
Abstract
Background
Uveal melanoma is a rare form of cancer with high mortality. The incidence of metastases is attributed to early seeding of micrometastases from the eye to distant organs, primarily the liver. Once these seeded clusters of dormant tumor cells grow into larger radiologically detectable macrometastases, median patient survival is about 1 year. Melatonin is an important hormone for synchronizing circadian rhythms. It is also involved in other aspects of human physiology and may offer therapeutic benefits for a variety of diseases including cancer.
Methods
Articles involving the physiological effects of melatonin, pharmacokinetics, and previous use in cancer studies were acquired using a comprehensive literature search in the Medline (PubMed) and Web of Science databases. In total, 147 publications were selected and included in the review.
Results
Melatonin has been observed to suppress the growth of cancer cells, inhibit metastatic spread, enhance immune system functions, and act as an anti-inflammatory in both in vitro and in vivo models. Melatonin may also enhance the efficacy of cancer treatments such as immuno- and chemotherapy. Numerous studies have shown promising results for oral melatonin supplementation in patients with other forms of cancer including cutaneous malignant melanoma. Cell line and animal studies support a hypothesis in which similar benefits may exist for uveal melanoma.
Conclusions
Given its low cost, good safety profile, and limited side effects, there may be potential for the use of melatonin as an adjuvant oncostatic treatment. Future avenues of research could include clinical trials to evaluate the effect of melatonin in prevention of macrometastases of uveal melanoma.
Similar content being viewed by others


Introduction
Uveal melanoma is the most common primary intraocular malignancy in adults and is associated with a high incidence of metastasis, a high mortality rate, and poor response to current treatments [1,2,3]. The cancer originates from uveal melanocytes and possesses several features seen in other melanocytic tumors of the central nervous system (CNS) [4]. Uveal melanoma is rarely present in children and the incidence increases with advancing age. Some studies suggest that uveal melanoma has a higher incidence rate in males over the age of 65, while other studies showed no sex predilection [5,6,7]. Strong evidence for incidence variation between different ethnic groups exists, where non-Hispanic white individuals have the highest incidence, with an overall non-Hispanic white:black incidence ratio of 18:1 [8]. Incidence rates for cases per million per year are 5.1 in the USA (1973–2008) and 1.3–8.6 in Europe (1983–1994) with generally increasing incidence from southern to northern regions [9, 10]. No major differences in survival have been observed when stratifying cumulative uveal melanoma-related mortality rates by year of diagnosis [11]. The estimated five, ten, 15, 20, 25 and 30-year relative survival rates are 79, 66, 60, 60, 62 and 67%, respectively [12].
Click or tap here to enter text.
There are two predominant types of risk factors for uveal melanoma: genetic and environmental. There are several low-risk genetic loci which include several single nucleotide polymorphisms (SNPs) in genes encoding pigment proteins (HERC2, OCA2, IRF4) as well as other genes including TERT which encodes for telomerase reverse transcriptase, and CLPTM1L, which encodes cleft lip and palate transmembrane protein 1-like protein. High-risk loci have been identified in the genes BAP1, MLH1, PALB2, and SMARCE1 [13]. Other risk factors include atypical or common cutaneous nevi, welding, occupational cooking, fair skin color, light eye color, sunburn, iris nevi, and cutaneous freckles [14]. The quantity and quality of melanin appears to play an important role in uveal melanocytes, where there are two types of melanin; black to brown eumelanin, and yellow to red pheomelanin. Eumelanin is associated with dark eyes while pheomelanin is associated with light eyes and it has been suggested that eumelanin acts as an antioxidant while pheomelanin acts as a pro-oxidant which may help explain its correlation to a higher incidence of uveal melanoma [15].
Inflammation has been suggested to play a key role in the development of uveal melanoma with high numbers of CD68+ macrophages being associated with heavy pigmentation, microvascular density, epithelioid cells, and increased 10-year uveal melanoma-related mortality rate [16].
The most common cytogenetic aberrations associated with uveal melanoma include monosomy of chromosome 3 and an isochromosome of 8q [17]. In turn, monosomy 3 is associated with mutations in the tumor suppressor gene BAP1, located on the short arm of chromosome 3 and is associated with uveal melanoma [18]. In the absence of BAP1 mutations, other gene mutations are usually present, such as SF3B1, a gene associated with late-onset metastasis and EIF1AX, associated with low-risk profile of uveal melanoma and a low risk for metastasis [19, 20]. Somatic mutations in GNAQ and GNA11 subunits of Gq-protein have been correlated to uveal melanoma oncogenesis, but do not correlate with metastatic risk [21]..
Primary uveal melanoma is typically treated by enucleation or eye-preserving plaque brachytherapy. Despite treatment, metastatic rates reach 30 to 50% even with removal of the eye due to the early seeding of micrometastases [22, 23]. These treatments result in similar survival rates for medium-sized tumors, which are defined as tumors between 2.5 mm and 10.0 mm in height, though no more than 8.0 mm in height whenever the tumor is near the disc, and no more than 16.0 mm in diameter [24].
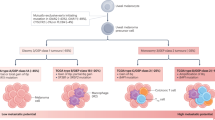
Tumor cells migrate from the intraocular tumor via a strictly hematogenous route to the systemic circulation from which they can extravasate and establish micrometastases (colonies of tumor cells that are too small to cause symptoms or to be detectable by radiological examinations), primarily in the spleen and liver [25]. The latter is the most common site for metastasis, which may be related to the affinity of cMET-ligands by uveal melanoma-cells to HGF-receptors of hepatic stellate cells [26]. Estimations based on tumor doubling times indicate that the seeding of metastases occurs well before detection of the average primary tumor [27, 28]. The micrometastases can remain dormant in their metastatic niche for years or decades before switching to proliferation and macrometastasis [29]. At that point, mean patient survival is ~ 1 year, with low response rates to immune- and chemo- therapy [13]. As such, additional adjuvant treatments for uveal melanoma and their metastases are of great therapeutic need.
Aims
Melatonin has been studied as an oncostatic agent and has shown promising results in breast, prostate, colon, and liver cancers [30,31,32,33]. However, research regarding melatonin’s potential therapeutic role in uveal melanoma is limited. This review aims to explore current data surrounding melatonin as an oncostatic agent and whether future clinical studies should be performed to further investigate the effects of melatonin as an adjuvant treatment in uveal melanoma.
Literature search
Data were acquired with a comprehensive literature search in the Medline (PubMed) and Web of Science databases for peer reviewed published articles that described relevant results. The following search terms were used and matched to appropriate medical subject headings: (“Melatonin” AND (“physiological effects” OR “cancer” OR “animal studies” OR “oncostatic effect” OR “mechanism of action” OR “pharmacokinetics” OR “clinical trial”)). The search strategy was restricted to titles and/or abstracts of human clinical studies published in English. No restrictions based on year of publication were made. The latest search was performed on August 1st, 2021. All available studies were included and could be accessed in full via the University Library, Karolinska Institutet. Trial registries, unpublished studies, grey literature, animal studies, laboratory studies, letters to the editor, correspondence, notes, editorials, and conference abstracts were not considered. Reference lists of included articles were searched for additional studies.
The selection of articles for this analysis was performed in three steps as illustrated in Fig. 1: Identification, title, abstract and full-text screening, and inclusion. Abstract screening of articles identified in the literature search was done independently by the two first authors, with any disagreements resolved by discussion. Publications were included for full-text screening if they reported relevant results. The search string resulted in a total of 1293 publications. After title, abstract and full-text screening, 147 publications remained in this review. Studies were excluded if they 1) were earlier versions of a series of articles from the same database or center or 2) reported patients that were already included in another publication. Studies including patients with primary conjunctival or orbital melanomas or metastatic lesions were not considered. The same inclusion and exclusion criteria applied to full-text screening (if not evident in title or abstract). The preferred reporting items for systematic reviews and meta-analyses (PRISMA) were not followed as this is a narrative review rather than a systematic review or a meta-analysis.

Literature search. The selection of articles for this analysis was performed in three steps. Articles were identified using specific search terms, abstracts were then screened, and lastly, full text articles were assessed for eligibility based on our exclusion criteria. Through this process 147 articles were selected and included in this review
Review
Melatonin
Rene Descartes described the pineal gland as the “seat of the soul”. The earliest indications that the pineal gland was biologically active were described ~ 2000 years ago by the Greek physician Galen in his 8th work De Usu Partium — On the Usefulness of the Parts of the Body [34]. Evidence that the pineal gland produced a biologically active substance was published as early as 1917 [35]. Later, this substance was named melatonin, based on the observation that extracts from bovine pineal glands produced a skin lightening response when applied to frog skin. In 1958, Lerner et al. isolated melatonin itself (5 methoxy-N-acetyltryptamine) [36]. Melatonin is involved in numerous biological processes, playing an important role in human physiology, and may offer therapeutic benefits for a variety of diseases. Systemic melatonin is mainly secreted by the pineal gland, however, additional sources include the retina, skin, bone marrow cells, and the gastrointestinal tract [37,38,39,40,41].
Melatonin was previously thought to be made in the cytosol, however, more recent research suggests that melatonin is primarily produced in the mitochondria [42]. Melatonin is synthesized from serotonin which in turn is synthesized from the amino acid tryptophan (Fig. 2). In the pineal gland, tryptophan is taken up from the blood and hydroxylated to 5-hydroxytryptophan which is subsequently decarboxylated to serotonin. Serotonin is in turn converted to N-acetylserotonin and then converted to melatonin ready to be released into the blood and cerebrospinal fluid [43,44,45].

Melatonin synthesis pathway. Tryptophan is hydroxylated by tryptophan hydroxylase (TPH) to 5-hydroxytryptophan which is subsequently decarboxylated by aromatic amino acid decarboxylase (AADC) resulting in serotonin. Serotonin is converted to N-acetylserotonin by Arylalkylamine N-acetyltransferase (AA-NAT) which is then catalyzed by N-acetylserotonin O-methyltransferase (ASMT) to produce melatonin
Melatonin is an evolutionarily preserved molecule found in all microorganisms, plants, and animals [46]. Key functions of melatonin include the regulation of circadian patterns, such as fluctuations in core body temperature and sleep-wake cycles, seasonal reproduction, immune system enhancement, and glucose regulation [47, 48]. While melatonin performs several hormonal functions, this unique molecule also exhibits paracrine and autocrine effects and acts as an antioxidant and a free radical scavenger [49,50,51].
Endogenous production
While melatonin is produced by cells in several types of tissue throughout the body, the primary source of systemic melatonin is the pineal gland. Unlike other endocrine glands, the pineal gland does not store the melatonin it produces but, instead, secretes it directly into the blood and cerebrospinal fluid. From the vasculature, melatonin is distributed across various fluids and tissues including saliva, urine, amniotic fluid, semen, and breast milk [48]. The secretion of melatonin is mediated by light and dark cycles via the suprachiasmatic nucleus (SCN) in the hypothalamus. Light is detected by intrinsically photoreceptive retinal ganglion cells (ipRGC; also named melanopsin sensitive retinal ganglion cells) in the retina which are particularly sensitive to blue light (480 nm), found in natural sunlight as well as in artificial light sources such as LEDs [52, 53]. This information is relayed to the SCN via the retinohypothalamic tract before forming a complex signaling pathway to the paraventricular nuclei (PVN), the intermediolateral nucleus of the spinal cord (IML), the superior cervical ganglion (SCG), and finally, the pineal gland [54]. Production of melatonin takes place at night, in the absence of light (Fig. 3) [41].

The anatomy of melatonin secretion. The production and release of melatonin are mainly mediated by postganglionic retinal nerve fibers that exits the eye through the optic nerve, pass through the retinohypothalamic tract to the suprachiasmatic nucleus and then to the pineal gland via the superior cervical ganglion. This axis is activated by darkness and suppressed by light. The circadian rhythm of melatonin secretion is to a lesser extent also controlled by the suprachiasmatic nucleus
During the day, serum concentrations of melatonin remain low (10 pg/ml), however, levels begin to increase after sunset, reaching a peak concentration of ~ 60 pg/ml between 2:00 and 4:00 a.m. [55]. Peak concentrations vary among individuals and have been found to be the highest in children between the ages of one and five, thereafter decreasing with age [56]. Meanwhile, melatonin concentrations have been found to be lower in individuals with certain conditions including depression, Alzheimer’s disease, age-related macular degeneration, idiopathic scoliosis, and autism spectrum disorders [57,58,59,60,61].
When taken orally, a 0.5 mg tablet of exogenous melatonin is equivalent to physiological levels of endogenous melatonin [62]. Exogenous melatonin is commonly taken as a supplement to combat insomnia as well as to ameliorate the effects of jetlag, however, there is increasing evidence suggesting that taking exogenous melatonin may have benefits in a variety of diseases including cancer [63].
Pharmacokinetics
Melatonin, whether administered orally or intravenously, is absorbed via first-order kinetics. Melatonin administered orally has a half-life of absorption of 6 min. Tmax of 10 mg orally administrated melatonin was 41 min in a study, which is in agreement with early studies where Tmax varied between 46 to 65 min when doses ranged between 0.5 to 6 mg. When administered intravenously, the elimination of 10 mg of melatonin was 39 min, which corresponded to early studies where the elimination varied between 28 and 60 min for doses ranging between 0.005 to 2 mg. Melatonin has low bioavailability due to hepatic first-passage metabolism, at about 3% with some inter-individual variability. Other studies have suggested higher bioavailability between 9 and 33% with similar findings in regards to inter-individual variability [64]. The metabolism of melatonin occurs primarily in the liver through hydroxylation by the enzyme CYP1A2 and conjugation with sulfuric or glucuronic acid. Melatonin undergoes secondary metabolism in the kidney and is excreted in the urine as an inactive metabolite; 6-sulfatoxymelatonin (6-SM), which is used as a reliable marker of melatonin production [62, 65].
Mechanism of action
Melatonin is lipophilic and interacts with a broad variety of receptors including specific membrane receptors, nuclear receptors, intracellular proteins such as calmodulin, in addition to acting as a direct antioxidant and free radical scavenger [66]. Primary receptors for melatonin include the G-coupled transmembrane receptors MT1 and MT2 which are widely expressed throughout the human body as described in Table 1 [67,68,69,70,71,72,73,74,75,76].
A third binding site with lower affinity to melatonin compared to MT1 and MT2 was described in 1988 and dubbed MT3 (ML-2), however, it was later discovered that MT3 was in fact the previously characterized enzyme, quinone reductase 2 (QR2), a protein with detoxifying properties [77,78,79]. It is of note that melatonin binding sites tend to be expressed in low densities throughout various tissues which may be due to the high affinity of melatonin for its receptors [72].
Melatonin may also activate certain nuclear receptors in the retinoid orphan receptor (ROR), or retinoid Z receptor (RZR), family, however, whether melatonin interacts with these receptors directly remains uncertain [80, 81].
In order for melatonin to perform intracellular actions, it must be available intracellularly. A study by Hevia et al. found that members of the SLC2/GLUT family glucose transporters contribute to melatonin uptake. This has potential consequences including that high blood glucose concentrations may impact a cell’s ability to take up melatonin by acting as a competitive ligand of GLUT1. On the other hand, melatonin may reduce the uptake of glucose to cells including tumor cells, as suggested by a study performed in prostate cancer cells [82].
Physiological effects
Melatonin is a chronobiotic molecule which synchronizes a vast variety of central and peripheral biological functions as an adaptation to the 24-h day-night cycle by binding to the aforementioned receptors. In the absence of light, signals from the SCN are relayed to the pineal gland and stimulate the secretion of melatonin. Melatonin then travels to the SCN and binds to MT1 and MT2 receptors where MT1 activation leads to an inhibition of neuronal activity which normally promotes wakefulness and MT2 activation causes a phase shift in circadian firing rhythms [83, 84]. A recent study conducted in mice demonstrated that the activation of MT1 receptors primarily regulate rapid eye movement (REM) sleep while the activation of MT2 receptors selectively increases non-rapid eye movement (NREM) sleep [85]. Whilst melatonin is most well-known for its regulation of the sleep-wake cycle, it also plays an important role in neural development, blood glucose regulation, immune system function, sexual maturation and the cardiovascular system [86,87,88,89,90,91,92,93,94,95,96].
Neurodevelopment
Melatonin plays a significant role in human development in utero where the hormone exhibits effects on placenta function and early nervous system development. Niles et al. demonstrated in vitro that melatonin activates glial cell-line derived neurotrophic factor (GDNF) and that neural stem and progenitor cells express MT1 receptors [86]. These results suggest a potential neuroprotective role of melatonin and may explain why sleep deprivation has been found to suppress neurogenesis in the mouse hippocampus while melatonin supplementation rescues these deficits [97].
Reproduction and sexual maturation
The process of sexual maturation may also be impacted by melatonin [87]. Existing evidence indicates that the decrease in nocturnal melatonin levels during adolescence correlates to sexual maturation. This is further supported by the inverse relationship between levels of luteinizing hormone and melatonin at various stages of puberty and the decline in salivary melatonin levels with advancing tanner stages in both boys and girls [56, 88]. There has been some concern that long-term exposure to exogenous melatonin may alter the timing of puberty onset in adolescents as seen in some animal studies, however more research is needed before drawing similar conclusions in humans [98, 99].
Cardiovascular system
Beneficial effects of melatonin on various cardiovascular risk factors such as hyperlipidemia, hypertension, and obesity have been demonstrated in numerous studies [89]. Celinski et al. demonstrated that melatonin supplementation at 5 mg twice a day for 14 months significantly lowered LDL cholesterol and triglyceride levels in patients with nonalcoholic fatty liver disease [90]. Supporting this, Scheer et al. demonstrated that melatonin supplementation may lower blood pressure in patients with untreated essential hypertension [91]. In kee** with these findings, a study performed in rats by Simko et al. showed that continuous light exposure can lead to hypertension, left ventricle hypertrophy and fibrosis, as well as increased oxidative stress in the left ventricle and aorta [100].
Immune system regulation
It is hypothesized that one of melatonin’s key roles throughout evolution involved protecting organisms from environmental stressors by acting as a potent antioxidant [50]. Melatonin detoxifies numerous reactive oxygen and nitrogen species including hydrogen peroxide, hydroxyl radical, singlet oxygen, peroxyl radicals, and nitric oxide radicals. Several in vivo studies have found melatonin to be a more effective antioxidant than the well-known vitamin C and vitamin E in terms of protection from tissue damage [92, 93]. In addition to acting as an antioxidant itself, melatonin upregulates the expression of anti-oxidative enzymes such as Cu/Zn-superoxide dismutase (CuZn-SOD), Mn-superoxide dismutase (Mn-SOD), catalase, and glutathione peroxidase (GPx) [94]. The hormone also enhances T- and NK- cell responses in addition to stimulating the production of cytokines and interleukins including IL-2, IL-6, and IL-12 [95, 96].
In uveal melanoma, the occurrence of micrometastases is dependent on a delicate balance between supportive and inhibitory interactions between the tumor cells, the surrounding stroma and the immune system [29, 101, 102]. While response rates are low for CTLA4 and PD-L1 immune checkpoint inhibition in metastatic uveal melanoma, other immune checkpoints are at play and increasing the T- and NK- cell response in the metastatic niche may inhibit both survival and growth of uveal melanoma micrometastases [103].
Melatonin and mitochondria
Melatonin is an effective antioxidant in mitochondria. Mitochondria synthesize melatonin locally in addition to performing active take-up from cytosolic pools derived from systemic circulation [104,105,106]. Melatonin counteracts oxidative stress and promotes the activity of enzymes involved in oxidative phosphorylation, increasing ATP production in neuronal and hepatic mitochondria in rats [107]. Yamamoto et al. demonstrated that melatonin reversed the deadly effects of cyanide, a complex IV inhibitor, in mice [108].
Blood glucose
Melatonin plays a role in the regulation of blood glucose levels via its association with MT1 and MT2 [109]. When melatonin binds MT1 and MT2 receptors present in the pancreas, insulin secretion is reduced via a receptor mediated inhibition of the cyclic adenosine monophosphate (cAMP) and cyclic guanosine monophosphate (cGMP) pathways [110, 111]. On the other hand, activation of MT1 and MT2 receptors on pancreatic alpha cells has been shown to increase glucagon secretion via Gαq-coupled and PI3K signalling pathways [112]. Further supporting the role of melatonin in blood glucose regulation, studies have found that pinealectomized rats display an altered daily rhythm in glucose levels with increased glucose concentrations at night and mice lacking MT1 receptors entirely have higher overall mean blood glucose levels [113, 114].
Therapeutic effects
Previous research indicates a therapeutic benefit of melatonin for multiple types of cancer. In patients with advanced and metastatic non-small cell lung cancer were treated with 10 mg of oral melatonin in a cycle consisting of 21 treatment days followed by 7 rest days, as compared to supportive care alone, there was a significant increase in both one-year survival and disease stabilization without drug-related toxicity [115]. In another study, melatonin had a small therapeutic effect on stage 3 and 4 human metastatic malignant melanoma and with few side effects despite a relatively high oral dose of 50 mg 4 times a day for a minimum of 2 months [116]. Lissoni et al. demonstrated that in patients with metastatic solid tumors, including non-small cell lung cancer, breast cancer, gastro-intestinal tract tumors as well as head and neck cancers, there was a higher survival rate and a longer mean time to progression for patients treated with 20 mg oral melatonin 7 days prior to chemotherapy compared to those treated with chemotherapy alone. This is potentially indicative of the antioxidative effects of melatonin which may help mediate chemotherapy toxicity and even enhance of the efficacy of chemotherapies on cancer cells [117]. Oral melatonin at 40 mg daily amplifies the anti-tumor activity of IL-2 treatments in patients with solid neoplasms when given 7 days before IL-2 is administered [118]. In glioblastoma patients treated with radiotherapy in addition to 20 mg oral melatonin daily until disease progression, there was an increase in one-year survival rate as well as lower therapy-related toxicity compared to radiotherapy alone [119]. When adding 20 mg of melatonin daily to 14 women with metastatic breast cancer who did not respond to tamoxifen therapy alone or progressed after an initially stable disease status, a partial response was achieved in four participants (29%). Mean serum levels of the breast cancer growth factor IGF-1 and patient anxiety levels were significantly decreased [120].
An in vitro study on endometrial adenocarcinomas demonstrated that a physiological concentration of melatonin at 10− 9 M for 4 h and 96 h incubation showed no inhibition on SNG-II estrogen receptor-negative, endometrial cancer cell growth. However, melatonin at the same concentration had a significant inhibition effect on Ishikawa estrogen receptor-positive endometrial cancer cell growth [121]. In ovarian adenocarcinoma BG-1, treatment with melatonin at a concentration of 10− 9 – 10− 7 M for 48 h resulted in a 20–25% reduction in the number of cancer cells in vitro [122].
Several studies have shown that melatonin is beneficial when used in conjunction with other anti-cancer drugs and radiation treatments by reducing their toxicity on healthy human cells while enhancing the drugs’ effect on cancer cells [123,124,125,126,127,128]. This may be due to the antioxidative properties of Melatonin which neutralizes toxic hydroxyl radicals responsible for cell damage at the nucleic acid level, protecting from ionized radiation as well as from single- and double- stranded breaks in DNA, offering resilience to DNA modifications in healthy cells [129]. At the same time melatonin’s enhancement of the efficacy of chemotherapy drugs may be related to its pro-apoptotic features [130]. A review by Wang et al. found that Melatonin decreased radiochemotherapy-related side effects such as thrombocytopenia (Relative risk, RR = 0.13), neurotoxicity (RR = 0.19), and fatigue (RR = 0.37) in patients with solid tumors including [125]. Another study found that melatonin treatment protects bone marrow when used prior to whole body radiation or when combined with proteasome-inhibitor drugs such as Bortezomib, melatonin reduces the drug’s toxic effects [131]. Similarly, when combining cisplatin with melatonin in the treatment of ovarian cancer melatonin protected normal ovarian cells from cisplatin’s toxicity [132, 133]. Furthermore, melatonin supports the apoptotic effects of Doxorubicin when both are applied to human breast cancer cells [132].
Several in vitro studies suggest that the concentration of melatonin used may impact its effects on malignancies. For example, a study conducted using rodent melanoma cells by Slominski et al. demonstrated that low concentrations of melatonin (0.1–10 nM) inhibited cell proliferation without impacting melanogenesis while high concentrations (> 0.10 μM) inhibited melanogenesis while having no effect on cell proliferation and that particularly high concentrations (100 μM) were found to stimulate cell proliferation [134]. Similarly, an in vitro study utilizing two cell sublines of mouse melanoma found that a melatonin concentration of 1 nM significantly inhibited cell proliferation while sub-physiologic concentrations (0.1 pM) and supraphysiologic concentrations (100 nM and 10 μM) had no impact [135]. Fisher et al. demonstrated that melatonin suppressed cell proliferation at all tested concentrations (1 mM – 100 nM) in four different human melanoma cell lines [136]. These studies suggest that further research is required to assess the dose dependent actions of melatonin in inhibiting cancer cell proliferation.
Oncostatic effects
In addition to melatonin’s immunoenhancing features and its synergistic effects when used in conjunction with anti-cancer drugs, it possesses several oncostatic properties. A meta-analysis published in 2005 evaluated the effects of melatonin in cancer treatment where melatonin was either used as the sole treatment or as an adjuvant therapy for patients with solid tumors. Ten randomly controlled trials were reviewed, and results showed an overall 44% reduction in the risk of death at 1 year [63]. Oncosuppressive mechanisms mediated by melatonin are summarized in Fig. 4.

Oncosuppressive mechanisms mediated by melatonin. Melatonin (MLT) activates the high-affinity G protein-coupled receptors MT1 and MT2 which reduce the transcriptional activity of NF-κB and activates phosphorylation cascades mediated by mitogen-activated protein kinases (MAPKs) including MEK1/2, ERK1/2, JNK, and p38. In turn, NF-κB inhibition and MAPKs activation inhibit cell growth and motility and promote apoptosis and DNA damage repair through accumulation of oncosuppressors such as p53, p27kip1, and p21. NF-κB inhibition and MAPKs activation also activates DNA repair complexes such as P53/PML/H2AX on DNA damage sites, and transcriptional control of genes involved in the cell cycle, apoptosis, and invasiveness. Further, melatonin can bind to the intracellular protein calmodulin (CaM) and reduce the Estrogen Receptor α (Erα) response in ER positive cells by impairing the formation of a proper E2–Erα–CaM complex on target genes. Melatonin downregulates the nuclear RZR receptors (RZR alpha, RZR beta, ROR alpha 1, RZR alpha 2, ROR alpha 3 and ROR gamma), inhibiting growth, angiogenesis and HIF-1α activity. Arrows indicate activation, while dashed blunt lines indicate inhibition. Activation indicates an increase in protein or activity levels, while inhibition indicates a decrease in protein or activity levels
Apoptosis and tumor invasion
Melatonin is involved in cell cycle regulation, apoptosis induction, and metastasis inhibition. A proposed mechanism for these processes involves melatonin’s inhibition of NF-kB and activation of MAPKs which suppresses cell growth and motility in addition to promoting apoptosis through an accumulation of oncosuppressors such as p53, p27kip1, and p21 [137]. NF-κB inhibition and MAPKs activation also activates DNA repair complexes which help prevent the accumulation of DNA mutations caused by chemotherapeutic agents or ionizing radiations [138]. Melatonin has also been shown to inhibit cancer cell migration by reducing the expression of proteins involved in tumor cell progression and metastasization [137, 139,140,141]. An in vitro and in vivo study by Borin et al. found that melatonin decreased the expression of the Rho-associated kinase protein (ROCK-1), a protein associated with tumor growth and metastasis [141, 142]. Results from an in vitro study by Zhou et al. suggest that melatonin increases the expression of occludin, a regulatory protein in tight junctions where decreased expression has been associated with cancer and increased tumor invasion [139, 143].
Angiogenesis
Tumors with high levels of vascular endothelial growth factor (VEGF), an endogenous pro-angiogenic protein, often grow faster and metastasize earlier resulting in poor prognosis [144, 145]. Melatonin has been found to have direct effects on angiogenesis, through inhibitory actions on VEGF as well as indirect anti-angiogenic effects through the suppression of tumor growth factors and the neutralization of ROS which normally stimulate angiogenesis in tumors [146,29]. When primary uveal melanoma is diagnosed in patients, the cancer could be considered to already be in an advanced stage. Melatonin pocesses a broad spectrum of qualities desired in an adjuvant treatment such as general oncostatic, immunoenhancing, anti-oxidative, and anti-angiogenic properties while also possessing anti-metastatic properties relevant to more advanced cancer stages. Furthermore, the incidence of uveal melanoma peaks in patients over 60 years, which coincides with an age-related reduction in endogenous melatonin production [56, 59]. Thus, the herein reviewed anti-cancer effects of melatonin supplementation could potientially be greater in this age group than in younger populations.
Most human studies have provided participants with doses ranging from 10 to 40 mg, including the studies analysed in the systematic review by Mills et al [63]. Twenty milligram at night has been selected for no less than 13 randomized cancer trials [119, 120, 167, 168, 170,171,172,173,174,175,176,177,178]. This dose range may therefore be a good starting point for a randomized trial with adjuvant melatonin for patients with uveal melanoma.
Discussion
In this review, we have explored the rationale for using melatonin as an adjuvant treatment for non-metastatic uveal melanoma. This cancer is characterized by a relatively high proportion of patients develo** metastatic disease even when the eye containing the primary tumor is immediately enucleated, with up to 50% of patients develo** metastases within 25 years [22, 179]. This trend has been attributed to the spreading of clusters of dormant tumor cells called micrometastases to distant organs, notably the liver. By the time these micrometastases grow large enough to be detected, prognosis is fatal with a median patient survival time of less than 1 year [180]. Consequently, micrometastases and dormancy are central to the progression of uveal melanoma. The concept of dormancy here can be described as a phenomenon where malignant cells enter the G0 phase of the cell cycle and experience a state of temporary mitotic arrest. These so called micrometastases, however, maintain the potential to reactivate after several years or even decades and begin to grow, as seen in uveal melanoma [181]. It is therefore vital in the treatment of uveal melanoma to inhibit the development and migration of these micrometastases as soon as possible upon detection of the primary tumor.
Melatonin has been studied for several decades and has been shown to suppress the growth of cancer cells by up to 50% in vitro [159] and melatonin analogues have been found to suppress the growth of cancer cells by up to 90% in vivo [163]. Despite the limited number of studies investigating uveal melanoma in humans, a positive response to treatment with melatonin on cancer progression and disease stability has been observed in other types of cancer at various doses [165]. Melatonin has also been found to increase the efficiency of other standard cancer treatments such as chemotherapy and radiation in addition to possessing oncostatic properties itself [123,124,125,126,127,128]. As discussed, melatonin enhances our immune system, stimulates apoptosis in cancerous cells, inhibits angiogenesis, and regulates estrogen receptor expression. Therefore, it may be regarded as a suitable candidate for patients with uveal melanoma, especially in cohorts with non-metastatic uveal melanoma but high risk of later metastatic progression [179].
In previous studies, treatment with melatonin has been associated with a higher survival rate, longer mean time to progression, and a better response cytotoxic chemotherapy with less drug-related toxicity [117,118,119,120]. In uveal melanoma, early micrometastasis migrate via the blood stream to other organs and are usually detected long time after the primary tumour diagnosis [23, 25, 27, 28]. In this sense, uveal melanoma might therefore be regarded as an advanced systematic disease at onset, for which adjuvant treatment regimes are undoubtedly needed. Approximately 50% of patients with uveal melanoma who develop metastases, do so within 5 years from diagnosis, therefore a clinical trial investigating the impact of melatonin as an adjuvant treatment would require continuous supplementation over an extended period of time [179]. Such a project is motivated given the few reported side effects of melatonin, which include mild symptoms such as light diarrhea, drowsiness, fatigue, and itching without any significant effects on sexual health, sleep or mental status [116]. In summary, the characteristics of uveal melanoma taken together with the oncostatic properties of melatonin, support the importance of conducting a clinical trial with melatonin as an adjuvant treatment in newly diagnosed patients.
Limitations
There are limitations to this review which should be taken into consideration. First, no studies were found which evaluated the effects of melatonin at high doses for a long period of time in humans. Second, while studies were found which observed anti-metastatic properties of melatonin in vitro and in animal subjects, no studies were found which specifically investigated metastasis prevention in human subjects. Third, this systematic review relies on a limited number of databases where few studies focusing specifically on uveal melanoma were found.
With regards to potential limitations in the use of melatonin as an adjuvant treatment, one might argue that there isn’t enough regulation at present as melatonin is widely accessible and in some countries such as the United States can be bought at grocery stores without a prescription. This may pose challenges for controlling the use of the molecule in patients. While this issue may have to be addressed in the future, melatonin has been found to be a particularly safe supplement with few side effects which are relatively mild.
Conclusions
Uveal melanoma is a rare but serious form of cancer with a high rate of metastasis which can appear years to decades after the primary tumor is identified and removed. Results from both animal and human studies suggest that melatonin has the potential to suppress the growth of malignant cells and inhibit the development of metastasis in numerous types of cancers. This may be particularly beneficial in uveal melanoma where protection from the development of micrometastasis is vital immediately upon diagnosis as the prognosis diminishes significantly with the detection of metastases. Animal and cell line studies suggest that melatonin may have these oncostatic effects on uveal melanoma, however, no clinical trial specific to uveal melanoma has been performed on human patients. Future studies investigating the impact of long-term melatonin supplementation in patients diagnosed with uveal melanoma is the next step in this investigation.
Availability of data and materials
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
References
Shields JA, Shields CL, Donoso LA. Management of posterior uveal melanoma. Surv Ophthalmol. 1991;36(3):161–95.
Rantala ES, Hernberg M, Kivelä TT. Overall survival after treatment for metastatic uveal melanoma: a systematic review and meta-analysis. Melanoma Res. 2019;29(6):561–8.
Khoja L, Atenafu EG, Suciu S, Leyvraz S, Sato T, Marshall E, et al. Meta-analysis in metastatic uveal melanoma to determine progression free and overall survival benchmarks: an international rare cancers initiative (IRCI) ocular melanoma study. Ann Oncol. 2019;30(8):1370–80.
Küsters-Vandevelde HVN, Küsters B, Van Engen-van Grunsven ACH, Groenen PJTA, Wesseling P, Blokx WAM. Primary melanocytic tumors of the central nervous system: a review with focus on molecular aspects. Brain Pathol. 2015;25(2):209–26.
Singh AD, Shields CL, Shields JA, Sato T. Uveal melanoma in young patients. Arch Ophthalmol (Chicago, Ill : 1960). 2000;118(7):918–23.
Kaliki S, Shields CL. Uveal melanoma: relatively rare but deadly cancer. Eye. 2017;31(2):241–57.
Stålhammar G, See TR, Filì M, Seregard S. No gender differences in long-term survival after brachytherapy of 1,541 patients with uveal melanoma. Ocular Oncol Pathol. 2019;5(6):432–9.
Hu D-N, Yu G-P, McCormick SA, Schneider S, Finger PT. Population-based incidence of uveal melanoma in various races and ethnic groups. Am J Ophthalmol. 2005;140(4):612.e1–8.
Singh AD, Turell ME, Topham AK. Uveal melanoma: trends in incidence, treatment, and survival. Ophthalmology. 2011;118(9):1881–5.
Virgili G, Gatta G, Ciccolallo L, Capocaccia R, Biggeri A, Crocetti E, et al. Incidence of uveal melanoma in Europe. Ophthalmology. 2007;114(12):2309–2315.e2.
Aronow ME, Topham AK, Singh AD. Uveal melanoma: 5-year update on incidence, treatment, and survival (SEER 1973-2013). Ocular Oncol Pathol. 2018;4(3):145–51.
Stålhammar G, Herrspiegel C. Long-term relative survival in uveal melanoma: a systematic review and meta-analysis. Commun Med. 2022;2(18).
Jager MJ, Shields CL, Cebulla CM, Abdel-Rahman MH, Grossniklaus HE, Stern M-H, et al. Uveal melanoma. Nat Rev Dis Primers. 2020;6(1):24.
Nayman T, Bostan C, Logan P, Burnier MN. Uveal melanoma risk factors: a systematic review of meta-analyses. Curr Eye Res. 2017;42(8):1085–93.
Wakamatsu K, Hu D-N, McCormick SA, Ito S. Characterization of melanin in human iridal and choroidal melanocytes from eyes with various colored irides. Pigment Cell Melanoma Res. 2007;21(1):97–105.
Mäkitie T, Summanen P, Tarkkanen A, Kivelä T. Tumor-infiltrating macrophages (CD68(+) cells) and prognosis in malignant uveal melanoma. Invest Ophthalmol Vis Sci. 2001;42(7):1414–21.
Horsman DE, Sroka H, Rootman J, White VA. Monosomy 3 and isochromosome 8q in a uveal melanoma. Cancer Genet Cytogenet. 1990;45(2):249–53.
Harbour JW, Onken MD, Roberson EDO, Duan S, Cao L, Worley LA, et al. Frequent mutation of BAP1 in metastasizing uveal melanomas. Science. 2010;330(6009):1410–3.
Yavuzyigitoglu S, Koopmans AE, Verdijk RM, Vaarwater J, Eussen B, van Bodegom A, et al. Uveal melanomas with SF3B1 mutations. Ophthalmology. 2016;123(5):1118–28.
Ewens KG, Kanetsky PA, Richards-Yutz J, Purrazzella J, Shields CL, Ganguly T, et al. Chromosome 3 status combined with BAP1 and EIF1AX mutation profiles are associated with metastasis in uveal melanoma. Investig Opthalmol Visual Sci. 2014;55(8):5160.
Vader MJC, Madigan MC, Versluis M, Suleiman HM, Gezgin G, Gruis NA, et al. GNAQ and GNA11 mutations and downstream YAP activation in choroidal nevi. Br J Cancer. 2017;117(6):884–7.
Kujala E, Mäkitie T, Kivela T. Very long-term prognosis of patients with malignant uveal melanoma. Investig Opthalmol Visual Sci. 2003;44(11):4651.
Eagle RC. The COMS randomized trial of iodine 125 brachytherapy for choroidal melanoma. Arch Ophthalmol. 2006;124(12):1684.
Melia BM, Abramson DH, Albert DM, Boldt HC, Earle JD, Hanson WF, Montague P, Moy CS, Schachat AP, Simpson ER, Straatsma BR, Vine AK, Weingeist TA. Collaborative ocular melanoma study (COMS) randomized trial of I-125 brachytherapy for medium choroidal melanoma I. visual acuity after 3 years COMS report no. 16. Ophthalmology. 2001;108(2):348–66.
Eide N, Hoifødt HK, Nesland JM, Faye RS, Qvale GA, Faber RT, et al. Disseminated tumour cells in bone marrow of patients with uveal melanoma. Acta Ophthalmol. 2013;91(4):343–8.
Yu G, **g Y, Kou X, Ye F, Gao L, Fan Q, et al. Hepatic stellate cells secreted hepatocyte growth factor contributes to the chemoresistance of hepatocellular carcinoma. PLoS One. 2013;8(9):e73312.
Singh AD. Uveal melanoma: implications of tumor doubling time. Ophthalmology. 2001;108(5):829–30.
Uner OE, See TRO, Szalai E, Grossniklaus HE, Stålhammar G. Author correction: estimation of the timing of BAP1 mutation in uveal melanoma progression. Sci Rep. 2021;11(1):17036.
Grossniklaus HE. Understanding uveal melanoma metastasis to the liver: the Zimmerman effect and the Zimmerman hypothesis. Ophthalmology. 2019;126(4):483–7.
Kubatka P, Zubor P, Busselberg D, Kwon TK, Adamek M, Petrovic D, et al. Melatonin and breast cancer: evidences from preclinical and human studies. Crit Rev Oncol Hematol. 2018;122:133–43.
Anisimov VN, Popovich IG, Shtylik AV, Zabezhinski MA, Ben-Huh H, Gurevich P, et al. Melatonin and colon carcinogenesis. Exp Toxicol Pathol. 2000;52(1):71–6.
Wang S, Tai H, Tang C, Lin L, Lin T, Chang A, et al. Melatonin impedes prostate cancer metastasis by suppressing MMP-13 expression. J Cell Physiol. 2021;236(5):3979–90.
Fernández-Palanca P, Méndez-Blanco C, Fondevila F, Tuñón MJ, Reiter RJ, Mauriz JL, et al. Melatonin as an antitumor agent against liver cancer: an updated systematic review. Antioxidants. 2021;10(1):103.
Galien. Galen on the usefulness of the parts of the body: De Usu Partium; 1968.
McCord CP, Allen FP. Evidences associating pineal gland function with alterations in pigmentation. J Exp Zool. 1917;23(1):207–24.
Lerner AB, Case JD, Takahashi Y, Lee TH, Mori W. Isolation of melatonin, the pineal gland factor that lightens melanocytes. J Am Chem Soc. 1958;80(10):2587.
Slominski A, Fischer TW, Zmijewski MA, Wortsman J, Semak I, Zbytek B, et al. On the role of melatonin in skin physiology and pathology. Endocrine. 2005;27(2):137–48.
Bubenik GA. Gastrointestinal melatonin: localization, function, and clinical relevance. Dig Dis Sci. 2002;47(10):2336–48.
Cahill GM, Besharse JC. Circadian clock functions localized in xenopus retinal photoreceptors. Neuron. 1993;10(4):573–7.
Conti A, Conconi S, Hertens E, Skwarlo-Sonta K, Markowska M, Maestroni GJM. Evidence for melatonin synthesis in mouse and human bone marrow cells. J Pineal Res. 2000;28(4):193–202.
Tan D, Xu B, Zhou X, Reiter R. Pineal calcification, melatonin production, aging, associated health consequences and rejuvenation of the pineal gland. Molecules. 2018;23(2):301.
Tan D-X, RusselJ R. Mitochondria: the birth place, battle ground and the site of melatonin metabolism in cells. Melatonin Res. 2019;2(1):44–66.
Axelrod J, Weissbach H. Enzymatic O-methylation of N-Acetylserotonin to melatonin. Science. 1960;131(3409):1312.
Weissbach H, Redfield BG, Axelrod J. Biosynthesis of melatonin: enzymic conversion of serotonin to N-acetylserotonin. Biochim Biophys Acta. 1960;43:352–3.
Tricoire H, Møller M, Chemineau P, Malpaux B. Origin of cerebrospinal fluid melatonin and possible function in the integration of photoperiod. Reprod (Cambridge, England) Supplement. 2003;61:311–21.
Zhao D, Yu Y, Shen Y, Liu Q, Zhao Z, Sharma R, et al. Melatonin synthesis and function: evolutionary history in animals and plants. Front Endocrinol. 2019;10.
Maestroni GJM. The immunoneuroendocrine role of melatonin. J Pineal Res. 1993;14(1):1–10.
Claustrat B, Leston J. Melatonin: physiological effects in humans. Neurochirurgie. 2015;61(2–3):77–84.
Tan D-X, Manchester LC, Hardeland R, Lopez-Burillo S, Mayo JC, Sainz RM, et al. Melatonin: a hormone, a tissue factor, an autocoid, a paracoid, and an antioxidant vitamin. J Pineal Res. 2003;34(1):75–8.
Tan D, Chen LD, Poeggeler B, Manchester LC, Reiter RJ. Melatonin: a potent endogenous hydroxyl radical scavenger. Endocr J. 1993;1:57–60.
Tan D-X, Manchester LC, Reiter RJ, Qi W-B, Karbownik M, Calvo JR. Significance of melatonin in antioxidative defense system: reactions and products. Neurosignals. 2000;9(3–4):137–59.
Nelson RJ, Chbeir S. Dark matters: effects of light at night on metabolism. Proc Nutr Soc. 2018;77(3):223–9.
Hattar S, Liao H-W, Takao M, Berson DM, Yau K-W. Melanopsin-containing retinal ganglion cells: architecture, projections, and intrinsic photosensitivity. Science. 2002;295(5557):1065–70.
Borjigin J, Samantha Zhang L, Calinescu A-A. Circadian regulation of pineal gland rhythmicity. Mol Cell Endocrinol. 2012;349(1):13–9.
Brzezinski A. Melatonin in humans. N Engl J Med. 1997;336(3):186–95.
Waldhauser F, Frisch H, Waldhauser M, Weiszenbacher G, Zeitlhuber U, RichardJ W. Fall in nocturnal serum melatonin during prepuberty and pubescense. Lancet. 1984;323(8373):362–5.
Rossignol D, Frye R. Melatonin in autism spectrum disorders: a systematic review and meta-analysis. Dev Med Child Neurol. 2011;53(9):783–92.
Sadat-Ali M, Al-Habdan I, Al-Othman A. Adolescent idiopathic scoliosis. Is low melatonin a cause? Joint Bone Spine. 2000;67(1):62–4.
Zhou J-N, Liu R-Y, Kamphorst W, Hofman MA, Swaab DF. Early neuropathological Alzheimer’s changes in aged individuals are accompanied by decreased cerebrospinal fluid melatonin levels. J Pineal Res. 2003;35(2):125–30.
Brown RP, Kocsis JH, Caroff S, Amsterdam J, Winokur A, Stokes P, et al. Depressed mood and reality disturbance correlate with decreased nocturnal melatonin in depressed patients. Acta Psychiatr Scand. 1987;76(3):272–5.
Rosen R, Hu D-N, Perez V, Tai K, Yu G-P, Chen M, et al. Urinary 6-sulfatoxymelatonin level in age-related macular degeneration patients. Mol Vis. 2009;15:1673–9.
Tordjman S, Chokron S, Delorme R, Charrier A, Bellissant E, Jaafari N, et al. Melatonin: pharmacology, functions and therapeutic benefits. Curr Neuropharmacol. 2017;15(3):434–43.
Mills E, Wu P, Seely D, Guyatt G. Melatonin in the treatment of cancer: a systematic review of randomized controlled trials and meta-analysis. J Pineal Res. 2005;39(4):360–6.
Andersen LPH, Werner MU, Rosenkilde MM, Harpsøe NG, Fuglsang H, Rosenberg J, et al. Pharmacokinetics of oral and intravenous melatonin in healthy volunteers. BMC Pharmacol Toxicol. 2016;17(1):8.
Zimmermann RC, McDougle CJ, Schumacher M, Olcese J, Heninger GR, Price LH. Urinary 6-hydroxymelatonin sulfate as a measure of melatonin secretion during acute tryptophan depletion. Psychoneuroendocrinology. 1993;18(8):567–78.
Cardinali DP, Golombek DA, Rosenstein RE, Cutrera RA, Esquifino AI. Melatonin site and mechanism of action: single or multiple? J Pineal Res. 1997;23(1):32–9.
Aust S, Thalhammer T, Humpeler S, Jager W, Klimpfinger M, Tucek G, et al. The melatonin receptor subtype MT1 is expressed in human gallbladder epithelia. J Pineal Res. 2004;36(1):43–8.
Fischer TW, Slominski A, Zmijewski MA, Reiter RJ, Paus R. Melatonin as a major skin protectant: from free radical scavenging to DNA damage repair. Exp Dermatol. 2008;17(9):713–30.
Dubocovich ML, Markowska M. Functional MT1 and MT2 melatonin receptors in mammals. Endocrine. 2005;27(2):101–10.
Pandi-Perumal SR, Trakht I, Srinivasan V, Spence DW, Maestroni GJM, Zisapel N, et al. Physiological effects of melatonin: role of melatonin receptors and signal transduction pathways. Prog Neurobiol. 2008;85(3):335–53.
Slominski A, Wortsman J, Tobin DJ. The cutaneous serotoninergic/melatoninergic system: securing a place under the sun. FASEB J. 2005;19(2):176–94.
Jockers R, Maurice P, Boutin JA, Delagrange P. Melatonin receptors, heterodimerization, signal transduction and binding sites: what’s new? Br J Pharmacol. 2008;154(6):1182–95.
Savaskan E, Ayoub MA, Ravid R, Angeloni D, Fraschini F, Meier F, et al. Reduced hippocampal MT2 melatonin receptor expression in Alzheimer’s disease. J Pineal Res. 2005;38(1):10–6.
Brunner P, Sözer-Topcular N, Jockers R, Ravid R, Angeloni D, Fraschini F, et al. Pineal and cortical melatonin receptors MT1 and MT2 are decreased in Alzheimer’s disease. Eur J Histochem. 2007;50(4):311–6.
Reppert SM, Godson C, Mahle CD, Weaver DR, Slaugenhaupt SA, Gusella JF. Molecular characterization of a second melatonin receptor expressed in human retina and brain: the Mel1b melatonin receptor. Proc Natl Acad Sci. 1995;92(19):8734–8.
Slaugenhaupt SA, Roca AL, Liebert CB, Altherr MR, Gusella JF, Reppert SM. Map** of the gene for the mel1a-melatonin receptor to human chromosome 4 (MTNR1A) and mouse chromosome 8 (Mtnr1a). Genomics. 1995;27(2):355–7.
Duncan MJ, Takahashi JS, Dubocovich ML. 2-[125I]iodomelatonin binding sites in hamster brain membranes: pharmacological characteristics and regional distribution*. Endocrinology. 1988;122(5):1825–33.
Zhao Q, Yang XL, Holtzclaw WD, Talalay P. Unexpected genetic and structural relationships of a long-forgotten flavoenzyme to NAD(P)H:quinone reductase (DT-diaphorase). Proc Natl Acad Sci. 1997;94(5):1669–74.
Nosjean O, Ferro M, Cogé F, Beauverger P, Henlin J-M, Lefoulon F, et al. Identification of the melatonin-binding SiteMT 3 as the Quinone reductase 2. J Biol Chem. 2000;275(40):31311–7.
Smirnov AN. Nuclear melatonin receptors. Biochemistry (Mosc). 2001;66(1):19–26.
Dai J, Ram PT, Yuan L, Spriggs LL, Hill SM. Transcriptional repression of RORα activity in human breast cancer cells by melatonin. Mol Cell Endocrinol. 2001;176(1–2):111–20.
Hevia D, González-Menéndez P, Quiros-González I, Miar A, Rodríguez-García A, Tan D-X, et al. Melatonin uptake through glucose transporters: a new target for melatonin inhibition of cancer. J Pineal Res. 2015;58(2):234–50.
Dubocovich ML. Melatonin receptors: role on sleep and circadian rhythm regulation. Sleep Med. 2007;8:34–42.
Liu C, Weaver DR, ** X, Shearman LP, Pieschl RL, Gribkoff VK, et al. Molecular dissection of two distinct actions of melatonin on the suprachiasmatic circadian clock. Neuron. 1997;19(1):91–102.
Gobbi G, Comai S. Differential function of melatonin MT1 and MT2 receptors in REM and NREM sleep. Front Endocrinol. 2019;10.
Niles LP, Armstrong KJ, Rincón Castro L, Dao CV, Sharma R, McMillan CR, et al. Neural stem cells express melatonin receptors and neurotrophic factors: colocalization of the MT 1 receptor with neuronal and glial markers. BMC Neurosci. 2004;5(1):41.
Luboshitzky R, Lavie P. Melatonin and sex hormone interrelationships - a review. J Pediatr Endocrinol Metab. 1999;12(3):355-62.
Crowley SJ, Acebo C, Carskadon MA. Human puberty: salivary melatonin profiles in constant conditions. Dev Psychobiol. 2012;54(4):468–73.
Imenshahidi M, Karimi G, Hosseinzadeh H. Effects of melatonin on cardiovascular risk factors and metabolic syndrome: a comprehensive review. Naunyn Schmiedebergs Arch Pharmacol. 2020;393(4):521–36.
Celinski K, Konturek PC, Slomka M, Cichoz-Lach H, Brzozowski T, Konturek SJ. Effects of treatment with melatonin and tryptophan on liver enzymes, parameters of fat metabolism and plasma levels of cytokines in patients with non-alcoholic fatty liver disease--14 months follow up. J Physiol Pharmacol. 2014;65:75–82.
Scheer FAJL, van Montfrans GA, van Someren EJW, Mairuhu G, Buijs RM. Daily nighttime melatonin reduces blood pressure in male patients with essential hypertension. Hypertension. 2004;43(2):192–7.
Mansouri A, Demeilliers C, Amsellem S, Pessayre D, Fromenty B. Acute ethanol administration oxidatively damages and depletes mitochondrial dna in mouse liver, brain, heart, and skeletal muscles: protective effects of antioxidants. J Pharmacol Exp Ther. 2001;298(2):737–43.
Tan D, Reiter R, Manchester L, Yan M, El-Sawi M, Sainz R, et al. Chemical and physical properties and potential mechanisms: melatonin as a broad spectrum antioxidant and free radical scavenger. Curr Top Med Chem. 2002;2(2):181–97.
Reiter RJ, Calvo JR, Karbownik M, Qi W, Tan DX. Melatonin and its relation to the immune system and inflammation. Ann N Y Acad Sci. 2006;917(1):376–86.
Srinivasan V, Pandi-Perumal SR, Maestroni GJM, Esquifino AI, Hardeland R, Cardinali DP. Role of melatonin in neurodegenerative diseases. Neurotox Res. 2005;7(4):293–318.
Miller SC, Pandi PSR, Esquifino AI, Cardinali DP, Maestroni GJM. The role of melatonin in immuno-enhancement: potential application in cancer. Int J Exp Pathol. 2006;87(2):81–7.
Sompol P, Liu X, Baba K, Paul KN, Tosini G, Iuvone PM, et al. N-acetylserotonin promotes hippocampal neuroprogenitor cell proliferation in sleep-deprived mice. Proc Natl Acad Sci. 2011;108(21):8844–9.
Buchanan KL, Yellon SM. Delayed puberty in the male Djungarian hamster: effect of short photoperiod or melatonin treatment on the GnRH neuronal system. Neuroendocrinology. 1991;54(2):96–102.
Boafo A, Greenham S, Alenezi S, Robillard R, Pajer K, Tavakoli P, et al. Could long-term administration of melatonin to prepubertal children affect timing of puberty? A clinician’s perspective. Nat Sci Sleep. 2019;11:1–10.
Simko F, Pechanova O, Repova Bednarova K, Krajcirovicova K, Celec P, Kamodyova N, et al. Hypertension and cardiovascular remodelling in rats exposed to continuous light: protection by ACE-inhibition and melatonin. Mediators Inflamm. 2014;2014:1–10.
Wessely A, Steeb T, Erdmann M, Heinzerling L, Vera J, Schlaak M, et al. The role of immune checkpoint blockade in uveal melanoma. Int J Mol Sci. 2020;21(3):879.
Stålhammar G, Seregard S, Grossniklaus HE. Expression of immune checkpoint receptors Indoleamine 2,3-dioxygenase and T cell Ig and ITIM domain in metastatic versus nonmetastatic choroidal melanoma. Cancer Med. 2019;8:2784–92.
Javed A, Milhem M. Role of natural killer cells in uveal melanoma. Cancers. 2020;12(12):3694.
Reiter RJ, Rosales-Corral S, Tan DX, Jou MJ, Galano A, Xu B. Melatonin as a mitochondria-targeted antioxidant: one of evolution’s best ideas. Cell Mol Life Sci. 2017;74(21):3863–81.
Mediavilla M, Sanchez-Barcelo E, Tan D, Manchester L, Reiter R. Basic mechanisms involved in the anti-cancer effects of melatonin. Curr Med Chem. 2010;17(36):4462–81.
Qi X, Wang J. Melatonin improves mitochondrial biogenesis through the AMPK/PGC1α pathway to attenuate ischemia/reperfusion-induced myocardial damage. Aging. 2020;12(8):7299–312.
Martin M, Macıas M, León J, Escames G, Khaldy H, Acuña-Castroviejo D. Melatonin increases the activity of the oxidative phosphorylation enzymes and the production of ATP in rat brain and liver mitochondria. Int J Biochem Cell Biol. 2002;34(4):348–57.
Yamamoto H, Tang H. Preventive effect of melatonin against cyanide-induced seizures and lipid peroxidation in mice. Neurosci Lett. 1996;207(2):89–92.
Owino S, Contreras-Alcantara S, Baba K, Tosini G. Melatonin signaling controls the daily rhythm in blood glucose levels independent of peripheral clocks. PLoS One. 2016;11(1):e0148214.
Peschke E, Mühlbauer E, Mußhoff U, Csernus VJ, Chankiewitz E, Peschke D. Receptor (MT1) mediated influence of melatonin on cAMP concentration and insulin secretion of rat insulinoma cells INS-1. J Pineal Res. 2002;33(2):63–71.
Stumpf I, Mühlbauer E, Peschke E. Involvement of the cGMP pathway in mediating the insulin-inhibitory effect of melatonin in pancreatic β-cells. J Pineal Res. 2008;45(3):318–27.
Bähr I, Mühlbauer E, Albrecht E, Peschke E. Evidence of the receptor-mediated influence of melatonin on pancreatic glucagon secretion via the Gαq protein-coupled and PI3K signaling pathways. J Pineal Res. 2012;53(4):390–8.
la Fleur SE, Kalsbeek A, Wortel J, van der Vliet J, Buijs RM. Role for the pineal and melatonin in glucose homeostasis: pinealectomy increases night-time glucose concentrations. J Neuroendocrinol. 2001;13(12):1025–32.
Mühlbauer E, Gross E, Labucay K, Wolgast S, Peschke E. Loss of melatonin signalling and its impact on circadian rhythms in mouse organs regulating blood glucose. Eur J Pharmacol. 2009;606(1–3):61–71.
Lissoni P, Barni S, Ardizzoia A, Tancini G, Conti A, Maestroni G. A randomized study with the pineal hormone melatonin versus supportive care alone in patients with brain metastases due to solid neoplasms. Cancer. 1994;73(3):699–701.
Robinson WA, Dreiling L, Gonzalez R, Balmer C. Treatment of human metastatic malignant melanoma with high dose oral melatonin. In: The pineal gland and its hormones. Boston: Springer US; 1995. p. 219–25.
Lissoni P, Barni S, Mandalà M, Ardizzoia A, Paolorossi F, Vaghi M, et al. Decreased toxicity and increased efficacy of cancer chemotherapy using the pineal hormone melatonin in metastatic solid tumour patients with poor clinical status. Eur J Cancer. 1999;35(12):1688–92.
Lissoni P, Barni S, Fossati V, Ardizzoia A, Cazzaniga M, Tancini G, et al. A randomized study of neuroimmunotherapy with low-dose subcutaneous interleukin-2 plus melatonin compared to supportive care alone in patients with untreatable metastatic solid tumour. Support Care Cancer. 1995;3(3):194–7.
Lissoni P, Meregalli S, Nosetto L, Barni S, Tancini G, Fossati V, et al. Increased survival time in brain glioblastomas by a radioneuroendocrine strategy with radiotherapy plus melatonin compared to radiotherapy alone. Oncology. 1996;53(1):43–6.
Lissoni P, Barni S, Meregalli S, Fossati V, Cazzaniga M, Esposti D, et al. Modulation of cancer endocrine therapy by melatonin: a phase II study of tamoxifen plus melatonin in metastatic breast cancer patients progressing under tamoxifen alone. Br J Cancer. 1995;71(4):854–6.
Kanishi Y, Kobayashi Y, Noda S, Ishizuka B, Saito K. Differential growth inhibitory effect of melatonin on two endometrial cancer cell lines. J Pineal Res. 2000;28(4):227–33.
Petranka J, Baldwin W, Biermann J, Jayadev S, Barrett JC, Murphy E. The oncostatic action of melatonin in an ovarian carcinoma cell line. J Pineal Res. 1999;26(3):129–36.
Najafi M, Salehi E, Farhood B, Nashtaei MS, Hashemi Goradel N, Khanlarkhani N, et al. Adjuvant chemotherapy with melatonin for targeting human cancers: a review. J Cell Physiol. 2019;234(3):2356–72.
Najafi M, Shirazi A, Motevaseli E, Geraily G, Norouzi F, Heidari M, et al. The melatonin immunomodulatory actions in radiotherapy. Biophys Rev. 2017;9(2):139–48.
Wang Y, ** B, Ai F, Duan C, Lu Y, Dong T, et al. The efficacy and safety of melatonin in concurrent chemotherapy or radiotherapy for solid tumors: a meta-analysis of randomized controlled trials. Cancer Chemother Pharmacol. 2012;69(5):1213–20.
Shen Y-Q, Guerra-Librero A, Fernandez-Gil BI, Florido J, García-López S, Martinez-Ruiz L, et al. Combination of melatonin and rapamycin for head and neck cancer therapy: suppression of AKT/mTOR pathway activation, and activation of mitophagy and apoptosis via mitochondrial function regulation. J Pineal Res. 2018;64(3):e12461.
Lissoni P. Biochemotherapy with standard chemotherapies plus the pineal hormone melatonin in the treatment of advanced solid neoplasms. Pathol Biol. 2007;55(3–4):201–4.
Alonso-González C, Menéndez-Menéndez J, González-González A, González A, Cos S, Martínez-Campa C. Melatonin enhances the apoptotic effects and modulates the changes in gene expression induced by docetaxel in MCF-7 human breast cancer cells. Int J Oncol. 2018;52(2):560–70.
Reiter R, Rosales-Corral S, Tan D-X, Acuna-Castroviejo D, Qin L, Yang S-F, et al. Melatonin, a full service anti-cancer agent: inhibition of initiation, progression and metastasis. Int J Mol Sci. 2017;18(4):843.
Casado-Zapico S, Rodriguez-Blanco J, GarcÃa-Santos G, MartÃn V, Sánchez‐Sánchez AM, Antolín I, et al. Synergistic antitumor effect of melatonin with several chemotherapeutic drugs on human Ewing sarcoma cancer cells: potentiation of the extrinsic apoptotic pathway. J Pineal Res. 2010;48(1):72–80.
Menashe J. Managing and avoiding bortezomib toxicity. Commun Oncol. 2007;4(8):480–4.
Koşar PA, Nazıroğlu M, Övey İS, Çiğ B. Synergic effects of doxorubicin and melatonin on apoptosis and mitochondrial oxidative stress in MCF-7 breast cancer cells: involvement of TRPV1 channels. J Membr Biol. 2016;249(1–2):129–40.
Kim J-H, Jeong S-J, Kim B, Yun S-M, Choi DY, Kim S-H. Melatonin synergistically enhances cisplatin-induced apoptosis via the dephosphorylation of ERK/p90 ribosomal S6 kinase/heat shock protein 27 in SK-OV-3 cells. J Pineal Res. 2012;52(2):244–52.
Slominski A, Pruski D. Melatonin inhibits proliferation and melanogenesis in rodent melanoma cells. Exp Cell Res. 1993;206(2):189–94.
Cos S, Garcia-Bolado A, Sánchez-Barceló EJ. Direct antiproliferative effects of melatonin on two metastatic cell sublines of mouse melanoma (B16BL6 and PG19). Melanoma Res. 2001;11(2):197–201.
Fischer T, Zmijewski M, Zbytek B, Sweatman T, Slominski R, Wortsman J, et al. Oncostatic effects of the indole melatonin and expression of its cytosolic and nuclear receptors in cultured human melanoma cell lines. Int J Oncol. 2006;29(3):665–72.
Sardo FL, Muti P, Blandino G, Strano S. Melatonin and hippo pathway: is there existing cross-talk? Int J Mol Sci. 2017;18(9):1913.
Santoro R, Marani M, Blandino G, Muti P, Strano S. Melatonin triggers p53Ser phosphorylation and prevents DNA damage accumulation. Oncogene. 2012;31(24):2931–42.
Zhou Q, Gui S, Zhou Q, Wang Y. Melatonin inhibits the migration of human lung adenocarcinoma A549 cell lines involving JNK/MAPK pathway. PLoS One. 2014;9(7).
Cos S, Fernández R, Güézmes A, Sánchez-Barceló EJ. Influence of melatonin on invasive and metastatic properties of MCF-7 human breast cancer cells. Cancer Res. 1998;58(19):4383–90.
Borin TF, Arbab AS, Gelaleti GB, Ferreira LC, Moschetta MG, Jardim-Perassi BV, et al. Melatonin decreases breast cancer metastasis by modulating rho-associated kinase protein-1 expression. J Pineal Res. 2016;60(1):3–15.
Liu S, Goldstein RH, Scepansky EM, Rosenblatt M. Inhibition of rho-associated kinase signaling prevents breast cancer metastasis to human bone. Cancer Res. 2009;69(22):8742–51.
Tobioka H, Isomura H, Kokai Y, Tokunaga Y, Yamaguchi J, Sawada N. Occludin expression decreases with the progression of human endometrial carcinoma. Hum Pathol. 2004;35(2):159–64.
Ferrara N, Houck K, Jakeman L, Leung DW. Molecular and biological properties of the vascular endothelial growth factor family of proteins. Endocr Rev. 1992;13(1):18–32.
Salven P, Mänpää H, Orpana A, Alitalo K, Joensuu H. Serum vascular endothelial growth factor is often elevated in disseminated cancer. Clin Cancer Res. 1997;3(5):647–51.
Fandrey J, Genius J. Reactive oxygen species as regulators of oxygen dependent gene expression. Adv Exp Med Biol. 2000;475:153–9.
Dai M, Cui P, Yu M, Han J, Li H, **u R. Melatonin modulates the expression of VEGF and HIF-1α induced by CoCl 2 in cultured cancer cells. J Pineal Res. 2008;44(2):121–6.
Cheng J, Yang H, Gu C, Liu Y, Shao J, Zhu R, et al. Melatonin restricts the viability and angiogenesis of vascular endothelial cells by suppressing HIF-1α/ROS/VEGF. Int J Mol Med. 2019;43:945–55.
Wang R-X, Liu H, Xu L, Zhang H, Zhou R-X. Involvement of nuclear receptor RZR/RORγ in melatonin-induced HIF-1α inactivation in SGC-7901 human gastric cancer cells. Oncol Rep. 2015;34(5):2541–6.
Wang R-X, Liu H, Xu L, Zhang H, Zhou R-X. Melatonin downregulates nuclear receptor RZR/RORγ expression causing growth-inhibitory and anti-angiogenesis activity in human gastric cancer cells in vitro and in vivo. Oncol Lett. 2016;12(2):897–903.
Dong L, You S, Zhang Q, Osuka S, Devi NS, Kaluz S, et al. Arylsulfonamide 64B inhibits hypoxia/HIF-induced expression of c-met and CXCR4 and reduces primary tumor growth and metastasis of uveal melanoma. Clin Cancer Res. 2019;25(7):2206–18.
Schoenfield L, Janse S, Kline D, Aronow ME, Singh AD, Craven C, et al. Estrogen receptor is expressed in uveal melanoma: a potential target for therapy. Ocular Oncol Pathol. 2021;7(4):303–10.
Rato AG, Pedrero JG, Martínez MA, del Rio B, Lazo PS, Ramos S. Melatonin blocks the activation of estrogen receptor for DNA binding. FASEB J. 1999;13(8):857–68.
Molis TM, Spriggs LL, Hill SM. Modulation of estrogen receptor mRNA expression by melatonin in MCF-7 human breast cancer cells. Mol Endocrinol. 1994;8(12):1681–90.
Lawson NO, Wee BEF, Blask DE, Castles CG, Spriggs LL, Hill SM. Melatonin decreases estrogen receptor expression in the medial preoptic area of inbred (LSH/SsLak) Golden hamsters. Biol Reprod. 1992;47(6):1082–90.
Bouhoute A, Leclercq G. Modulation of estradiol and DNA binding to estrogen receptor upon association with calmodulin. Biochem Biophys Res Commun. 1995;208(2):748–55.
Castoria G, Migliaccio A, Nola E, Auricchio F. In vitro interaction of estradiol receptor with Ca2+ −calmodulin. Mol Endocrinol. 1988;2(2):167–74.
García Pedrero JM, del Rio B, Martínez-Campa C, Muramatsu M, Lazo PS, Ramos S. Calmodulin is a selective modulator of estrogen receptors. Mol Endocrinol. 2002;16(5):947–60.
Hu D-N, Roberts JE. Melatonin inhibits growth of cultured human uveal melanoma cells. Melanoma Res. 1997;7(1):27–31.
Hu DN, McCormick SA, Roberts JE. Effects of melatonin, its precursors and derivatives on the growth of cultured human uveal melanoma cells. Melanoma Res. 1998;8(3):205–10.
Roberts JE, Wiechmann AF, Hu D-N. Melatonin receptors in human uveal melanocytes and melanoma cells. J Pineal Res. 2002;28(3):165–71.
Alvarez-Artime A, Cernuda-Cernuda R, Francisco-Artime-Naveda CV, Gonzalez-Menendez P, Fernadez-Vega S, et al. Melatonin-induced cytoskeleton reorganization leads to inhibition of melanoma cancer cell proliferation. Int J Mol Sci. 2020;21(2):548.
Gatti G, Lucini V, Dugnani S, Calastretti A, Spadoni G, Bedini A, et al. Antiproliferative and pro-apoptotic activity of melatonin analogues on melanoma and breast cancer cells. Oncotarget. 2017;8(40):68338–53.
Narita T, Kudo H. Effect of melatonin on B16 melanoma growth in athymic mice. Cancer Res. 1985;45(9):4175–7.
Gonzalez R, Sanchez A, Ferguson JA, Balmer C, Daniel C, Cohn A, et al. Melatonin therapy of advanced human malignant melanoma. Melanoma Res. 1991;1(4):237–44.
Kane MA, Johnson A, Nash AE, Boose D, Mathai G, Balmer C, et al. Serum melatonin levels in melanoma patients after repeated oral administration. Melanoma Res. 1994;4(1):59–65.
Seely D, Legacy M, Auer RC, Fazekas A, Delic E, Anstee C, et al. Adjuvant melatonin for the prevention of recurrence and mortality following lung cancer resection (AMPLCaRe): a randomized placebo controlled clinical trial. EClinicalMedicine. 2021;33.
Hrushesky WJM, Lis CG, Levin RD, Grutsch JF, Birdsall T, Wood PA, et al. Daily evening melatonin prolongs survival among patients with advanced non-small-cell lung cancer. Biol Rhythm Res. 2021:1–15.
Uner OE, See TRO, Szalai E, Grossniklaus HE, Stålhammar G. Estimation of the timing of BAP1 mutation in uveal melanoma progression. Sci Rep. 2021;11(1):8923.
Kartini D, Taher A, Panigoro SS, Setiabudy R, Jusman SW, Haryana SM, et al. Effect of melatonin supplementation in combination with neoadjuvant chemotherapy to miR-210 and CD44 expression and clinical response improvement in locally advanced oral squamous cell carcinoma: a randomized controlled trial. J Egypt Natl Canc Inst. 2020;32(1):12.
Lissoni P, Chilelli M, Villa S, Cerizza L, Tancini G. Five years survival in metastatic non-small cell lung cancer patients treated with chemotherapy alone or chemotherapy and melatonin: a randomized trial. J Pineal Res. 2003;35(1):12–5.
Cerea G, Vaghi M, Ardizzoia A, Villa S, Bucovec R, Mengo S, et al. Biomodulation of cancer chemotherapy for metastatic colorectal cancer: a randomized study of weekly low-dose irinotecan alone versus irinotecan plus the oncostatic pineal hormone melatonin in metastatic colorectal cancer patients progressing on 5-fluorouracil-containing combinations. Anticancer Res. 2003;23(2C):1951–4.
Berk L, Berkey B, Rich T, Hrushesky W, Blask D, Gallagher M, et al. Randomized phase II trial of high-dose melatonin and radiation therapy for RPA class 2 patients with brain metastases (RTOG 0119). Int J Radiat Oncol Biol Phys. 2007;68(3):852–7.
Lissoni P, Barni S, Cattaneo G, Tancini G, Esposti G, Esposti D, et al. Clinical results with the pineal hormone melatonin in advanced cancer resistant to standard antitumor therapies. Oncology. 1991;48(6):448–50.
Lissoni P, Brivio O, Brivio F, Barni S, Tancini G, Crippa D, et al. Adjuvant therapy with the pineal hormone melatonin in patients with lymph node relapse due to malignant melanoma. J Pineal Res. 1996;21(4):239–42.
Lissoni P, Brivio F, Fumagalli L, Messina G, Vigoré L, Parolini D, et al. Neuroimmunomodulation in medical oncology: application of psychoneuroimmunology with subcutaneous low-dose IL-2 and the pineal hormone melatonin in patients with untreatable metastatic solid tumors. Anticancer Res. 2008;28(2B):1377–81.
Lissoni P, Bolis S, Brivio F, Fumagalli L. A phase II study of neuroimmunotherapy with subcutaneous low-dose IL-2 plus the pineal hormone melatonin in untreatable advanced hematologic malignancies. Anticancer Res. 2000;20(3B):2103–5.
Yan J-J, Shen F, Wang K, Wu M-C. Patients with advanced primary hepatocellular carcinoma treated by melatonin and transcatheter arterial chemoembolization: a prospective study. Hepatobiliary Pancreat Dis Int. 2002;1(2):183–6.
Stålhammar G. Forty-year prognosis after plaque brachytherapy of uveal melanoma. Sci Rep. 2020;10(1):11297.
Augsburger JJ, Corrêa ZM, Shaikh AH. Effectiveness of treatments for metastatic uveal melanoma. Am J Ophthalmol. 2009;148(1):119–27.
Nichols EE, Richmond A, Daniels AB. Micrometastatic dormancy in uveal melanoma: a comprehensive review of the evidence, mechanisms, and implications for future adjuvant therapies. Int Ophthalmol Clin. 2017;57(1):1–10.
Acknowledgements
Not applicable.
Funding
Open access funding provided by Karolinska Institute. Gustav Stålhammar is supported by The Royal Swedish Academy of Sciences (reference ME2019-0036); The Swedish Cancer Society (20 0798 Fk); The Swedish Society of Medicine (Cronqvists stiftelse, SLS 934014); The Swedish Eye Foundation (reference 2021-04-28); Karolinska Institutet (2019-01900, 2020-013333 and 2020-02517); Region Stockholm (20200356); The Crown Princess Margareta Foundation for the Visually Impaired (157); Carmen and Bertil Regnér Foundation (2020-00062). Pete Williams is supported by Karolinska Institutet in the form of a Board of Research Faculty Funded Career Position, by St. Erik Eye Hospital philanthropic donations, and by Vetenskapsrådet (2018-02124). The funding organizations had no role in the design or conduct of this research.
Author information
Authors and Affiliations
Contributions
Conceptualization, Gustav Stålhammar; methodology, Gustav Stålhammar; investigation, Anna Hagström and Ruba Kal Omar; writing – original draft preparation, Anna Hagström and Ruba Kal Omar; writing – review & editing, Anna Hagström, Ruba Kal Omar, Gustav Stålhammar and Pete A. Williams; supervision, Gustav Stålhammar and Pete A. Williams; project administration; Gustav Stålhammar. The author(s) read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
Gustav Stålhammar has received honoraria for consultancy and service on an advisory board (Santen SA). The other authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Hagström, A., Kal Omar, R., Williams, P.A. et al. The rationale for treating uveal melanoma with adjuvant melatonin: a review of the literature. BMC Cancer 22, 398 (2022). https://doi.org/10.1186/s12885-022-09464-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12885-022-09464-w