
Abstract
Background
A significant fraction of the human proteome is still inaccessible to in vitro studies since the recombinant production of several proteins failed in conventional cell factories. Eukaryotic protein kinases are difficult-to-express in heterologous hosts due to folding issues both related to their catalytic and regulatory domains. Human CDKL5 belongs to this category. It is a serine/threonine protein kinase whose mutations are involved in CDKL5 Deficiency Disorder (CDD), a severe neurodevelopmental pathology still lacking a therapeutic intervention. The lack of successful CDKL5 manufacture hampered the exploitation of the otherwise highly promising enzyme replacement therapy. As almost two-thirds of the enzyme sequence is predicted to be intrinsically disordered, the recombinant product is either subjected to a massive proteolytic attack by host-encoded proteases or tends to form aggregates. Therefore, the use of an unconventional expression system can constitute a valid alternative to solve these issues.
Results
Using a multiparametric approach we managed to optimize the transcription of the CDKL5 gene and the synthesis of the recombinant protein in the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125 applying a bicistronic expression strategy, whose generalization for recombinant expression in the cold has been here confirmed with the use of a fluorescent reporter. The recombinant protein largely accumulated as a full-length product in the soluble cell lysate. We also demonstrated for the first time that full-length CDKL5 produced in Antarctic bacteria is catalytically active by using two independent assays, making feasible its recovery in native conditions from bacterial lysates as an active product, a result unmet in other bacteria so far. Finally, the setup of an in cellulo kinase assay allowed us to measure the impact of several CDD missense mutations on the kinase activity, providing new information towards a better understanding of CDD pathophysiology.
Conclusions
Collectively, our data indicate that P. haloplanktis TAC125 can be a valuable platform for both the preparation of soluble active human CDKL5 and the study of structural–functional relationships in wild type and mutant CDKL5 forms. Furthermore, this paper further confirms the more general potentialities of exploitation of Antarctic bacteria to produce “intractable” proteins, especially those containing large intrinsically disordered regions.
Similar content being viewed by others
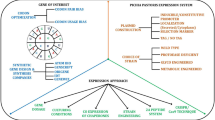


Background
Cyclin-dependent kinase-like 5 (CDKL5) is a serine/threonine protein kinase involved in the development of the human brain. Dozens of mutations of the CDKL5 gene are causative of CDKL5 Deficiency Disorder (CDD; OMIM 300203; 300672) [1, 2], a severe condition that is manifested with intellectual disability, autistic behavior, motor and visual impairments, infantile-onset refractory epilepsy, and many other symptoms [3]. Although no cure for CDD exists today, some studies proved that the restoration of CDKL5 activity through either protein or genetic intervention can revert CDD symptoms in mice and human models [4,49]
Measurement of pGFP fluorescence
After recombinant production, the equivalent of 1 OD cells was harvested and washed with PBS. Then, each sample was diluted with the same buffer to achieve the best signal-to-noise ratio in the fluorescence measurements that were carried out with a JASCO FP-750 spectrofluorometer at 25 °C (excitation at 488 nm and emission at 509 nm).
Analysis of the production of flCDKL5 and EB2 proteins
To analyze total protein productions, 1 OD pellets were solubilized in Laemmli Sample buffer and heated at 90 °C for 20 min. Then, total cellular extracts were resolved by SDS-PAGE and analyzed either by Coomassie staining or Western blot. For the solubility analysis, cells were lysed in 20 mM sodium phosphate buffer pH 7.0, supplemented with 0.5 M NaCl, 10% Glycerol, 0.1% Triton X-100, 20 U/mL DNAse I, 0.1 mg/mL lysozyme, 1 mM DTT and a protease inhibitor cocktail. Then, the soluble and insoluble fractions were segregated by centrifugation. Finally, the insoluble fraction was resuspended with lysis buffer in the same volume as the soluble fraction. 10 µg of soluble extract were analyzed by SDS-PAGE and the same volume of insoluble fraction was used as a control of solubility.
After SDS-PAGE runs, proteins were electroblotted to PVDF membranes using a semidry system. After the incubation with specific antibodies, the chemiluminescent signals were developed with the ECL method.
To detect flCDKL5, the membrane was blocked with PBS, 0.05% Triton X-100, 5% w/v milk for one hour. Then, CDKL5 (D-12): sc-376314 antibody (Santa Cruz Biotechnology) was diluted 1:1000 in the same buffer. After one hour of incubation at room temperature with the primary antibody, the membrane was washed with PBS, 0.05% v/v Triton X-100 three times (5 min each) and incubated with an anti-mouse antibody diluted 1:10,000 in PBS, 0.05% v/v Triton X-100, 5% w/v milk for one hour. Then, the membrane was washed again with PBS, 0.05% v/v Triton X-100 three times and the secondary antibody was detected using the ECL method.
For anti-FLAG Western blots, the membrane was blocked with PBS, 0.2% Tween 20, 5% w/v milk, for one hour. Then, Monoclonal ANTI-FLAG M2, Clone M2 (F1804, Sigma) was diluted 1:1000 in the same buffer. After overnight incubation at 4 °C with the primary antibody, the membrane was washed with PBS, 0.2% Tween 20 three times and incubated with an anti-mouse antibody diluted 1:5000 in PBS, 0.2% Tween 20, 5% w/v milk for one hour at room temperature. Then, the membrane was washed again with PBS, 0.2% Tween 20 three times and the secondary antibody was detected using the ECL method.
In the case of anti-His Western blots, the membrane was blocked with PBS, 5% w/v milk for one hour. Then, Monoclonal Anti-polyHistidine-Peroxidase clone HIS-1 antibody (A7058, Merck) was diluted 1:2000 in PBS, 0.05% Tween 20, 5% w/v milk. After one hour of incubation at room temperature with the antibody, the membrane was washed with PBS, 0.05% Tween 20 three times and it was developed.
To measure EB2 phosphorylation of Ser222, the membrane was blocked with TBST, 5% w/v milk for one hour. Then, anti-EB2 pS222 antibody (00117739, Covalab) was diluted 1:4000 in the same buffer. After overnight incubation at 4 °C with the primary antibody, the membrane was washed with TBST three times and incubated with an anti-rabbit antibody diluted 1:2000 in TBST and 5% w/v milk for one hour at room temperature. Then, the membrane was washed again with TBST three times and was developed.
Parallel Coomassie stained polyacrylamide gels were always used to ascertain that complex samples (i.e. total and soluble lysates) were correctly balanced in Western blot analyses.
Preparation of samples for N-terminal sequencing
To produce the catalytic domain of CDKL5 in E. coli, the primers named PhSumoCDKL5_NdeI_fw and PhCDKL5dC_XhoI_rv were used in a PCR on pB40-BCD2-107 (L). This gene encoding a Sumo-tagged version of CDKL5(1–352) was cloned into the pET40-b vector in frame with a C-terminal 8xHis tag using NdeI/XhoI double digestion. After recombinant expression in E. coli BL21(DE3), the recombinant cells were resuspended in 50 mM TrisHCl pH 8.0, 0.5 M NaCl, 20 mM imidazole and lysed by sonication in the presence of a protease inhibitor cocktail. After centrifugation (14,000 g, 4 °C, 60 min), the soluble fraction was loaded onto a HisTrap of 1 mL (Cytiva) and both the full-length protein, and its N-terminally truncated fragment were collected with a linear gradient of imidazole. A sample containing approximately 30 µg of the intact catalytic domain and 6 µg of the N-terminally truncated fragment were loaded onto SDS-PAGE and then electroblotted onto a PVDF membrane using 10 mM CAPS, 10% methanol pH 11.0 as the transfer buffer. The protein bands were made visible by Ponceau S staining and submitted to Edman sequencing at the Institute of Biosciences and Bioresources (CNR, Naples).
Enrichment of 107 (L) M10V from P. haloplanktis TAC125 lysate
Recombinant P. haloplanktis TAC125 was lysed with a chemical-enzymatic method. Briefly, the cell paste was resuspended in 20 mM sodium phosphate buffer pH 7.0, supplemented with 0.5 M NaCl, 10% glycerol, 0.1% Triton X-100, 20 U/mL DNAse I, 0.1 mg/mL lysozyme, 1 mM DTT, and a protease inhibitor cocktail to reach a final concentration of 14 OD/mL. After incubation at 4 °C for 20 min, the suspension was centrifugated (14,000 g for 45 min at 4 °C) to separate the soluble fraction from the cellular debris. Then, the soluble lysate was incubated with 0.15 mL of ANTI-FLAG M2 Affinity gel (Millipore) at 4 °C for 4 h and a gravity flow column chromatography was performed. The resin was washed with lysis buffer containing 1% Triton X-100, while the elution was performed using in 0.5 mL of the same buffer containing 175 µM 3xFLAG peptide.
EB2 purification
A codon optimized EB2 gene was synthesized by an external company and cloned into the pET40-b with NdeI/BamHI double digestion. The resulting gene encodes human EB2 with an N-terminal 6xHis tag (see the Additional file 1: Information for the nucleotide sequence). After recombinant expression, E. coli BL21(DE3) recombinant cells were lysed by sonication in 50 mM TrisHCl pH 8.0, 0.5 M NaCl, 5% glycerol, 20 mM imidazole supplemented with a protease inhibitor cocktail. The soluble fraction was recovered after a centrifugation (14,000 g for 45 min at 4 °C) and loaded onto a HisTrap of 1 mL (Cytiva). The target protein was eluted with 250 mM imidazole and loaded onto a a Hiload 16/600 Superdex 200 pg using 50 mM TrisHCl pH 8.0, 0.18 M NaCl as a running buffer for a final polishing step. The final preparation was stored at – 80 °C in 40 mM TrisHCl pH 8.0, 0.15 M NaCl, 1 mM DTT, 15% glycerol at 1.8 mg/mL protein concentration.
In vitro kinase assay
EB2 phosphorylation assays with enriched CDKL5 proteins were carried out using 200 nM EB2 and 100 nM of enzyme in 30 μL of 20 mM TrisHCl pH 7.7, 0.5 M NaCl, 10 mM MgCl2, 1 mM DTT, 0.7 mM ATP, complete protease inhibitor cocktail (Roche) and Halt phosphatase inhibitor cocktail (Thermo Fisher Scientific). The reactions were stopped after 30 min with 10 μL Laemmli Sample buffer 4 × and denatured at 70 °C for 20 min. 10 μL of each reaction were analyzed via either SDS-PAGE (for total CDKL5 and EB2 detection) or anti-EB2 pSer222 Western blot (for phosphorylated EB2 detection). As a negative control, a reaction was set up with flCDKL5 KD, a catalytically inactive CDKL5 variant. As a positive control, a reaction with commercial GST-CDKL5(1–498) (ab131695, abcam) was performed.
Statistics and reproducibility of results
The Data from the in cellulo kinase assays were statistically validated using the t-Student test comparing the mean measurements of experimental and control samples, both carried out as technical triplicates. The significance of differences between mean values was calculated using a two-tailed Student’s t-test. A p value of < 0.05 was considered significant.
Availability of data and materials
The sequences of the original flCDKL5 encoding genes (107 (B), 107 (G), 107 (H)) cloned into pUC18 have been deposited in GenBank with the accession codes ON605205, ON605206 and ON605207, respectively. The synthetic plasmid pMK-T-H6EB2 from which the EB2 encoding gene was taken for expression in E. coli is available in GenBank with the accession code ON605208, while the sequence for EB2 co-expression with flCDKL5 was obtained from pMK-RQ-BCD_LacZ-cmyc-EB2-H6, whose accession code in GenBank is ON605209. All the other constructs described in this work were derived from such sequences and the ones described in refs. [30, 31]. Correspondence and material requests should be addressed to MLT (tutino@unina.it) and AC (andrea.colarusso@unina.it).
References
Hector RD, Kalscheuer VM, Hennig F, Leonard H, Downs J, Clarke A, et al. CDKL5 variants. Neurol Genet. 2017;3(6): e200.
Krishnaraj R, Ho G, Christodoulou J. RettBASE: Rett syndrome database update. Hum Mutat. 2017;38(8):922–31.
Olson HE, Demarest ST, Pestana-Knight EM, Swanson LC, Iqbal S, Lal D, et al. Cyclin-dependent kinase-like 5 deficiency disorder: clinical review. Pediatr Neurol. 2019;97:18–25.
Trazzi S, De Franceschi M, Fuchs C, Bastianini S, Viggiano R, Lupori L, et al. CDKL5 protein substitution therapy rescues neurological phenotypes of a mouse model of CDKL5 disorder. Hum Mol Genet. 2018;27(9):1572–92.
Terzic B, Felicia Davatolhagh M, Ho Y, Tang S, Liu YT, **a Z, et al. Temporal manipulation of Cdkl5 reveals essential postdevelopmental functions and reversible CDKL5 deficiency disorder-related deficits. J Clin Invest. 2021. https://doi.org/10.1172/JCI143655.
Gao Y, Irvine EE, Eleftheriadou I, Naranjo CJ, Hearn-Yeates F, Bosch L, et al. Gene replacement ameliorates deficits in mouse and human models of cyclin-dependent kinase-like 5 disorder. Brain. 2020;143(3):811–32.
Kim JY, Bai Y, Jayne LA, Hector RD, Persaud AK, Ong SS, et al. A kinome-wide screen identifies a CDKL5-SOX9 regulatory axis in epithelial cell death and kidney injury. Nat Commun. 2020. https://doi.org/10.1038/s41467-020-15638-6.
Muñoz IM, Morgan ME, Peltier J, Weiland F, Gregorczyk M, Brown FC, et al. Phosphoproteomic screening identifies physiological substrates of the CDKL 5 kinase. EMBO J. 2018;37(24):1–19.
Baltussen LL, Negraes PD, Silvestre M, Claxton S, Moeskops M, Christodoulou E, et al. Chemical genetic identification of CDKL 5 substrates reveals its role in neuronal microtubule dynamics. EMBO J. 2018;37(24):1–18.
Khanam T, Muñoz I, Weiland F, Carroll T, Morgan M, Borsos BN, et al. CDKL5 kinase controls transcription-coupled responses to DNA damage. EMBO J. 2021;40(23): e108271.
Nawaz MS, Giarda E, Bedogni F, La MP, Ricciardi S, Ciceri D, et al. CDKL5 and shootin1 interact and concur in regulating neuronal polarization. PLoS ONE. 2016. https://doi.org/10.1371/journal.pone.0148634.
Barbiero I, Peroni D, Tramarin M, Chandola C, Rusconi L, Landsberger N, et al. The neurosteroid pregnenolone reverts microtubule derangement induced by the loss of a functional CDKL5-IQGAP1 complex. Hum Mol Genet. 2017;26(18):3520–30.
Khokhlatchev A, Xu S, English J, Wu P, Schaefer E, Cobb MH. Reconstitution of mitogen-activated protein kinase phosphorylation cascades in bacteria. Efficient synthesis of active protein kinases. J Biol Chem. 1997;272(17):11057–62.
Fu Z, Schroeder MJ, Shabanowitz J, Kaldis P, Togawa K, Rustgi AK, et al. Activation of a nuclear Cdc2-related kinase within a mitogen-activated protein kinase-like TDY motif by autophosphorylation and cyclin-dependent protein kinase-activating kinase. Mol Cell Biol. 2005;25(14):6047–64.
Fu Z, Larson KA, Chitta RK, Parker SA, Turk BE, Lawrence MW, et al. Identification of Yin-Yang regulators and a phosphorylation consensus for male germ cell-associated kinase (MAK)-related kinase. Mol Cell Biol. 2006;26(22):8639–54.
Hector RD, Dando O, Landsberger N, Kilstrup-Nielsen C, Kind PC, Bailey MES, et al. Characterisation of CDKL5 transcript isoforms in human and mouse. PLoS ONE. 2016;11(6):1–22.
Fahmi M, Yasui G, Seki K, Katayama S, Kaneko-Kawano T, Inazu T, et al. In silico study of Rett Syndrome treatment-related genes, MECP2, CDKL5, and FOXG1, by evolutionary classification and disordered region assessment. Int J Mol Sci. 2019;20(22):1–19.
Altmeyer M, Neelsen KJ, Teloni F, Pozdnyakova I, Pellegrino S, Grøfte M, et al. Liquid demixing of intrinsically disordered proteins is seeded by poly(ADP-ribose). Nat Commun. 2015;6.
Paz A, Zeev-Ben-Mordehai T, Sussman JL, Silman I. Purification of intrinsically disordered proteins [Internet]. Instrumental Analysis of Intrinsically Disordered Proteins. 2010. p. 695–704. (Wiley Online Books).
Canning P, Park K, Gonçalves J, Li C, Howard CJ, Sharpe TD, et al. CDKL family kinases have evolved distinct structural features and ciliary function. Cell Rep. 2018;22(4):885–94.
Katayama S, Sueyoshi N, Kameshita I. Critical determinants of substrate recognition by cyclin-dependent kinase-like 5 (CDKL5). Biochemistry. 2015;54(19):2975–87.
Kameshita I, Sekiguchi M, Hamasaki D, Sugiyama Y, Hatano N, Suetake I, et al. Cyclin-dependent kinase-like 5 binds and phosphorylates DNA methyltransferase 1. Biochem Biophys Res Commun. 2008;377(4):1162–7.
Katayama S, Inazu T. Straightforward and rapid method for detection of cyclin-dependent kinase-like 5 activity. Anal Biochem. 2019;566(November 2018):58–61.
Vigentini I, Merico A, Tutino ML, Compagno C, Marino G. Optimization of recombinant human nerve growth factor production in the psychrophilic Pseudoalteromonas haloplanktis. J Biotechnol. 2006;127(1):141–50.
Papa R, Rippa V, Sannia G, Marino G, Duilio A. An effective cold inducible expression system developed in Pseudoalteromonas haloplanktis TAC125. J Biotechnol. 2007;127(2):199–210.
Unzueta U, Vázquez F, Accardi G, Mendoza R, Toledo-Rubio V, Giuliani M, et al. Strategies for the production of difficult-to-express full-length eukaryotic proteins using microbial cell factories: production of human alpha-galactosidase A. Appl Microbiol Biotechnol. 2015;99(14):5863–74.
Calvanese M, Colarusso A, Lauro C, Parrilli E, Tutino ML. Soluble recombinant protein production in Pseudoalteromonas haloplanktis TAC125: the case study of the full-length human CDKL5 protein. Methods Mol Biol. 2022;2406:219–32.
Fondi M, Maida I, Perrin E, Mellera A, Mocali S, Parrilli E, et al. Genome-scale metabolic reconstruction and constraint-based modelling of the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125. Environ Microbiol. 2015;17(3):751–66.
Fondi M, Gonzi S, Dziurzynski M, Turano P, Ghini V, Calvanese M, et al. Modelling hCDKL5 heterologous expression in bacteria. Metabolites. 2021;11(8):1–16.
Sannino F, Giuliani M, Salvatore U, Apuzzo GA, de Pascale D, Fani R, et al. A novel synthetic medium and expression system for subzero growth and recombinant protein production in Pseudoalteromonas haloplanktis TAC125. Appl Microbiol Biotechnol. 2017;101(2):725–34.
Colarusso A, Lauro C, Calvanese M, Parrilli E, Tutino ML. Improvement of Pseudoalteromonas haloplanktis TAC125 as a cell factory: IPTG-Inducible plasmid construction and strain engineering. Microorganisms. 2020;8(10):1–24.
Mutalik VK, Guimaraes JC, Cambray G, Lam C, Christoffersen MJ, Mai QA, et al. Precise and reliable gene expression via standard transcription and translation initiation elements. Nat Methods. 2013;10(4):354–60.
Tutino ML, Duilio A, Parrilli E, Remaut E, Sannia G, Marino G. A novel replication element from an Antarctic plasmid as a tool for the expression of proteins at low temperature. Extremophiles. 2001;5(4):257–64.
Puigbò P, Guzmán E, Romeu A, Garcia-Vallvé S. OPTIMIZER: a web server for optimizing the codon usage of DNA sequences. Nucleic Acids Res. 2007;35(SUPPL2):126–31.
Qi W, Colarusso A, Olombrada M, Parrilli E, Patrignani A, Tutino ML, et al. New insights on Pseudoalteromonas haloplanktis TAC125 genome organization and benchmarks of genome assembly applications using next and third generation sequencing technologies. Sci Rep. 2019;9(1):16444.
Koutmou KS, Schuller AP, Brunelle JL, Radhakrishnan A, Djuranovic S, Green R. Ribosomes slide on lysine-encoding homopolymeric A stretches. Elife. 2015;2015(4):1–18.
Arthur LL, Pavlovic-djuranovic S, Koutmou KS, Green R. Translational control by lysine-encoding A-rich sequences. Sci Adv. 2015. https://doi.org/10.1126/sciadv.1500154.
Buskirk AR, Green R. Ribosome pausing, arrest and rescue in bacteria and eukaryotes. Philos Trans R Soc B Biol Sci. 2017. https://doi.org/10.1098/rstb.2016.0183.
Suskiewicz MJ, Sussman JL, Silman I, Shaul Y. Context-dependent resistance to proteolysis of intrinsically disordered proteins. Protein Sci. 2011;20(8):1285–97.
Zhang G, Ignatova Z. Folding at the birth of the nascent chain: coordinating translation with co-translational folding. Curr Opin Struct Biol. 2011;21(1):25–31.
Verma M, Choi J, Cottrell KA, Lavagnino Z, Thomas EN, Pavlovic-Djuranovic S, et al. A short translational ramp determines the efficiency of protein synthesis. Nat Commun. 2019;10(1):1–15.
Leith EM, O’Dell WB, Ke N, McClung C, Berkmen M, Bergonzo C, et al. Characterization of the internal translation initiation region in monoclonal antibodies expressed in Escherichia coli. J Biol Chem. 2019;294(48):18046–56.
Zhao Z, Liu X, Zhang W, Yang Y, Dai X, Bai Z. Construction of genetic parts from the Corynebacterium glutamicum genome with high expression activities. Biotechnol Lett. 2016;38(12):2119–26.
Whitaker WR, Lee H, Arkin AP, Dueber JE. Avoidance of truncated proteins from unintended ribosome binding sites within heterologous protein coding sequences. ACS Synth Biol. 2015;4(3):249–57.
Bertani I, Rusconi L, Bolognese F, Forlani G, Conca B, De Monte L, et al. Functional consequences of mutations in CDKL5, an X-linked gene involved in infantile spasms and mental retardation. J Biol Chem. 2006;281(42):32048–56.
Lin C, Franco B, Rosner MR. CDKL5/Stk9 kinase inactivation is associated with neuronal developmental disorders. Hum Mol Genet. 2005;14(24):3775–86.
Parrilli E, Giuliani M, Tutino ML. General secretory pathway from marine antarctic Pseudoalteromonas haloplanktis TAC125. Mar Genomics. 2008;1(3–4):123–8.
Engler C, Kandzia R, Marillonnet S. A one pot, one step, precision cloning method with high throughput capability. PLoS ONE. 2008;3(11): e3647.
Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001;29(9):45.
Acknowledgements
This work is dedicated to Elettra Yvonne, Sofia, Elena, Arianna, Filippo, Raina, Gianluigi and all the beloved CDD patients all over the world, to their extraordinary parents and relatives. We feel responsible for your hope. Open access publication fees was kindly financed by the Department of Chemical Sciences, “Federico II” University of Naples.
Funding
This research was funded by a research grant from the University of Pennsylvania Orphan Disease Center on behalf of the Loulou Foundation to MLT (pilot award no. CDKL5-20-101-08), by the Italian Parents’ Association “La fabbrica dei sogni 2—New developments for Rett syndrome” (to A.C., M.C., C.L., E.P. and M.L.T.), and by the Italian Parents’ Association “CONRETT ONLUS” (to A.C. and C.L.).
Author information
Authors and Affiliations
Contributions
A.C., E.P. and M.L.T. designed the project, A.C. was responsible of its experimental execution, of data collection and analysis, and wrote the manuscript. C.L. developed some bicistronic constructs for the expression of CDKL5 and pGFP, set up the in vitro activity assay and purified flCDKL5. M.C. handled the isolation of B40 origin of replication, the production of some flCDKL5 missense mutated genes, and the setup of the first in cellulo co-expression assay. C.L. and M.C. established the EB2 production and purification protocol. All the authors revised the manuscript and approved the content.
Corresponding author
Ethics declarations
Consent for publication
not applicable.
Competing interests
Authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Characteristics of BCD constructs for pGFP expression. Table S2. List of primers used in this work. Fig. S1. Average plasmid copy number (PCN) of pP79-107 (B). Fig. S2. Ranking of bicistronic designs (BCDs) with a fluorescent reporter. Fig. S3. flCDKL5 production profiles with pBCD-107 (L) plasmids. Fig. S4. Development of Bicistronic Entry Clones. Fig. S5. Development of Bicistronic Designs. Fig. S6. Development of Tricistronic Designs.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Colarusso, A., Lauro, C., Calvanese, M. et al. Active human full-length CDKL5 produced in the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125. Microb Cell Fact 21, 211 (2022). https://doi.org/10.1186/s12934-022-01939-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-022-01939-6