
Abstract
Background
Pancreatic ductal adenocarcinoma (PDAC) is one of the deadliest cancers based on five-year survival rates. Genes contributing to chemoresistance represent novel therapeutic targets that can improve treatment response. Increased expression of ANGPTL4 in tumors correlates with poor outcomes in pancreatic cancer.
Methods
We used statistical analysis of publicly available gene expression data (TCGA-PAAD) to test whether expression of ANGPTL4 and its downstream targets, ITGB4 and APOL1, were correlated with patient survival. We measured the impact of ANGPTL4 overexpression in a common pancreatic cancer cell line, MIA PaCa-2 cells, using CRISPRa for overexpression and DsiRNA for knockdown. We characterized global gene expression changes associated with high levels of ANGPTL4 and response to gemcitabine treatment using RNA-sequencing. Gemcitabine dose response curves were calculated on modified cell lines by measuring cell viability with CellTiter-Glo (Promega). Impacts on cell migration were measured using a time course scratch assay.
Results
We show that ANGPTL4 overexpression leads to in vitro resistance to gemcitabine and reduced survival times in patients. Overexpression of ANGPTL4 induces transcriptional signatures of tumor invasion and metastasis, proliferation and differentiation, and inhibition of apoptosis. Analyses revealed an overlap** signature of genes associated with both ANGPTL4 activation and gemcitabine response. Increased expression of the genes in this signature in patient PDAC tissues was significantly associated with shorter patient survival. We identified 42 genes that were both co-regulated with ANGPTL4 and were responsive to gemcitabine treatment. ITGB4 and APOL1 were among these genes. Knockdown of either of these genes in cell lines overexpressing ANGPTL4 reversed the observed gemcitabine resistance and inhibited cellular migration associated with epithelial to mesenchymal transition (EMT) and ANGPTL4 overexpression.
Conclusions
These data suggest that ANGPTL4 promotes EMT and regulates the genes APOL1 and ITGB4. Importantly, we show that inhibition of both targets reverses chemoresistance and decreases migratory potential. Our findings have revealed a novel pathway regulating tumor response to treatment and suggest relevant therapeutic targets in pancreatic cancer.
Similar content being viewed by others


Background
Pancreatic ductal adenocarcinoma (PDAC) is among the deadliest cancers with a five-year survival of only 11% (SEER) [1]. The high mortality of pancreatic cancer patients is explained by the lack of early diagnostic markers leading to late-stage diagnosis of the disease and the high rate of treatment resistance [2]. Standard of care includes resection for about 20% of patients with localized disease who are eligible and treatment with cytotoxic chemotherapies such as gemcitabine or combination agents like FOLFIRINOX (5-FU, oxaliplatin, folinic acid, and irinotecan). This combination, when tolerated, improves progression-free survival by 11 months [3]. Despite the aggressive treatment of this disease with combinations of cytotoxic drugs, the vast majority of patients do not have an enduring response to the treatment. Through improvements in precision cancer treatments based on genetic mutations, implementation of combination therapies like FOLFIRINOX, and advances in immunotherapy over the last decade the five-year survival has increased from 6 to 11%, nevertheless, pancreatic cancer remains extremely deadly. These factors combined with the increased incidence of risk factors like diabetes and obesity. It is predicted that pancreatic cancer will be the second leading cause of cancer deaths by 2030 [4].
Doubling the five-year survival rate is remarkable progress, but novel treatment strategies are still needed. One effective strategy is to identify cellular markers or pathways that are enriched in pancreatic cancer cells. For example, over 90% of PDAC tumors have a mutation in the KRAS gene, however, no KRAS targeting treatments are available and existing targeted therapies like PARP inhibitors [23]. Heatmaps were generated using iDEP.95 [24].
Scratch-wound assay
Cells were seeded at 100,000 cells per well of a 96-well plate. After 24 h uniform scratches were made across the diameter of the wells using a multichannel pipette with 200ul pipette tips and even pressure applied across the wells to cause a wound. The media was then vacuumed off and 200ul of new media was added. Cells were imaged on a Lionheart FX Automated Microscope every 8 h for 48 h. Cell culture growth conditions of 37 °C and 5% CO2 were maintained throughout. Forty images were taken of each scratch (4 wide by 10 long) with no overlap, autofocus, and auto brightness.
Images were integrated using R and the “tiff” package. Images were further processed using GIMP 2.10. ImageJ and the MRI_wound_Healing_Tool.ijm macro plugin [25] were used to process the images and calculate the area of the wound the cells have not covered. Relative wound closure over time using time 0 for each condition as the control was plotted and a curve line equation was formed by fitting the curve to a non-linear fit one phase decay least squares fit with Yo = 0 as a constraint. This base equation (Y=(Y0 - Plateau)*exp(-K*X) + Plateau) where Y0 is the Y value when X (time) is zero, Plateau is the Y value at infinite times, and K is the rate constant, was used to determine the value of X or half the time it takes to close the wound. Data were analyzed using GraphPad Prism 9.
Drug resistance screening
Cells were seeded in 96-well plates at 2000 cells/well. Seeded cells were treated with a range of gemcitabine concentrations. Cells were treated again 48 h later. The number of viable cells surviving drug treatment were measured with CellTiter-Glo (Promega #G7571) 24 h after the last drug treatment per the manufacturer’s protocol using a BioTek Synergy H5 plate reader. Sample size ranged from 4 to 11.
Pathway analysis
1198 DEG from the ANGPTL4 OE vs. KD analysis were imported into the KEGG Mapper-Color [26] and Panther GO Enrichment Analysis [27]. The following parameters were used for KEGG: Search mode: hsa, used uncolored diagrams, included aliases. The following parameters were used for PANTHER: overrepresentation test, homo sapiens reference list, GO biological process complete, Fisher’s exact test, and calculate FDR. Top pathways were merged manually and drawn using BioRender.
Overall survival analysis (OS)
To conduct survival analysis, clinical and RNA-seq expression data was retrieved from The Cancer Genome Atlas for 178 PDAC (TCGA-PAAD) patients (https://portal.gdc.cancer.gov/). Data were normalized using the R package DESeq2 and differentially expressed genes with an FDR < 0.1 were used to generate Kaplan-Meier survival curves. We classified tissues based on their average expression of a given gene set (bottom 25%, middle 50%, and top 25% of gene expression). We compared the patients with the lowest and highest quartile of average gene expression and performed survival analysis. Survival curves and analyses were generated using the “ggplot2”, “survminer”, and “survival” R packages. P values were generated using a log rank test.
Recurrence free survival (RFS)
A Kaplan Meier curve of recurrence-free survival survival plots for ANGPTL4 was created using GEPIA (http://gepia.cancer-pku.cn/) single gene analysis. The relevant parameters were as follows: Disease-free Survival (RFS), Group Cutoff: Quartile (75% high, 25% low), Hazards Ratio: Yes, 95% Confidence Interval: Yes, Axis Units: Months, and datasets Selection: PAAD.
Correlation analysis
To compute correlation matrices, RNA-seq expression data was retrieved from The Cancer Genome Atlas for 178 PDAC (TCGA-PAAD) patients (https://portal.gdc.cancer.gov/).
Rank-based correlation coefficients were computed for each of the DEG with log2 fold change greater than 0.7 and baseMean > 10 (MP2_ANGPTL4_KD vs. MP2_ANGPTL4_OE with or without treatment) for (1) all TCGA-PAAD data DEG per gene vs. ANGPTL4 expression and (2) all TCGA-PAAD data DEG gene versus OS time. These correlation values were used to generate a list of genes that are co-expressed with ANGPTL4 is overexpressed.
Correlation coefficients and p-values were computed using the “Hmisc” (version 4.6-0) R package. We classified tissues based on their ANGPTL4 expression of a given gene set (bottom 25%, middle 50%, and top 25% of gene expression). We computed correlation matrices using the expression data for patients with the lowest and highest quartile of ANGPTL4 expression which was used to divide samples into quantiles with highest and lowest average gene expression; based on expression of these 42 genes. (40 genes were measured in TCGA: KDM7a and STN1 were omitted).
Statistical analysis
Statistical analysis was conducted in R (version 3.6.1 and R version 4.1.2 for RNAseq analysis). The following R packages and software were used for analysis:
Results
ANGPTL4 overexpression increases gemcitabine resistance genes in PDAC
We previously showed that increased ANGPTL4 expression in tumors is associated with poor survival in PDAC patients (n = 51) [8]. Here, we confirm the association with survival in an independent cohort of 178 PDAC patients from The Cancer Genome Atlas (TCGA, PAAD expression dataset). Patients with the highest (top 25%) expression of ANGPTL4 in tumor tissue had reduced overall survival (OS) and disease-free survival, (p = 0.043 and p = 0.059 (log-rank test) (Fig. 1A, Supplementary Fig. S1A).

Expression of ANGPTL4 is associated with patient outcomes and modifies expression of resistance-associated genes. (a) A Kaplan Meier curve for overall survival using TCGA-PAAD data. The patients with the top (red) and bottom (blue) 25% average ANGPTL4 gene expression have significantly different OS. (p = 0.043, log-rank test) (b) Relative expression of ANGPTL4 transcript in MP2_ANGPTL4_KD (black circles), MP2 (pink squares), and MP2_ANGPTL4_OE (turquoise triangles) as measured by qPCR. P-values from unpaired, two-tailed parametric t-tests performed with 95% confidence intervals are displayed. Values are normalized with ACTB as the housekeeper. (P-values ≤ 0.0001=****) (c) Heatmap displaying RNA-seq results from MP2_ANGPTL4_KD (black), MP2_NTC (pink), and MP2_ANGPTL4_OE (turquoise) cell lines. Displayed are 15 genes associated with both ANGPTL4 overexpression and gemcitabine resistance. Normalized expression for each of the 15 genes is plotted in the heatmap. Red (*) indicates ITGB4 used for later experiments
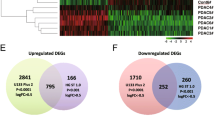
Our previous findings motivated the current study to understand possible mechanisms by which ANGPTL4 contributes to patient survival and specifically how it alters survival outcomes through a possible role in drug resistance. We measured the impact of ANGPTL4 overexpression and knockdown in the MIA PaCa-2 cell line using CRISPRa (MP2_ANGPTL4_OE) [21] and siRNA knockdown (MP2_ANGPTL4_KD). qPCR showed CRISPRa successfully increased expression of the ANGPTL4 transcript by 8-fold compared to a non-targeting control guide. Similarly, siRNA knockdown reduced expression by over 90% of the control line (Fig. 1B, Supplementary Table S1). An ELISA assay showed that protein levels were affected similarly (Supplementary Fig. S1B). We measured global gene expression changes using RNA-sequencing on the modified cell lines and controls which revealed 1198 differentially expressed genes (DEG) with a mean read count greater than 10, absolute log2Fold Change (log2FC) of at least 0.7, and adjusted p-value (padj) less than 0.05) when comparing overexpressed (OE) ANGPTL4 cells with the knockdown (KD) ANGPTL4 cells (Supplementary Fig. S1C, Supplementary Table S2). Given the observed association with drug resistance, we asked whether these DEGs were also associated with gemcitabine resistance by overlap** the 1198 DEG with 114 known gemcitabine resistance genes (Supplementary Table S3) [35,36,37,38]. This revealed 15 gemcitabine resistance genes that are altered with overexpression of ANGPTL4, 10 of which overlap genes that are part of the EMTome (Fig. 1C) [39]. Several of these genes are linked to activation of TGFβ and its downstream signaling pathways: ERK, PI3K/AKT, and JNK (SH3RF2, TGFBR3, PPP1R15A, INPP4B, WWTR1, and FGF2), contributors to EMT (SOX9, CUL7, and TGFBR3), and transcriptional regulation of pluripotent stem cells (STAT3, KLF4, and FGF2) [26, 27]. Additional pathway analysis showed a connection between the expression of ANGPTL4 and EMT (GO:0001837) in the presence (3.24-fold enrichment, 0.0273 FDR) or absence (3.04-fold enrichment, 0.195 FDR) of gemcitabine treatment (Supplementary Table S4-S5).
Increased ANGPTL4 expression alters cellular response to gemcitabine
We assessed whether altering ANGPTL4 expression impacted response to gemcitabine to explore the potential impact of ANGPTL4 overexpression on treatment response in patients. We measured cell viability in MP2_ANGPTL4_KD and MP2_ANGPTL4_OE cell lines before and after treatment with gemcitabine. As expected, treatment with 12.5nM gemcitabine reduces viability compared to untreated cells. Comparing cells with overexpression of ANGPTL4 to controls, we observed an increase in cell viability with the same gemcitabine concentrations. Similarly, there was a significant reduction in viability with the ANGPTL4 knockdown compared to cells overexpressing ANGPTL4. This difference in viability is not observed in the untreated cells (Fig. 2A, Supplementary Table S1). These results demonstrate a greater sensitivity to gemcitabine with reduction of ANGPTL4. To understand the link between ANGPTL4 expression and gemcitabine response, we measured transcriptional response to gemcitabine in MP2_ANGPTL4_OE cells and MP2_ANGPTL4_KD cells using RNA-sequencing. We were not surprised to note that 955 genes were differentially expressed upon treatment with the cytotoxic gemcitabine alone. We explored the intersection of the 1198 DEGs identified upon ANGPTL4 overexpression and the 955 DEGs associated with gemcitabine treatment. This analysis revealed 42 genes associated with gemcitabine response and ANGPTL4 overexpression (Fig. 2B, Supplementary Table S6). This strategy nominated genes that might contribute to altered response to gemcitabine in cells overexpressing ANGPTL4 and are potentially important for patient outcomes. We tested whether these genes were predictive of patient overall survival using data from TCGA. Since all 42 transcripts were positively correlated with ANGPTL4 expression, we calculated the mean gene expression of these genes and found that patients whose tumors had the highest average expression had significantly reduced overall survival (p = 0.0073, Fig. 2C). The fraction of patients surviving 1500 days or more is dramatically increased in the individuals with low expression of these 42 ANGPTL4 and gemcitabine response genes. This compares favorably to the average 3-year relative survival for PDAC patients in the United States with resectable stage I/II disease- 17% [40].

Gemcitabine resistance signature in PDAC cells with high ANGPTL4. (a) Cell viability was measured for MP2_ANGPTL4_KD (black), MP2_NTC (pink), and MP2_ANGPTL4_OE (turquoise) cells treated with 0, 5nM, or 12.5nM gemcitabine. Relative viability reflects normalization to cells not treated with gemcitabine. P-value asterisks: ≤0.01=**, ≤ 0.001=*** & ≤0.0001=****, unpaired, two-tailed t-test (b) Heatmap of gene expression measured by RNA-seq in MP2_ANGPTL4_KD (black/gray), MP2 (pink/light pink), and MP2_ANGPTL4_OE (turquoise/aqua) cell lines both untreated and treated with 1.5nM gemcitabine. Normalized expression of the 42 genes that resulted from an intersection of ANGPTL4 expression DEGs and gemcitabine treatment response DEGs are included in the heatmap. (c) Kaplan-Meier curve for overall survival using TCGA-PAAD data where patients were divided into quartiles with highest (red) and lowest (blue) using mean expression of the 42 genes in the intersection (40 genes were available in TCGA: KDM7a and STN1 were omitted). P-values were derived from a log-rank test
ITGB4 and APOL1 inhibition sensitizes high ANGPTL4 cells to gemcitabine
In vitro overexpression of ANGPTL4 alters cellular drug response and correlates with patient survival but the pathway(s) by which this resistance is achieved are unknown. We reasoned that genes co-regulated with ANGPTL4 may function with ANGPTL4 and contribute to resistance and found those genes by identifying genes that are co-expressed with ANGPTL4 in patient tissues. For each of the 1198 DEGs identified when ANGPTL4 was overexpressed in vitro, we calculated rank-based correlation coefficients and identified co-expressed genes in patient tissues (TCGA-PAAD). For each gene with a significant correlation with ANGPTL4 in patient tissues, we determined whether expression of that gene alone was significantly correlated with patient overall survival (Supplementary Table S7). This allowed further filtering of the list based on the assumption that true resistance-associated genes would impact patient survival. From this list, ITGB4 and APOL1 are both positively correlated with ANGPTL4 expression in vitro and in patients, altered with gemcitabine response in vitro, and correlated with patient survival in tissues. While there are several genes of interest, we focused on ITGB4 and APOL1 in this study, in part based on their drugability (pharos.nih.gov) and availability of efficient siRNA knockdown reagents. Based on the co-expression with ANGPTL4 and the link to patient survival, we tested whether expression of these genes in combination with ANGPTL4 was relevant for patient outcomes. Comparing tissues that were among the top 25% expression of both ANGPTL4 and APOL1 (Fig. 3A) or ITGB4 (Fig. 3B) to those that were among the bottom 25% expression of both ANGPTL4 and APOL1 or ITGB4 we saw significant differences in overall survival, with overall survival being worse for TCGA-PAAD patients with high ANGPTL4 combined with high APOL1 (p = 0.014) or ITGB4 (p = 0.01) expression. Tumors with high levels (top 25%) of all three genes; APOL1, ITGB4, and ANGPTL4, also had reduced survival compared with the bottom 25%, but when APOL1 and ITGB4 are combined with ANGPTL4 expression the reduction in survival did not become more significant (p = 0.015) (Supplementary Fig. S1D).

ITGB4 and APOL1 inhibition sensitizes cells overexpressing ANGPTL4 to gemcitabine. a-b) Kaplan-Meier plot for overall survival using TCGA-PAAD data. Patients with the top (red) and bottom (blue) 25% average ANGPTL4 gene expression and APOL1 or ITGB4 gene expression (p = 0.01 and p = 0.014, respectively, log-rank test). c) ANGPTL4 expression measured by qPCR normalized to MP2_ANGPTL4_OE + DsiRNA_NTC (turquoise) control: MP2 (pink), MP2_ANGPTL4_OE_ITGB4_KD (lavender), and MP2_ANGPTL4_OE_APOL1_KD (purple) compared to mean of MP2_ANGPTL4_OE + DsiRNA_NTC (MP2_ANGPTL4_OE cells transduced with non-targeting control siRNA). Expression is normalized to the housekee** gene ACTB. P-values ≤ 0.0001=****; unpaired two-tailed parametric t-test d) Relative viability is plotted by normalizing all data to cells not treated with gemcitabine. MP2_ANGPTL4_KD (black), MP2_NTC (pink), MP2_ANGPTL4_OE + DsiRNA_NTC (turquoise), MP2_ANGPTL4_OE_ITGB4_KD (lavender), and MP2_ANGPTL4_OE_APOL1_KD (purple) cells treated with 0, 5nM, or 12.5nM gemcitabine. Sample size ranges from 4 to 11 per condition. P-value asterisks: ≤0.001=*** & ≤0.0001=****, unpaired, two-tailed parametric t-tests
Given the association of ANGPTL4 with resistance and patient outcome, we tested whether, like ANGPTL4, loss of ITGB4 or APOL1 expression increased sensitivity to gemcitabine in vitro. Using siRNAs, we knocked down APOL1 and ITGB4 in the MP2_ANGPTL4_OE cell line. We showed that treatment with the siRNAs reduced mRNA levels of the target genes (Supplementary Figure S2A-B), and we also found that knockdown of ITGB4 or APOL1 reduced expression of ANGPTL4 significantly compared to a non-targeting siRNA control with ANGPTL4 overexpression background; expression was reduced to 2% in MP2, 5% in ITGB4 knockdown, and 25% in APOL1 knockdown (Fig. 3C, Supplementary Table S1). Given our previous finding that overexpression of ANGPTL4 leads to resistance, we hypothesized that knockdown of APOL1 and ITGB4 might reverse resistance given the impact on ANGPTL4 expression. We tested the impact of ITGB4 and APOL1 knockout on gemcitabine sensitivity in the MP2_ANGPTL4_OE line. After gemcitabine treatment, cells with knockdown of ITGB4 or APOL1 in the ANGPTL4_OE background (MP2_ANGPTL4_OE_ITGB4_KD or MP2_ANGPTL4_OE_APOL1_KD) showed reversal of drug resistance when compared to the ANGPTL4 OE cells (Fig. 3D). There was no difference in growth rate among the lines that explained the differential viability observed with gemcitabine treatment (Supplementary Fig. S2C).
ANGPTL4 overexpression increases cell migration
To further understand the pathways that might be involved in resistance, we performed pathway enrichment analysis on the 1198 DEG associated with overexpression of ANGPTL4 (OE v KD). Those included tumor invasion and metastasis, proliferation, differentiation, inhibition of apoptosis, and cell membrane formation. Genes participating in invasion and metastasis included RhoEGF which was increased with ANGPTL4 expression. RhoEGF is linked to tumor invasion and metastasis [41]. Many genes in the Ras-ARF6 and Ras-MEK2 signaling pathways are also upregulated (e.g. FGF, APOL1, ITGB4, EGFR, MPK, CREB, cFOS, JUN, and VEGF) impacting proliferation and differentiation. In addition, ANTPGL4 overexpression is associated with the upregulation of anti-apoptotic pathways including JAK-STAT, Bcl_XS, and SOCS pathways triggered by upregulation of cytokine, cytokine receptors, CNTF, and IL22RA2. The formation of cellular structure was also affected by ANGPTL4 and increase in Claudin via the decrease in cell permeability (GEF-H1, Claudin2), increased actin assembly (ARP2/3), and cell polarity (AMPK, DLG1) (Fig. 4A, Supplementary Table S8).

ANGPTL4 overexpression correlation with EMT. (a) Pathway analysis of 1198 DEG revealed common cancer pathways. Red = log2FC > 0.7 and Blue = log2FC < 0.7. (b) Wound healing assay results at 0 h (after initial wound) and 48 h MP2_NTC (pink), MP2_ANGPTL4_OE + DsiRNA_NTC (turquoise), MP2_ANGPTL4_OE_ITGB4_KD (lavender), and MP2_ANGPTL4_OE_APOL1_KD (purple). Each set was performed in triplicate or quadruplicate. (c) Time to close half the gap (t1/2 gap) for each of the four cell types is plotted. t1/2: MP2 = 57.7 h, MP2_ANGPTL4_OE = 31.6 h., MP2_ANGPTL4_OE_ITGB4_KD = 192 h., and MP2_ANGPTL4_OE_APOL1_KD = 52.96 h. P-values ≤ 0.01=**, unpaired parametric t-test
Others have shown that on their own ANGPTL4, APOL1, and ITGB4 are each associated with cell proliferation and contribute to PDAC progression [17, The RNA-seq data generated in this study are available in Gene Expression Omnibus (GEO) at GSE207359. Clinical data and RNA-sequencing data for TCGA-PAAD (PDAC) samples were retrieved on 04/01/2020 using the GDC Data Portal (https://portal.gdc.cancer.gov/projects/TCGA-PAAD). Our analyses included 178 samples in this cohort that had matched clinical and RNA sequencing data. AKT Serine/Threonine Kinase 1 Protein Kinase AMP-Activated Catalytic Subunit Alpha 1 Angiopoietin Like 4 Actin Related Protein 2/3 Apolipoprotein L1 BCL2 Apoptosis Regulator Fos Proto-Oncogene, AP-1 Transcription Factor Subunit Ciliary Neurotrophic Factor Carbon dioxide CAMP Responsive Element Binding Protein dCas9 activation system Colony-stimulating factor 3 Cullin 7 Database for Annotation, Visualization, and Integrated Discovery Differentially expressed genes Discoidin Domain Receptor Tyrosine Kinase 2 Discs Large MAGUK Scaffold Protein 1 Dicer-substrate small interfering RNA Epithelial to mesenchymal transition Epidermal Growth Factor Receptor Mitogen-Activated Protein Kinase 1 Mitogen-Activated Protein Kinase 1/2 Fibroblast Growth Factor 1 Fibroblast Growth Factor Receptor 2 Fos Proto-Oncogene, AP-1 Transcription Factor Subunit Gene Expression Omnibus Gene Expression Profiling Interactive Analysis Rho/Rac Guanine Nucleotide Exchange Factor 2 Heparin Binding EGF-Like Growth Factor Inositol Polyphosphate-4-Phosphatase Type II B Integrin Subunit Beta 4 Janus Kinase 1 Mitogen-Activated Protein Kinase 8 Jun Proto-Oncogene, AP-1 Transcription Factor Subunit Knockdown Kyoto Encyclopedia of Genes and Genomes Lysine Demethylase 7 A KLF Transcription Factor 4 KRAS Proto-Oncogene, GTPase Interleukin 22 Receptor Subunit Alpha 2 Mitogen-Activated Protein Kinase MIA PaCa-2 cells Mitogen-Activated Protein Kinase Number Nanograms Nanomolar Notch Receptor Overexpression Overall survival P-value Pancreatic ductal adenocarcinoma Adjusted p-value Poly(ADP-Ribose) Polymerase Pancreatic ductal adenocarcinoma Phosphatidylinositol-4,5-Bisphosphate 3-Kinase Catalytic Subunit Delta Protein Phosphatase 1 Regulatory Subunit 15 A Quantitative polymerase chain reaction Recurrence-free survival Surveillance, Epidemiology, and End Results SH3 Domain Containing Ring Finger 2 Silencing RNA Suppressor of Cytokine Signaling SRY-Box Transcription Factor 9 Signal Transducer and Activator of Transcription 3 Signal Transducer and Activator of Transcription 3 STN1 subunit of CTS complex The Cancer Genome Atlas TGFB Induced Factor Homeobox 1 Transforming Growth Factor Beta Receptor 3 Microliter Versus Vascular Endothelial Growth Factor A Vascular Endothelial Growth Factor A WW Domain Containing Transcription Regulator 1 U.S. Mortality Data 1969–2019 when Using SEER*Stat: Surveillance, Epidemiology, and End Results (SEER) Program (www.seer.cancer.gov) SEER*Stat Database: Mortality - All COD, Aggregated With State, Total U.S. (1969–2019) [Internet]. 2021. Available from: http://www.cdc.gov/nchs. Sturm N, Ettrich TJ, Perkhofer L. The impact of biomarkers in pancreatic ductal adenocarcinoma on diagnosis, Surveillance and Therapy. Cancers. 2022;14(1):217. https://doi.org/10.3390/cancers14010217. Conroy T, Desseigne F, Ychou M, Bouché O, Guimbaud R, Bécouarn Y, Adenis A, Raoul JL, Gourgou-Bourgade S, de la Fouchardière C, Bennouna J, Bachet JB, Khemissa-Akouz F, Péré-Vergé D, Delbaldo C, Assenat E, Chauffert B, Michel P, Montoto-Grillot C, Ducreux M. ; Groupe Tumeurs Digestives of Unicancer; PRODIGE Intergroup. FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N Engl J Med. 2011;364(19):1817-25. doi: https://doi.org/10.1056/NEJMoa1011923. PMID: 21561347. American Cancer Society. Cancer Facts & Fig. 2022. Atlanta; 2022. Javle M, Shacham-Shmueli E, **ao L, Varadhachary G, Halpern N, Fogelman D, et al. Olaparib Monotherapy for previously treated pancreatic Cancer with DNA damage repair genetic alterations other than germline BRCA variants: findings from 2 phase 2 Nonrandomized clinical trials. JAMA Oncol. 2021;7:693–9. Moore MJ, Goldstein D, Hamm J, Figer A, Hecht JR, Gallinger S, et al. Erlotinib plus gemcitabine compared with gemcitabine alone in patients with advanced pancreatic cancer: a phase III trial of the National Cancer Institute of Canada clinical trials Group. J Clin Oncol American Society of Clinical Oncology. 2007;25:1960–6. Pishvaian MJ, Bender RJ, Halverson D, Rahib L, Hendifar AE, Mikhail S, et al. Molecular profiling of patients with pancreatic cancer: initial results from the know your tumor initiative. Clin Cancer Res American Association for Cancer Research Inc. 2018;24:5018–27. Kirby MK, Ramaker RC, Gertz J, Davis NS, Johnston BE, Oliver PG, et al. RNA sequencing of pancreatic adenocarcinoma tumors yields novel expression patterns associated with long-term survival and reveals a role for ANGPTL4. Mol Oncol. 2016;10:1169–82. Fernández-Hernando C, Suárez Y. ANGPTL4: A multifunctional protein involved in metabolism and vascular homeostasis. Current Opinion in Hematology 2020. page 206–13. Available from: https://doi.org/10.1097/MOH.0000000000000580. La Paglia L, Listì A, Caruso S, Amodeo V, Passiglia F, Bazan V, et al. Potential role of ANGPTL4 in the Cross talk between metabolism and Cancer through PPAR signaling pathway. PPAR Res. 2017. https://doi.org/10.1155/2017/8187235. Lei X, Shi F, Basu D, Huq A, Routhier S, Day R, et al. Proteolytic processing of angiopoietin-like protein 4 by proprotein convertases modulates its inhibitory effects on lipoprotein lipase activity. J Biol Chem American Society for Biochemistry and Molecular Biology Inc. 2011;286:15747–56. Kim SH, Park YY, Kim SW, Lee JS, Wang D, DuBois RN. ANGPTL4 induction by prostaglandin E 2 under hypoxic conditions promotes colorectal cancer progression. Cancer Res. 2011;71:7010–20. Tan MJ, Teo Z, Sng MK, Zhu P, Tan NS. Emerging roles of angiopoietin-like 4 in human cancer. Mol Cancer Res. 2012;10(6):677–88. https://doi.org/10.1158/1541-7786.MCR-11-0519. Zhu P, Tan MJ, Huang RL, Tan CK, Chong HC, Pal M, et al. Angiopoietin-like 4 protein elevates the Prosurvival Intracellular O2-:H2O2 ratio and confers Anoikis Resistance to Tumors. Cancer Cell Cell Press. 2011;19:401–15. Teo Z, Sng MK, Chan JSK, Lim MMK, Li Y, Li L, et al. Elevation of adenylate energy charge by angiopoietin-like 4 enhances epithelial-mesenchymal transition by inducing 14-3-3γ expression. Oncogene. 2017;36:6408–19. Zhang K, Zhai Z, Yu S, Tao Y. DNA methylation mediated down-regulation of ANGPTL4 promotes colorectal cancer metastasis by activating the ERK pathway. J Cancer. 2021;12:5473–85. Zhao J, Liu J, Wu N, Zhang H, Zhang S, Li L, et al. ANGPTL4 overexpression is associated with progression and poor prognosis in breast cancer. Oncol Lett. 2020;20:2499–505. Cai YC, Yang H, Wang KF, Chen TH, Jiang WQ, Shi YX. ANGPTL4 overexpression inhibits tumor cell adhesion and migration and predicts favorable prognosis of triple-negative breast cancer. BMC Cancer. 2020;20. https://doi.org/10.1186/s12885-020-07343-w. Yan HH, Jung KH, Lee JE, Son MK, Fang Z, Park JH, et al. ANGPTL4 accelerates KRASG12D-Induced acinar to ductal metaplasia and pancreatic carcinogenesis. Cancer Lett. 2021;519:185–98. Gupta N, Park JE, Tse W, Keem Low J, Kon OL, Mccarthy N, et al. ERO1α promotes hypoxic tumor progression and is associated with poor prognosis in pancreatic cancer. Oncotarget. 2019;10(5):5870–982. https://www.oncotarget.com/article/27235/text/. Joung J, Konermann S, Gootenberg JS, Abudayyeh OO, Platt RJ, Brigham MD, Sanjana NE, Zhang F. Genome-scale CRISPR-Cas9 knockout and transcriptional activation screening. Nat Protoc. 2017;12(4):828–63. https://doi.org/10.1038/nprot.2017.016. Larson B, Held P, Banks P. Optimization of Perkin Elmer’s AlphaScreen® cAMP assay on the Synergy™ H4 hybrid Multi-Mode Microplate Reader [Internet]. 2010. http://www.biotek.com. Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15. https://doi.org/10.1186/s13059-014-0550-8. Ge SX, Son EW, Yao R, iDEP. An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinformatics. 2018;19. https://doi.org/10.1186/s12859-018-2486-6. Baecker V. ImageJ Macro Tool Sets for Biological Image Analysis. Conference. Kanehisa M, Sato Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020;29:28–35. Mi H, Huang X, Muruganujan A, Tang H, Mills C, Kang D, Thomas PD. PANTHER version 14: more genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019;47(D1):D419–26. Therneau T. A Package for Survival Analysis in R. R package version 3.4-0 [Internet]. 2022. https://CRAN.R-project.org/package=survival. Kassambara A, Kosinski M, Biecek F. S.Package “survminer” Type Package Title Drawing Survival Curves using “ggplot2;” 2022. Wickham H. Ggplot2: elegant graphics for data analysis. Springer; 2009. Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15(12). https://doi.org/10.1186/s13059-014-0550-8. Kolde R. Pretty Heatmaps. R package version 1.0.12 [Internet]. 2019. Available from: https://CRAN.R-project.org/package=pheatmap. Frank M, Harrell E, Hmisc. A package of miscellaneous R functions [Internet]. 2020. Available from: http://biostat.mc.vanderbilt.edu/Hmisc. Urbanek S, Johnson K, tiff. Read and Write TIFF Images. R package version 0.1–11. 2020; Available from: https://CRAN.R-project.org/package=tiff. Jia Y, **e J. Promising molecular mechanisms responsible for gemcitabine resistance in cancer. Genes and Diseases. 2015;2(4):299–306. https://doi.org/10.1016/j.gendis.2015.07.003. Wei X, Zhou X, Zhao Y, He Y, Weng Z, Xu C. A 14-gene gemcitabine resistance gene signature is significantly associated with the prognosis of pancreatic cancer patients. Sci Rep. 2021;11(6087). https://doi.org/10.1038/s41598-021-85680-x. Akada M, Crnogorac-Jurcevic T, Lattimore S, Mahon P, Lopes R, Sunamura M, et al. Intrinsic chemoresistance to Gemcitabine is Associated with decreased expression of BNIP3 in pancreatic Cancer. Clin Cancer Res. 2005;11(8):3094–101. https://aacrjournals.org/clincancerres/article/11/8/3094/188241/Intrinsic-Chemoresistance-to-Gemcitabine-Is. Nakano Y, Tanno S, Koizumi K, Nishikawa T, Nakamura K, Minoguchi M, et al. Gemcitabine chemoresistance and molecular markers associated with gemcitabine transport and metabolism in human pancreatic cancer cells. Br J Cancer. 2007;96:457–63. Vasaikar SV, Deshmukh AP, den Hollander P, Addanki S, Kuburich NA, Kudaravalli S, Joseph R, Chang JT, Soundararajan R, Mani SA. EMTome: a resource for pan-cancer analysis of epithelial-mesenchymal transition genes and signatures. Br J Cancer. 2021;124(1):259–69. https://doi.org/10.1038/s41416-020-01178-9. Epub 2020 Dec 10. PMID: 33299129; PMCID: PMC7782839. Huang L, Jansen L, Balavarca Y, Babaei M, van der Geest L, Lemmens V, Van Eycken L, De Schutter H, Johannesen TB, Primic-Žakelj M, Zadnik V, Besselink MG, Schrotz-King P, Brenner H. Stratified survival of resected and overall pancreatic cancer patients in Europe and the USA in the early twenty-first century: a large, international population-based study. BMC Med. 2018 Aug 21;16(1):125. doi: https://doi.org/10.1186/s12916-018-1120-9. PMID: 30126408; PMCID: PMC6102804. Ghanem NZ, Matter ML, Ramos JW. Regulation of Leukaemia Associated rho GEF (LARG/ARHGEF12). Small GTPases. 2022;13(1):196–204. https://doi.org/10.1080/21541248.2021.1951590. Lin J, Xu Z, **e J, Deng X, Jiang L, Chen H, Peng C, Li H, Zhang J, Shen B. Oncogene APOL1 promotes proliferation and inhibits apoptosis via activating NOTCH1 signaling pathway in pancreatic cancer. Cell Death Dis. 2021;12(8). https://doi.org/10.1038/s41419-021-03985-1. Cruz-Monserrate Z, Qiu S, Evers BM, O’Connor KL. Upregulation and redistribution of integrin α6β4 expression occurs at an early stage in pancreatic adenocarcinoma progression. Mod Pathol. 2007;20:656–67. Duchateau PN, Pullinger CR, Orellana RE, Kunitake ST, Naya-Vigne J, O’Connor PM, et al. Apolipoprotein L, a new human high density lipoprotein apolipoprotein expressed by the pancreas. Identification, cloning, characterization, and plasma distribution of apolipoprotein L. J Biol Chem. 1997;272:25576–82. Cho S, Lu M, He X, Ee PLR, Bhat U, Schneider E, et al. Notch1 regulates the expression of the multidrug resistance gene ABCC1/MRP1 in cultured cancer cells. Proc Natl Acad Sci. 2011;108:20778–83. Sangphech N, Keawvilai P, Palaga T. Notch signaling increases PPARγ protein stability and enhances lipid uptake through AKT in IL-4-stimulated THP-1 and primary human macrophages. FEBS Open Bio. 2020;10:1082–95. Mohanty A, Nam A, Pozhitkov A, Yang L, Srivastava S, Nathan A et al. A Non-genetic Mechanism Involving the Integrin β4/Paxillin Axis Contributes to Chemoresistance in Lung Cancer. iScience 2020;23. https://doi.org/10.1016/j.isci.2020.101496. Yoshioka T, Otero J, Chen Y, Kim YM, Koutcher JA, Satagopan J, et al. β4 integrin signaling induces expansion of prostate tumor progenitors. J Clin Invest. 2013;123:682–99. Masugi Y, Yamazaki K, Emoto K, Effendi K, Tsujikawa H, Kitago M, Itano O, Kitagawa Y, Sakamoto M. Upregulation of integrin β4 promotes epithelial-mesenchymal transition and is a novel prognostic marker in pancreatic ductal adenocarcinoma. Lab Invest; 2015;95(3):308 – 19. doi: https://doi.org/10.1038/labinvest.2014.166. PMID: 25599535. Giancotti FG, Ruoslahti E. Integrin signaling. Science. 2017;285:1028–32. Meng X, Liu P, Wu Y, Liu X, Huang Y, Yu B, et al. Integrin beta 4 (ITGB4) and its tyrosine-1510 phosphorylation promote pancreatic tumorigenesis and regulate the MEK1-ERK1/2 signaling pathway. Bosn J Basic Med Sci. 2020;20:106–16. This work was funded by NIH 1R43CA232844-01A1 and the State of Alabama Cancer Research Fund. SJC is supported by UL1TR003096. We would like to thank Anuj Singhal and his team at CFD Research as well as our HudsonAlpha colleagues, especially those in the Myers lab for their feedback, support, and discussions of this work. Figure 4 A created with BioRender.com agreement number RD24SQPZ3K. This work was funded by NIH 1R43CA232844-01A1 and the State of Alabama Cancer Research Fund. ERG, CAW, and SJC designed the experiments. ERG, CAW, and MJ collected data. ERG and CAW analyzed the data. ERG and SJC wrote the first draft. All authors contributed to the writing of the paper and read and approved the final manuscript. The authors declare that they have no competing interests. All methods were carried out in accordance with relevant guidelines and regulations. Not applicable. Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. Below is the link to the electronic supplementary material. Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data. Gordon, E.R., Wright, C.A., James, M. et al. Transcriptomic and functional analysis of ANGPTL4 overexpression in pancreatic cancer nominates targets that reverse chemoresistance.
BMC Cancer 23, 524 (2023). https://doi.org/10.1186/s12885-023-11010-1 Received: Accepted: Published: DOI: https://doi.org/10.1186/s12885-023-11010-1Data Availability
Abbreviations
References
Acknowledgements
Funding
Author information
Authors and Affiliations
Contributions
Corresponding author
Ethics declarations
Competing interests
Ethics approval and consent to participate
Consent for Publication
Additional information
Publisher’s Note
Electronic supplementary material
Rights and permissions
About this article
Cite this article
Keywords
