
Abstract
Multiple studies have shown that extracellular vesicles (EVs) play a key role in the process of information transfer and material transport between cells. EVs are classified into different types according to their sizes, which includes the class of exosomes. In comparison to normal EVs, tumor-derived EVs (TDEs) have both altered components and quantities of contents. TDEs have been shown to help facilitate an environment conducive to the occurrence and development of tumor by regulation of glucose, lipids and amino acids. Furthermore, TDEs can also affect the host metabolism and immune system. EVs have been shown to have multiple clinically useful properties, including the use of TDEs as biomarkers for the early diagnosis of diseases and using the transport properties of exosomes for drug delivery. Targeting the key bioactive cargoes of exosomes could be applied to provide new strategies for the treatment of tumors. In this review, we summarize the finding of studies focused on measuring the effects of TDE on tumor-related microenvironment and systemic metabolism.
Video Abstract
Similar content being viewed by others


Introduction
In the last few years, there has been increased interest in the roles and biological functions of extracellular vesicles (EVs). Exosomes are one type of EVs and are 30-100 nm in diameter [1]. The outer membrane of exosomes is a lipid bilayer that protects the exosome contents from various stimuli in the circulating fluid. This protection allows the exosomes and its contents to achieve long-distance transport in circulating body fluids [2]. Exosomes can contain multiple soluble bioactive substances, such as DNA, RNA, metabolites, lipids and proteins. The specific contents of the exosomes are determined by the cellular source. The content of EVs can be affected by biological factors such as age, gender, and race [3]. Exosome-mediated transfer of DNA, proteins, mRNA and noncoding RNA can lead to the phenotypic change in target cells, which can induce physiological or pathological states [4].
The formation process of exosomes is thought to begin with the establishment of intraluminal vesicles (ILVs), which accumulate in the lumen and then lead to the formation of multivesicular bodies (MVBs). While a proportion of MVBs are degraded by lysosomal fusion, MVBs could also bind to the plasma membrane and could be secreted extracellularly [5]. Exosomes can be secreted into the extracellular environment by a variety of cells, including tumor cells [6], to act on local or distal target cells [7]. Endosomal sorting complex required by ESCRT (endosomal sorting complex required for transport) machinery controls the biogenesis and formation of exosomes. The ESCRT is composed of five different proteins (ESCRT-0, ESCRT-I, ESCRT-II, ESCRT-III and the AAA ATPase Vps4 complex) [8, 9]. In addition to the ESCRT mechanism, sphingolipids ceramide [10], small GTPase ADP ribosylation factor 6 (ARF6) and its effector phospholipase D2 (PLD2) [11], the tetraspanin CD63 in melanocytes [12] can also directly regulate the biogenesis and material sorting of exosomes.
Exosomes act on receptor cells in three main ways: first, proteins on the exosome membrane directly contact with proteins on the receptor cell membrane to trigger intracellular signaling cascades; second, the contents of the exosome membrane are delivered to the receptor cells after fusion with the receptor cell membrane; and third, the target cells directly engulf exosomes [13]. Uptake of exosomes is not random but depends on the interaction between recipient cells and proteins on the surface of exosomes. Some reports have shown that adhesion related molecules on exosomal surfaces determine which cells can receive exosomes. These adhesion molecules include tetraspanins, glycoproteins, and integrins [14, 15]. As a bridge for material and information transfer between cells, exosomes play a key role in local and systemic cancer cell communications. Increasing evidences have shown that exosomes play an important role in tumor proliferation, metastasis, apoptosis, and resistance to drug therapy [16, 17].
In comparison to the normal cell, the tumor cell have an increased consumption of glucose and also undergo metabolism alterations required to sustain growth and reproduction in a limited nutritional environment. Even under oxygen-rich conditions, cancer cells have a much higher rate of glycolysis than tricarboxylic acid (TCA) cycle-mediated oxidative phosphorylation (OXPHOS), a phenomenon known as the “Warburg effect” [18]. Increased glucose uptake and enhanced glycolysis, as well as high lactate production under aerobic conditions, are all considered markers of tumors. The increased demands for lipids and cholesterol in highly proliferating cancer cells also lead to changes in lipid metabolism [19]. Similarly, amino acid metabolism is increased in cancer cells, particularly in the metabolism of the major energy substrates glutamine and serine [20]. Metabolism does not exist independently but acts in concert to provide fertile soil for tumor reproduction and growth.
Studies have shown that in order to survive in the complex tumor microenvironment (TME), tumor cells increase the number of secreted exosomes to actively construct conditions suitable for their growth [21]. Tumor cells release exosomes through various regulatory mechanisms to transmit signals to other cells that trigger subsequent cancer-promoting effects, which include signals that induce invasion, metastasis, angiogenesis, or defensive effects [22]. Exosomes play a key role as a link of information and material transfer between cells in the process of tumor metabolic changes. Therefore, this review focuses on the relationship between tumor-derived exosomes and tumor metabolism, body metabolism, and their impacts on the tumorigenesis and development. It is hoped that this can provide new diagnosis and treatment ideas and strategies for tumors.
Tumor-derived exosomes
Multiple cell types, including tumor cells, can secret exosomes. The exosomes secreted by tumor cells are called tumor-derived exosomes (TDEs). Studies have estimated that the blood from cancer patients contains twice as many exosomes as the blood from healthy individuals [23]. TDE can transport information and materials not only between tumor cells, but also between stromal cells and tumor cells. Stromal cells receive exosomes from tumor cells and generate a tumor-promoting microenvironment. In turn, exosomes secreted by stromal cells act on tumor cells to promote their proliferation and invasion [40]. Moreover, lactate, a glycolytic metabolite, has been shown to directly inhibit the cytolytic activity of NK cells and indirectly inhibit the function of NK cells by increasing the number of bone marrow derived suppressor cells (MDSCs) [63]. These findings suggest that immune suppression induced by metabolic recoding mediated by TDEs may be a potential trigger for tumor development.
Lipid metabolism
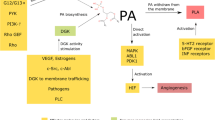
Exosome-mediated disruption of lipid metabolism is increasingly recognized as a feature of tumor cells and may be a key factor in the progression and metastatic behavior of malignant tumors [64]. Studies have shown that lipid metabolism disorders can upregulate oncogenes such as Mtor, cyclin-E, c-Jun, Notch, c-Myb, and c-Myc to promote tumor invasion and metastasis [65]. Exosome membranes contain many molecules which may include phosphatidic acid (PA), phosphatidyl inositol (PI), phosphatidyl ethanolamine (PE), phosphatidyl choline (PC), phosphatidyl serine (PS), ceramide, cholesterol, sphingomyelin, glycosphingomyelin, and other lipids in low abundance. Some have suggested that PS and PE appear to be involved in the biogenesis of exosomes [66]. The enrichment of specific lipids has been shown to significantly increased exosome membrane hardness. Moreover, these lipids exist in the outer membrane of exosomes and play a crucial role in the recognition and internalization of exosomes, enabling them to deliver metabolites to recipient cell [67]. Depletion of a cholesterol lipid efflux pump ABCG1 (ATP-binding cassette transporter G1), leads to the accumulation of EVs and their derivatives, thereby triggering tumor regression [68]. Tafelmeier et al. have demonstrated that ABCG1-mediated cholesterol efflux promotes exosome release, while SRB1-mediated cholesterol efflux inhibits exosome uptake by recipient cells [69].
The role of lipids in cell communication is an interesting emerging topic of research and is worth further investigation. Exosomes are known to carry bioactive lipids, such as prostaglandins and leukotrienes, which have been shown to promote the development of tumors [70]. Furthermore, Lydic et al. found that TDEs from colorectal cancer cell line LIM1215 have higher levels of glycerolipids, cholesterol, glycerol, and sphingolipids [27]. Others have shown that TDEs from prostate cancer cells are rich in phosphatidylserine, glycosphingolipids, sphingomyelin, and cholesterol [28]. High fat content appears to be more conducive to the uptake of TDEs by normal cells, inducing the transformation of normal cells into tumor cells [65, 71, 72].
It has been shown that tumor derived signaling molecules trigger lipolysis in cancer-associated adipocytes (CAAs), which results in lipoatrophy in humans [73], a form of cancer cachexia [74]. Phospho-hormone-sensitive lipase (P-HSL), a marker that activates lipolysis, was found at higher levels in TDEs from Lewis lung Cancer (LLC). Adipocytes exposed to TDEs from LLC showed lower levels of lipid droplets and higher levels of glycerol release [75]. Another study showed that TDEs from pancreatic cancer cells containing adrenomedullin (AM) interact with adrenomedullin receptors (ADMRs) in adipocytes and activate ERK 1/2 and MAPKs p38 signaling pathways to induce lipolysis via HSL phosphorylation [76]. Wang et al. found that TDEs from lung cancer could be internalized by human adipose tissue-derived MSCs and participate in the inhibition of adipogenesis of MSCs through TGFβ signaling pathway [77]. The effect between the two is reciprocal, TDEs regulate the metabolism of MSCs, and MSCs affected by TDEs secrete more exosomes as a kind of feedback to promote tumor angiogenesis [78]. Whether MSC-derived EVs promote or inhibit cancer seems to depend on the contents of cytokines and miRNA in exosomes [79,80,81]. Interestingly, lipids carried in exosomes have also been found to be important for inducing tumor drug resistance. For example, through regulating lipid metabolism, studies have shown that high expression of acid sphingomyelinase (ASM) by multiple myeloma (MM) derived exosomes can transfer drug-resistant phenotypes to drug-sensitive MM cells. The expression and protein level of ASM in MM cells and exosomes increased after antitumor drug stimulation, reflecting the tumor protective effect of ASM and promoting the occurrence of drug resistance [82].
Lipids in TDEs have also been shown to alter immune responses. For example, Jiang et al. found that overexpression of FASN (Fatty Acid Synthase) in ovarian cancer led to lipid accumulation in TME, resulting in T cell dysfunction, and then impaired anti-tumor immune responses [83]. It has also been shown that high cholesterol is more conducive for exosomes to bind to CD8 ( +) T cells, as the enrichment of cholesterol in cell membranes can improve the fluidity of cell membranes [84].
Amino acid metabolism and nucleotide metabolism
Studies on exosomes and amino acid metabolism mainly focus on tumor cells and CAFs and provides evidence that TDEs could induce CAFs production [85]. Liu et al. found that CAFs mainly regulated amino acid metabolism in an exosome-dependent manner in lung adenocarcinoma (LUAD) cells. Stimulated by tumor-derived proinflammatory cytokines, the specific long noncoding RNA LINC01614 secreted from CAFs, was up-regulated. CAF-derived exosomes could promote NF-κB activation through transport of LINC01614 to LUAD cells, which then interacted with ANXA2 and p65, leading to upregulation of glutamine transporters SLC7A5 and SLC38A2. Ultimately, LINC01614 enhanced glutamine uptake in LUAD cells [86]. Moreover, Zhao et al. found that exosomes secreted by CAFs could significantly inhibit electron transport chains after being absorbed by prostate and pancreatic cancer cells, thus increasing glutamine dependent reduction carboxylation [52]. The above studies suggest that exosomes play a key role in regulating the metabolism of glutamine. Furthermore, because glutamine is a nitrogen donor for nucleotide synthesis [87] TDEs effects on glutamine is likely to also affect nucleotide synthesis. Therefore, it could be speculated that TDEs may regulate the generation and degradation of nucleic acids by reacting to nucleotide metabolites and metabolic wastes. While the effects of TDEs on nucleotide metabolism is interesting, the current understanding about this biology is limited and further studies are warranted.
Effects on systemic metabolism
The normal state of the body is one of equilibrium and homeostasis. The development of neoplasms can upset this balance, for example emaciation is considered to be a manifestation of tumor cachexia. It has been shown that TDEs from pancreatic cancer could induce subcutaneous adipose tissue lipolysis through AM as a mediator, suggesting that TDEs induced lipolysis may be associated with weight loss in patients [76]. In addition, Fong et al. found that inhibition of exosomal miR-122 could normalize glucose metabolism of the brain, lung and other distal organs and reduce the incidence of tumor metastasis, indicating that exosomal miR-122 could reshape the whole body energy metabolism and promote tumor progression [58]. It has been found that TDEs from prostate cancer (PCa) containing PKM2 could be transferred to bone marrow stromal cells (BMSCs) and promote tumor metastasis by altering bone metabolism [88]. And It has been found that TDEs from PCa transfer LncRNA nuclear-enriched abundant transcript 1 (NEAT1) to human bone marrow-derived MSCs promote osteogenic differentiation, suggesting that this might be one of the reasons why patients with PCa often present with osteoblast bone metastases [89]. In summary, TDEs have been shown to have significant effects on the whole-body metabolism, although the whole-body metabolism is complex, and requires further exploration.
Recent studies have found that metabolism alterations caused by TDEs can affect the immune system which can aid in the ability of the tumor to escape the immune system. Studies have shown that TDEs can lead to increased glucose uptake by macrophages in the pre-metastasis niche through TLR2 and NF-κB signaling pathways, thus increasing the expression of PD-L1 and promoting the polarization of macrophages towards immunosuppressive phenotypes [90]. In addition, TDEs activate Tregs cells to form immunotolerant premetastatic niches by regulating the interaction of CCL1 + fibroblasts and CCR8 + Treg cells [91]. This provides immune conditions for tumor metastasis. Peroxisome proliferator activated receptor (PPAR) α response to fatty acids delivered by TDEs and leads to excess lipid droplet formation and enhanced fatty acid oxidation, ultimately leading to metabolic shift to mitochondrial oxidative phosphorylation, which drives dendritic cell immune dysfunction [92]. In summary, multiple studies have shown that TDEs mediated metabolism alterations can cause deleterious effects for the ability of the immune system to target tumor cells.
Application
Diagnostic biomarkers
A large number of tumor markers carried by cancer cell exosomes have set off a boom in liquid biopsy. The application of TDEs was shown in Fig. 2. Depending on the chemical, physical, and biological properties of exosomes, different methods have been used to isolate and purify exosomes, including ultra-centrifugation, size exclusion chromatography, ultrafiltration, and microplate-based magnetic immunocapture. Using multiple techniques based on membrane protein composition, size and density, rich exosome populations could be isolated from the various biological fluids and cell media [93]. The liquid biopsy strategy assesses factors including circulating tumor cells, circulating tumor DNA (ctDNA), EVs or exosomes, and other biochemical substances [94,95,96]. Since exosomes are detectable in almost all body fluids, they are more readily available, making them ideal biomarkers for monitoring dynamic intratumoral heterogeneity (ITH), enabling early detection and minimizing treatment side effects and toxicity [22, 97]. There has been a growing number of clinical trials related to exosomes in multiple cancer types, a summary of these trials are shown in Table 1.

The application of tumor-derived exosomes (TDEs). The application of tumor-derived exosomes (TDEs) is mainly divided into two kinds: as a biomarker for diagnosis and participating in treatment. Starting from the diagnostic biomarkers, the biomarkers related to diagnosis or prognosis should be determined first, and then the isolated and purified exosomes can be obtained by centrifugation, filtration and other methods, and substances required for detection in the exosomes can be detected to help diagnosis and treatment. From the perspective of treatment, the tumor-promoting function can be blocked by inhibiting the release of TDEs or inhibiting the absorption of TDEs by other cells. TDEs could also be used to load drugs or prepare tumor vaccines
Although the development of science and technology for exosome detection is growing, there are still many problems to be solved. First, each exosome assay technique is known to have its own bias in estimating exosome size. For example, Nanoparticle Tracking Analysis (NTA) is widely used for exosome size detection in bioparticle applications, and its detection limit for bioparticles is about 70 nm. NTA and Transmission Electron Microscopy are sensitive to different sizes [98]. Moreover, the process of exosome preparation may result in swelling, shrinkage, or obesity of the exosome, and these changes have a significant impact on true size analysis [66]. In addition, due to technical limitations, it is not possible to obtain fully purified exosomes, which also affects the accuracy of EV analysis. For example, EVs in the typical exosome size range are known to include exosomes (20—100 nm), microvesicles (100–1000 nm) and apoptotic bodies (1–5 μm), VLDL, chylomicrons, and retroviruses cannot be effectively separated from exosomes completely by centrifugation because of their similar membrane orientation and density [99]. Furthermore, Exosome size may vary considerably even within the same single cell line, perhaps due to the inhomogeneous invagination inherent in the restrictive membrane during exosome biogenesis [66]. Interestingly, exosome size has been reported to be related with certain diseases. For example, in patients with NSCLC, tumor-draining pulmonary vein blood secrete body size (< 112 nm) is associated with a shorter time to recurrence and shorter overall survival [100].
The exosome population is made up of exosomes of different internal and external carriers, different sizes, different cellular origins, different functional effects on the recipient cell, and resulting in uneven application functions [66]. The content of exosomes reflects the state of secretory cells to some extent, making them potentially useful for assessing normal and pathophysiological status [3, 101].
Recent studies have shown that exosomes can be used as biomarkers to diagnose, identify the stage and the subtype of tumor cells. In 2015, Dr. Raghu Kallur’s team found that GPC1 protein contained in exosomes of pancreatic cancer cells could be used as a non-invasive method to diagnose and screen early pancreatic cancer at a stage suitable for surgical treatment. Most importantly, it can distinguish chronic pancreatitis from early or advanced pancreatic cancer, providing a new approach to the diagnosis of early pancreatic cancer, which is difficult to detect clinically [102]. Moreover, through the bone marrow, PCa cells create a pre-metastatic niche through primary PCa TDEs mediated PKM2 transfer to BMSCs and subsequent CXCL12 upregulation. This novel mechanism suggests that exosomal PKM2 may serve as a therapeutic biomarker target for PCa bone metastases [88]. Furthermore, Roberg-Larsen et al. showed the TDEs from MCF-7 cell line (estrogen receptor (ER +) breast cancer cell line) had increased levels of 27-OHC in compared with TDEs from the ER- breast cancer cell line (MDA-MB-231), providing evidence that TDEs may contain additional information of diagnostic value [103].
Exosomes are stable sources of miRNA in body fluids, which prevent the degradation of biomacromolecules under fluctuating body conditions [104]. These highly stabile miRNAs in exosomes are attractive non-invasive biomarker targets for cancer screening and disease surveillance. Analysis of exosome miRNAs in the sera of healthy individuals and cancer patients revealed important differences related to tumor progression, while highlighting the potential value of these miRNAs as biomarkers of disease prognosis [105]. miRNA detected in serum TDEs from breast cancer patients can be used to discriminate between specific molecular subtypes. Furthermore, it has been found that high level of miR-373 in TDEs from breast cancer correlates with triple-negative or other highly aggressive breast cancer types, highlighting the potential role of serum-specific exosomal miR-373 as an aggressive tumor biomarker [106]. The identification of in vitro miRNAs that associate with tumor metastases could provide an additional diagnostic tool to assess disease stage and monitor its progression. High levels of miR-105 have been found in serum-derived exosomes from breast cancer patients who later develop metastatic disease [107]. Similarly, down-regulation of miR-19a and/or miR-29c and up-regulation of miR-210 have been detected in TDEs from brain metastatic breast cancer cells [108]. The overexpression of miR-483-3p occurs in the early development of PDAC and exists in precancerous PanIN lesions, providing evidence that miR-483-3p might be a biomarker for early diagnosis and prognosis of PDAC [109].
Treatment
One interesting therapeutic idea involves harnessing the functionality of exosomes, which transport proteins, lipids, and nucleic acids to mediate cell-to-cell communication between TMEs components, for anticancer therapy. Therefore, research focused on targeting exosome biogenesis and loading is required before the exosome could be used as a viable strategy to treat cancers.
Recent studies have investigated if manipulating the release of TDEs could be used for therapeutics. Inhibition of Rab27 with targeted shRNAs has been shown to reduce exosome release, but this manipulation also leads to a significant increase in smaller endosome-sized vesicles (50 nm), implying that Rab proteins have the ability to alter the size distribution of exosomes [110]. Others have shown that GW4869 (exosome-release inhibitor) can block the secretion of exosomes and reverse metabolic changes in breast cancer cells. GW4869 inhibits glycolysis and receptor cell activation in tumor cells, thereby inhibiting cancer progression [111, 112]. Fanny et al. found that reducing exosomes production with dimethyl amiloride enhanced the antitumor efficacy of the chemotherapy drug cyclophosphamide in vivo in three different mouse tumor models [113]. Another possible mechanism to inhibit the tumor-promoting function of TDEs is to prevent exosome fusion or uptake by target cells. One study suggested that TDEs uptake by cells could be prevented by targeting specific exosome biomarkers [114]. For example, a recent study showed that positively charged mesoporous silica nanoparticles (MSNs) with EGFR-targeting aptamers (MSN-AP) could interact and eliminate circulating cancer-derived negatively charged exosomes by allowing them to enter the small intestine, thereby reducing metastasis formation [115].
Exosomes have also been used to build cancer vaccines. For example, Huang et al. loaded Hiltonol (TLR3 agonist) and the immunogenic cell death inducers human neutrophil elastase into α-LA (α-lactalbumin)-engineered breast cancer-derived exosomes to form an in situ DC vaccine (HELA-Exos). HELA-Exos has been shown to exhibit potent antitumor activity in both mouse models and human breast cancer organoids, which improves subsequent tumor-reactive CD8 + T-cell response by promoting the activation of type one conventional dendritic cells in situ [116]. Taking advantage of the property that TDEs can specifically deliver drugs to the tumor site, Zhu et al. found that TDEs could be developed for combined delivery of AIEgen and proton pump inhibitors (PPI) for the combined treatment of gastric cancer [117]. In addition, a recent study found successful evaluation of drug efficacy by using exosome-synthesized probes to detect drug occupancy [118]. While the early studies of exosome cancer vaccines are promising, more researches are needed to determine the feasibility of antitumor exosome for use in human and mass production. In addition to using cell exosomes, artificial engineering such as freeze–thaw cycles, electroporation, ultrasound, reagent transfection, or saponin methods, are also methods for loading drugs or functional cargo into exosomes [119]. Hypoxia might influence the suitability of exosome cargo as a scaffold for fusion of functional molecules and other drugs, thus affecting the efficiency of treatment. In addition, exosomes in TME exhibit specific uptake under hypoxic conditions, which might provide a pathway for specific targeting of malignant tumors [22].
Conclusion and prospects
Tumor-derived exosomes play a key role in the development of tumors. By regulating the glucose and lipid metabolism of tumor cells and other cells in TME, TDEs promotes more suitable soil and materials for tumor growth. From this point of view, many researchers hope to provide new strategies for the diagnosis and treatment of cancer patients by improving the technology of exosomes detection and isolation, inhibiting the secretion of TDEs, or blocking the binding of TDEs with targeted cells. However, there are also problems that need attention, including how to quickly and effectively load drugs into exosomes, how to enhance the stability, and how to improve the specificity and targeting. It is believed that in the near future, with the progress of science and technology and the continuous efforts of researchers, these questions will get addressed, providing new benefits to patients.
Availability of data and materials
Not applicable.
Abbreviations
- EVs:
-
Extracellular vesicles
- TDE:
-
Tumor-derived EVs
- ILVs:
-
Intraluminal vesicles
- MVBs:
-
Multivesicular bodies
- ESCRT:
-
Endosomal sorting complex required for transport
- ARF6:
-
ADP ribosylation factor 6
- PLD2:
-
Phospholipase D2
- TCA:
-
Tricarboxylic acid cycle
- OXPHOS:
-
Oxidative phosphorylation
- TME:
-
Tumor microenvironment
- NSCLC:
-
Non-small cell lung cancer
- MSCs:
-
Mesenchymal stem cells
- PKM2:
-
Pyruvate kinase M2
- SNAP-23:
-
Synaptosome associated protein 23
- PtNPs:
-
Platinum nanoparticles
- CAFs:
-
Cancer associated fibroblasts
- ECM:
-
Extracellular matrix
- Cav-1:
-
Caveolin-1
- MCT4:
-
Monocarboxylate transporter 4
- TAM:
-
Tumor-associated macrophages
- FAO:
-
Fatty acid oxidation
- ECs:
-
Endothelial cells
- PDAC:
-
Pancreatic Ductal Adenocarcinoma
- MDSCs:
-
Marrow derived suppressor cells
- PA:
-
Phosphatidic acid
- PI:
-
Phosphatidyl inositol
- PE:
-
Phosphatidyl ethanolamine
- PC:
-
Phosphatidyl choline
- PS:
-
Phosphatidyl serine
- ABCG1:
-
ATP-binding cassette transporter G1
- CAAs:
-
Cancer-associated adipocytes
- P-HSL:
-
Phospho-hormone-sensitive lipase
- ADMRs:
-
Adrenomedullin receptors
- ASM:
-
Acid sphingomyelinase
- FASN:
-
Fatty Acid Synthase
- LUAD:
-
Lung adenocarcinoma
- PCa:
-
Prostate cancer
- BMSCs:
-
Bone marrow stromal cells
- NEAT1:
-
Nuclear-enriched abundant transcript 1
- ctDNA:
-
Circulating tumor DNA
- ITH:
-
Intratumoral heterogeneity
- NTA:
-
Nanoparticle Tracking Analysis
- MSN-AP:
-
Mesoporous silica nanoparticles with EGFR-targeting aptamers
References
Hoshino A, Costa-Silva B, Shen TL, et al. Tumour exosome integrins determine organotropic metastasis. Nature. 2015;527(7578):329–35. https://doi.org/10.1038/nature15756.
Wang W, Zhu N, Yan T, et al. The crosstalk: exosomes and lipid metabolism. Cell Commun Signal. 2020;18(1):119. https://doi.org/10.1186/s12964-020-00581-2.
Noren Hooten N, Byappanahalli AM, Vannoy M, Omoniyi V, Evans MK. Influences of age, race, and sex on extracellular vesicle characteristics. Theranostics. 2022;12(9):4459–76. https://doi.org/10.7150/thno.72676.
Ma P, Pan Y, Li W, et al. Extracellular vesicles-mediated noncoding RNAs transfer in cancer. J Hematol Oncol. 2017;10(1):57. https://doi.org/10.1186/s13045-017-0426-y.
Thery C, Zitvogel L, Amigorena S. Exosomes: composition, biogenesis and function. Nat Rev Immunol. 2002;2(8):569–79. https://doi.org/10.1038/nri855.
Colombo M, Raposo G, Thery C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30:255–89. https://doi.org/10.1146/annurev-cellbio-101512-122326.
Milane L, Singh A, Mattheolabakis G, Suresh M, Amiji MM. Exosome mediated communication within the tumor microenvironment. J Control Release. 2015;219:278–94. https://doi.org/10.1016/j.jconrel.2015.06.029.
Hurley JH. The ESCRT complexes. Crit Rev Biochem Mol Biol. 2010;45(6):463–87. https://doi.org/10.3109/10409238.2010.502516.
Katzmann DJ, Babst M, Emr SD. Ubiquitin-dependent sorting into the multivesicular body pathway requires the function of a conserved endosomal protein sorting complex. ESCRT-I Cell. 2001;106(2):145–55. https://doi.org/10.1016/s0092-8674(01)00434-2.
Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 2008;319(5867):1244–7. https://doi.org/10.1126/science.1153124.
Ghossoub R, Lembo F, Rubio A, et al. Syntenin-ALIX exosome biogenesis and budding into multivesicular bodies are controlled by ARF6 and PLD2. Nat Commun. 2014;5:3477. https://doi.org/10.1038/ncomms4477.
van Niel G, Charrin S, Simoes S, et al. The tetraspanin CD63 regulates ESCRT-independent and -dependent endosomal sorting during melanogenesis. Dev Cell. 2011;21(4):708–21. https://doi.org/10.1016/j.devcel.2011.08.019.
Barros FM, Carneiro F, Machado JC, Melo SA. Exosomes and Immune response in Cancer: friends or foes? Front Immunol. 2018;9:730. https://doi.org/10.3389/fimmu.2018.00730.
Rana S, Yue S, Stadel D, Zoller M. Toward tailored exosomes: the exosomal tetraspanin web contributes to target cell selection. Int J Biochem Cell Biol. 2012;44(9):1574–84. https://doi.org/10.1016/j.biocel.2012.06.018.
Nazarenko I, Rana S, Baumann A, et al. Cell surface tetraspanin Tspan8 contributes to molecular pathways of exosome-induced endothelial cell activation. Cancer Res. 2010;70(4):1668–78. https://doi.org/10.1158/0008-5472.CAN-09-2470.
Zhang L, Yu D. Exosomes in cancer development, metastasis, and immunity. Biochim Biophys Acta Rev Cancer. 2019;1871(2):455–68. https://doi.org/10.1016/j.bbcan.2019.04.004.
Milman N, Ginini L, Gil Z. Exosomes and their role in tumorigenesis and anticancer drug resistance. Drug Resist Updat. 2019;45:1–12. https://doi.org/10.1016/j.drup.2019.07.003.
Warburg O. On the origin of cancer cells. Science. 1956;123(3191):309–14. https://doi.org/10.1126/science.123.3191.309.
Currie E, Schulze A, Zechner R, Walther TC, Farese RV Jr. Cellular fatty acid metabolism and cancer. Cell Metab. 2013;18(2):153–61. https://doi.org/10.1016/j.cmet.2013.05.017.
Hensley CT, Wasti AT, DeBerardinis RJ. Glutamine and cancer: cell biology, physiology, and clinical opportunities. J Clin Invest. 2013;123(9):3678–84. https://doi.org/10.1172/JCI69600.
Rodriguez M, Silva J, Lopez-Alfonso A, et al. Different exosome cargo from plasma/bronchoalveolar lavage in non-small-cell lung cancer. Genes Chromosomes Cancer. 2014;53(9):713–24. https://doi.org/10.1002/gcc.22181.
He G, Peng X, Wei S, et al. Exosomes in the hypoxic TME: from release, uptake and biofunctions to clinical applications. Mol Cancer. 2022;21(1):19. https://doi.org/10.1186/s12943-021-01440-5.
Kalluri R. The biology and function of exosomes in cancer. J Clin Invest. 2016;126(4):1208–15. https://doi.org/10.1172/JCI81135.
Tan S, **a L, Yi P, et al. Exosomal miRNAs in tumor microenvironment. J Exp Clin Cancer Res. 2020;39(1):67. https://doi.org/10.1186/s13046-020-01570-6.
Carnino JM, Ni K, ** Y. Post-translational modification regulates formation and cargo-loading of extracellular vesicles. Front Immunol. 2020;11:948. https://doi.org/10.3389/fimmu.2020.00948.
Phoonsawat W, Aoki-Yoshida A, Tsuruta T, Sonoyama K. Adiponectin is partially associated with exosomes in mouse serum. Biochem Biophys Res Commun. 2014;448(3):261–6. https://doi.org/10.1016/j.bbrc.2014.04.114.
Lydic TA, Townsend S, Adda CG, Collins C, Mathivanan S, Reid GE. Rapid and comprehensive ‘shotgun’ lipidome profiling of colorectal cancer cell derived exosomes. Methods. 2015;87:83–95. https://doi.org/10.1016/j.ymeth.2015.04.014.
Llorente A, Skotland T, Sylvanne T, et al. Molecular lipidomics of exosomes released by PC-3 prostate cancer cells. Biochim Biophys Acta. 2013;1831(7):1302–9. https://doi.org/10.1016/j.bbalip.2013.04.011.
Kim DH, Kim H, Choi YJ, et al. Exosomal PD-L1 promotes tumor growth through immune escape in non-small cell lung cancer. Exp Mol Med. 2019;51(8):1–13. https://doi.org/10.1038/s12276-019-0295-2.
Rahman MA, Barger JF, Lovat F, Gao M, Otterson GA, Nana-Sinkam P. Lung cancer exosomes as drivers of epithelial mesenchymal transition. Oncotarget. 2016;7(34):54852–66. https://doi.org/10.18632/oncotarget.10243.
Li X, Wang S, Zhu R, Li H, Han Q, Zhao RC. Lung tumor exosomes induce a pro-inflammatory phenotype in mesenchymal stem cells via NFkappaB-TLR signaling pathway. J Hematol Oncol. 2016;9:42. https://doi.org/10.1186/s13045-016-0269-y.
Gurunathan S, Kang MH, Kim JH. A comprehensive review on factors influences biogenesis, functions, therapeutic and clinical implications of exosomes. Int J Nanomedicine. 2021;16:1281–312. https://doi.org/10.2147/IJN.S291956.
Villarroya-Beltri C, Baixauli F, Gutierrez-Vazquez C, Sanchez-Madrid F, Mittelbrunn M. Sorting it out: regulation of exosome loading. Semin Cancer Biol. 2014;28:3–13. https://doi.org/10.1016/j.semcancer.2014.04.009.
Gong C, Zhang X, Shi M, et al. Tumor exosomes reprogrammed by low pH are efficient targeting vehicles for smart drug delivery and personalized therapy against their homologous tumor. Adv Sci (Weinh). 2021;8(10):2002787. https://doi.org/10.1002/advs.202002787.
Ban JJ, Lee M, Im W, Kim M. Low pH increases the yield of exosome isolation. Biochem Biophys Res Commun. 2015;461(1):76–9. https://doi.org/10.1016/j.bbrc.2015.03.172.
Luo W, Hu H, Chang R, et al. Pyruvate kinase M2 is a PHD3-stimulated coactivator for hypoxia-inducible factor 1. Cell. 2011;145(5):732–44. https://doi.org/10.1016/j.cell.2011.03.054.
Wei Y, Wang D, ** F, et al. Pyruvate kinase type M2 promotes tumour cell exosome release via phosphorylating synaptosome-associated protein 23. Nat Commun. 2017;8:14041. https://doi.org/10.1038/ncomms14041.
Gurunathan S, Kang MH, Jeyaraj M, Kim JH. Platinum nanoparticles enhance exosome release in human lung epithelial adenocarcinoma cancer cells (A549): oxidative stress and the ceramide pathway are key players. Int J Nanomedicine. 2021;16:515–38. https://doi.org/10.2147/IJN.S291138.
Shu S, Yang Y, Allen CL, et al. Metabolic reprogramming of stromal fibroblasts by melanoma exosome microRNA favours a pre-metastatic microenvironment. Sci Rep. 2018;8(1):12905. https://doi.org/10.1038/s41598-018-31323-7.
Cascone T, Mckenzie JA, Mbofung RM, et al. Increased tumor glycolysis characterizes immune resistance to adoptive t cell therapy. Cell Metab. 2018;27(5):977-987 e4. https://doi.org/10.1016/j.cmet.2018.02.024.
Li F, Zhan L, Dong Q, et al. Tumor-derived exosome-educated hepatic stellate cells regulate lactate metabolism of hypoxic colorectal tumor cells via the IL-6/STAT3 pathway to confer drug resistance. Onco Targets Ther. 2020;13:7851–64. https://doi.org/10.2147/OTT.S253485.
Wang D, Zhao C, Xu F, et al. Cisplatin-resistant NSCLC cells induced by hypoxia transmit resistance to sensitive cells through exosomal PKM2. Theranostics. 2021;11(6):2860–75. https://doi.org/10.7150/thno.51797.
Belli C, Trapani D, Viale G, et al. Targeting the microenvironment in solid tumors. Cancer Treat Rev. 2018;65:22–32. https://doi.org/10.1016/j.ctrv.2018.02.004.
Martinez-Outschoorn UE, Pavlides S, Howell A, et al. Stromal-epithelial metabolic coupling in cancer: integrating autophagy and metabolism in the tumor microenvironment. Int J Biochem Cell Biol. 2011;43(7):1045–51. https://doi.org/10.1016/j.biocel.2011.01.023.
Rai A, Greening DW, Chen M, Xu R, Ji H, Simpson RJ. Exosomes derived from human primary and metastatic colorectal cancer cells contribute to functional heterogeneity of activated fibroblasts by reprogramming their proteome. Proteomics. 2019;19(8):e1800148. https://doi.org/10.1002/pmic.201800148.
Sotgia F, Martinez-Outschoorn UE, Howell A, Pestell RG, Pavlides S, Lisanti MP. Caveolin-1 and cancer metabolism in the tumor microenvironment: markers, models, and mechanisms. Annu Rev Pathol. 2012;7:423–67. https://doi.org/10.1146/annurev-pathol-011811-120856.
Zhuang G, Wu X, Jiang Z, et al. Tumour-secreted miR-9 promotes endothelial cell migration and angiogenesis by activating the JAK-STAT pathway. EMBO J. 2012;31(17):3513–23. https://doi.org/10.1038/emboj.2012.183.
Yan W, Wu X, Zhou W, et al. Cancer-cell-secreted exosomal miR-105 promotes tumour growth through the MYC-dependent metabolic reprogramming of stromal cells. Nat Cell Biol. 2018;20(5):597–609. https://doi.org/10.1038/s41556-018-0083-6.
Hong CS, Graham NA, Gu W, et al. MCT1 Modulates Cancer cell pyruvate export and growth of tumors that co-express MCT1 and MCT4. Cell Rep. 2016;14(7):1590–601. https://doi.org/10.1016/j.celrep.2016.01.057.
Pertega-Gomes N, Felisbino S, Massie CE, et al. A glycolytic phenotype is associated with prostate cancer progression and aggressiveness: a role for monocarboxylate transporters as metabolic targets for therapy. J Pathol. 2015;236(4):517–30. https://doi.org/10.1002/path.4547.
Thakur A, Qiu G, Xu C, et al. Label-free sensing of exosomal MCT1 and CD147 for tracking metabolic reprogramming and malignant progression in glioma. Sci Adv. 2020;6(26):eaaz6119. https://doi.org/10.1126/sciadv.aaz6119.
Zhao H, Yang L, Baddour J, et al. Tumor microenvironment derived exosomes pleiotropically modulate cancer cell metabolism. Elife. 2016;5:e10250. https://doi.org/10.7554/eLife.10250.
Achreja A, Zhao H, Yang L, Yun TH, Marini J, Nagrath D. Exo-MFA - A 13C metabolic flux analysis framework to dissect tumor microenvironment-secreted exosome contributions towards cancer cell metabolism. Metab Eng. 2017;43(Pt B):156–72. https://doi.org/10.1016/j.ymben.2017.01.001.
Rabold K, Netea MG, Adema GJ, Netea-Maier RT. Cellular metabolism of tumor-associated macrophages - functional impact and consequences. FEBS Lett. 2017;591(19):3022–41. https://doi.org/10.1002/1873-3468.12771.
Zhu L, Zhao Q, Yang T, Ding W, Zhao Y. Cellular metabolism and macrophage functional polarization. Int Rev Immunol. 2015;34(1):82–100. https://doi.org/10.3109/08830185.2014.969421.
Park JE, Dutta B, Tse SW, et al. Hypoxia-induced tumor exosomes promote M2-like macrophage polarization of infiltrating myeloid cells and microRNA-mediated metabolic shift. Oncogene. 2019;38(26):5158–73. https://doi.org/10.1038/s41388-019-0782-x.
Netea-Maier RT, Smit JWA, Netea MG. Metabolic changes in tumor cells and tumor-associated macrophages: a mutual relationship. Cancer Lett. 2018;413:102–9. https://doi.org/10.1016/j.canlet.2017.10.037.
Fong MY, Zhou W, Liu L, et al. Breast-cancer-secreted miR-122 reprograms glucose metabolism in premetastatic niche to promote metastasis. Nat Cell Biol. 2015;17(2):183–94. https://doi.org/10.1038/ncb3094.
Peinado H, Lavotshkin S, Lyden D. The secreted factors responsible for pre-metastatic niche formation: old sayings and new thoughts. Semin Cancer Biol. 2011;21(2):139–46. https://doi.org/10.1016/j.semcancer.2011.01.002.
Wang B, Wang X, Hou D, et al. Exosomes derived from acute myeloid leukemia cells promote chemoresistance by enhancing glycolysis-mediated vascular remodeling. J Cell Physiol. 2019;234(7):10602–14. https://doi.org/10.1002/jcp.27735.
Rohlenova K, Veys K, Miranda-Santos I, De Bock K, Carmeliet P. Endothelial cell metabolism in health and disease. Trends Cell Biol. 2018;28(3):224–36. https://doi.org/10.1016/j.tcb.2017.10.010.
Basso D, Gnatta E, Padoan A, et al. PDAC-derived exosomes enrich the microenvironment in MDSCs in a SMAD4-dependent manner through a new calcium related axis. Oncotarget. 2017;8(49):84928–44. https://doi.org/10.18632/oncotarget.20863.
Husain Z, Huang Y, Seth P, Sukhatme VP. Tumor-derived lactate modifies antitumor immune response: effect on myeloid-derived suppressor cells and NK cells. J Immunol. 2013;191(3):1486–95. https://doi.org/10.4049/jimmunol.1202702.
Luo X, Zhao X, Cheng C, Li N, Liu Y, Cao Y. The implications of signaling lipids in cancer metastasis. Exp Mol Med. 2018;50(9):1–10. https://doi.org/10.1038/s12276-018-0150-x.
Liu Z, Liu X, Liu S, Cao Q. Cholesterol promotes the migration and invasion of renal carcinoma cells by regulating the KLF5/miR-27a/FBXW7 pathway. Biochem Biophys Res Commun. 2018;502(1):69–75. https://doi.org/10.1016/j.bbrc.2018.05.122.
Pegtel DM, Gould SJ. Exosomes. Annu Rev Biochem. 2019;88:487–514. https://doi.org/10.1146/annurev-biochem-013118-111902.
Zakharova L, Svetlova M, Fomina AF. T cell exosomes induce cholesterol accumulation in human monocytes via phosphatidylserine receptor. J Cell Physiol. 2007;212(1):174–81. https://doi.org/10.1002/jcp.21013.
Namba Y, Sogawa C, Okusha Y, et al. Depletion of lipid efflux pump abcg1 triggers the intracellular accumulation of extracellular vesicles and reduces aggregation and tumorigenesis of metastatic Cancer cells. Front Oncol. 2018;8:376. https://doi.org/10.3389/fonc.2018.00376.
Tafelmeier M, Fischer A, Orso E, et al. Mildly oxidized HDL decrease agonist-induced platelet aggregation and release of pro-coagulant platelet extracellular vesicles. J Steroid Biochem Mol Biol. 2017;169:176–88. https://doi.org/10.1016/j.jsbmb.2016.05.003.
**ang X, Poliakov A, Liu C, et al. Induction of myeloid-derived suppressor cells by tumor exosomes. Int J Cancer. 2009;124(11):2621–33. https://doi.org/10.1002/ijc.24249.
Wu B, Cui J, Yang XM, et al. Cytoplasmic fragment of CD147 generated by regulated intramembrane proteolysis contributes to HCC by promoting autophagy. Cell Death Dis. 2017;8(7):e2925. https://doi.org/10.1038/cddis.2017.251.
Hosseini-Beheshti E, Choi W, Weiswald LB, et al. Exosomes confer pro-survival signals to alter the phenotype of prostate cells in their surrounding environment. Oncotarget. 2016;7(12):14639–58. https://doi.org/10.18632/oncotarget.7052.
Cao Y. Adipocyte and lipid metabolism in cancer drug resistance. J Clin Invest. 2019;129(8):3006–17. https://doi.org/10.1172/JCI127201.
Argiles JM, Busquets S, Stemmler B, Lopez-Soriano FJ. Cancer cachexia: understanding the molecular basis. Nat Rev Cancer. 2014;14(11):754–62. https://doi.org/10.1038/nrc3829.
Hu W, Ru Z, **ao W, et al. Adipose tissue browning in cancer-associated cachexia can be attenuated by inhibition of exosome generation. Biochem Biophys Res Commun. 2018;506(1):122–9. https://doi.org/10.1016/j.bbrc.2018.09.139.
Sagar G, Sah RP, Javeed N, et al. Pathogenesis of pancreatic cancer exosome-induced lipolysis in adipose tissue. Gut. 2016;65(7):1165–74. https://doi.org/10.1136/gutjnl-2014-308350.
Wang S, Li X, Xu M, Wang J, Zhao RC. Reduced adipogenesis after lung tumor exosomes priming in human mesenchymal stem cells via TGFbeta signaling pathway. Mol Cell Biochem. 2017;435(1–2):59–66. https://doi.org/10.1007/s11010-017-3056-3.
Dostert G, Mesure B, Menu P, Velot E. How do mesenchymal stem cells influence or are influenced by microenvironment through extracellular vesicles communication? Front Cell Dev Biol. 2017;5:6. https://doi.org/10.3389/fcell.2017.00006.
Lopatina T, Gai C, Deregibus MC, Kholia S, Camussi G. Cross talk between cancer and mesenchymal stem cells through extracellular vesicles carrying nucleic acids. Front Oncol. 2016;6:125. https://doi.org/10.3389/fonc.2016.00125.
Lee JK, Park SR, Jung BK, et al. Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells. PLoS One. 2013;8(12):e84256. https://doi.org/10.1371/journal.pone.0084256.
Shi S, Zhang Q, **a Y, et al. Mesenchymal stem cell-derived exosomes facilitate nasopharyngeal carcinoma progression. Am J Cancer Res. 2016;6(2):459–72.
Faict S, Oudaert I, D’Auria L, et al. The transfer of sphingomyelinase contributes to drug resistance in multiple Myeloma. Cancers (Basel). 2019;11(12):1823. https://doi.org/10.3390/cancers11121823.
Jiang L, Fang X, Wang H, Li D, Wang X. Ovarian Cancer-Intrinsic fatty acid synthase prevents anti-tumor immunity by disrupting tumor-infiltrating dendritic cells. Front Immunol. 2018;9:2927. https://doi.org/10.3389/fimmu.2018.02927.
Yang W, Bai Y, **ong Y, et al. Potentiating the antitumour response of CD8(+) T cells by modulating cholesterol metabolism. Nature. 2016;531(7596):651–5. https://doi.org/10.1038/nature17412.
Yeon JH, Jeong HE, Seo H, et al. Cancer-derived exosomes trigger endothelial to mesenchymal transition followed by the induction of cancer-associated fibroblasts. Acta Biomater. 2018;76:146–53. https://doi.org/10.1016/j.actbio.2018.07.001.
Liu T, Han C, Fang P, et al. Cancer-associated fibroblast-specific lncRNA LINC01614 enhances glutamine uptake in lung adenocarcinoma. J Hematol Oncol. 2022;15(1):141. https://doi.org/10.1186/s13045-022-01359-4.
Hosios AM, Hecht VC, Danai LV, et al. Amino acids rather than glucose account for the majority of cell mass in proliferating mammalian cells. Dev Cell. 2016;36(5):540–9. https://doi.org/10.1016/j.devcel.2016.02.012.
Dai J, Escara-Wilke J, Keller JM, et al. Primary prostate cancer educates bone stroma through exosomal pyruvate kinase M2 to promote bone metastasis. J Exp Med. 2019;216(12):2883–99. https://doi.org/10.1084/jem.20190158.
Mo C, Huang B, Zhuang J, Jiang S, Guo S, Mao X. Clin Transl Med. 2021;11(8):e493. https://doi.org/10.1002/ctm2.493.
Morrissey SM, Zhang F, Ding C, et al. Tumor-derived exosomes drive immunosuppressive macrophages in a pre-metastatic niche through glycolytic dominant metabolic reprogramming. Cell Metab. 2021;33(10):2040-2058 e10. https://doi.org/10.1016/j.cmet.2021.09.002.
Wang M, Qin Z, Wan J, et al. Tumor-derived exosomes drive pre-metastatic niche formation in lung via modulating CCL1(+) fibroblast and CCR8(+) Treg cell interactions. Cancer Immunol Immunother. 2022;71(11):2717–30. https://doi.org/10.1007/s00262-022-03196-3.
Yin X, Zeng W, Wu B, et al. PPARalpha inhibition overcomes tumor-derived Exosomal lipid-induced dendritic cell dysfunction. Cell Rep. 2020;33(3):108278. https://doi.org/10.1016/j.celrep.2020.108278.
Dang XTT, Kavishka JM, Zhang DX, Pirisinu M, Le MTN. Extracellular vesicles as an efficient and versatile system for drug delivery. Cells. 2020;9(10):2191. https://doi.org/10.3390/cells9102191.
Pantel K, Alix-Panabieres C. Liquid biopsy and minimal residual disease - latest advances and implications for cure. Nat Rev Clin Oncol. 2019;16(7):409–24. https://doi.org/10.1038/s41571-019-0187-3.
Nakamura Y, Taniguchi H, Ikeda M, et al. Clinical utility of circulating tumor DNA sequencing in advanced gastrointestinal cancer: SCRUM-Japan GI-SCREEN and GOZILA studies. Nat Med. 2020;26(12):1859–64. https://doi.org/10.1038/s41591-020-1063-5.
Sun N, Lee YT, Zhang RY, et al. Purification of HCC-specific extracellular vesicles on nanosubstrates for early HCC detection by digital scoring. Nat Commun. 2020;11(1):4489. https://doi.org/10.1038/s41467-020-18311-0.
Vitale I, Shema E, Loi S, Galluzzi L. Intratumoral heterogeneity in cancer progression and response to immunotherapy. Nat Med. 2021;27(2):212–24. https://doi.org/10.1038/s41591-021-01233-9.
Mathieu M, Nevo N, Jouve M, et al. Specificities of exosome versus small ectosome secretion revealed by live intracellular tracking of CD63 and CD9. Nat Commun. 2021;12(1):4389. https://doi.org/10.1038/s41467-021-24384-2.
Zhang H, Freitas D, Kim HS, et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat Cell Biol. 2018;20(3):332–43. https://doi.org/10.1038/s41556-018-0040-4.
Navarro A, Molins L, Marrades RM, et al. Exosome analysis in tumor-draining pulmonary vein identifies NSCLC patients with higher risk of relapse after curative surgery. Cancers (Basel). 2019;11(2):249. https://doi.org/10.3390/cancers11020249.
Kucharzewska P, Christianson HC, Welch JE, et al. Exosomes reflect the hypoxic status of glioma cells and mediate hypoxia-dependent activation of vascular cells during tumor development. Proc Natl Acad Sci U S A. 2013;110(18):7312–7. https://doi.org/10.1073/pnas.1220998110.
Melo SA, Luecke LB, Kahlert C, et al. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature. 2015;523(7559):177–82. https://doi.org/10.1038/nature14581.
Roberg-Larsen H, Lund K, Seterdal KE, et al. Mass spectrometric detection of 27-hydroxycholesterol in breast cancer exosomes. J Steroid Biochem Mol Biol. 2017;169:22–8. https://doi.org/10.1016/j.jsbmb.2016.02.006.
Fabris L, Calin GA. Circulating free xeno-microRNAs - The new kids on the block. Mol Oncol. 2016;10(3):503–8. https://doi.org/10.1016/j.molonc.2016.01.005.
Ye SB, Li ZL, Luo DH, et al. Tumor-derived exosomes promote tumor progression and T-cell dysfunction through the regulation of enriched exosomal microRNAs in human nasopharyngeal carcinoma. Oncotarget. 2014;5(14):5439–52. https://doi.org/10.18632/oncotarget.2118.
Eichelser C, Stuckrath I, Muller V, et al. Increased serum levels of circulating exosomal microRNA-373 in receptor-negative breast cancer patients. Oncotarget. 2014;5(20):9650–63. https://doi.org/10.18632/oncotarget.2520.
Zhou W, Fong MY, Min Y, et al. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell. 2014;25(4):501–15. https://doi.org/10.1016/j.ccr.2014.03.007.
Camacho L, Guerrero P, Marchetti D. MicroRNA and protein profiling of brain metastasis competent cell-derived exosomes. PLoS One. 2013;8(9):e73790. https://doi.org/10.1371/journal.pone.0073790.
Shao H, Zhang Y, Yan J, et al. Upregulated MicroRNA-483-3p is an early event in pancreatic ductal adenocarcinoma (PDAC) and as a powerful liquid biopsy biomarker in PDAC. Onco Targets Ther. 2021;14:2163–75. https://doi.org/10.2147/OTT.S288936.
Zheng Y, Campbell EC, Lucocq J, Riches A, Powis SJ. Monitoring the Rab27 associated exosome pathway using nanoparticle tracking analysis. Exp Cell Res. 2013;319(12):1706–13. https://doi.org/10.1016/j.yexcr.2012.10.006.
Li Y, Zhao Z, Liu W, Li X. SNHG3 Functions as miRNA sponge to promote breast cancer cells growth through the metabolic reprogramming. Appl Biochem Biotechnol. 2020;191(3):1084–99. https://doi.org/10.1007/s12010-020-03244-7.
Wan L, **a T, Du Y, et al. Exosomes from activated hepatic stellate cells contain GLUT1 and PKM2: a role for exosomes in metabolic switch of liver nonparenchymal cells. FASEB J. 2019;33(7):8530–42. https://doi.org/10.1096/fj.201802675R.
Chalmin F, Ladoire S, Mignot G, et al. Membrane-associated Hsp72 from tumor-derived exosomes mediates STAT3-dependent immunosuppressive function of mouse and human myeloid-derived suppressor cells. J Clin Invest. 2010;120(2):457–71. https://doi.org/10.1172/JCI40483.
Lima LG, Chammas R, Monteiro RQ, Moreira ME, Barcinski MA. Tumor-derived microvesicles modulate the establishment of metastatic melanoma in a phosphatidylserine-dependent manner. Cancer Lett. 2009;283(2):168–75. https://doi.org/10.1016/j.canlet.2009.03.041.
**e X, Nie H, Zhou Y, et al. Eliminating blood oncogenic exosomes into the small intestine with aptamer-functionalized nanoparticles. Nat Commun. 2019;10(1):5476. https://doi.org/10.1038/s41467-019-13316-w.
Huang L, Rong Y, Tang X, et al. Engineered exosomes as an in situ DC-primed vaccine to boost antitumor immunity in breast cancer. Mol Cancer. 2022;21(1):45. https://doi.org/10.1186/s12943-022-01515-x.
Zhu D, Zhang T, Li Y, et al. Tumor-derived exosomes co-delivering aggregation-induced emission luminogens and proton pump inhibitors for tumor glutamine starvation therapy and enhanced type-I photodynamic therapy. Biomaterials. 2022;283:121462. https://doi.org/10.1016/j.biomaterials.2022.121462.
Pan S, Zhang Y, Natalia A, et al. Extracellular vesicle drug occupancy enables real-time monitoring of targeted cancer therapy. Nat Nanotechnol. 2021;16(6):734–42. https://doi.org/10.1038/s41565-021-00872-w.
Moller A, Lobb RJ. The evolving translational potential of small extracellular vesicles in cancer. Nat Rev Cancer. 2020;20(12):697–709. https://doi.org/10.1038/s41568-020-00299-w.
Acknowledgements
Not applicable.
Funding
No funders.
Author information
Authors and Affiliations
Contributions
All authors contributed to the conception and the main idea of the work. XF and JS wrote the original draft and prepared Figs. 1 and 2. B.M.D prepared Table 1. JL, WY and WW reviewed and edited the manuscript; project administration. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Fu, X., Song, J., Yan, W. et al. The biological function of tumor-derived extracellular vesicles on metabolism. Cell Commun Signal 21, 150 (2023). https://doi.org/10.1186/s12964-023-01111-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12964-023-01111-6