughachiche et al. (2016). A novel thermostable keratinolytic alkaline protease producing Streptomyces sp. Al-Dhabi-82 was reported by Al-Dhabi et al. (2020). Similarly, Masi et al. (2017) reported alkaline protease production from Enterococcus hirae and Pseudomonas aeruginosa isolated from dairy effluents. They further carried out formation of nanocomposite of the bacterial protease with super paramagnetic nanoparticles leading to protease stability enhancement. Not only the bacteria like Bacillus, Pseudomonas, etc. have the capability of producing alkaline proteases, but several fungi have also been reported to produce these enzymes (Sharma 2019). Meshram et al (2016) reported that a metalloprotease (with 8.0 pH optima) produced by endophytic fungus Xylaria curta possessed appreciable fibrinolytic properties, and thus has the potential of being used for therapeutic purposes. da Silva et al. (2018) have carried out thermodynamic investigations on the serine alkaline protease produced by the fungus Aspergillus tamarii URM4634. Nadeem et al. (2019) reported significant alkaline protease production from Chetomium globosum.
The bacterial alkaline proteases have generally pH optima in the range of 8–12. Many of these enzymes also display higher thermostability and activity at elevated temperatures (Naveed et al. 2021). Being capable of showing activity in the alkaline pH range, the alkaline proteases have been very much in use as an additive to the detergents, for enhancement of the stain removal properties (Guleria et al. 2016a; Asha and Palaniswamy 2018). Similarly, leather processing is carried out at high pH, hence the alkaline proteases can act as suitable additives for the tanning industries (Hussain et al. 2017; Hakim et al. 2018).
Acidic proteases
As the name itself indicates, the acidic proteases tend to show their activity in the acidic pH range, mostly between pH 3.8 and 5.6, with the optimum pH being 3–4 and isoelectric point between 3 and 4.5 and belong to the group of aspartic proteases (Machado et al. 2016; Razzaq et al. 2019). The microbial acidic proteases are broadly categorized into pepsin-like and rennin-like (Razzaq et al. 2019). Unlike the alkaline proteases, which are mostly produced by the bacteria, the fungi are the primary producers of acid proteases, with the members of the genera Aspergillus, Penicillium, Endothia, Mucor, etc. being the most commonly reported ones (Mammo and Assefa 2018).
There are several reports of novel acid proteases being produced by the molds as well as some yeasts (Mandujano-González et al. 2016). An aspartic protease of Rhizomucor miehei, which can serve as an alternative enzyme for milk clotting during cheese production, was characterized by da Silva et al. (2016a, b). Souza et al. (2017) characterized an acidic protease with optimum pH of 5.0 and optimum temperature of 55 °C, from Aspergillus foetidus isolated from soil sample. Similarly, da Silva et al. (2017) characterized an aspartic protease with promising milk clotting properties from Phanerochaete chrysosporium. Sun et al. (2018) cloned the aspartic protease gene from Rhizomucor miehei into Pichia pastoris and reported better meat tenderization as compared to papain (which is a plant origin protease, commonly used for meat tenderization). Deng et al. (2018) reported that recombinant aspartic protease from Trichoderma harzianum expressed in Pichia pastoris was effective against several pathogenic fungi like Botrytis cinerea, Mucor circinelloides, Aspergillus fumigatus, Aspergillus flavus, Rhizoctonia solani, and Candida albicans. Song et al. (2020) carried out biochemical characterization of two new Aspergillus niger aspartic proteases and reported both the enzymes had 50 °C as the optimum temperature, with the pH optima of PepAb and PepAc being 2.5 and 3.0 respectively.
A few bacteria have also been reported to produce acidic proteases. Proteases from Enterococcus faecium strain 1.15 and Lactobacillus plantarum strain 1.13, isolated from Bakasam (a type of fermented meat) by Putranto et al. (2017) and (2020a), displayed rennin-like activity. Similar activity was also shown by the protease produced by L. paracasei strain 2.12, which was isolated by Putranto et al. (2020b) from goat milk.
The acidic proteases play a very significant role in the food and beverage industry. Animal origin enzymes like rennin were originally used for milk clotting during cheese production, but these are now being replaced by microbial origin proteases (Mammo and Assefa 2018). Similarly, the acid proteases are also used for preparation of protein hydrolysates as well as in bakeries for improving the characteristics of dough (Singh and Bajaj 2017). The acid proteases are also important in removal of haze from juices and beer.
Neutral proteases
Neutral proteases refer to those proteolytic enzymes which show their activity in either in neutral pH or slightly alkaline pH or slightly acidic pH, with the maximal activity within the pH range of 5–8 (Razzaq et al 2019). Like the alkaline proteases, the neutral proteases are also contributed by the genus Bacillus (Contesini et al. 2017; Razzaq et al. 2019). Many of the neutral proteases fall into the category of metalloprotease and have requirement of a divalent positively charged ion for their activity.
The neutral proteases have been utilized for various purposes including the brewing and baking industries (Razzaq et al. 2019). Aissaoui et al. (2016) characterized a neutral protease from Trichoderma harzianum and carried out hydrolysis of Scorpaena notata (fish) viscera. They reported anti-bacterial peptide production in the fish viscera hydrolysate. Ao et al. (2018) undertook the characterization of neutral protease from Aspergillus oryzae Y1 isolated from naturally fermented broad beans and reported it to be a 45 kD protein with 7.0 optimum pH and 55 °C optimum temperature. De Oliveira et al. (2020) reported about production of a relatively thermostable protease by the fungus Moorella speciosa, with 6.5 as the optimum pH. However, on immobilization on MAT540 microspheres, the optimum pH changed to 5.74. Deng et al. (2021) reported about the possible application of the recombinant neutral protease rNpI as fish feed additive to improve protein digestion and growth.
Microbial protease production technology
Microorganisms and source
The first step in the production of microbial proteases is the isolation and selection of candidate microorganisms. The microorganisms are isolated from a variety of sources/ecosystems followed by selection for the desired character. These sources/ecosystems have specific or adverse feature which reflects the characteristics of the enzyme to be produced; for example, dum** sites, hot springs, soda lake, soil samples from milk processing plants, poultry waste sites, meat waste contaminated soil, tannery waste, detergent industry, leather industry, and wood factory (Anandharaj et al. 2016; Kshetri et al. 2016; Suleiman et al. 2020; Sharma et al. 2017; Chu 2007; Kandasamy et al. 2016; Ramakodi et al. 2020). Other sources includes saltern Qingdao at China, Izmir Gulf at Turkey, composting soil, sewage sludge, vermicomposting pit soil, sea mud, marine sediments, southern ocean, and mangrove sediments (Baweja et al. 2017; Li et al. 2019; Ammasi et al. 2020). Such biobased processes would help reducing emissions. Thus, exploitation of enzymes for leather processing is beneficial in terms of improved process efficiency, highly specific enzyme-based catalysis, enhanced leather output and superior quality of leather.
Photographic industry
Silver recovery from waste photofilms is a big business. The exposed nearly 2.0 billion radiographs are taken each year, including chest X-rays, mammograms and CT scans (Cavello et al. 2015). X-ray films have approximately 5–15 g of silver per kg of film. Around 18–20% of the world’s silver needs are supplied by recycling photographic waste (Asgher et al. 2018). Since silver is linked to gelatin in the emulsion layer, it is possible to break the same and release the silver which could be used as source of secondary silver. Enzyme-based method may be developed that may not only recover silver efficiently with minimal impact on the environment (Al-Abdalall and Al-Khaldi 2016). Proteases have been reported to possess excellent gelatinolytic activity for successful recovery of silver from X-ray films. Proteases from Purpureocillium lilacinum (Cavello et al. 2015) have also shown good gelatin hydrolysing ability. Alkaline proteases produced by B. subtilis, Streptomyces avermectnus, and Conidiobolus coronatus have been successfully reported to recover silver from X-ray films, ensuring that the process is eco-friendlier over the use of chemicals. The enzyme-based silver extraction from X-ray films relies more on renewable energy resources than on fossil fuel and, thus, might offer an overall eco-safe process.
Silk degumming
Silk fabric is composed of two fibroin filaments surrounded by a cementing layer of proteinaceous sericin and this gives a rough texture to silk but after its removal silk fibres turns very soft (More et al. 2017). Proteases which can cleave the peptide bonds of sericin without destroying the fibroin may have potential for application as degumming agents (Suwannaphan et al. 2017). Traditionally harsh chemicals used in degumming process that reduce the strength of silk, shorten shelf life of silk, not eco-friendly and even harmful to the environment. Enzymes like trypsin, papain and some other bacterial proteases are used for degumming process. Plant proteases like papain and bromelain (are the effective cocoon cooking enzymes that are commonly used for the processing of cocoons whereas some bacterial protease also has been found to be very effective in hydrolysing sericin (Singh et al. 2016). Joshi and Satyanarayana (2013) reported a recombinant alkaline serine protease from B. lehensis which enhanced the softness and shine of silk fibres. Thus, the enzyme-based technology could be used effectively for silk degumming in industries as an eco-friendly alternative (More et al. 2017). These enzymes not only increases the strength of the yarn but also prevent various environmental issues; therefore, proteases are greatly used for degumming of silk and projected as a replacement of the harsh and energy demanding chemicals for treatment process. Thus, enzyme-based silk degumming is an eco-friendly approach that may potentially help reducing usage of hazardous chemicals and undesired emissions due to usage of chemicals and fossil fuels.
Biopolishing of wool
Alkaline proteases are used for the manufacture of shrink-proof wool. Wool fibres are covered in overlap** scales pointing towards fibre tips, which could be hydrolysed by protease action. However, protease treatment in general damages the wool by causing excessive loss of strength, and lowering the anti-felting ability, which in turn may lead to additional damage to the fibre interior during wool processing. Therefore, specific proteases whose action is limited to cuticle scale of wool fibre are desired. Keratinase can be used in textile processing for efficient production of shrink-resistant fibre that has improved handling properties (Lateef et al. 2015). A keratinase from Brevibacillus parabrevis CGMCC10798 was purified and characterized for its excellent potential for wool processing (Zhang et al. 2016a, b). A novel recombinant keratinase expressed in E. coli BL21 (DE3) exhibited high specificity towards some substrates including wool. The protease has the potential for application in wool processing (Su et al. 2020). Bacillus sp. G51 exhibited the highest wool-keratinolytic activity. Thus, eco-friendly treatment approaches based on enzyme cock-tail (with protease combination) may help designing the organic wool processing (Srivastava et al. 2020).
Biofilm removal
Biofilms are formed by extracellular polymeric substances (EPS) which are mainly consist mostly of polysaccharides, proteins and nucleic acids form structural matrix which help the bacteria to attach to the surface to facilitate the survival in the adverse conditions and environments (Lohse et al. 2020). Biofilms are detrimental to both human life and industrial processes due to their association with infection, pathogen contamination, biofouling, and slime formation. Harmful biofilms may cause grave economic losses due to reduced productivity, decreased product quality and loss of time and expenses for biofilm removal and impose an economic burden on companies besides contamination and biofouling. The most effective method for removing a biofilm is by the clean-in-place (CIP) method in combination with chemicals involving manual scrubbing of the affected area. But the method is impractical for larger structures where regions like joints, filters or gaskets are not easily accessible (Nahar et al. 2018). Microbial enzymes may be safer and more efficient alternatives to traditional chemical means of biofilm removal. Proteases are the most commonly used biofilm removal agents however, other hydrolases in combination with proteases, help to eliminate existing biofilms and prevent bacteria from adhering to surfaces (Lohse et al. 2020).
Food industry
The use of chemicals in the food industry is increasingly being recognized as hazardous and natural alternatives are being explored by food technologists around the globe. Use of eco-friendly microbial proteases has shown prominent potential in replacing chemical agents, while improving piquant properties of the food products. The potential of microbial proteases as biocatalysts has been employed since time immemorial in the fermentation of food items (Singh et al. 2016). It is also known that proteases of microbial origin are more stable than their counterparts from animals or plants, with better consistency and easier potential for optimization or process modification (Gurung et al. 2013). The microbial species mainly employed in production of proteases for food industry (Table 2) have been isolated from the traditional sources of fermented foods. The US Food and Drug Administration GRAS list may be referred to identify the Bacillus (Bacillus subtilis, B. licheniformis) and Aspergillus (Aspergillus niger, A. oryzae) species, whose products are labelled as ‘generally safe’.
Medical field
Proteases show promising therapeutic properties in medical field such as gauze, non-woven tissues, and ointment composition. Certain lytic enzyme deficiency syndromes are diagnosed to be aided by an oral administration of alkaline proteases (Joshi and Satyanarayana 2013). The use of this fibrinolytic enzyme suggests its future application as an anticancer drug and in thrombolytic therapy (Jaouadi et al. 2012). Slow-release dosage form preparation containing collagenases with alkaline proteases is extensively used in therapeutic applications. The hydrolysis of collagen by the enzyme liberates low molecular weight peptides without any amino acid release for therapeutic use (Suwannaphan et al. 2017). For the treatment of various diseases, such as burns, carbuncles, furuncles, and wounds, a preparation of elastoterase immobilized on bandage is used. Proteases possess distinct therapeutic properties which are helpful in develo** various drugs against fatal diseases like anticancer, antimicrobial, anti-inflammatory infections and dissolving clots class with diverse other applications (Srilakshmi et al. 2015; Kumar and Jain 2018). Nowadays proteases are useful in treatment of cystic fibrosis, sepsis, digestive disorders, retinal disorder, and many more disease. Beside this they are also help in reducing the discomfort of breast engorgement in lactating women (Banerjee and Ray 2017). Asparaginase from Escherichia coli and collagenase also play a major role in removing asparagine from the blood in forms of lymphocytic leukemia, burns and wounds, respectively (Muneer et al. 2020). Nattokinase from Bacillus subtilis is also helpful against cardiovascular disease (Lan et al. 2020). Microbial proteases also act as the therapeutic targets for protease inhibitor drugs in various disorders and conditions, including bacterial and viral diseases (Table 3).
Prion protein degradation
Prion protein or Scrapie protein (PrPScis) is a misfolded infectious form of protein and the causative agents of transmissible spongiform encephalopathies in a variety of mammals, including bovine spongiform encephalopathy in cattle. In humans, prions cause Creutzfeldt–Jakob disease, variant Creutzfeldt–Jakob disease, Gerstmann–Straussler–Scheinker syndrome, fatal familial insomnia and kuru (Silveira et al. 2005). All known prion diseases affect the structure of the brain or other neural tissue, and all are currently untreatable and universally fatal. Proteases may help degrading wrongly folded proteins. The protease (MSK103) was also effective against dried PrPSc. Thermostable keratinase from B. pumilus KS12 has potential for degradation of Sup35NM (Rajput and Gupta 2013). It maybe envisaged that potentially novel proteases may be discovered that may help develo** bio based therapeutics for prion diseases in humans and animals.
Peptide synthesis
Enzymatic peptide synthesis has several advantages, including enantioselectivity, free racemization, ecologically benign reaction conditions, over chemical synthesis that requires expensive defensive groups, solvents, or reagents (Agyei et al. 2016; Antink et al. 2019). Protease from viral, fungal, plant and animal origins has received a great deal of interest in recent years for the synthesis of many small peptides, especially dipeptides and tripeptides utilizing either a thermodynamically or a kinetically controlled bioprocess. (Kumar and Bhalla 2005; Motyan et al. 2013; Yang et al. 2020). Organic solvent-tolerant alkaline proteases from the genera Aspergillus, Bacillus, and Pseudomonas have shown promise for peptide synthesis (Song et al. 2020). Protease-produced therapeutic bioactive peptides that serve as nutraceuticals, digestive aids, antioxidants, blood pressure regulators, and foe diet and medications ( (Nongonierma and FitzGerald 2015; Nandan and Nampoothiri 2020). Computer-aided drug design, in conjunction with genetic and protein engineering techniques, can also aid in the discovery of new peptides.
Nematicidal activity
Protease can also be used as biocontrol agents against pathogenic nematodes. Traditional chemical-based methods are not only toxic to humans and cause significant environmental pollution but also lead to the emergence of nematicide resistance (Singh and Bajaj 2017; Yang et al. 2019). Nematicidal microbial proteases, on the other hand, are natural biocontrol agents that offer several advantages like being non-toxic, environmental friendly, economically sustainable, besides improving soil characteristics and plant growth.
There are several reports of proteases from non-Bacillus spp. being used as biocontrol agents against nematodes. Alkaline protease from B. lehensis has been found to be useful as a biocontrol agent for plant pathogenic nematode Meloidogyne incognita (Joshi and Satyanarayana 2013). Saccharomonospora viridis isolated from soil sample secreted extra-cellular cuticle-degrading protease that had remarkable nematotoxic activity against Panagrellus redivivus (Darwesh et al. 2019). One of the major mechanisms for nematicidal capacities was observed to be the ability of bacteria to produce proteases and/orchitinases. Thus, development of bio-based strategies for controlling and managing the pests may contribute substantively towards environmental health as this would lead to reduced application of chemical-based pesticides (Table 4).
Contact lens cleansing
Proteases are important components in biopharmaceutical products such as contact-lens enzyme cleaners and enzymatic debriders. It is well known that the deposits on contact lenses consist of proteins, lipids, and mucin as tear components. Protein deposits are natural deposits on contact lenses which are unavoidable as they are formed by the interaction of the protein in our natural tears and the contact lenses. Generally, contact lens cleaning solutions have been prepared using plant (papain) and animal (pancreatin, trypsin and chemotrypsin) proteases but most of them impart an unpleasant odor to the cleansing bath or develop an odor after a few hours of use (Liu et al. 2018). Several microbial proteases from Bacillus species, Streptomyces sp., and Aspergillus sp. were reported to clean tear films and debris of contact lenses to overcome these drawbacks and to make cleaning composition odorless and safe, i.e., not producing an allergic response or causing irritation to the eyes, bacterial proteases are gaining importance (Singh and Bajaj 2017). Several reports are available on production of proteases from bacterial cultures with Bacillus sp. as the dominating organism. Protease from Bacillus sp.158 exhibited ability for cleaning of tear films and efficiently removed the protein deposits from contact lenses (Razzaq et al. 2019). The enzyme could effectively be used to remove protein deposits from contact lenses and, thus, help increasing the transmittance of lenses. Proteases from several Bacillus sp., viz. B. subtilis, B. licheniformis, B. thermophiles and B. cereus, exhibited excellent activity against artificial tear solution and, thus, could be of importance for contact lens cleansing (Singh et al. 2016; Contesini et al. 2018).
Animal feed
Animal feeds supplemented with proteases enzymes can enhance the nutrient bioavailability of the food constituents or as an element in an animal diet formulation to help protein digestion in the gastrointestinal tract (Hejdysz et al. 2020). The influence of a supplemental dietary mixture of different proteases in granular form cysteine protease and a serine endopeptidase on the productivity variables in dairy cattle, using as a model lactating Holstein cows was studied by Park et al. (2020). The outcome of the experiment showed that supplemental protease enhances milk and meat production effectiveness and enhances parameters of nitrogen status (Yang et al. 2020). A novel serine protease, expressed in Bacillus licheniformis as feed protease, permitted a significant augmentation in the degree of the hydrolysis, solubilization, and digestibility of proteins, improving broiler performance by enhancing protein and energy digestibility (Naveed et al. 2021).
Market potential of proteases
Changing cleaning habits, higher advancements in protein engineering technologies to produce high performing products, and rapid industrialization are driving forces behind the woofing market growth for proteases, which is further fueled by rapid industrialization and growing environmental concern in food and beverages, livestock feed, soaps and detergents, pharmaceuticals, and other industries. The protease market is expected to reach USD 2.21 billion by 2021, with a CAGR of 6% from 2016 to 2021 (Protease Market 2021). The protease market is divided into three segments based on their sources: microorganisms, plants, and animals, with the microbial sector increasing at the quickest pace in terms of value due to the inability of plant and animal sources to fulfil global protease demand. Because of its high-temperature functional characteristics, the microbial protease market is expected to gain momentum due to its commercial viability to produce large revenues with extensive penetration across the textile manufacturing, detergent, food and beverage, and paper and pulp sectors (Gupta et al. 2013). For example, in 2019, DuPont Nutrition & Biosciences released OPTIMASE, an unique range of biobased enzymes for liquid detergents in medical equipment cleaning applications. Novozymes released Formea Prime, a new protease enzyme, in 2021 to enhance its Formea range of protein drink ingredients and to assist beverage formulators in overcoming the problems associated with utilizing whey protein hydrolysates. Similarly, Amunix Pharmaceuticals, Inc. raised $117 million in a Series B fundraising round to advance AMX-818, a masked and protease-activated T-cell engager that targets HER2-expressing solid tumors, into the clinic. Some of the key participants are Advanced Enzyme Technologies, Specialty Enzymes and Biotechnologies (Advanced Enzymes USA), Biocatalysts (BRAIN AG), DuPont and DSM, Enzyme Technical Associations (ETA), and others are develo** new and robust protease for various applications in food and beverages, livestock feed, soaps and detergent, pharmaceuticals, and other industries such as textile and leather, waste management, chemical industries, silver recovery, photography, and biofuel (Protease Market 2021).
Protease engineering
Although there has been tremendous increase in our knowledge of proteases in terms of their sequence and evolutionary significant relationships, as well as technologies to engineer recombinant proteins as per our convenience, still there is a lot to know when producing a protein with ideal applications in biotechnology industry. Naturally occurring proteases might not be optimal for industrial application but with progress in bioengineering technology, they can be specifically “edited” for desirable applications. Challenges being dealt by the geneticists and protein engineering scientists while designing desirable proteases are.
-
Discovering or engineering novel proteases with industrial or pharmaceutical applications.
-
Increasing the yield, output, or cost-effectiveness of the product of the enzyme.
-
Improving the characteristics of the enzyme like increasing thermal stability or widening/altering substrate specificity, optimal pH, etc., or preventing inactivation by auto-proteolysis.
Exploring the data from available plethora of biodiversity is the invaluable resource based on which the proteases or microorganisms can be engineered for the industry. For instance, metagenomics data of the enzymes isolated from extremophiles, or ‘extremozymes’, may possess unique activities ideally required for commercial applications (Bilal et al. 2018; Bruno et al. 2019). The only limitation will be that maintaining these extremophiles in laboratory conditions might reduce their cost-effectiveness (Table 5).
The methods of protein engineering that are employed for engineering novel enzymes for industrial or pharmaceutical applications may be categorized into mainly two approaches—rational redesigning and directed evolution (Baweja et al. 2016a, b; Fig. 2). Rational redesigning approach is based on the method of ‘site-directed mutagenesis’ whereas directed evolution approach is based on natural unbiased evolution through ‘random mutagenesis’. Microbial hosts, such as Escherichia coli or Bacillus sp. have been ideal tools for such metabolic engineering approaches.

Site-directed mutagenesis
Selected amino acid of a particular site is edited creating a mutated protein. It is preferred for those proteins whose 3-D structures and functional characteristics are well established. The evaluation of the mutant protein is quicker since the variants produced are few. Site-directed mutagenesis is done generally to impart the proteins with desirable properties like improved catalysis or activity, stability like thermostability, substrate specificity, solubility, expression, etc. while also being utilized for immobilization of enzymes for the targeted industrial application (Baweja et al. 2016a, b; Chen et al. 2018a, b).
Random mutagenesis
The technique is based on introducing random mutations in a gene. The mutated protein products are produced within weeks in the form a library of mutants and require high throughput screening strategies for selecting the desired variant. The quality of library decides the engineered protein formed. This technique may also be employed for those proteins whose structural and functional characterization is not fully known. This is an indiscriminate method of mutagenesis and involves various techniques summarized in Fig. 2. Error prone PCR is generally the technique employed for random mutagenesis. Examples of applications include enhanced high/low pH tolerance, thermostability, substrate or product specificity, and so on (Baweja et al. 2016a, b).
The selection of method is based on the desirable protein product and may involve more than one method, including combination of both site-directed and random mutagenesis techniques (Chen et al. 2018a, b; Weng et al. 2015).
Future prospects of proteases
Proteases are a unique class of enzymes that bear both degradative and synthetic potential, along with the advantage over chemical processes of being biological in nature. Hence, being a part of clean and green technology, the environmentally friendly applications of proteases are vast, holding promising potential for the future. Their therapeutic potential against controlling the spread of epidemics like malaria and AIDS and diseases like cancer is being researched. Last few decades have enhanced our knowledge to explore the applications of these enzymes in industries beyond the food technology, like textiles, leather, detergent, agriculture, and pharmaceutical industry. The ‘era of OMICS’ has further provided future directions of utilizing proteolytic potential of these enzymes in nanotechnology and site-specific or targeted drug delivery systems (Zhao et al. 2020), as well as for imaging technology in cancer diagnostics (Abousaway et al. 2021). Novel engineered proteases activated by small molecules hold the future of specifically controlling the activity of human proteases, and significantly changing the course of cell cycle and probably even cell death (Gerry and Schreiber 2020). These molecules present the potential strategies of targeting the otherwise unreachable targets inside the cells, promising great clinical applications in future. Engineering the proteases in accordance with their prerequisite application is the challenge for future.
Conclusion
Applications of proteases in many areas of industry are already well established, while they are being explored as promising tools in pharmaceuticals (therapeutic and diagnostics), waste management, production of bioenergy, agriculture and food industry. The bottlenecks in the production of novel and desirable microbial proteases can be narrowed down to their substrate specificity, stability, and production costs. New age technologies are increasingly being used to discover or engineer novel proteases overcoming these obstacles. While directed evolution is being utilized for optimizing available enzymes, rational redesigning approaches aim at producing new ones. Future efforts of replacing the questionable practices in industry with enzyme or protease-based methods will significantly benefit both environment and survival of human species.
References
Aballay E, Prodan S, Zamorano A, Castaneda-Alvarez C (2017) Nematicidal effect of rhizobacteria on plant-parasitic nematodes associated with vineyards. World J Microbiol Biotechnol 33:131. https://doi.org/10.1007/s11274-017-2303-9
Abidi F, Aissaoui N, Chobert J-M, Haertlé T, Marzouki MN (2014) Neutral serine protease from Penicillium italicum. Purification, biochemical characterization, and use for antioxidative peptide preparation from Scorpaena notata muscle. Appl Biochem Biotechnol 174(1):186–205. https://doi.org/10.1007/s12010-014-1052-6
Abousaway O, Rakhshandehroo T, Van den Abbeele AD, Kircher MF, Rashidian M (2021) Noninvasive imaging of cancer immunotherapy. Nanotheranostics 5:90–112. https://doi.org/10.7150/ntno.50860
Abu-Tahon MA, Arafat HH, Isaac GS (2020) Laundry detergent compatibility and dehairing efficiency of alkaline thermostable protease produced from Aspergillus terreus under solid-state fermentation. J Oleo Sci 69(3):241–254. https://doi.org/10.5650/jos.ess19315
Agbowuro AA, Huston WM, Gamble AB, Tyndall JDA (2018) Proteases and protease inhibitors in infectious diseases. Med Res Rev 38:1295–1331. https://doi.org/10.1002/med.21475
Agyei D, Ongkudon CM, Wei CY, Chan AS, Danquah MK (2016) Bioprocess challenges to the isolation and purification of bioactive peptides. Food Bioprod Proc 98:244–256. https://doi.org/10.1016/j.fbp.2016.02.003
Ahmad W, Tayyab M, Aftab MN, Hashmi AS, Ahmad MD, Firyal S, Wasim M, Awan AR (2020) Optimization of conditions for the higher level production of protease: characterization of protease from Geobacillus SBS-4S. Waste Biomass Valori. https://doi.org/10.1016/j.bcab.2020.101631
Aissaoui N, Chobert J-M, Haertlé T, Marzouki MN, Abidi F (2016) Purification and biochemical characterization of a neutral serine protease from Trichoderma harzianum. use in antibacterial peptide production from a fish by-product hydrolysate. Appl Biochem Biotechnol 182:831–845. https://doi.org/10.1007/s12010-016-2365-4
Al-Abdalall AH, Al-Khaldi EM (2016) Recovery of silver from used X-ray film using alkaline protease from Bacillus subtilis sub sp. subtilis. Afr J Biotechnol 15:1413–1416. https://doi.org/10.5897/AJB2016.15340
Al-Dhabi NA, Esmail GA, Ghilan A-KM, Arasu MV, Duraipandiyan V, Ponmurugan K (2020) Characterization and fermentation optimization of novel thermo stable alkaline protease from Streptomyces sp. Al-Dhabi-82 from the Saudi Arabian environment for eco-friendly and industrial applications. J King Saud Univ Sci 32:1258–1264. https://doi.org/10.1016/j.jksus.2019.11.011
Ali N, Ullah N, Qasim M, Rahman H, Khan SN, Sadig A et al (2016) Molecular characterization and growth optimization of halo-tolerant protease producing Bacillus subtilis Strain BLK-1.5 isolated from salt mines of Karak, Pakistan. Extremophiles 20:395–402. https://doi.org/10.1007/s00792-016-0830-1
Alma’abadi AD, Gojobori T, Mineta K (2015) Marine metagenome as a resource for novel enzymes. Genom Proteom Bioinform 13(5):290–295. https://doi.org/10.1016/j.gpb.2015.10.001
Alves MP, Salgado RL, Eller MR, Vidigal PMP, de Carvalho AF (2016) Characterization of a heat-resistant extracellular protease from Pseudomonas fluorescens 07A shows that low temperature treatments are more effective in deactivating its proteolytic activity. J Dairy Sci 99(10):7842–7851. https://doi.org/10.3168/jds.2016-11236
Ammasi R, Victor JS, Chellan R, Chellappa M (2020) Alkaline protease for an efficacious rehydration of skin matrix by a novel Bacillus crolab MTCC 5468 in sustainable leather production: a green approach. Biotechnol Lett 42:249–267. https://doi.org/10.1007/s10529-019-02769-0
Anandharaj M, Sivasankari B, Siddharthan N, Rani RP, Sivakumar S (2016) Production, purification, and biochemical characterization of thermostable metallo-protease from novel Bacillus alkalitelluris TWI3 isolated from tannery waste. Appl Biochem Biotechnol 178(8):1666–1686. https://doi.org/10.1007/s12010-015-1974-7
Antink MMH, Sewczyk T, Kroll S, Árki P, Beutel S, Rezwan K et al (2019) Proteolytic ceramic capillary membranes for the production of peptides under flow. Biochem Eng J 147:89–99. https://doi.org/10.1016/j.bej.2019.04.005
Ao X, Yu X, Wu D et al (2018) Purification and characterization of neutral protease from Aspergillus oryzae Y1 isolated from naturally fermented broad beans. AMB Express 8:96. https://doi.org/10.1186/s13568-018-0611-6
Asgher M, Bashir F, Iqbal HM (2018) Protease-based cross-linked enzyme aggregates with improved catalytic stability, silver removal, dehairing potentials. Int J Biol Macromol 118:1247–1256. https://doi.org/10.1016/j.ijbiomac.2018.06.107
Ash K, Sushma, Ramteke PW (2018) Optimization of extracellular alkaline protease production from Pseudomonas aeruginosa isolated from soil samples. Inter J Agric Environ Biotechnol 11:187–194. https://doi.org/10.30954/0974-1712.2018.00178.24
Asha B, Palaniswamy M (2018) Optimization of alkaline protease production by Bacillus cereus FT 1 isolated from soil. J Appl Pharma Sci 8:119–127. https://doi.org/10.7324/JAPS.2018.8219
Bajaj BK, Sharma P (2011) An alkali-thermotolerant extracellular protease from a newly isolated Streptomyces sp. DP2. New Biotechnol 28(6):725–732. https://doi.org/10.1016/j.nbt.2011.01.001
Banerjee G, Ray AK (2017) Impact of microbial proteases on biotechnological industries. Biotechnol Genetic Engg Rev 33:119–143. https://doi.org/10.1080/02648725.2017.1408256
Baweja M, Nain L, Kawarabayasi Y, Shukla P (2016a) Current technological improvements in enzymes toward their biotechnological applications. Front Microbiol 7:965. https://doi.org/10.3389/fmicb.2016.00965
Baweja M, Tiwari R, Singh PK, Nain L, Shukla P (2016b) An alkaline protease from Bacillus pumilus MP 27: functional analysis of its binding model toward its applications as detergent additive. Front Microbiol 7:1195. https://doi.org/10.3389/fmicb.2016.01195
Baweja M, Singh PK, Sadaf A, Tiwari R, Nain L, Khare SK, Shukla P (2017) Cost effective characterization process and molecular dynamic simulation of detergent compatible alkaline protease from Bacillus pumilus strain MP27. Proc Biochem 58:199–203. https://doi.org/10.1016/j.procbio.2017.04.024
Benmrad MO, Moujehed E, Elhoul MB, Mechri S, Bejar S, Zouari R, Baffoun A, Jaouadi B (2018) Production, purification, and biochemical characterization of serine alkaline protease from Penicillium chrysogenium strain X5 used as excellent bio-additive for textile processing. Int J Biol Macromol 119:1002–1016. https://doi.org/10.1016/j.ijbiomac.2018.07.194
Bernardo R, Hongying S, Fabio P, Antonio GJ (2018) Plant viral proteases: beyond the role of peptide cutters. Front Plant Sci 9:666. https://doi.org/10.3389/fpls.2018.00666
Bhagwat PK, Dandge PB (2018) Collagen and collagenolytic proteases: a review. Biocatal Agric Biotechnol 15:43–55. https://doi.org/10.1016/j.bcab.2018.05.005
Bhatia RK, Ullah S, Hoque HZ, Ahmad I, Yang YH, Bhatt AK, Bhatia SK (2021) Psychrophiles: a source of cold-adapted enzymes for energy efficient biotechnological industrial processes. J Envir Chem Eng 9:104607. https://doi.org/10.1016/j.jece.2020.104607
Białkowska AM, Morawski K, Florczak T (2017) Extremophilic proteases as novel and efficient tools in short peptide synthesis. J Ind Microbiol Biotechnol 44:1325–1342. https://doi.org/10.1007/s10295-017-1961-9
Bilal T, Malik B, Hakeem KR (2018) Metagenomic analysis of uncultured microorganisms and their enzymatic attributes. J Microbiol Methods 155:65–69. https://doi.org/10.1016/j.mimet.2018.11.014
Bond JS (2019) Proteases: history, discovery, and roles in health and disease. J Biol Chem 294:1643–1651. https://doi.org/10.1074/jbc.TM118.004156
Boughachiche F, Rachedi K, Duran R et al (2016) Optimization of alkaline protease production by Streptomyces sp. strain isolated from saltpan environment. Afr J Biotechnol 15:1401–1412. https://doi.org/10.5897/AJB2016.15259
Briki S, Hamdi O, Landoulsi A (2016) Enzymatic dehairing of goat skins using alkaline protease from Bacillus sp. SB12. Protein Expr Purif 121:9–16. https://doi.org/10.1016/j.pep.2015.12.021
Bruno S, Coppola D, di Prisco G, Giordano D, Verde C (2019) Enzymes from marine polar regions and their biotechnological applications. Mar Drugs 17:544. https://doi.org/10.3390/md17100544
Cavello IA, Crespo JM, Garcia SS, Zapiola JM, Luna MF, Cavalitto SF (2015) Plant growth promotion activity of keratinolytic fungi growing on a recalcitrant waste known as “Hair Waste.” Biotechnol Res Int. https://doi.org/10.1155/2015/952921
Chatha SAS, Asgher M, Iqbal HMN (2017) Enzyme-based solutions for textile processing and dye contaminant biodegradation—a review. Environ Sci Pollut Res 24:14005–14018. https://doi.org/10.1007/s11356-017-8998-1
Chen D, Wang D, Xu C, Chen C, Li J, Wu W, Huang X, **e H (2018a) Nematicidal protease genes screened from a soil metagenomic library to control Radopholus similis mediated by Pseudomonas fluorescens pf36. Appl Microbiol Biotechnol 102:3301–3314. https://doi.org/10.1007/s00253-018-8869-9
Chen H, Li M, Liu C, Zhang H, **an M, Liu H (2018b) Enhancement of the catalytic activity of Isopentenyl diphosphate isomerase (IDI) from Saccharomyces cerevisiae through random and site-directed mutagenesis. Microb Cell Fact 17:65. https://doi.org/10.1186/s12934-018-0913-z
Chiliveri SR, Koti S, Linga VR (2016) Retting and degumming of natural fibers by pectinolytic enzymes produced from Bacillus tequilensis SV11-UV37 using solid state fermentation. Springerplus 5(1):1–17. https://doi.org/10.1186/s40064-016-2173-x
Chu W-H (2007) Optimization of extracellular alkaline protease production from species of Bacillus. J Ind Microbiol Biotechnol 34(3):241–245. https://doi.org/10.1007/s10295-006-0192-2
Contesini FJ, Rodrigues de Melo R, Sato HH (2017) An overview of Bacillus proteases: from production to application. Crit Rev Biotechnol. https://doi.org/10.1080/07388551.2017.1354354
Contesini FJ, Melo RR, Sato HH (2018) An overview of Bacillus proteases: from production to application. Crit Rev Biotechnol 38:321–334. https://doi.org/10.1080/07388551.2017.1354354
Culp E, Wright G (2017) Bacterial proteases, untapped antimicrobial drug targets. J Antibiot 70:366–377. https://doi.org/10.1038/ja.2016.138
da Silva RR (2017) Bacterial and fungal proteolytic enzymes: production, catalysis and potential applications. Appl Biochem Biotechnol 183:1–19. https://doi.org/10.1007/s12010-017-2427-2
da Silva RR, Cabral TPdF, Rodrigues A, Hamilton C (2013) Production and partial characterization of serine and metallo peptidases secreted by Aspergillus fumigatus Fresenius in submerged and solid state fermentatio. Braz J Microbiol 44(1):235–243. https://doi.org/10.1590/S1517-83822013000100034
da Silva RR, Souto TB, Oliveira TB et al (2016a) Evaluation of the catalytic specificity, biochemical properties, and milk clotting abilities of an aspartic peptidase from Rhizomucor miehei. J Ind Microbiol Biotechnol 43:1059–1069. https://doi.org/10.1007/s10295-016-1780-4
da Silva RR, Souto TB, Oliveira TB, Oliveira LCG, Karcher D, Juliano MA et al (2016b) Evaluation of the catalytic specificity, biochemical properties, and milk clotting abilities of an aspartic peptidase from Rhizomucor miehei. J Ind Microbiol Biotechnol 43:1059–1069. https://doi.org/10.1016/j.foodchem.2017.01.009
da Silva RR, de Oliveira LC, Juliano MA, Juliano L, de Oliveira AH, Rosa JC, Cabrod H (2017) Biochemical and milk clotting properties and map** of catalytic subsites of an extra cellular aspartic peptidase from Basidiomycete fungus, Phanerochete chrysosporium. Food Chem 225:45–54. https://doi.org/10.1016/j.foodchem.2017.01.009
da Silva OS, de Oliveira RL, de Carvalho SJ, Converti A, Porto TS (2018) Thermodynamic investigation of an alkaline protease from Aspergillus tamarii URM4634: a comparative approach between crude extract and purified enzyme. Int J Biol Macromol 109:1039–1044. https://doi.org/10.1016/j.ijbiomac.2017.11.081
Daoud L, Jlidi M, Hmani H, Hadj Brahim A, El Arbi M, Ben Ali M (2017) Characterization of thermo-solvent stable protease from Halobacillus sp. CJ4 isolated from Chott Eldjerid hypersaline lake in Tunisia. J Basic Microbiol 57:104–113. https://doi.org/10.1002/jobm.201600391
Darwesh OM, El-Hawary AS, El Kelany US, El-Sherbiny GM (2019) Nematicidal activity of thermostable alkaline protease produced by Saccharomonospora viridis strain Hw G550. Biotechnol Rep (amst) 24:e00386. https://doi.org/10.1016/j.btre.2019.e00386
de Castro RJS, Ohara A, Nishide TG, Albernaz JRM, Soares MH, Sato HH (2015) A new approach for proteases production by Aspergillus niger based on the kinetic and thermodynamic parameters of the enzymes obtained. Biocatal Agric Biotechnol 4(2):199–207. https://doi.org/10.1016/j.bcab.2014.12.001
de Oliveira CT, Rieger TJ, Daroit DJ (2017) Catalytic properties and thermal stability of a crude protease from the keratinolytic Bacillus sp. CL33A. Biocatal Agric Biotechnol 10:270–277. https://doi.org/10.1016/j.bcab.2017.04.004
De Oliveira JM, Fernandes P, Benevides RG, de Assis SA (2020) Characterization and immobilization of protease secreted by the fungus Moorella speciosa. 3 Biotech. https://doi.org/10.1007/s13205-020-02412-0
Deng JJ, Huang WQ, Li ZW, Lu DL, Zhang Y, Luo XC (2018) Biocontrol activity of recombinant aspartic protease from Trichoderma harzianum against pathogenic fungi. Enzyme Microb Technol 112:35–42. https://doi.org/10.1016/j.enzmictec.2018.02.002
Devi SG, Fathima AA, Sanitha M, Iyappan S, Curtis WR, Ramya M (2016) Expression and characterization o falkaline protease from the metagenomic library of tannery activated sludge. J Biosci Bioengineer 122(6):694–700
Deng JJ, Shi D, Zhao M et al (2021) Recombinant neutral protease rNpI as fish feed additive to improve protein digestion and growth. Aquacult Res 52:273–281. https://doi.org/10.1111/are.14890
Doriya K, Jose N, Gowda M, Kumar D (2016) Solid-state fermentation vs submerged fermentation for the production of l-asparaginase. Adv Food Nutr Res 78:115–135. https://doi.org/10.1016/bs.afnr.2016.05.003
Dorra G, Ines K, Imen BS, Laurent C, Sana A, Olfa T, Pascal C, Thierry J, Ferid L (2018) Purification and characterization of a novel high molecular weight alkaline protease produced by an endophytic Bacillus halotolerans strain CT2. Inter J Biol Macromol 111:342–351. https://doi.org/10.1016/j.ijbiomac.2018.01.024
dos Santos Aguilar JG, Sato HH (2018) Microbial proteases: production and application in obtaining protein hydrolysates. Food Res Inter 103:253–262. https://doi.org/10.1016/j.foodres.2017.10.044
Dudani JS, Warren AD, Bhatia SN (2018) Harnessing protease activity to improve cancer care. Annu Rev Cancer Biol 2:353–376. https://doi.org/10.1146/annurev-cancerbio030617-050549
Fang Z, Yong YC, Zhang J, Du G, Chen J (2017) Keratinolytic protease: a green biocatalyst for leather industry. Appl Microbiol Biotechnol 101:7771–7779. https://doi.org/10.1007/s00253-017-8484-1
Far EB, Khosroushahi YA, Dilmaghani A (2020) In silico study of alkaline serine protease and production optimization in Bacillus sp. khoz1 klosed Bacillus safensis isolated from honey. Int J Pept Res Ther 26:2241–2251. https://doi.org/10.1007/s10989-020-10016-8
García-Moyano A, Larsen Ø, Gaykawad S, Christakou E, Boccadoro C, Puntervoll P, Bjerga GEK (2020) Fragment exchange plasmid tools for CRISPR/Cas9-mediated gene integration and protease production in Bacillus subtilis. Appl Environ Microbiol 87(1):e02090-e2120. https://doi.org/10.1128/AEM.02090-20
García-Moyano A, Diaz Y, Navarro J, Almendral D, Puntervoll P, Ferrer M, Bjerga GEK (2021) Two-step functional screen on multiple proteinaceous substrates reveals temperature-robust proteases with a broad-substrate range. Appl Microbiol Biotechnol. 105(8):3195–3209
Gerry CJ, Schreiber SL (2020) Unifying principles of bifunctional, proximity-inducing small molecules. Nat Chem Biol 16:369–378. https://doi.org/10.1038/s41589-020-0469-1
Gimenes NC, Silveira E, Tambourgi EB (2019) An overview of proteases: production, downstream processes and industrial applications. Sep Purif Rev 50(3):223–243. https://doi.org/10.1080/15422119.2019.1677249
Golunski S, Astolfi V, Carniel N, de Oliveira D, Di Luccio M, Mazutti MA, Treichel H (2011) Ethanol precipitation and ultrafiltration of inulinases from Kluyveromyces marxianus. Sep Purif Technol 78(3):261–265. https://doi.org/10.1016/j.seppur.2011.02.019
Guleria S, Walia A, Chauhan A, Shirkot CK (2014) Genotypic and phenotypic diversity analysis of alkalophilic proteolytic Bacillus sp. associated with rhizosphere of apple trees in trans Himalayan region of Himachal Pradesh. Proc Natl Acad Sci India B Biol Sci 86(2):331–341. https://doi.org/10.1007/s40011-014-0447-z
Guleria S, Walia A, Chauhan A, Shirkot CK (2015) Purification and characterization of detergent stable alkaline protease from Bacillus amyloliquefaciens SP1 isolated from apple rhizosphere. J Basic Microbiol 55:1–15. https://doi.org/10.1002/jobm.201500341
Guleria S, Walia A, Chauhan A, Shirkot CK (2016a) Immobilization of Bacillus amyloliquefaciens SP1 and its alkaline protease in various matrices for effective hydrolysis of casein. 3 Biotech 6:208. https://doi.org/10.1007/s13205-016-0519-2
Guleria S, Walia A, Chauhan A, Shirkot CK (2016b) Molecular characterization of alkaline protease of Bacillus amyloliquefaciens SP1 involved in biocontrol of Fusarium oxysporum. Int J Food Microbiol 232:134–143. https://doi.org/10.1016/j.ijfoodmicro.2016.05.030
Guleria S, Walia A, Chauhan A, Shirkot CK (2016c) Optimization of milk clotting enzyme production by Bacillus amyloliquefaciens SP1 isolated from apple rhizosphere. Bioresour Bioprocess 3:30. https://doi.org/10.1186/s40643-016-0108-6
Guleria S, Kumar A, Sharma S, Kulshrestha S, Chauhan A (2017) Metagenomics of fermented foods: implications on probiotic development. In: Mining of microbial wealth and metagenomics. Springer, pp 333–355. https://doi.org/10.1007/978-981-10-5708-3_19
Guleria S, Walia A, Chauhan A, Shirkot CK (2018) Production and eco-friendly application of alkaline protease from Bacillus amyloliquefaciens sp1. Indian J Biotechnol 17:448–458. http://nopr.niscair.res.in/handle/123456789/45281
Gupta R, Beg Q, Khan S, Chauhan B (2002) An overview on fermentation, downstream processing and properties of microbial alkaline proteases. Appl Microbiol Biotechnol 60(4):381–395. https://doi.org/10.1007/s00253-002-1142-1
Gupta R, Rajput R, Sharma R, Gupta N (2013) Biotechnological applications and prospective market of microbial keratinases. Appl Microbiol Biotechnol 97:9931–9940. https://doi.org/10.1007/s00253-013-5292-0
Gurumallesh P, Alagu K, Ramakrishnan B, Muthusamy S (2019) A systematic reconsideration on proteases. Inter J Bio Macromol 128:254–267. https://doi.org/10.1016/j.ijbiomac.2019.01.081
Gurung N, Ray S, Bose S, Rai V (2013) A broader view: microbial enzymes and their relevance in industries, medicine, and beyond. Biomed Res Int 2013:329121. https://doi.org/10.1155/2013/329121
Hakim A, Bhuiyan FR, Iqbal A, Emon TH, Ahmed J, Azad AK (2018) Production and partial characterization of dehairing alkaline protease from Bacillus subtilis AKAL7 and Exiguobacterium indicum AKAL11 by using organic municipal solid wastes. Heliyon 4:e00646. https://doi.org/10.1016/j.heliyon.2018.e00646
Hammami A, Bayoudh A, Hadrich B, Abdelhedi O, Jridi M, Nasri M (2020) Response-surface methodology for the production and the purification of a new H2O2-tolerant alkaline protease from Bacillus invictae AH1 strain. Biotech Progress 36:e2965. https://doi.org/10.1002/btpr.2965
Hamza TA (2017) Bacterial protease enzyme: safe and good alternative for industrial and commercial use. Int J Chem Biomol Sci 3:1–10
Hejdysz M, Kaczmarek SA, Kubis M, Wiśniewska Z, Peris S, Budnik S, Rutkowski A (2020) The effect of protease and Bacillus licheniformis on nutritional value of pea, faba bean, yellow lupin and narrow-leaved lupin in broiler chicken diets. Br Poultry Sci 61(3):287–293. https://doi.org/10.1080/00071668.2020.1716303
Homaei A, Lavajoo F, Sariri R (2016) Development of marine biotechnology as a resource for novel proteases and their role in modern biotechnology. Int J Biol Macromol 88:542–552. https://doi.org/10.1016/j.ijbiomac.2016.04.023
Hosokawa M, Hoshino Y, Nishikawa Y, Hirose T, Yoon DH, Mori T, Sekiguchi T, Shoji S, Takeyama H (2015) Droplet-based microfluidics for high-throughput screening of a metagenomic library for isolation of microbial enzymes. Biosens Bioelectron 67:379–385. https://doi.org/10.1016/j.bios.2014.08.059
Hu H, Gao Y, Li X, Chen S, Yan S, Tian X (2020) Identification and nematicidal characterization of proteases secreted by endophytic bacteria Bacillus cereus BCM2. Phytopathology 110(2):336–344. https://doi.org/10.1094/PHYTO-05-19-0164-R
Huang Y, Wang Y, Xu Y (2017) Purification and characterisation of an acid protease from the Aspergillus hennebergii HX08 and its potential in traditional fermentation. J Inst Brew 123(3):432–441. https://doi.org/10.1002/jib.427
Hussain F, Kamal S, Rehman S, Azeem M, Bibi I, Ahmed T, Iqbal HMN (2017) Alkaline protease production using response surface methodology, characterization and industrial exploitation of alkaline protease of Bacillus subtilis sp. Catal Lett 147:1204–1213. https://doi.org/10.1007/s10562-017-2017-5
Ibrahim ASS, Elbadawi YB, El-Tayeb MA et al (2019) Alkaline serine protease from the new halotolerant alkaliphilic Salipaludibacillus agaradhaerens strain AK-R: purification and properties. 3 Biotech 9:391. https://doi.org/10.1007/s13205-019-1928-9
Ida ÉL, da Silva RR, de Oliveira TB, Souto TB, Leite JA, Rodrigues A, Cabral H (2017) Biochemical properties and evaluation of washing performance in commercial detergent compatibility of two collagenolytic serine peptidases secreted by Aspergillus fischeri and Penicillium citrinum. Prep Biochem Biotechnol 47(3):282–290. https://doi.org/10.1080/10826068.2016.1224247
Imdakim MM, Hassan Z, Aween MM, Elshaafi I, Muhialdin BJ (2015) Milk clotting and proteolytic activity of enzyme preparation from Pediococcus acidilactici SH for dairy products. Afr J Biotechnol 14(2):133–142
Jadhav HP, Sonawane MS, Khairnar MH et al (2020) Production of alkaline protease by rhizospheric Bacillus cereus HP_RZ17 and Paenibacillus xylanilyticus HP_RZ19. Environ Sustain 3:5–13. https://doi.org/10.1007/s42398-020-00096-z
Jaouadi NZ, Jaouadi B, Aghajari N, Bejar S (2012) The overexpression of the SAPB of Bacillus pumilus CBS and mutated sapB-L31I/T33S/N99Y alkaline proteases in Bacillus subtilis DB430: new attractive properties for the mutant enzyme. Bioresour Technol 105:142–151. https://doi.org/10.1016/j.biortech.2011.11.115
Jensen K, Østergaard PR, Wilting R, Lassen SF (2010) Identification and characterization of a bacterial glutamic peptidase. BMC Biochem 11(1):1–12. https://doi.org/10.1186/1471-2091-11-47
Jo S, Kim S, Shin DH, Kim M-S (2020) Inhibition of SARS-CoV 3CL protease by flavonoids. J Enz Inhib Med Chem 35:145–151. https://doi.org/10.1080/14756366.2019.1690480
Joshi S, Satyanarayana T (2013) Characteristics and applications of a recombinant alkaline serine protease from a novel bacterium Bacillus lehensis. Bioresour Technol 131:76–85. https://doi.org/10.1016/j.biortech.2012.12.124
Ju S, Lin J, Zheng J, Wang S, Zhou H, Sun M (2016) Alcaligenes faecalis ZD02, a novel nematicidal bacterium with an extracellular serine protease virulence factor. Appl Environ Microbiol 82(7):2112–2120. https://doi.org/10.1128/AEM.03444-15
Kalaikumari SS, Vennila T, Monika V, Chandraraj K, Gunasekaran P, Rajendhran J (2019) Bioutilization of poultry feather for keratinase production and its application in leather industry. J Clean Prod 208:44–53. https://doi.org/10.1016/j.jclepro.2018.10.076
Kamarudin NB, Sharma S, Gupta A et al (2017) Statistical investigation of extraction parameters of keratin from chicken feather using Design-Expert. 3 Biotech 7:127. https://doi.org/10.1007/s13205-017-0767-9
Kandasamy S, Muthusamy G, Balakrishnan S, Duraisamy S, Thangasamy S, Seralathan K-K, Chinnappan S (2016) Optimization of protease production from surface-modified coffee pulp waste and corncobs using Bacillus sp. by SSF. 3 Biotech 6(2):167. https://doi.org/10.1007/s13205-016-0481-z
Kanmani R, Dhivya S, Jayalakshmi S, Vijayabaskar P (2011) Studies on detergent additives of protease enzyme from an estuarine bacterium Bacillus cereus. Int Res J Biotechnol 2(7):157–163
Karray A, Alonazi M, Horchani H, Ben BA (2021) A novel thermostable and alkaline protease produced from Bacillus stearothermophilus isolated from olive oil mill sols suitable to industrial biotechnology. Molecules 26:1139. https://doi.org/10.3390/molecules26041139
Kasana RC, Salwan R, Yadav SK (2011) Microbial proteases: detection, production, and genetic improvement. Crit Rev Microbiol 37(3):262–276. https://doi.org/10.3109/1040841X.2011.577029
Kshetri P, Ningombam O, Ningombam D (2016) Optimization of alkaline protease production by alkaliphilic Bacillus sp. KW2 in low cost medium using statistical approaches. Appl Micro Open Access 2(3):1–8. https://doi.org/10.4172/2471-9315.1000117
Kumar D, Bhalla TC (2005) Microbial proteases in peptide synthesis: approaches and applications. Appl Microbiol Biotechnol 68:726–736. https://doi.org/10.1007/s00253-005-0094-7
Kumar L, Jain SK (2018) Proteases: a beneficial degradative enzyme in therapeutic applications. Inter J Sci Res Biol Sci 5:114–118. https://doi.org/10.26438/ijsrbs/v5i4.114118
Kumar DM, Premavathi V, Govindarajan N, Balakumaran M, Kalaichelvan P (2012) Production and purification of alkaline protease from Bacillus sp. MPTK 712 isolated from dairy sludge. Glob Veterinaria 8(5):433–439
Kumawat TK, Sharma A, Sharma V, Chandra S (2018) Keratin waste: the biodegradable polymers. Keratin Miroslav Blumenberg IntechOpen. https://doi.org/10.5772/intechopen.79502
Lam MQ, Nik Mut NN, Thevarajoo S et al (2018) Characterization of detergent compatible protease from halophilic Virgibacillus sp. CD6. 3 Biotech 8:104. https://doi.org/10.1007/s13205-018-1133-2
Lan G, Li C, He L, Zeng X, Zhu Q (2020) Effects of different strains and fermentation method on nattokinase activity, biogenic amines, and sensory characteristics of natto. J Food Sci Technol 57(12):4414–4423. https://doi.org/10.1007/s13197-020-04478-3
Lateef A, Adelere IA, Gueguim-Kana EB (2015) Bacillus safensis LAU 13: a new source of keratinase and its multi-functional biocatalytic applications. Biotechnol Biotechnol Equip 29:54–63. https://doi.org/10.1080/13102818.2014.986360
Li Y, Wu C, Zhou M, Wang ET, Zhang Z, Liu W, Ning J, **e Z (2017) Diversity of cultivable protease-producing bacteria in laizhou bay sediments, Bohai Sea. China Front Microbiol 8:405. https://doi.org/10.3389/fmicb.2017.00405
Li C, Zhang R, Wang J, Wilson LM, Yan Y (2020) Protein engineering for improving and diversifying natural product biosynthesis. Trends Biotechnol 38(7):729–744. https://doi.org/10.1016/j.tibtech.2019.12.008
Limkar MB, Pawar SV, Rathod VK (2019) Statistical optimization of xylanase and alkaline protease co-production by Bacillus sp. using Box-Behnken Design under submerged fermentation using wheat bran as a substrate. Biocatal Agric Biotechnol 17:455–464. https://doi.org/10.1016/j.bcab.2018.12.008
Liu J, Sharma A, Niewiara MJ, Singh R, Ming R, Yu Q (2018) Papain-like cysteine proteases in Carica papaya: lineage-specific gene duplication and expansion. BMC Genom 19:26. https://doi.org/10.1186/s12864-017-4394-y
Lohse MB, Gulati M, Craik CS, Johnson AD, Nobile CJ (2020) Combination of antifungal drugs and protease inhibitors prevent Candida albicans biofilm formation and disrupt mature biofilms. Front Microbiol 11:1027. https://doi.org/10.3389/fmicb.2020.01027
Machado ARGM, Teixeira MFS, de Souza Kirsch L, Campelo MDCL, de Aguiar Oliveira IM (2016) Nutritional value and proteases of Lentinus citrinus produced by solid state fermentation of lignocellulosic waste from tropical region. Saudi J Biol Sci 23:621–627. https://doi.org/10.1016/j.sjbs.2015.07.002
Majumder R, Banik SP, Ramrakhiani L, Khowala S (2015) Bioremediation by alkaline protease (AkP) from edible mushroom Termitomyces clypeatus: optimization approach based on statistical design and characterization for diverse applications. J Chem Technol Biotechnol 90:1886–1896. https://doi.org/10.1002/jctb.4500
Mamo J, Assefa F (2018) The role of microbial aspartic protease enzyme in food and beverage industries. J Food Qual 2018:7957269. https://doi.org/10.1155/2018/7957269
Mandal S, Banerjee D (2019) Proteases from endophytic fungi with potential industrial applications. In: Yadav A, Mishra S, Singh S, Gupta A (eds) Recent advancement in white biotechnology through fungi. Fungal biology. Springer, Cham. https://doi.org/10.1007/978-3-030-10480-1_10
Mandujano-González V, Villa-Tanaca L, Anducho-Reyes MA, Mercado-Flores Y (2016) Secreted fungal aspartic proteases: a review. Rev Iberoam Micol 33:76–82. https://doi.org/10.1016/j.riam.2015.10.003
Martínez V, Lauritsen I, Hobel T, Li S, Nielsen AT, Nørholm MH (2017) CRISPR/Cas9-based genome editing for simultaneous interference with gene expression and protein stability. Nucleic Acids Res 45(20):e171
Maruthiah T, Somanath B, Jasmin JV, Immanuel G, Palavesam A (2016) Production, purification and characterization of halophilic organic solvent tolerant protease from marine crustacean shell wastes and its efficacy on deproteinization. 3 Biotech 6:157. https://doi.org/10.1007/s13205-016-0474-y
Masi C, Chandramohan C, Fazil AM (2017) Immobilization of the magnetic nanoparticles with alkaline protease enzyme produced by Enterococcus hirae and Pseudomonas aeruginosa isolated from dairy effluents. Braz Archive Biol Technol 60:e17160572. https://doi.org/10.1590/1678-4324-2017160572
Matkawala F, Nighojkar S, Kumar A, Nighojkar A (2021) Microbial alkaline serine proteases: production, properties and applications. World J Microbiol Biotechnol 37(4):1–12. https://doi.org/10.1007/s11274-021-03036-z
Meshram V, Saxena S (2016) Potential fibrinolytic activity of an endophytic Lasiodiplodia pseudotheobromae species. 3 Biotech 6:114. https://doi.org/10.1007/s13205-016-0428-4
Meshram V, Saxena S, Paul K (2016) Xylarinase: a novel clot busting enzyme from an endophytic fungus Xylaria curta. J Enzyme Inhib Med Chem 31(6):1502–1511. https://doi.org/10.3109/14756366.2016.1151013
Mhamdi S, Bkhairia I, Nasri R, Mechichi T, Nasri M, Kamoun AS (2017) Evaluation of the biotechnological potential of a novel purified protease BS1 from Bacillus safensis S406 on the chitin extraction and detergent formulation. Int J Biol Macromol 104:739–747. https://doi.org/10.1016/j.ijbiomac.2017.06.062
Mitra P, Chakrabartty PK (2005) An extracellular protease with depilation activity from Streptomyces nogalator. Microbial Technol 112:35–42
More SV, Chavan S, Prabhune A (2017) Silk degumming and utilization of silk sericin by hydrolysis using alkaline protease from Beauveria sp. (MTCC 5184): a green approach. J Nat Fibers 15(1):1–11. https://doi.org/10.1080/15440478.2017.1330718
Moridshahi R, Bahreini M, Sharifmoghaddam M, Asoodeh A (2020) Biochemical characterization of an alkaline surfactant-stable keratinase from a new keratinase producer, Bacillus zhangzhouensis. Extremophiles 24:693–704. https://doi.org/10.1007/s00792-020-01187-9
Mothe T, Sultanpuram VR (2016) Production, purification and characterization of a thermotolerant alkaline serine protease from a novel species Bacillus caseinilyticus. 3 Biotech 6:53. https://doi.org/10.1007/s13205-016-0377-y
Motyan JA, Toth F, Tozser J (2013) Research applications of proteolytic enzymes in molecular biology. Biomolecules 3(4):923–942. https://doi.org/10.3390/biom3040923
Muneer F, Siddique MH, Azeem F, Rasul I, Muzammil S, Zubair M, Nadeem H (2020) Microbial l-asparaginase: purification, characterization and applications. Arch Microbiol 5:1–15. https://doi.org/10.1007/s00203-020-01814-1
Mukhtar H, Haq I (2013) Comparative evaluation of agroindustrial by products for the production of alkaline protease by wild and mutant strains of Bacillus subtilis in submerged and solid state fermentation. Scientific World J 2013:6. https://doi.org/10.1155/2013/538067
Nadeem F, Mehmood T, Naveed M et al (2019) Protease production from Cheotomium globusum through central composite design using agricultural wastes and its immobilization for industrial exploitation. Waste Biomass Valori. https://doi.org/10.1007/s12649-019-00890-9
Nahar S, Mizan MFR, Ha AJW, Ha SD (2018) Advances and future prospects of enzyme-based biofilm prevention approaches in the food industry. Compr Rev Food Sci Food Saf 17(6):1484–1502. https://doi.org/10.1111/1541-4337.12382
Nandan A, Nampoothiri KM (2020) Therapeutic and biotechnological applications of substrate specific microbial aminopeptidases. Appl Microbiol Biotechnol 104:5243–5257. https://doi.org/10.1007/s00253-020-10641-9
Naveed M, Nadeem F, Mehmood T, Bilal M, Anwar Z, Amjad F (2021) Protease-a versatile and ecofriendly biocatalyst with multi-industrial applications: an updated review. Catal Lett 151(2):307–323. https://doi.org/10.1007/s10562-020-03316-7
Niyonzima FN, More SS (2013) Screening and optimization of cultural parameters for an alkaline protease production by Aspergillus terreus gr. under submerged fermentation. Int J Pharm Bio Sci 4(1):1016–1028
Nongonierma AB, FitzGerald RJ (2015) The scientific evidence for the role of milk protein-derived bioactive peptides in humans: a review. J Funct Foods 17:640–656. https://doi.org/10.1016/j.jff.2015.06.021
Oh M, Han JW, Lee C, Choi GJ, Kim H (2018) Nematicidal and plant growth-promoting activity of Enterobacter asburiae HK169: genome analysis provides insight into its biological activities. J Microbiol Biotechnol 28:968–975. https://doi.org/10.4014/jmb.1801.01021
Omodamilola OI, Ibrahim AU (2018) CRISPR technology: advantages, limitations and future direction. J Biomed Pharm Sci 1(115):2
Osmolovskiy AA, Popova EA, Kreyer VG, Baranova NA, Egorov NS (2021) Vermiculite as a new carrier for extracellular protease production by Aspergillus spp. under solid-state fermentation. Biotechnol Rep 29:e00576. https://doi.org/10.1016/j.btre.2020.e00576
Pal GK, Suresh PV (2016) Microbial collagenases: challenges and prospects in production and potential applications in food and nutrition. RSC Adv 6:33763–33780. https://doi.org/10.1039/c5ra23316j
Park S, Lee JJ, Yang BM, Cho JH, Kim S, Kang J, Song M (2020) Dietary protease improves growth performance, nutrient digestibility, and intestinal morphology of weaned pigs. J Animal Sci Technol 62(1):21. https://doi.org/10.5187/jast.2020.62.1.21
Patel S, Homaei A, El-Seedide HR, Akhtar N (2018) Cathepsins: proteases that are vital for survival but can also be fatal. Biomed Pharmacother 105:526–553. https://doi.org/10.1016/j.biopha.2018.05.148
Pathak AP, Rathod MG (2018) A review on alkaline protease producers and their biotechnological perspectives. Ind J Geo Mar Sci 47:1113–1119
Peng D, Lin J, Huang Q, Zheng W, Liu G, Zheng J, Zhu L, Sun M (2016) A novel metalloproteinase virulence factor is involved in Bacillus thuringiensis pathogenesis in nematodes and insects. Environ Microbiol 18:846–862. https://doi.org/10.1111/1462-2920.13069
Pessoa TB, Rezende RP, Marques EdLS, Pirovani CP, Dos Santos TF, dos Santos Gonçalves AC, Romano CC, Dotivo NC, Freitas AC, Salay LC (2017) Metagenomic alkaline protease from mangrove sediment. J Basic Microbiol 57(11):962–973. https://doi.org/10.1002/jobm.201700159
Prabhavathy G, Rajasekara M, Senthilkumar B (2013) Identification of industrially important alkaline protease producing Bacillus subtilis by 16s rRNA sequence analysis and its applications. Int J Res Pharma Biomed Sci 4:332–338
Prajapati VS, Ray S, Narayan J, Joshi CC, Patel KC, Trivedi UB, Patel RM (2017) Draft genome sequence of a thermostable, alkaliphilic α-amylase and protease producing Bacillus amyloliquefaciens strain KCP2. 3 Biotech. https://doi.org/10.1007/s13205-017-1005-1
Prayogo FA, Budiharjo A, Kusumaningrum HP, Wijanarka W, Suprihadi A, Nurhayati N (2020) Metagenomic applications in exploration and development of novel enzymes from nature: a review. J Genet Eng Biotechnol 18(1):1–10. https://doi.org/10.1186/s43141-020-00043-9
Proteases Market size, regional outlook, application growth potential, COVID-19 impact analysis, COVID-19 impact analysis, competitive market growth & forecast, 2021–2027. https://www.gminsights.com/industry-analysis/proteases-market
Putatunda C, Kundu BS, Bhatia R (2019) Purification and characterization of alkaline protease from Bacillus sp. HD292. Proc Natl Acad Sci India B Biol Sci 89:957–965. https://doi.org/10.1007/s40011-018-1011-z
Putranto WS, Kusmajadi S, Hartati C, Apon ZM, Puspo EG, Harsi DK, Maggy TS (2017) Enterococcus faecium 1.15 isolated from bakasam showed milk clotting activity. Annales Bogoriensis 21:9–14
Putranto WS, Maggy TS, Harsi DK, Puspo EG, Apon ZM (2020a) A novel rennin like protease from Lactobacillus plantarum 1.13 isolated from Indonesian fermented meat (Bakasam). Biocatal Agric Biotechnol. https://doi.org/10.1016/j.bcab.2020.101818
Putranto WS, Mustopa AZ, Arizah K, Prastyowati A (2020b) The purification of rennin-like protease from Lactobacillus paracasei isolated from ettawa goat milk. Annales Bogorienses 24:74–80. https://doi.org/10.14203/ann.bogor.2020.v24.n2.74-80
Qamar SA, Asgher M, Bilal M (2020) Immobilization of alkaline protease from Bacillus brevis using Ca-Alginate entrapment strategy for improved catalytic stability, silver recovery, and dehairing potentialities. Catal Lett 150:3572–3583. https://doi.org/10.1007/s10562-020-03268-y
Qureshi AS, Khushk I, Ali CH, Chisti Y, Ahmad A, Majeed H (2016) Coproduction of protease and amylase by thermophilic Bacillus sp. BBXS-2 using open solid-state fermentation of lignocellulosic biomass. Biocatal Agric Biotechnol 8:146–151. https://doi.org/10.1016/j.bcab.2016.09.006
Rahem FZ, Badis A, Zenati B, Mechri S, Hadjidj R, Rekik H, Eddouaouda K, Annane R, Jaouadi B (2021) Characterization of a novel serine alkaline protease from Bacillus atrophaeus NIJ as a thermophilic hydrocarbonoclastic strain and its application in laundry detergent formulations. Algeria J Environ Sci Technol 7:1707–1724
Rajput R, Gupta R (2013) Thermostable keratinase from Bacillus pumilus KS12: production, chitin crosslinking and degradation of Sup35NM aggregates. Bioresour Technol 133:118–126. https://doi.org/10.1016/j.biortech.2013.01.091
Ramakodi MP, Santhosh N, Pragadeesh T, Mohan SV, Basha S (2020) Production of protease enzyme from slaughterhouse effluent: an approach to generate value-added products from waste. Bioresour Technol Rep 12:100552
Rawlings ND, Barrett AJ, Thomas PD, Huang X, Bateman A, Finn RD (2018) The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res 46:D624–D632. https://doi.org/10.1093/nar/gkx1134
Razzaq A, Shamsi S, Ali A, Ali Q, Sajjad M, Malik A, Ashraf M (2019) Microbial proteases applications. Front Bioeng Biotechnol 7:110. https://doi.org/10.3389/fbioe.2019.00110
Rehman R, Ahmed M, Siddique A, Hasan F, Hameed A, Jamal A (2017) Catalytic role of thermostable metalloproteases from Bacillus subtilis KT004404 as dehairing and destaining agent. Appl Biochem Biotechnol 181(1):434–450. https://doi.org/10.1007/s12010-016-2222-5
Rekik H, Jaouadi NZ, Gargouri F, Bejar W, Frikha F, Jmal N, Bejar S, Jaouadi B (2019) Production, purification and biochemical characterization of a novel detergent-stable serine alkaline protease from Bacillus safensis strain RH12. Int J Biol Macromol 12:1227–1239. https://doi.org/10.1016/j.ijbiomac.2018.10.139
Sa A, Singh A, Garg S, Kumar A, Kumar H (2012) Screening, isolation and characterisation of protease producing moderately halophilic microorganisms. Asian J Microbiol Biotech Env Sci 14(4):603–612
Sahay H, Yadav AN, Singh AK et al (2017) Hot springs of Indian Himalayas: potential sources of microbial diversity and thermostable hydrolytic enzymes. 3 Biotech 7:118. https://doi.org/10.1007/s13205-017-0762-1
Salwan R, Sharma V (2019) Trends in extracellular serine proteases of bacteria as detergent bioadditive: alternate and environmental friendly tool for detergent industry. Arch Microbiol 201:863–877. https://doi.org/10.1007/s00203-019-01662-8
Sandhya C, Sumantha A, Szakacs G, Pandey A (2005) Comparative evaluation of neutral protease production by Aspergillus oryzae in submerged and solid-state fermentation. Proc Biochem 40(8):2689–2694. https://doi.org/10.1016/j.procbio.2004.12.001
Sattar H, Bibi Z, Kamran A, Aman A, Qader SAU (2019) Degradation of complex casein polymer: production and optimization of a novel serine metalloprotease from Aspergillus niger KIBGE-IB36. Biocatal Agric Biotechnol 21:101256. https://doi.org/10.1016/j.bcab.2019.101256
Sharma N (2019) A review on fungal alkaline protease. J Emerg Tech Innov Res 6(6):1–14
Sharma A, Gupta SP (2017) Fundamentals of viruses and their proteases. Viral Protease Inhib. https://doi.org/10.1016/B978-0-12-809712-0.00001-0
Sharma KM, Kumar R, Panwar S, Kumar A (2017) Microbial alkaline proteases: optimization of production parameters and their properties. J Genet Eng Biotechnol 15(1):115–126. https://doi.org/10.1016/j.jgeb.2017.02.001
Silveira JR, Raymond GJ, Hughson AG, Race RE, Sim VL, Hayes SF, Caughey B (2005) The most infectious prion protein particles. Nature 437(7056):257–261. https://doi.org/10.1038/nature03989
Singh S, Bajaj BK (2017) Potential application spectrum of microbial proteases for clean and green industrial production. Energ Ecol Environ 2:370–386. https://doi.org/10.1007/s40974-017-0076-5
Singh R, Kumar M, Mittal A, Mehta PK (2016) Microbial enzymes: industrial progress in 21st century. 3 Biotech 6:174. https://doi.org/10.1007/s13205-016-0485-8
Singhania RR, Patel AK, Soccol CR, Pandey A (2009) Recent advances in solid-state fermentation. Biochem Eng J 44(1):13–18. https://doi.org/10.1016/j.bej.2008.10.019
Soccol CR, da Costa ESF, Letti LAJ, Karp SG, Woiciechowski AL, de Souza Vandenberghe LP (2017) Recent developments and innovations in solid state fermentation. Biotechnol Res Innov 1(1):52–71. https://doi.org/10.1016/j.biori.2017.01.002
Song P, Cheng L, Tian K et al (2020) Biochemical characterization of two new Aspergillus niger aspartic proteases. 3 Biotech 10:303. https://doi.org/10.1007/s13205-020-02292-4
Sonune N, Garode A (2018) Isolation, characterization and identification of extracellular enzyme producer Bacillus licheniformis from municipal wastewater and evaluation of their biodegradability. Biotechnol Res Innov 2(1):37–44. https://doi.org/10.1016/j.biori.2018.03.001
Souza PM, Werneck G, Aliakbarian B, Siqueira F, Filho EXF, Perego P, Converti A, Magalhães PO, Pessoa Junior A (2017) Production, purification and characterization of an aspartic protease from Aspergillus foetidus. Food Chem Toxicol 109:1103–1110. https://doi.org/10.1016/j.fct.2017.03.055
Srilakshmi J, Madhavi J, Lavanya S, Ammani K (2015) Commercial potential of fungal protease: past, present and future prospects. J Pharm Chem Biol Sci 2(4):218–234
Srivastava B, Khatri M, Singh G, Arya SK (2020) Microbial keratinases: an overview of biochemical characterization and its eco-friendly approach for industrial applications. J Clean Prod 252:119847. https://doi.org/10.1016/j.jclepro.2019.119847
Steele HL, Wolfgang RS (2005) Metagenomics: advances in ecology and biotechnology. FEMS Microbiol Lett 247(2):105–111. https://doi.org/10.1016/j.femsle.2005.05.011
Su C, Gong JS, Qin J, Li H, Li H, Xu ZH, Shi JS (2020) The tale of a versatile enzyme: molecular insights into keratinase for its industrial dissemination. Biotechnol Adv. https://doi.org/10.1016/j.biotechadv.2020.107655
Suleiman AD, Rahman A, Mohd Yusof H, Mohd Shariff F, Yasid NA (2020) Effect of cultural conditions on protease production by a thermophilic Geobacillus thermoglucosidasius SKF4 isolated from Sungai Klah hot Spring Park, Malaysia. Molecules 25(11):2609. https://doi.org/10.3390/molecules25112609
Sun J, Wang M, Cao J, Zhao Y, Jiang W (2010) Characterization of three novel alkaline serine proteases from tomato (Lycopersicum esculentum Mill.) Fruit and their potential application. J Food Biochem 34(5):1014–1031. https://doi.org/10.1111/j.1745-4514.2010.00346.x
Sun Q, Chen F, Geng F, Luo Y, Gong S, Jiang Z (2018) A novel aspartic protease from Rhizomucor miehei expressed in Pichia pastoris and its application on meat tenderization and preparation of turtle peptides. Food Chem 245:570–577. https://doi.org/10.1016/j.foodchem.2017.10.113
Suwannaphan S, Fufeungsombut E, Promboon A, Chim-Anage P (2017) A serine protease from newly isolated Bacillus sp. for efficient silk degumming, sericin degrading and colour bleaching activities. Int Biodeter Biodegr 117:141–149. https://doi.org/10.1016/j.ibiod.2016.12.009
Tavano OL, Berenguer-Murcia A, Secundo F, Fernandez-Lafuente R (2018) Biotechnological applications of proteases in food technology. Comp Rev Food Sci Food Safety 17:412–436. https://doi.org/10.1111/1541-4337.12326
Tekin A, Uzuner U, Sezen K (2020) Homology modeling and heterologous expression of highly alkaline subtilisin-like serine protease from Bacillus halodurans C-125. Biotechnol Lett. https://doi.org/10.1007/s10529-020-03025-6
Thakur A, Putatunda C, Sharma R, Mehta R, Solanki P, Bhatia K (2020) Innovative techniques for improving microbial enzyme production. In: Microbial diversity, interventions and scope. Springer, pp 157–184. https://doi.org/10.1007/978-981-15-4099-8
Tigabu BM, Agide FD, Mohraz M, Nikfar S (2020) Atazanavir/ritonavir versus Lopinavir/ritonavir-based combined antiretroviral therapy (cART) for HIV-1 infection: a systematic review and meta-analysis. Afr Health Sci 20:91–101. https://doi.org/10.4314/ahs.v20i1.14
Uddin F, Rudin CM, Sen T (2020) CRISPR gene therapy: applications, limitations, and implications for the future. Front Oncol 10:1387. https://doi.org/10.3389/fonc.2020.01387
Verma R, Schwaneberg U, Roccatano D (2012) Computer-aided protein directed evolution: a review of web servers, databases and other computational tools for protein engineering. Comput Struct Biotechnol J 2(3):e201209008. https://doi.org/10.5936/csbj.201209008
Verma SK, Kaur S, Tevetia A, Chatterjee S, Sharma PC (2021) Structural characterization and functional annotation of microbial proteases mined from solid tannery waste metagenome. Biologia 76(6):1829–1842. https://doi.org/10.1007/s11756-021-00727-8
Vijayaraghavan P, Lazarus S, Vincent SGP (2014) De-hairing protease production by an isolated Bacillus cereus strain AT under solid-state fermentation using cow dung: biosynthesis and properties. Saudi J Biol Sci 21(1):27–34. https://doi.org/10.1016/j.sjbs.2013.04.010
Villa ALV, Aragao MRS, Santos EPD, Mazotto AM, Zingali RB, de Souza EP, Vermelho AB (2013) Feather keratin hydrolysates obtained from microbial keratinases: effect on hair fiber. BMC Biotechnol 18:13–15. https://doi.org/10.1186/1472-6750-13-15
Vojcic L, Pitzler C, Körfer G, Jakob F, Martinez R, Maurer K-H, Schwaneberg U (2015) Advances in protease engineering for laundry detergents. New Biotechnol 32(6):629–634. https://doi.org/10.1016/j.nbt.2014.12.010
Waschkowitz T, Rockstroh S, Daniel R (2009) Isolation and characterization of metalloproteases with a novel domain structure by construction and screening of metagenomic libraries. Appl Environ Microbiol 75(8):2506–2516. https://doi.org/10.1128/AEM.02136-08
Weng M, Deng X, Bao W, Zhu L, Wu J, Cai Y, Jia Y, Zheng Z, Zou G (2015) Improving the activity of the subtilisin nattokinase by site-directed mutagenesis and molecular dynamics simulation. Biochem Biophys Res Commun 465:580–586. https://doi.org/10.1016/j.bbrc.2015.08.063
**n X, Ambati RR, Cai Z, Lei B (2018) Purification and characterization of fibrinolytic enzyme from a bacterium isolated from soil. 3 Biotech 8:90. https://doi.org/10.1007/s13205-018-1115-4
Xu W, Shao R, Wang Z, Yan X (2015) Improving the neutral phytase activity from Bacillus amyloliquefaciens DSM 1061 by site-directed mutagenesis. Appl Biochem Biotechnol 175(6):3184–3194. https://doi.org/10.1007/s12010-015-1495-4
Yang X, Cong H, Song J, Zhang J (2013) Heterologous expression of an aspartic protease gene from biocontrol fungus Trichoderma asperellum in Pichia pastoris. World J Microbiol Biotechnol 29(11):2087–2094
Yang LQ, Chen GY, Li Y, Zhang RP, Liu SQ, Sang P (2019) Insight derived from molecular dynamics simulation into dynamics and molecular motions of cuticle-degrading serine protease Ver112. J Biomol Struct Dyn 37:2004–2016. https://doi.org/10.1080/07391102.2018.1471418
Yang H, Liu Y, Ning Y, Wang C, Zhang X, Weng P, Wu Z (2020) Characterization of an intracellular alkaline serine protease from Bacillus velezensis SW5 with fibrinolytic activity. Curr Microbiol 77:1610–1621. https://doi.org/10.1007/s00284-020-01977-6
Yusuf I, Ahmad SA, Phang LY et al (2019) Effective production of keratinase by gellan gum-immobilised Alcaligenes sp. AQ05-001 using heavy metal-free and polluted feather wastes as substrates. 3 Biotech 9:32. https://doi.org/10.1007/s13205-018-1555-x
Zanutto-Elgui MR, Vieira JCS, do Prado DZ, Buzalaf MAR, de Magalhães PP, De Oliveira DE, Fleuri LF (2018) Production of milk peptides with antimicrobial and antioxidant properties through fungal proteases. Food Chem. https://doi.org/10.1016/j.foodchem.2018.11.119
Zhang XH, Tee LY, Wang XG, Huang QS, Yang SH (2015) Off-target effects in CRISPR/Cas9-mediated genome engineering. Mol Therapy-Nucleic Acids 4:e264. https://doi.org/10.1038/mtna.2015.37
Zhang K, Duan X, Wu J (2016a) Multigene disruption in undomesticated Bacillus subtilis ATCC 6051a using the CRISPR/Cas9 system. Sci Rep 6:27943. https://doi.org/10.1038/srep27943
Zhang RX, Gong JS, Dou WF et al (2016b) Production and characterization of surfactant-stable fungal keratinase from Gibberella intermedia CA3-1 with application potential in detergent industry. Chem Pap 70:1460–1470. https://doi.org/10.1515/chempap-2016-0086
Zhang S, Xu Z, Sun H, Sun L, Shaban M, Yang X et al (2019) Genome-wide identification of papain-like cysteine proteases in Gossypium hirsutum and functional characterization in response to Verticillium dahliae. Front Plant Sci 10:134. https://doi.org/10.3389/fpls.2019.00134
Zhao G, Ding L-L, Pan Z-H, Kong D-H, Hadiatullah H, Fan Z-C (2019) Proteinase and glycoside hydrolase production is enhanced in solid-state fermentation by manipulating the carbon and nitrogen fluxes in Aspergillus oryzae. Food Chem 271:606–613. https://doi.org/10.1016/j.foodchem.2018.07.199
Zhao Y, Yu X, Li J (2020) Manipulation of immune-vascular crosstalk: new strategies towards cancer treatment. Acta Pharm Sin B 10:2018–2036. https://doi.org/10.1016/j.apsb.2020.09.014
Zhou C, Qin H, Chen X, Zhang Y, Xue Y, Ma Y (2018) A novel alkaline protease from alkaliphilic Idiomarina sp. C9–1 with potential application for eco-friendly enzymatic dehairing in the leather industry. Sci Rep 8:16467. https://doi.org/10.1038/s41598-018-34416-5
Zhou C, Zhou H, Li D, Zhang H, Wang H, Lu F (2020) Optimized expression and enhanced production of alkaline protease by genetically modified Bacillus licheniformis 2709. Microb Cell Fact 19:45. https://doi.org/10.1186/s12934-020-01307-2
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Solanki, P., Putatunda, C., Kumar, A. et al. Microbial proteases: ubiquitous enzymes with innumerable uses. 3 Biotech 11, 428 (2021). https://doi.org/10.1007/s13205-021-02928-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13205-021-02928-z