
Abstract
Background
Current animal tracking studies are most often based on the application of external geolocators such as GPS and radio transmitters. While these technologies provide detailed movement data, they are costly to acquire and maintain, which often restricts sample sizes. Furthermore, deploying external geolocators requires physically capturing and recapturing of animals, which poses an additional welfare concern. Natural biomarkers provide an alternative, non-invasive approach for addressing a range of geolocation questions and can, because of relatively low cost, be collected from many individuals thereby broadening the scope for population-wide inference.
Methods
We developed a low-cost, minimally invasive method for distinguishing between local versus non-local movements of cattle using sulfur isotope ratios (δ34S) in cattle tail hair collected in the Greater Serengeti Ecosystem, Tanzania.
Results
We used a Generalized Additive Model to generate a predicted δ34S isoscape across the study area. This isoscape was constructed using spatial smoothers and underpinned by the positive relationship between δ34S values and lithology. We then established a strong relationship between δ34S from recent sections of cattle tail hair and the δ34S from grasses sampled in the immediate vicinity of an individual’s location, suggesting δ34S in the hair reflects the δ34S in the environment. By combining uncertainty in estimation of the isoscape, with predictions of tail hair δ34S given an animal’s position in the isoscape we estimated the anisotropic distribution of travel distances across the Serengeti ecosystem sufficient to detect movement using sulfur stable isotopes.
Conclusions
While the focus of our study was on cattle, this approach can be modified to understand movements in other mobile organisms where the sulfur isoscape is sufficiently heterogeneous relative to the spatial scale of animal movements and where tracking with traditional methods is difficult.
Similar content being viewed by others

Background
Movement is a fundamental characteristic of life [1], yet its quantification across individuals and populations has remained a major methodological challenge in the field of movement ecology. Common animal tracking techniques such as GPS and radio transmitters [2] are time-intensive to collect, require expensive equipment and pose welfare concerns [3], and therefore may not be available in all settings nor necessary for addressing certain questions. For instance, characterizing population-level variation in movement patterns, such as the proportion of residents versus migrants, is likely to be inaccurate if one can only tag a handful of individuals over a relatively short period of time [4]. Forensically recreating movement paths from dead animals, or studying historical connectivity from archived specimens, or studying landscape connectivity, can provide useful insights into the drivers of population dynamics. There are few techniques currently available for retrospective animal movement tracking that allow the characterization of population-wide movements. The use of intrinsic markers to infer location of animals [5] offers a relatively low-cost, noninvasive solution to study animal movement patterns.
Intrinsic markers are natural biological or biogeochemical tags that can be retrieved from animals’ tissues [5]. Biogeochemical markers are particularly promising for studying movements because they form links between seasons and across populations, and they give time-integrated information which can directly be linked to geographical regions [5]. Stable isotopes of key elements, particularly hydrogen (δ2H) and sulfur (δ34S), are popular geolocators [5]. Hydrogen isotopes are used to make inferences of long-distance migration because they vary over latitudinal and continental gradients, reflecting local precipitation patterns [6]. Sulfur isotopes have been popular in movement studies such as tracking distance to the sea [7] and in dietary studies for making inferences about marine and marsh food webs [8]. Sulfur isotope fractionation between soil and plants [9,10,11] is around − 2‰ and from − 1‰ to + 2‰ between animal diet and different tissues [12,13,14,15,16], suggesting that sulfur is a useful geolocator in animal tissue because it is largely reflective of δ34S of the local geology [12, 17]. There are only two published values of the offset between diet and mammalian keratin: − 1‰ [12] and 1.2 ± 0.3 ‰ [18], with the latter study suggesting that offsets are higher for hair than other tissues. The combination of inert biological material that acts as a natural biologger for geolocating animals and continuously growing tissue that does not erode easily, can provide unique time series information about animal movement. This is because as individual animals move across distinct soils or between food webs, tissues that grow continuously (e.g. hair) retain the isotopic signatures of their present and previous feeding locations [17], potentially enabling ecologists to infer movement patterns from them.
This study demonstrates how variation in δ34S along cattle tail hair can be used to study animal movement retrospectively. We first assess the variation of δ34S across environmental space in the Serengeti landscape, and develop a sulfur isoscape for the ecosystem. We test the hypothesis that variation in δ34S in grass samples is reflected in sections of the most recent growth of tail hair, which indicates whether tail hair has the potential to be used as a natural bio-logger of geolocation in cattle. Lastly, we establish the distribution of travel distances across the Serengeti ecosystem sufficient to detect movement using sulfur stable isotopes. Agropastoral cattle provide an ideal system for develo** these methods because 1) these animals often move long distances; 2) their movements play important roles in human-wildlife conflict and the epidemiology of livestock diseases; 3) cattle owners can help verify the animal’s movement history; and 4) cattle are easier and cheaper to capture and sample than wild animals.
Materials and methods
Study area
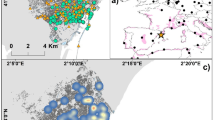
The study was conducted in the Serengeti ecosystem, Tanzania, (Fig. 1a), a landscape that is approximately 25,000 km2 in area [19, 20] and located between 34o and 36o E, and 1o and 3o N [21]. The ecosystem is characterized by a subtropical climate, with a dry and relatively cool season from late May to August, and a warmer dry season from September to October. Rainfall is highly variable but normally peaks in December, and between March and May [21, 22]. The area’s savanna vegetation is strongly influenced by soil type and rainfall [22] (Fig. 1a). The ecosystem is home to a diverse assemblage of both wild and domestic ungulates, including the largest terrestrial mammal migration in the world [23]. The soils underlying the ecosystem are highly heterogeneous and largely volcanic in nature [24]. The Eastern portion of the ecosystem, including the Serengeti plains, comprises alkaline soils derived from tephra deposited more than twenty thousand years ago from Ngorongoro rift region and more recently from the Oldonyo Lengai volcano [25], the world’s only active carbonatite volcano [26, 27]. The Western portions of the ecosystem are dominated by older alluvial soils, formed from the erosion of Precambrian volcanic rocks and banded ironstones [25, 27, 28]. Other parts of the ecosystem including north of the Mara river and crater highlands are dominated by heavily leached soils from older parent materials including complex granite, and volcanic rocks [25, 29, 30] (Fig. 1c). In addition to a number of protected areas, such as Serengeti National Park in Tanzania and the Masai Mara National Reserve in Kenya [31], the ecosystem supports a large human population consisting of pastoralists and agro-pastoralists living in close proximity to protected areas boundaries.

a The mean annual precipitation (b) the interpolated sulfur isoscape based on the output of a spatial GAM and (c) the underlying geology of the area. The boundaries of protected areas in the Serengeti ecosystem are illustrated with black lines. Sampling locations for (a) grass and (b) cattle are illustrated with black points
Data collection
All data collection including questionnaires, hair and grass samples was conducted from July 2017 to April 2018.
Environmental sampling
We collected grass samples from across the ecosystem to establish the spatial pattern of sulfur in the landscape. Grass samples were collected from 20 randomly selected villages bordering the Serengeti National Park but still within the Serengeti ecosystem (Fig. 1a). A total of 116 grass samples were collected and analyzed for δ34S to create a landscape-level isoscape. To collect grass samples, a 4 × 4 m plot was laid out and five sub samples (one from each corner and one at the middle) in 25 × 25 cm quadrats were clipped to ground level and pooled together to make one single sample from each site [32]. In addition, grass samples were collected (using the same sampling method) from each village where cattle were sampled to compare local versus landscape level variation. At each village, grass samples were collected from three random points located in non-cultivated grazing fields and at least 100 m away from roads and rivers. This minimized any potential sulfur contamination from industrial fertilizers, vehicle exhaust and road dust. Collected grass samples were kept in a paper envelope and stored in an open area at room temperature to prevent microbial activities and fungal development.
Cattle hair sampling
In each of the 20 villages where grass samples were collected, between one and three cattle from random households were sampled for tail hairs, making a total of 46 tail hair samples. Tail hair samples were obtained by pulling hairs from the base of the tail of each animal. Pulling helped to remove the entire hair root, which represents the most recent feeding history of an animal. Cattle age, sex and color were recorded during sampling. After hair collection, all hairs from an individual were aligned by root to standardize time zero (i.e. the most recent time), tied together and stored at room temperature [33, 34].
Questionnaire
A set of eight close-ended questions (See Additional information 1: Appendix 1) focused on exploring the movement history of cattle was posed to each cattle owner in relation to the sampled cattle. Questionnaires to cattle owners provided supplementary information on whether cattle had moved beyond the grazing area of the village or had been recently purchased from a neighboring village during the 5 months period prior to hair sampling. In addition to this binary response variable (moved versus not moved), these questionnaires also provided an estimate of distance travelled based on descriptions of the villages from which cattle had reportedly been moved.
Sample preparation for stable isotope analysis
For grass samples, all non-grass species and debris were removed from samples prior to analysis. Grasses were thoroughly washed in double distilled water (DDS) to remove any soil. Grass samples were oven dried at 60 °C for 48 h, pulverized into a fine powder and weighed (6.1–6.5 mg) into tin capsules [32], ready for isotopic analysis.
The range of hair bundles’ length varied between 10 and 25 cm for adult cattle and between 5 and 7 cm for calves. The growth rate of cattle tail hair has been estimated to be 0.76 mm per day [35]. In this study, tail hairs were sectioned into 10 × 8 mm segments representing 105 days of growth for adult cattle and 5–9 × 8 mm segments representing 52 to 95 days for calves. All samples were thoroughly washed in 2:1 chloroform:methanol to remove the impurities [36], and then rinsed with DDS to remove the remnants of solvent. This process was repeated twice to ensure all possible contaminants had been removed from the samples. Samples were then oven dried, ground to powder and weighed (1.0–1.3 mg) into tin capsules as described above.
Stable isotope ratio analysis
All laboratory analyses for stable isotope ratios were performed at the NERC Life Sciences Mass Spectrometry Facility hosted by the Scottish Universities Environmental Research Centre (SUERC). All sample analyses were undertaken using a Pyrocube elemental analyser (Elementar nalysensysteme, Langenselbold, Germany) coupled to a VisION isotope ratio mass spectrometer (Elementar UK, Cheadle Hulme, Stockport, UK). Laboratory standards methanesulfonamide/Gelatine (MSAG2), methionine/alanine/glycine/gelatine (M2) and sulfanilamide/alanine/gelatine (SAAG2) were repeated after every 10 samples and were used to correct for linearity and instrument drift over a 72-h analytical run. The isotope ratios for lab standards are determined relative to a range of international standards from IAEA (Vienna, Austria) and USGS (Reston, VA, USA). The analytical precision for sulfur isotopes was better than 0.7‰. The isotope ratios are expressed in the delta (δ) notation in parts per million (‰): δX = [(Rsample/Rstandard)-1] where X = 34S and R = the ratio of 34S/32S isotopes in a given sample compared with V- CDT (Vienna - Canyon Diablo Troilite).
Sulfur isoscape
To develop a sulfur isoscape for Serengeti, we fitted a spatial Generalized Additive Model (GAM) using the mgcv package [37, 38]. The fitted GAM of sulfur isotope ratios was modeled as a function of spatial and environmental variables including mean annual precipitation (MAP), Normalized Difference Vegetation Index (NDVI), soil exchangeable bases (CEC), the underlying geology (geology layer), elevation and the longitude and latitude. Longitude and latitude were modelled as a tensor product smoother to allow for construction of a single model matrix with multiple penalties [38]. All environmental variables were prepared in R [39] using the sp, raster, rdgal and rgeos packages [40,41,42,43,44]. To characterize mean annual rainfall (MAP), we averaged African monthly data from The Climate Hazards Group Infrared Precipitation with station data (CHIRPS) 1981 and 2018 [45, 46]. CHIRPS integrates 0.05° resolution satellite imagery with in-situ station data to create gridded rainfall time series for trend analysis and seasonal drought monitoring [45]. The underlying geology and parent material for different soil types across the Serengeti ecosystem were characterized from the Minerogenic Map of Tanzania layer, from Geological Survey of Tanzania [30]. The underlying geology was classified using lithology (Fig. 1c) of the parent material. Model selection was based on Akaike Information Criterion (AIC, weight = 0.99) (See Additional file 1: Appendix 2) from the stats package [47]. Our final model included lithology as the main environmental predictor and longitude and latitude as a tensor product smoother. The model was analyzed for goodness of fit with the gam.check function from mgcv package [38] and was subsequently used to predict sulfur isotope values (δ34SL, j) across the entire ecosystem (i.e. the ‘isoscape’) together with the standard deviation that captured the prediction uncertainty (σL). The predicted δ34S isoscape was at the same spatial resolution as the geology layer (5 km2 per pixel).
Validation of δ34S methodology
To understand whether δ34S in the ith tail hair sample from the jth individual (δ34S T,i,j) linearly reflects the local δ34S signature in vegetation (δ34SL,j), and to account for multiple observations of tail hair from the same individual, and that the true values of explanatory variable (ML,j) are latent, and only observed with error (σM) we constructed a latent ‘error in variables’ model wherein:
The model was fitted in Stan (Stan version 2.23, [48] using the R interface RStan version 2.19), using 3 chains for 10,000 iterations after 5000 as warmup, thinning to generate 3000 posterior samples per parameter. We used weakly informative priors [49] for all parameters: for regression coefficients we used t distributions with 3 degrees of freedom and for standard deviations half t with 3 degrees of freedom, except for σM where the prior was modelled as N(σL, 0.25). We used the fitted model to generate predictions and associated uncertainty in these predictions for each observation from the most recent segment of tail hair.
Studying movements using δ34S across the Serengeti sulfur isoscape
Relocation data from 49 GPS collared cattle in Western Serengeti (Ekwem, 2020 [50];) suggested that cattle rarely moved farther than 5 km from their home bomas (i.e. only 1.7% of relocations). Therefore, we expected longer distance movement to be relatively rare. To determine how far an animal would need to travel in order to robustly detect movement from δ34S tail hair signatures, we identified 50,000 random pairs of points across the isoscape, predicted δ34SL for each point, and estimated the mean distance an animal needed to move in order to detect statistically significant differences in isotope values in its tail hair, given the propagated uncertainty in predicting values of δ34SL from the landscape data, and δ34ST from δ34SL. We then compared the output with our actual distances travelled by cattle established from the questionnaire (above).
Results
Variation of sulfur stable isotope ratios across the Serengeti ecosystem
δ34S values of grass range between + 2.82 ‰ and + 13.04 ‰ (See Additional file 1: Appendix 3), consistent with the terrestrial nature of the ecosystem [51]. Our final predicted model of δ34S values in Serengeti included lithology as the main environmental predictor, and the latitudes and longitudes and their interaction as spatial smoothing parameters (AIC weight = 0.99: See Additional file 1: Appendix 2). From the GAM, we identified the following statistically significant relationships; a) a positive relationship between sulfur isotope ratios and pyroclastic-alkaline volcanic lavas (β = 0.079 ± 0.029, t = 2.699, p = 0.008) and b) a negative relationship between sulfur isotope ratios and volcanic ash/tephra (β = -0.05 ± 0.023, t = 2.175, p = 0.032). Other relationships that were not statistically significant from the model included: a negative relationship between sulfur isotopes ratios and granitoids (β = -0.021 ± 0.015, t = 1.345, p = 0.181) and between sulfur isotope ratios and volcanic lava (β = -0.016 ± 0.017, t = 0.975, p = 0.332), as well as a positive relationship between sulfur isotope ratios and mafic volcanic meta-basalts (β = 0.032 ± 0.021, t = 1.533, p = 0.128). The mean standard deviation of predictions estimated directly from the GAM was σL = 1.00, and inferred from the full model, σM = 1.810.
Relationship between cattle locations and tail hair isotope values
The ‘error in variables’ model showed good convergence and effective sample sizes for all posteriors (Rhat < 1.01, neff > 1400). The δ34S values of grasses from locations where tail hair was sampled, and the most recent tail hair section were strongly and positively related (Fig. 2) with slope β = 1.736 (95% credible interval (CIs) 1.466–2.058) confirming that fractionation of δ34S in the hair reflects the δ34S in the surrounding grass [7, 12]. Furthermore, the intercept (i.e. the baseline fractionation rate between grass to herbivore tissue) was − 4.670 (95% CIs − 7.563 - -2.279) coincident with the estimated range of fractionation rates from previous studies [12, 18]. With segment-level standard deviation of σε = 0.64, and individual level standard deviation of σα = 0.292, segment level variation in δ34ST accounted for 83% of overall variation in recorded δ34ST values.

The observed relationship between δ34S in grass and the most recent segment of the cattle tail hair, suggesting that δ34S in the hair reflects the δ34S in the landscape and can be used as a reliable biomarker of location. Red points (slightly ‘jittered’ for clearer visualization), show the most recent segment of a tail hair (i.e. the root), and black points show the rest of the segments in the tail hair
Power analysis result
The standard deviation on predicting tail hair δ34S values from the isoscape was typically very close to 1 (range 0.987–1.184). Our power analysis results suggested that an animal needed to move typically about 100 km (Fig. 3) in order to generate a difference in tail hair δ34S values likely to be judged significant. However, the nature of our sulfur isoclines across the Serengeti isoscape suggests effective distance is not equal in all cardinal directions. For instance, animals moving in a north-east to south-west pattern would in general have to move less than animals moving due north or south or east-west for the movement to be reliably detectable using δ34S (Fig. 3). Additionally, a questionnaire report suggested that our sampled cattle had not moved enough distance to establish their movement pattern using sulfur stable isotopes ratios, because the highest distance travelled by the cows was 52 Km with the majority having travelled less than 10 Km (See Additional file 1; Appendix 4).

Black polygon shows mean distance required to move to detect movement across the Serengeti isoscape in different directions; green lines show the actual distance travelled by our cattle; red points correspond to distances and directions in which sulfur values in tail hair are not predicted to change significantly (based on comparing two segments, P > 0.05), while blue points correspond to distances and directions in which movement is predicted to result in a statistically significant change in sulfur tail values (P < 0.05)
Discussion
The application of sulfur isotope ratios to quantify the movement of animals delivers considerable benefits. The major findings from our study suggest that: 1) sulfur isotope ratios in the vegetation across our study area vary across lithology; 2) sulfur isotope ratios in the vegetation are accurately reflected in cattle tail hairs (Fig. 2), and therefore 3) variation in tail hair could be used to potentially identify non-local movements of animals.
Feeding studies for grazing animals have derived fractionation factors of − 1‰ [12] and 1.2 ± 0.3 ‰ [18] between fodder and animal keratin. This is a relatively small change when compared with the range of values we have from grass samples across the Serengeti ecosystem and indicates the potential of sulfur isotope values as a dietary tracer within this system. The mean distance required to detect movement is about 100 km and modest compared to how far cattle or other animal species can move across this system and beyond. For example, pastoralist cattle can be transported by lorry many hundreds of kilometers across the country, and wildebeest which undertake a routinely cyclic migratory pattern every year can move about 250 km from southern to northern part of the ecosystem. The effectiveness of δ34S in studying movements for animals moving less than this distance in the Serengeti ecosystem would be dependent on local isotopic gradients.
Variation of sulfur stable isotope ratios across the Serengeti ecosystem
The major source of sulfur in terrestrial plants is from soil, which is mostly derived from bedrock weathering in the form of sulphate [52]. In addition, the δ34S of soil is influenced by local lithology and rainfall [13, 51], aerobic and anaerobic growing conditions [5], microbial processes [13, 16], fertilization procedures, as well as atmospheric deposition including the sea-spray effect [53]. Normally, the δ34S value for bedrock varies with rock type and age [16, 54], and terrestrial plants exhibit a wide range of δ34S values between ∼ − 10 and + 35 ‰ [18, 53, 55]. In the Serengeti ecosystem, grass samples have δ34S values of 2.82‰ to + 13.04‰ (See Additional file 1; Appendix 3), midway in the global range. The lowest δ34S values in the Serengeti were observed on the eastern side of the ecosystem (Fig. 1b), which is dominated by volcanic ash tephra. The highest δ34S values in the isoscape come from the extreme south-eastern side of the Serengeti ecosystem beyond the Ngorongoro rain shadow (Fig. 1b) which is characterized by soils from pyroclastic-alkaline and volcanic lavas. The areas with the strongest gradients in the isoscape are south-eastern and north-western parts of the ecosystem (Fig. 1b), suggesting that animal movement across these isoclines could be detected over relatively short distances. In addition, the areas exhibit intermediate levels of mean annual precipitation of 800-1000 mm [56] (Fig. 1a) providing plants growing in the area with an important nutrient for their growth.
GAM results suggest the high values of sulfur in this region are mainly correlated with lithology and rainfall. However, microbial processes and volcanic activity in the Rift Valley [57] likely contribute to these high δ34S values. For example, the southern Serengeti plains are composed of tephra from volcanic eruptions from Ngorongoro highlands and other adjacent volcanoes including Lemagurut and Olmoti [58]. Volcanic gases and rocks have a wider range of δ34S values caused by inorganic chemical reactions, and this can change the δ34S signature of the soil [54, 59]. Aerosols and dust from volcanic fumaroles and eruptions can be dispersed by both prevailing winds and rainfall. The potential to include environmental variables in predicting sulfur variation, offers a useful predictive power to the isoscape, suggesting the method could be applied to other landscapes, particularly where there is soil heterogeneity and geological gradients.
The nature of the Serengeti sulfur isoscape suggests that sulfur isotopes ratios are relatively invariant across the centre and more heterogeneous across North East (NE) and South West (SW) gradients (Fig. 1b). Such a pattern indicates that an animal needs to travel longer distances along the East-West direction than NE-SW direction to detect movement (Fig. 3).
Sulfur stable isotopes in ecological studies
This study has demonstrated the applicability of sulfur stable isotopes ratios in studying the movement of animals. Historically, sulfur stable isotopes have been difficult and expensive to analyse compared to carbon and nitrogen [16, 60, 61], which limited their applications to different studies [62]. The recent advances in continuous-flow isotope-ratio mass spectrometry (CF-IRMS) have enabled sulfur isotopes to be measured and analysed from both organic and inorganic materials in relatively small amounts [12, 63, 64]. This has increased their applicability in different fields of study including archaeology and ecology [12].
Low variation in δ34S values of our cattle tail hairs (Fig. 4) suggests that cattle either foraged in a single location or foraged within an isotopically similar region of the isoscape (i.e. along the same isocline). Conversely, high variation in δ34S values in the tail hair (Fig. 4) should be taken as a primary indicator of movement or supplementary feeding using forage grown in a different isotopic setting [7]. This finding agrees with other recent works including that of Zazzo et al. [7], which revealed changes in δ34S in hair following the movement of sheep relative to the proximity to the sea.

Variation of δ34S across length of tail hairs for representative individual cattle in Serengeti, showing sulfur isotopes profiles for reportedly moved (with their associated distance moved. A1 = 10.607 Kms, A2 = 13.636 Kms, A3 = 3.79 Kms and A4 = 9.318 Kms) and animals that did not move
Estimating movement and wildlife home ranges of domestic and wild grazers
This study provides a template for understanding animal movement in and outside the Serengeti ecosystem. The question of where, how, and when livestock move remains critical for informing policies aimed at maximizing livestock productivity, minimizing disease spread while also maintaining traditional pastoralist livelihoods. Quantifying livestock movements, such as those occurring in remote agro-pastoralist settings, or through illegal transboundary trade or theft, is challenging to study using other methods. Our approach potentially provides a non-invasive way to infer whether an animal has moved long-distances across sulfur gradients in the previous ~5 months. The same methodology could be applied in other areas with sufficient variation in δ34S values where tail hair can be readily sampled (e.g. from carcasses) to study wide-ranging wildlife species (e.g. wildebeest and zebra). For example, this method could distinguish between migratory and non-migratory individuals and establish the frequency of different movement strategies in a partially migratory system such as Serengeti.
Conclusions and recommendations
This study has shown the potential for using sulfur stable isotopes ratios in studying movement ecology of herbivores. For example, the δ34S isoscape of the Serengeti provides baseline information on how δ34S can be applied to understand spatial and movement ecology of livestock across the landscape. However, the use of a single element, such as δ34S does not capture all the isotopic variation across the landscape and could limit our ability to recreate detailed movement patterns of individual animals. The inclusion of additional isotope tracers such as strontium (87Sr/86Sr), which tends to have a well-defined geological distribution [65], or δ2H which tends to differ by watershed [6], could improve the technique by adding isotopic axes from which the location can be more accurately triangulated. Future studies could incorporate data from other tracking techniques, such as GPS telemetry or genetic markers, to further calibrate these isotopic techniques which would ultimately improve our ability to forensically determine the movement history of unmarked animals.
Availability of data and materials
All data used in the development of this manuscript can be found here: https://doi.org/10.5525/gla.researchdata.1065.
References
Nathan R, Getz WM, Revilla E, Holyoak M, Kadmon R, Saltz D, et al. A movement ecology paradigm for unifying organismal movement research. PNAS. 2008;105(49):19052–9.
Coppolillo PBP. The landscape ecology of pastoral herding: spatial analysis of land use and livestock production in East Africa. Hum Ecol. 2000;28(4):527–60.
Bailey DW, Trotter MG, Knight CW, Thomas MG. Use of GPS tracking collars and accelerometers for rangeland livestock production research. Transl Anim Sci. 2017;2(1):81–8.
Hurme E, Gurarie E, Greif S, M LGH, Flores-martínez JJ, Wilkinson GS, et al. Acoustic evaluation of behavioral states predicted from GPS tracking: a case study of a marine fishing bat. Mov Ecol. 2019;7(21):1–14.
Rubenstein DR, Hobson KA. From birds to butterflies: animal movement patterns and stable isotopes. Trends Ecol Evol. 2004;19(5):256–63.
Hobson KA. Tracing origins and migration of wildlife using stable isotopes: a review. Oecologia. 1999;120(3):314–26.
Zazzo A, Monahan FJ, Moloney AP, Green S, Schmidt O. Sulphur isotopes in animal hair track distance to sea. Rapid Commun Mass Spectrom. 2011;25(17):2371–8.
Krouse HR. Sulfur isotope studies of the pedosphere and biosphere. In: Rundel PW, Ehleringer JR, Nagy KA, editors. Stable isotopes in ecological research. New York: Springer Berlin Heidelberg; 1988. p. 424–44.
Harrison SM, Schmidt O, Moloney AP, Kelly SD, Rossmann A, Schellenberg A, et al. Tissue turnover in ovine muscles and lipids as recorded by multiple (H, C, O, S) stable isotope ratios. Food Chem. 2011;124(1):291–7.
Trust BA, Fry B. Stable Sulphur isotopes in plants: a review. Plant Cell Environ. 1992;15(9):1105–10.
Novák M, Bottrell SH, Přechová E. Sulfur isotope inventories of atmospheric deposition, spruce forest floor and living sphagnum along a NW-SE transect across Europe. Biogeochemistry. 2001;53:23–50.
Richards MP, Fuller BT, Sponheimer M, Robinson T, Ayliffe L. Sulphur isotopes in Palaeodietary studies: a review and results from a controlled feeding experiment. IntJOsteoarchaeol. 2003;45(13):37–45.
Peterson BJ, Fry B. Stable isotopes in ecosystem studies. Annu Rev Ecol Sytematics. 1987;18:293–320.
Krajcarz MT, Krajcarz M, Drucker DG, Bocherens H. Prey-to-fox isotopic enrichment of 34S in bone collagen: implications for paleoecological studies. Rapid Commun Mass Spectrom. 2019;33(16):1311–7.
McCutchan JH, Lewis WM, Kendall C, McGrath CC. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos. 2003;102:378–90.
Peterson BJ, Howarth RW. Sulfur, carbon, and nitro- gen isotopes used to trace organic matter flow in the salt- marsh estuaries of Sapelo Island, Georgia. LimnolOceanogr. 1987;32:1195−213.
Richards MP, Fuller BT, Hedges REM. Sulphur isotopic variation in ancient bone collagen from Europe: implications for human palaeodiet, residence mobility, and modern pollutant studies. Earth Planet Sci Lett. 2001;191(3):185–90.
Webb EC, Newton J, Lewis J, Stewart A, Miller B, Tarlton JF, et al. Sulphur-isotope compositions of pig tissues from a controlled feeding study. Sci Technol Archaeol Res. 2017;3(1):71–9.
Emerton L, Mfunda I. Making Wildlife Economically Viable for Communities Living Around the Western Serengeti, Tanzania. Integration The Vlsi Journal. 1999;34.
Sinclair ARE, Metzger KL, Mduma SAR, Fryxell JM, Serengeti IV. Sustaining biodiversity in a coupled human-natural system. Sinclairl ARE, Metzger KL, Mduma SAR, Fryxell JM, editors. Chicago: Chicago University Press; 2015.
Sinclair EARE, Arcese P. Serengeti II: dynamics, management, and conservation of an ecosystem edited; 1995.
Mwakatobe A, Nyahongo J, Røskaft E. Livestock Depredation by Carnivores in the Serengeti Ecosystem. Environ Nat Resour Res. 2013;3(4).
Hopcraft JGC. Why are wildebeest the most abundant herbivore in the Serengeti ecosystem? In: Sinclair ARE, Metzger KL, Mduma SAR, Fryxell JM, editors. Serengeti IV. Chicago: University of Chicago Press; 2015. p. 9–11.
Reed DN, Anderson TM, Dempewolf J, Metzger K, Serneels S. The spatial distribution of vegetation types in the Serengeti ecosystem: the influence of rainfall and topographic relief on vegetation patch characteristics. J Biogeogr. 2009;36(4):770–82.
Krafft M, Keller J. Temperature measurements in Carbonatite Lava Lakes and flows from Oldoinyo Lengai, Tanzania. Science (80- ). 1989;245(4914):168–70.
Sinclair ARE, Hopcraft JGC, Olff H, Mduma SAR, Galvin KA, Sharam GJ. Historical and future changes to the Serengeti ecosystem. Serengeti III: human impacts on ecosystem dynamics. In: Sinclair ARE, Packer C, Mduma SAR, Fryxell JM, editors. Historical and future changes to the Serengeti ecosystem Serengeti III. Chicago: Chicago University Press, Chicago; 2008.
Gerresheim, K. The Serengeti Landscape Classification. Serengeti Research Institute Publication. Serengeti Ecological Monitoring Programme, African Wildlife Leadership Foundation. 1974;165. 26pp + colored map (1:125,000).
Sinclair AR E, Metzger Kristine L. Advances in wildlife ecology and the influence of Graeme Caughley. Wildl Res. 2009;36:8–15.
Scoon RN. Geology of National Parks of Central / Southern Kenya and Northern Tanzania: Geotourism of the Gregory Rift Valley, Active Volcanism and Regional Plateaus. Springer. 2018.
Leger C, Barth A, Falk D, Mruma AH, Magigita M, Boniface N, et al. Minerogenic map of Tanzania and explanatory notes for the Minerogenic map of Tanzania 1: 1.5 M. geological survey of Tanzania; 2015.
Reid RS, Galvin KA, Knapp EJ, Ogutu JO, Kelo DS. Sustainability of the Serengeti-Mara Ecosystem for Wildlife and People. Serengeti IV: Sustaining biodiversity in a coupled human-natural system. The University of Chicago Press; 2015.
Hellmann C, Rascher KG, Oldeland J, Werner C. Isoscapes resolve species-specific spatial patterns in plant-plant interactions in an invaded Mediterranean dune ecosystem. Tree Physiol. 2016;36(12):1460–70.
Horacek M, Burnik-Sturm M, Kaczensky P. First stable isotope analysis of Asiatic wild ass tail hair from the Mongolian Gobi. Erforsch Biol Ressourcen der Mongolei. 2012;12:85–92.
Rysava K, McGill RAR, Matthiopoulos J, Hopcraft JGC. Re-constructing nutritional history of Serengeti wildebeest from stable isotopes in tail hair: seasonal starvation patterns in an obligate grazer. Rapid Commun Mass Spectrom. 2016 Jul 15;30(13):1461–8.
Chen G, Schnyder H, Auerswald K. Model explanation of the seasonal variation of δ18O in cow (Bos taurus) hair under temperate conditions. Sci Rep. 2017;7(1):1–15.
Mekota AM, Grupe G, Ufer S, Cuntz U. Identifikation von Hungerepisoden mithilfe stabile isotope in Haarproben: Forensischer Ansatz an seriellen Haaranalysen. Rechtsmedizin. 2009;19(6):431–40.
Wood S. Generalized additive models: an introduction with R. J Am Stat Assoc. 2007;102(478):760–1.
Wood S. Generalized Additive Models. New York: Chapman and Hall/CRC; 2017.
R-Core-Team. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing; 2017.
Bivand R. Package “rgeos”; 2018.
Roger A, Rowlingson B, Sumner M, Hijmans R. Package ‘rgdal’ R topics documented; 2018. 0(March 2012).
Bivand RS, Pebesma EJ, Gomez-Rubio V. Applied spatial data analysis with R. applied spatial data analysis with R; 2013. p. 21–54.
Pebesma E, Bivand RS. S classes and methods for spatial data: the sp package. Econ Geogr. 2005;50(1):1–21.
Hijmans RJ. Introduction to the ‘raster’ package (version 2. 3–24). R-CRAN Proj; 20172008. p. 1–27.
Funk C, Peterson P, Landsfeld M, Pedreros D, Verdin J, Shukla S, et al. The climate hazards infrared precipitation with stations - a new environmental record for monitoring extremes. Sci Data. 2015;2:1–21.
Dinku T, Funk C, Peterson P, Maidment R, Tadesse T, Gadain H, et al. Validation of the CHIRPS satellite rainfall estimates over eastern Africa. Q J Roy Meteorol Soc. 2018;144(January):292–312.
SakamotoY, Ishiguro M, Kitagawa G. Akaike information criterion statistics. KTK Scientific Publishers; Dordrecht; Boston: D. Reidel; Hingham, MA: Sold and distributed in the U.S.A. and Canada by Kluwer Academic Publishers, Tokyo; 1986.
Carpenter B, Gelman A, Hoffman MD, Lee D, Goodrich B, Betancourt M, Brubaker M, Guo J, Li P, Riddell A. Stan: A probabilistic programming language. J Stat Softw. 2017;76(1).
Gelman A. Bayesian data analysis Gelman. J Chem Inf Model. 2013;53:1689–99.
Ekwem D. The determinants of foot-and-mouth disease virus circulation in livestock populations in northern Tanzania. University of Glasgow; 2020.
Coplen TB, Hopple J a, Böhlke JK, Peiser HS, Rieder SE, Krouse HR, Rosman KJR, Ding T, Vocke RD, Revesz KM, Lamberty A, Taylor P, De Bievre P. Compilation of minimum and maximum isotope ratios of selected elements in naturally occurring terrestrial materials and reagents. Usgs. 2002;110.
Krouse HR. Stable isotopes: natural and anthropogenic Sulphur in the environment. Scope (Kalamazoo). 1991;43:465.
Krouse HR, Mayer B. In: Cook PG, Herczeg AL, editors. Sulphur and oxygen isotopes in Sulphate BT - environmental tracers in subsurface hydrology. Boston: Springer US; 2000. p. 195–231.
Thode HG. Sulphur Isotopes in Nature and the Environment: An Overview. SCOPE. 1991;1–26.
Nehlich O, Richards MP. Establishing collagen quality criteria for Sulphur isotope analysis of archaeological bone collagen. Archaeol Anthr Sci. 2009;1:59–75.
Prins HHT, Loth PE. Rainfall patterns as background to plant phenology in northern Tanzania. J Biogeogr. 1988;15(3):451–63.
Ueda A, Sakai H. Sulfur isotope study of quaternary volcanic rocks from the Japanese Islands arc. Geochim Cosmochim Acta. 1984;48(9):1837–48.
McHenry LJ, Mollel GF, Swisher CC. Compositional and textural correlations between Olduvai Gorge bed I tephra and volcanic sources in the Ngorongoro volcanic highlands, Tanzaniar. Quat Int. 2008;178(1):306–19.
Puchelt H, Hoefs J, Nielsen H. Sulphur isotope investigations of the Aegean volcanoes. Acta First Int Sci Congr Volcano Thera, Athens. 1971;1969(15–23):303–17.
Hesslein RH, Capel MJ, Fox DE, Hallard KA. Stable isotopes of sulfur, carbon, and nitrogen as ndicators of trophic level and fish migration in the lower Mackenzie River basin, Canada. Can J Fish Aquat Sci. 1991;48:2258–65.
Connolly RM, Schlacher TA. Sample acidification significantly alters stable isotope ratios of sulfur in aquatic plants and animals. Mar Ecol Prog Ser. 2013;493(4):1–8.
Connolly RM, Guest MA, Melville AJ, Oakes JM. Sulfur stable isotopes separate producers in marine food-web analysis. Oecologia. 2004;138(2):161–7.
Richardson DM. Fifty Years of Invasion Ecology: The Legacy of Charles Elton. Blackwell publishing; 2011. p. 432.
Sayle KL, Cook GT, Ascough PL, Hastie HR, Einarsson Á, McGovern TH, et al. Application of 34S analysis for elucidating terrestrial, marine and freshwater ecosystems: evidence of animal movement/husbandry practices in an early Viking community around Lake Mývatn, Iceland. Geochim Cosmochim Acta. 2013;120(September):531–44.
Wassenaar LI. Introduction to Conducting Stable Isotope Measurements for Animal Migration Studies. In: Tracking Animal Migration with Stable Isotopes. Elsevier Inc.; 2019. p. 25–52.
Acknowledgements
The authors would like to acknowledge the SUERC stable isotope lab for their extensive feedback. The authors appreciate support from NM-AIST, TAWIRI, TANAPA, TAWA and the Serengeti Biodiversity Program during fieldwork and laboratory sample preparations. Special thanks to Alex Morton, Joseph Masoy, Majaliwa Masolele, Markus Borner, Sarah Cleaveland, Barbara Mable, Jonathan Lindsay, Neil Burns and all IBAHCM staff for their support in accomplishing this work.
Funding
This research has been funded by Karimjee Jivanjee Foundation and NERC ‘Grant-in-Kind’ support EK293–16/17. TAM, DH and JGCH were supported by the European Union’s Horizon 2020 research and innovation program under grant agreement 641918 African Bio Services.
Author information
Authors and Affiliations
Contributions
ZK, JGCH, TAM, RAM, JN and DH designed the study, ZK did field work, RAM and WLM did sample preparation and lab analysis, ZK, JGCH, TAM, RAM, JN, JMM and DH did statistical analyses. ZK, JGCH, TAM, RAM, JN, DH, WLM, LKM, DE and AL did the manuscript writing. The authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not Applicable
Competing interests
Authors Declare that no competing interest exist.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1 Appendix 1.
Questionnaires. List of questions asked to cattle owners for exploring movement history of cattle. Appendix 2: Summary statistics and model selection table. Appendix 3: Grass sulfur isotopes across the Serengeti ecosystem. Laboratory results for δ34S data used to interpolate the isoscape. Appendix 4: Distance in Km moved by cattle as per questionnaire report. Distance is the straight-line measure between two points.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kabalika, Z., Morrison, T.A., McGill, R.A.R. et al. Tracking animal movements using biomarkers in tail hairs: a novel approach for animal geolocating from sulfur isoscapes. Mov Ecol 8, 37 (2020). https://doi.org/10.1186/s40462-020-00222-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40462-020-00222-w