
Abstract
Background
The intensified global warming during grain filling deteriorated rice quality, in particular increasing the frequency of chalky grains which markedly impact market value. The formation of rice quality is a complex process influenced by multiple genes, proteins and physiological metabolic processes. Proteins responsive to stimulus can adjust the ability of plants to respond to unfavorable environments, which may be an important protein involved in the regulation of quality formation under elevated temperature. However, relatively few studies have hindered our further understanding of rice quality formation under elevated temperature.
Results
We conducted the actual field elevated temperature experiment and performed proteomic analysis of rice grains at the early stage of grain filling. Starting with the response to stimulus in GO annotation, 22 key proteins responsive to stimulus were identified in the regulation of grain filling and response to elevated temperature. Among the proteins responsive to stimulus, during grain filling, an increased abundance of signal transduction and other stress response proteins, a decreased abundance of reactive oxygen species-related proteins, and an increased accumulation of storage substance metabolism proteins consistently contributed to grain filling. However, the abundance of probable indole-3-acetic acid-amido synthetase GH3.4, probable indole-3-acetic acid-amido synthetase GH3.8 and CBL-interacting protein kinase 9 belonged to signal transduction were inhibited under elevated temperature. In the reactive oxygen species-related protein, elevated temperature increased the accumulation of cationic peroxidase SPC4 and persulfide dioxygenase ETHE1 homolog to maintain normal physiological homeostasis. The increased abundance of alpha-amylase isozyme 3E and seed allergy protein RA5 was related to the storage substance metabolism, which regulated starch and protein accumulation under elevated temperature.
Conclusion
Auxin synthesis and calcium signal associated with signal transduction, other stress responses, protein transport and modification, and reactive oxygen species-related proteins may be key proteins responsive to stimulus in response to elevated temperature. Alpha-amylase isozyme 3E and seed allergy protein RA5 may be the key proteins to regulate grain storage substance accumulation and further influence quality under elevated temperature. This study enriched the regulatory factors involved in the response to elevated temperature and provided a new idea for a better understanding of grain response to temperature.
Similar content being viewed by others
Background
The global mean surface temperature was predicted to increase by 1.0 °C to 5.7 °C by 2100 compared to 1850–1900 [1]. The temperature rise in China was faster than the global average temperature in the same period. In recent years, the occurrence of high temperature events in China remarkably increased due to the effect of climate change [2,3,4]. As one of the major rice-growing countries, China contributed more than 30% of global rice production [5]. However, with the increased frequency and magnitude of high temperature in the rice growing season, rice cultivation had encountered great challenges [6].
The temperature rise will impose stressful influences on different growth and developmental stages throughout the rice growing season [7]. However, elevated temperature has a greater effect on the reproductive stage, especially in the grain-filling stage [8]. From the biological point of view, grain filling refers that the zygote develops into an embryo through cell division and differentiation and the polar nucleus develops into endosperm by photosynthetic assimilation deposition and storage protein synthesis [9]. An et al. [21,22]. Increased temperature also reduced amylose content, changed amylopectin content, and increased protein accumulation to further change rice cooking and nutritional quality [23,24,25,26,27]. However, the molecular mechanism by which elevated temperature changes the physicochemical properties of rice storage substances and quality formation has not been elucidated. Hence, the lack of molecular mechanism pushes researchers to explore insight into the responses of rice grain to high temperature [24, 28, 43] found considerable proteins involved in redox homeostasis and carbohydrate biosynthetic pathways were important to rice grains in response to high temperature. Liao et al. [44] conducted a proteomic analysis with different heat-tolerant rice lines to elucidate the differential abundance proteins related to the Calvin cycle, the glycolytic pathway, the tricarboxylic acid cycle which regulated rice grain substance accumulation under high temperature. The identification of the proteins which are responsive to elevated temperature is vital to understanding the molecular mechanisms of temperature regulation effects. Numerous studies also inspired researchers that some stress-related proteins were pivotal in rice grain development, such as heat shock proteins (HSP), peroxidases and lipid transfer proteins [45,46,47]. Kumar et al. [48] revealed that 72% of the identified differential proteins belonged to defense and stress responses under elevated temperature by two-dimensional electrophoresis (2-DE) and MALDI-TOF/MS based proteomics approaches. Although several genes and proteins have been identified that were involved in the response to high temperature stress, much research remains to be done to fully understand the regulatory networks of stress-responsive proteins [24, 49, 50].
Since 2012, we continuously conducted actual field warming treatments by the Free-Air Temperature Enhancement (FATE) facility. In our previous study, we have found the changes in the physicochemical properties of starch, and the imbalance of storage protein may cause deteriorated rice quality [30, 31, 51,52,53]. A deeper understanding of the internal regulatory network is essential to explain the rice quality formation under warming. A number of studies have shown that stress proteins can adjust the ability of plants to respond to unfavorable environments. However, the lack of research content limits our understanding of stress proteins that regulate grain substance accumulation in rice under elevated temperature. Therefore, we performed proteomic analysis of rice grains at the early stage of grain filling under natural and elevated temperature. Our results identified some candidate proteins responsive to stimulus and revealed these proteins on the regulatory pathway for grain substance accumulation under elevated temperature, which enriches the protein regulatory network for grain quality formation under elevated temperature.
Results
Elevated temperature affected the grain filling and deteriorated rice quality
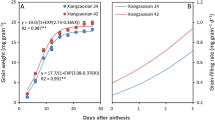
Figure 1 showed grain weight dynamics and grain filling rates of W3 at natural temperature and elevated temperature for two years. The effect of temperature on grain weight was consistent for two years. Elevated temperature increased grain weight before 25d after flowering. Subsequently, the up-regulated trend of grain weight disappeared and presented an opposite trend under elevated temperature. The effects of warming on the grain filling rate were consistent with the grain weight. Elevated temperature accelerated the grain filling in the early filling stage (3d to 12d after flowering). However, in the middle and late filling stages, insufficient filling stamina under elevated temperature was shown. Ultimately, the active grain-filling period was shortened under elevated temperature (Table S1). The effect of temperature on grain filling process ultimately reduced the seed setting rate and grain weight which led to a decrease in the theoretical yield (Table S2).

Effects of elevated temperature on grain filling. A, C Grain filling rate was in 2020 and 2021 under elevated temperature, respectively. B, D Grain weight was in 2020 and 2021, respectively. Note: CK Natural temperature, ET Elevated temperature
The effect of elevated temperature on rice quality was shown in Fig. 2. There was no significant influence on brown rice rate and milled rice rate under elevated temperature in 2020, and the head rice rate decreased significantly by 9.29%. However, in 2021, elevated temperature resulted in a decrease in milled rice rate at the significance level. Similarly, the head rice decreased significantly by 17.90% in 2021. The length of grain significantly reduced in 2020 and 2021, whereas the width did not change significantly in 2021, only a decreasing trend. However, the length/width was not affected under elevated temperature (Table S3). Chalkiness was the most sensitive appearance trait and changed significantly under elevated temperature. The two-year results consistently showed that increased temperature deteriorated chalky characteristics, especially the chalkiness, which increased by 527.36% and 596.89% in 2020 and 2021, respectively (Fig. 2).

Effects of elevated temperature on rice quality. A Milling quality; B Scan imagery on rice grain length and width; C Chalkiness characteristic; D The content of starch components and total protein. Note: CK: natural temperature; ET: elevated temperature. Bars are the stand deviation. Values followed by different letters are significantly different at 5% probability
The effect of temperature on filling ultimately also led to change in the two main storage substances, starch and storage protein. Elevated temperature had no significant effect on the total starch content, but significantly reduced the amylose content by 7.99% and 6.84% in 2020 and 2021, respectively. Elevated temperature significantly increased the content of amylopectin only in 2021. Elevated temperature had no significant influence on total starch, and led to an increase in total protein content, especially in 2020 which increased by 7.12%.
Under elevated temperature, the changes in Rapid viscosity analyzer (RVA) profile characteristics in 2020 and 2021 were consistent (Table 1). Elevated temperature had a positive effect on peak viscosity, breakdown viscosity and past temperature. Among them, compared with natural temperature, the breakdown viscosity increased by 49.56% and 72.87% in 2020 and 2021, respectively. Hot paste viscosity, final viscosity, setback viscosity and past time decreased significantly. Compared to 2020, the decline was even greater in 2021, especially the setback viscosity, which dropped by 363.27%.
The quality of the cooking and eating was measured by taste meter and hardness viscometer (Table 2). The appearance and taste of rice significantly decreased under elevated temperature, especially in 2020. Elevated temperature had a negative impact on rice viscosity, balance and elasticity. The overall score of rice under elevated temperature decreased by 4.63% and 3.25% in 2020 and 2021, respectively.
Proteins responsive to stimulus in co** with elevated temperature
We attempted to further investigate the effect of temperature on grain storage substance accumulation and quality formation by proteomics. Based on the DIA (Data Independent Acquisition) experiment, a total of 22,930 unique peptides and 6650 proteins were identified in grains at 3, 6, 9, 12 and 15d after flowering under natural temperature and elevated temperature (Fig. S1A, B). Fig. S1C indicated most protein coverage was < 30%. In addition, PCA (Principal Component Analysis) 1 and PCA2 accounted for 48.64% and 11.44% of the variability, which indicated good sample reproducibility for proteomics (Fig. S1D).
Figure 3A showed the identified differentially expressed proteins were mainly divided into molecular function, cellular components and biological processes by GO (Gene Ontology) analysis. Among them, the identified differential proteins were mostly involved in the biological processes, and more than 1000 proteins were enriched in response to stimulus, single-organism process, cellular process and metabolic process which belonged to biological processes. we selected 1119 proteins that belonged to responsive to stimulus for further analysis.

Changes of differential proteins under elevated temperature. A GO annotation statistics of differentially expressed proteins during grain substance accumulation and under elevated temperature; B Changes in the number of differential proteins under warming on different days after flowering; C Venn diagram analysis of differential proteins in response to elevated temperature on different days after flowering. Note: E3/C3 represents differentially responsive to stimulus proteins at 3d after flowering under elevated temperature. Other representations are similar
Figure 3B showed the number of differential proteins under elevated temperature (Fold change > = 2; P value < 0.05). At 9d after flowering, the highest number of differential proteins was found under elevated temperatures, including 11 up-regulated differential proteins and 34 down-regulated differential proteins. The number of total differential proteins was equal at 12d and 15d after flowering, but the number of down-regulated differential proteins at 15d after flowering were more than that at 12d. A Venn diagram analysis was conducted to determine the dynamics of differential proteins under elevated temperature at different days after flowering (Fig. 3C). We further analyzed the KEGG (Kyoto Encyclopedia of Genes and Genomes)-enriched bubble plot of differential proteins under elevated temperature (Fig. 4). In the KEGG-enriched bubble plots, differential proteins responsive to stimulus were more enriched at 12d and 15d after flowering, especially at 15d after flowering. Protein processing in endoplasmic reticulum (folding, classification and degradation), fructose and mannose metabolism (carbohydrate metabolism) were significantly enriched at 12d after flowering. At 15d after flowering, it was significantly enriched in fructose and mannose metabolism (carbohydrate metabolism), flavonoid biosynthesis (biosynthesis of other secondary metabolites), glycolysis/gluconeogenesis (carbohydrate metabolism), metabolic pathways and biosynthesis of secondary metabolites. Therefore, differential proteins responded to elevated temperature by different metabolic pathways at different days after flowering.

KEGG-enriched bubble plots of differential proteins under elevated temperature at different days after flowering. A 3d after flowering; B 6d after flowering; C 9d after flowering; D 12d after flowering; E 15d after flowering
The differential proteins under elevated temperature can be divided into seven types according to GO categories (Table S4). The differential proteins under elevated temperature were diversified, among which proteins involved in stress response, chemical stress and abiotic stress were the most, accounting for 26.51%, 17.87% and 17.58%, respectively.
Proteins responsive to stimulus during grain substance accumulation
After studying the proteins responsive to stimulus under elevated temperature, we further investigated the role of proteins in grain filling. Figure 5A showed the number of differential proteins which were at 6, 9, 12 and 15d after flowering compared with 3d after flowering (Fold change > = 2; P value < 0.05). The total differential proteins gradually increased as the grain substance accumulation progressed. There was no significant difference in the number of up-regulated differential proteins. However, the number of down-regulated differential proteins gradually increased with the progress of grain substance accumulation. Figure 5B indicated a total of 109 differential proteins which were significantly up- or down-regulated at 6, 9, 12 and 15d after flowering (Table S6). The 109 differential proteins were mainly involved in carbohydrate metabolism, amino acid metabolism, energy metabolism and lipid metabolism and other metabolic processes which were necessary for grain substance accumulation by KEGG metabolic pathway analysis (Fig. 5C).

The number, metabolic pathways and classification of differential proteins during grain substance accumulation. A The number of differential proteins during grain substance accumulation; B 109 proteins were consistently differentially expressed during grain substance accumulation by venn diagram analysis; C KEGG metabolic pathway analysis of 109 proteins that are consistently differentially expressed during grain substance accumulation; D Cluster analysis of expression patterns of 109 differential proteins. Red indicates the abundance of protein was up-regulated (the log2 relative expression level was more than 0), and green indicates the abundance of protein was down-regulated (the log2 relative expression level was less than 0). The down-regulated expression of differential proteins was classified as A class, and the up-regulated expression was classified as B class. Note: C6/C3 represents significantly differential proteins at 6d after flowering compared to 3d after flowering. Other representations are similar
To further clarify the role of differential proteins in grain filling, the expression patterns of 109 differential proteins were distinguished by hierarchical clustering analysis (Fig. 5D). With the grain substance accumulation, the down-regulated expression of differential proteins was classified as A class, and the up-regulated expression of differential proteins was classified as B class. The differential proteins with different expression patterns were further classified according to protein functions in Table 3. The function of differential proteins mainly participated in carbohydrate metabolism, other stress response and protein modification. In the class A expression pattern, the differential proteins were mainly involved in other stress response, protein modification and reactive oxygen species-related. Under the class B expression pattern, the differential proteins were mainly involved in carbohydrate metabolism, signal transduction, lipid metabolism and amino acid metabolism.
Key differential proteins responsive to stimulus were involved in both response to elevated temperature and grain substance accumulation
We screened for differential proteins which responded to both grain development (3-15d after flowering) and elevated temperature (Fold change > = 2; P-value < 0.05). Finally, 22 differential proteins were selected for further analysis (Fig. 6). The 22 proteins were divided into 5 categories according to functions: signal transduction, protein modification and transport, storage substance metabolism, reactive oxygen species-related and other stress responses (Table S7).

Expression of 22 important proteins which responded to both grain substance accumulation (3-15d after flowering) and elevated temperature (Fold change > = 2; P-value < 0.05). The 22 proteins were divided into 5 categories according to functions: signal transduction, protein modification and transport, storage substance metabolism, reactive oxygen species-related and other stress responses. a-d The protein functions belonged to signal transduction; e–h The protein functions belonged to protein transport and modification; i-p The protein functions belonged to storage substance metabolism; q-s The protein functions belonged to reactive oxygen species-related; t-v The protein functions belonged to other stress responses. Note: CK Natural temperature, ET Elevated temperature. Protein ID was obtained from NCBI
Probable indole-3-acetic acid-amide synthetase GH3.4 (1,002,267,580), probable indole-3-acetic amide synthetase GH3.8 (1,002,286,982), universal stress protein A-like protein (1,002,226,615) and CBL-interacting protein kinase 9 (1,002,252,959), which involved in signal transduction, gradually increased in protein abundance during grain substance accumulation. Warming decreased the abundance of the four proteins on different days after flowering, especially CBL-interacting protein kinase 9 (1,002,252,959), which decreased by 0.46 and 0.49-folds under elevated temperature at 6d and 15d after flowering, respectively.
Different from the expression pattern of differentially expressed proteins involved in signal transduction, the four proteins involved in protein modification and transport were all down-regulated as grain filling. Under elevated temperature, the abundance of basic 7S globulin (1,002,266,396) was up-regulated from 3d-15d after flowering, especially at 12d after flowering, in which the abundance was significantly up-regulated by 2.42 times. Unknown protein LOC4352441 (1,002,313,067), ATP-dependent DNA helicase 2 subunit KU80 isoform X1 (1,002,253,784) and deoxyuridine 5'-triphosphate nucleotide hydrolase (1,002,249,448) were down-regulated under elevated temperature.
A total of 8 proteins were involved in the storage substance metabolism, and 6 of these proteins were all continuously up-regulated with the grain substance accumulation. The abundance of aquaporin PIP2-4 (1,002,280,567) was significantly up-regulated by 3.34 times at 3d after flowering by elevated temperature. Elevated temperature significantly increased the abundance of Alpha-amylase isoenzyme 3E (1,002,292,698) by 3.02 times at 9d after flowering.
There were three proteins belonged to reactive oxygen species-related, of which the accumulation of superoxide dismutase [Fe] 1 (1,002,273,847) and persulfide dioxygenase ETHE1 homolog (1,002,230,966) was gradually down-regulated during grain substance accumulation. Under elevated temperature, the superoxide dismutase [Fe] 1 (1,002,273,847) was significantly down-regulated to 0.22 times of natural temperature at 15 d after flowering, while cationic peroxidase SPC4 (1,002,235,766) and persulfide dioxygenase ETHE1 homolog (1,002,230,966) were significantly increased by 2.15 and 2.20 times at 12d and 6d after flowering, respectively.
HEAT-STRESS-ASSOCIATED 32 (1,002,274,021), oil body-associated protein 1A (1,002,266,326) and copper transport protein ATX1 isoform X1 (1,002,287,567) were classified as other stress responses protein and gradually increased in relative protein abundance with grain substance accumulation. Elevated temperature resulted in a decreased accumulation of HEAT-STRESS-ASSOCIATED 32 (1,002,274,021) and copper transport protein ATX1 isoform X1 (1,002,287,567) at 3d and 15d after flowering, respectively. However, oil body-associated protein 1A (1,002,266,326) was 2.36-fold and 2.18-fold higher than that under natural temperature at 3d and 9d after flowering.
To verify the reliability of the proteome results, we performed a verification experiment by RT-PCR at the transcriptional level. 10 proteins were selected including the above five functional categories. Except for aquaporin PIP2-4 protein (LOC4343119), the other 9 proteins showed the same tendency in the relative expression level of RT-PCR and DIA proteomic level (Fig. S2).
Discussion
The important role of signal transduction and ROS related proteins in grain substance accumulation and response to elevated temperature
To withstand stresses, plants have formatted regulatory pathways and produce various proteins including protein kinases, molecular chaperones, ROS-scavenging proteins, etc. to confer tolerance to stresses [54]. The adverse environment firstly is sensed by plants which further triggers stress-specific signal transduction [55]. Plant hormones are important signal compounds that mediate plant responses to stress [56]. In our study, we identified several signal transduction proteins including probable indole-3-acetic acid-amido synthetase GH3.4 (1,002,267,580) and probable indole-3-acetic acid-amido synthetase GH3.8 (1,002,286,982) which were related to auxin synthesis during grain substance accumulation and response to elevated temperature (Fig. 7). The abundance of probable indole-3-acetic acid-amido synthetase GH3.8 was significantly accumulated at 6d-15d after flowering. However, elevated temperature suppressed the accumulation of probable indole-3-acetic acid-amido synthetase GH3.4 and probable indole-3-acetic acid-amido synthetase GH3.8 which decreased by 0.45-fold and 0.48-fold, respectively. The results showed that the up-regulation of auxin-related proteins may promote grain substance accumulation. Study has indicated auxin synthesis was necessary but not sufficient for heat tolerance [57]. However, elevated temperature inhibited the accumulation of auxin-related proteins. It may be our elevated temperatures are beyond the range that grain can actively regulate.

Heatmaps of differentially expressed proteins during grain substance accumulation. According to the functional classification, the important differential proteins were classified as signal transduction, protein transport, translation and modification, storage substance metabolism, reactive oxygen species-related and other stress responses (Fold change > = 2; P-value < 0. 05). Four consecutive squares from left to right indicates different days after flowering (6, 9, 12 and 15d after flowering).Red indicates the abundance of protein was low, and green indicates the abundance of protein was high
Protein kinases also play vital roles in signal transduction under stress conditions [58]. It is a central component in plant responses to environmental stresses by adding phosphate groups to control the concentration of ions or molecules which further initiate the corresponding physiological and biochemical reactions to alleviate adversity stress [59, 60]. We identified 3 differential expression protein kinases during grain substance accumulation and 1 differential expression protein kinase under elevated temperature. Under natural temperature, the abundance of serine/threonine-protein kinase EDR1 (1,002,302,490) was up-regulated by 4.44-fold and 3.98-fold at 12d and 15d after flowering compared to 3d. Mitogen-activated protein kinase (MAPK) belongs to serine/threonine-protein kinase and has been validated to join in biotic and abiotic stress responses [61]. Mitogen-activated protein kinase kinase kinase 1 (1,002,252,173) was differentially expressed in grain substance accumulation and the abundance was consistently up-regulated. CBL-interacting protein kinase 9 was the only signal transduction related protein that was differentially expressed during grain substance accumulation and elevated temperature. Calmodulin B-like (CBL) proteins specifically interacted with CBL-Interacting Protein Kinases (CIPKs) to transduce calcium signals [62, 63]. Piao et al. [64] used the loss-of-function mutation of OsCIPK31 to elucidate OsCIPK31 is involved in rice germination and seedling growth under abiotic stress. During grain development, CBL-interacting protein kinase 9 obviously accumulated. However, the CBL-interacting protein kinase 9 (1,002,252,959) was down-regulated under elevated temperature especially at 6d-15d after flowering. The decreased expression of signal transduction related proteins under warming was likely to impair the ability of rice grains to respond to elevated temperature.
Reactive oxygen species (ROS) are considered unavoidable byproducts of the aerobic metabolism of plants [65]. During stress, ROS accumulates significantly in chloroplast, mitochondria, peroxisome and apoplast leading to oxidative damage [66,67,68]. However, plants have a sophisticated antioxidative mechanism that consists of enzymatic and non-enzymatic components [83]. During grain substance accumulation, the accumulation of 19 other stress response proteins was changed significantly. For example, non-specific lipid-transfer protein 1(1,002,307,908), defensin-like protein (1,002,249,243) and protein HEAT-STRESS-ASSOCIATED 32(1,002,274,021). Non-specific lipid-transfer proteins play a crucial role in plant growth and development and resistance to biotic and abiotic stress [84]. However, the abundance of non-specific lipid-transfer protein 1 was obviously reduced in our study. Different from non-specific lipid-transfer protein 1, defensin-like protein accumulated continuously. At 12d and 15d after flowering, the content of defensin-like protein was 56.89 and 147.03 folds compared to 3d. Luo et al. [85, 92]. The accumulation of pyruvate, phosphate dikinase 1, and chloroplastic isoform X1 (1,002,269,961) increased to 18, 28.64, 29.04, 22.16 folds at 6d, 9d, 12d and 15d after flowering, respectively. Sucrose synthase catalyzes the conversion of sucrose and uridine diphosphate (UDP) into fructose and UDP-glucose, which is an important starting material for storage starch biosynthesis [93,94,95]. Starch is the most abundant storage substance in the rice endosperm [96]. Therefore, sucrose synthase is vital for grain substance accumulation. Fan et al. [93] demonstrated that overexpression of sucrose synthase coding genes could significantly increase rice starch accumulation in transgenic rice to improve yield and quality. The three sucrose synthases in our study accumulated significantly during grain substance accumulation. In 3d–15d after flowering, the increase in grain weight reflected the accumulation of starch (Fig. 1 B, D). In rice, alpha-amylase isozymes are crucial for the formation of starch granules during grain maturation [97]. However, alpha-amylase may involve in the production of chalky by impairing starch accumulation in the develo** endosperm [18]. Under elevated temperature, the abundance of alpha-amylase isozyme 3E (1,002,292,698) was increased by 2.02 folds compared to natural temperature at 9 d after flowering. The up-regulation of alpha-amylase isozyme may be responsible for chalk formation. During grain substance accumulation, glutamine synthetase cytosolic isozyme 1–3 (1,002,249,042) and aspartate aminotransferase (1,002,273,225) were continuously accumulated. Glutamine synthetase catalyzes the assimilation of ammonium into glutamine which is the main way to synthesize nitrogenous organic matter by ammonium [98]. Aspartate aminotransferase, which catalyzes to generate glutamate and oxaloacetate, plays a key role in the metabolic regulation of carbon and nitrogen metabolism [99]. The accumulation of the two proteins may promote the utilization of nitrogen in rice grains, promote the synthesis of amino acids and facilitate the accumulation of proteins (Fig. 2D). In lipid metabolism, we identified some seed allergenic proteins which significantly accumulated during grain filling and response to elevated temperature (Table S6). Zhou et al. [100] found that the overexpression of RAG2 gene significantly increased grain the content of storage protein and crude lipid, and improved grain chalkiness. During grain substance accumulation, the four seed allergenic protein continuously accumulated. Under elevated temperature, the four seed allergenic proteins accumulated slightly at all times except for a slight decrease in 6d after flowering. The seed allergenic protein RA5 (1,002,280,859) was significant accumulation at 3d after flowering, which may be the key seed allergenic protein to regulate the protein and crude lipid under elevated temperature. The relationship between seed allergenic protein accumulation and grain weight and quality under elevated temperature still needs further study.
Elevated temperature triggers a series of events in plants, involving heat sensing, signal transduction, sophisticated regulatory networks or pathways, and ultimately improved plant physiological and metabolic processes in response to warming [101]. In our study, auxin synthesis and calcium signal associated with signal transduction, protein transport and modification, and reactive oxygen species-related proteins may be key proteins responsive to stimulus under elevated temperature. Plant hormones as signaling compounds are not only involved in the regulation of plant growth and development, but are also able to respond to stress response [102]. In previous studies, auxin signaling can be involved in the plant response to temperature to influence plant growth [103, 104]. In addition, the auxin signaling transduction was also affected by reactive oxygen species which are essential chemical entities and as the key secondary messengers to activate signal transduction to regulate lateral root formation [105, 106]. In our study, the accumulation of probable indole-3-acetic acid-amido synthetase GH3.4 (1,002,267,580) and probable indole-3-acetic acid-amido synthetase GH3.8 (1,002,286,982) which were related to auxin synthesis was decreased, and the abundance of reactive oxygen species-related protein was up-regulated under elevated temperature. We hypothesize that the down-regulation of auxin synthesis-related protein and the increase in reactive oxygen species accumulation under elevated temperature is a strategy for rice to cope with elevated temperature. However, whether reactive oxygen species and auxin synthesis-related proteins are in the same regulatory pathway and jointly involved in specific pathways to regulate rice response to elevated temperature still requires further study. In Protein–protein interaction networks, deoxyuridine 5'-triphosphate nucleotidohydrolase which belongs to protein modification and transport, and superoxide dismutase [Fe] 1, which belongs to reactive oxygen species-related protein, did not have direct interconnections, and the function of proteins with indirect connections remains unknown (Fig. S3). Therefore, in our study, auxin synthesis associated with signal transduction, protein transport and modification, and reactive oxygen species-related proteins may be key proteins responsive to stimulus to further regulate the accumulation of storage substance under elevated temperature. However, the closer interactions among signal transduction, protein transport and modification, and reactive oxygen species-related proteins still need to be further investigated.
Conclusion
Elevated temperature affected the accumulation of storage substance and severely deteriorated rice quality, especially significantly increased the chalkiness. We identified some differential expression proteins responsive to stimulus by proteomic and classified into signal transduction, protein transport and modification, storage substance metabolism, reactive oxygen species-related and other stress responses. Under elevated temperature, the abundance of signal transduction and other stress response-related proteins decreased and the accumulation of protein transport and modification and reactive oxygen species-related proteins increased. Together with accumulated alpha-amylase isozyme and seed allergy protein, these proteins regulated the storage substance accumulation and possibly affected rice quality formation (Fig. 9). Among the proteins we identified, the functions of related proteins in grain substance accumulation and response to elevated temperatures still need to be further revealed. The results provide new insights into the formation of rice quality under elevated temperatures and provide a new strategy for quality improvement in rice breeding.

Key proteins responsive to stimulus may regulated grain quality formation under elevated temperature. Five consecutive squares from left to right indicates different days after flowering (3, 6, 9, 12 and 15d after flowering). Values of protein abundance from minimal to maximal are colored from red to green
Materials and methods
Crop management
The field experiment was carried out in 2020 and 2021 in Danyang City at the experimental base of Nan**g Agricultural University (31。56’N, 118。59’ E). Wuyu**g 3 (W3), was a conventional japonica rice variety widely cultivated in the middle and lower reaches of the Yangtze River. W3 was sown on May 26 and May 25 and transplanted on June 13 and June 16 in 2020 and 2021 respectively. Three seedlings per hill were artificially transplanted into fields at 13.3 cm × 30 cm spacing. Nitrogen fertilizer was applied three times (120 kg N ha−1 as basal fertilizer, 60 kg N ha−1 as tiller fertilizer, and 120 kg N ha−1 as panicle fertilizer). Phosphorus (150 kg P ha−1 as phosphorus pentoxide) was used as basal fertilizer. Potassium oxide was used as potassium fertilizer with the application rate of 240 kg K ha−1 split-applied at basal fertilizer (50%), and panicle fertilizer (50%). Basal fertilizer was manually applied one day before transplanting. Water irrigation and other field management were carried out in accordance with local high-yield cultivation measures. The average temperature and precipitation during grain filling in 2020 and 2021 were shown in Fig. S4.
Temperature treatment
This study was a real field temperature experiment with two treatments CK: natural temperature and ET: elevated temperature. The elevated temperature experiment was conducted under actual field warming from the beginning of grain filling to physiological maturity by the Free-Air Temperature Enhancement (FATE). As shown in Fig. S5A, infrared radiation heating was carried out through 12 ceramic infrared heaters (240 V × 1000W) in plots of warming. The ceramic infrared heaters were installed 1.2 m above the rice canopy at an angle of 45° from the horizontal and 30° from the vertical direction. A temperature sensor (HOBO, Onset computer Corp., Bourne, MA, USA) was placed in the middle of the plot to record temperature all day. The effect of elevated temperature was shown in Fig. 10. The mean temperature of rice canopy steadily increased by 3.13 ℃ and 3.24 ℃ in 2020 and 2021 under elevated temperature. The daytime temperature increased by 1.99 ℃ (2020) and 2.41 ℃ (2021), and the nighttime temperature increased by 4.27 ℃ (2020) and 4.08 ℃ (2021) respectively. The effect of elevated temperature on the spikelet temperature was shown in Fig. 10 G and H. The thermal images were photographed between 2:00–3:00 pm at 12 d after flowering using a FLIR ThermaCAMTM S65 system (FLIR Systems Inc., Portland, OR, USA). The experiment was a randomized block design with three replicates.

Day mean temperature (A,D) and Night mean temperature (B,E) and Mean temperature (C,F) of rice canopy under natural temperature (CK) and elevated temperature (ET) during grain filling in 2020 and 2021. Thermal images of CK (G) and ET (H) spikelets at 12d after flowering in the natural state in 2020
Filling rate and yield determination
Panicles were labeled when about one third of spikelets were flowering in 2020 and 2021. The marked panicles were collected at 3, 6, 9, 12, 15, 20, 25, 35 and 45 d after flowering. Samples were put into envelope bags and dried at 80 ℃ to a constant weight in oven. The grain weight after threshing was recorded. According to the grain weight dynamics, the Richards equation was used to fit the grain filling process, and calculated the grain filling rate and key parameters [107].
Ten hills from each replicate plot were sampled at the mature stage to determine yield components in 2020 and 2021. Calculate the number of panicles to determine the panicle. The number of filled spikelets and unfilled spikelets were counted, and the spikelets per panicle and seed setting rate were calculated. The manually separated filled spikelets were dried to constant weight to be weighed and used to calculate the grain weight. Finally, the theoretical yield was calculated by the formula: yield (t ·ha−1) = panicles per hectare × spikelets per panicle × grain weight (mg) × seed setting rate × 10–6.
Determination of related indicators of rice quality
Rice grains at maturity stage were collected to determine the rice quality in 2020 and 2021.
Milling quality: 100 g rice grains with three repetitions were shelled by machine JLGJ4.5 of Taizhou Grain Industry instrument Company (Taizhou Grain Industrial Instrument Company, Zhejiang, China). Brown rice rate was obtained by weighing brown rice. Brown rice was milled for 90 s by JNMJ3 rice mill (Taizhou Grain Industry instrument Company, Zhejiang, China) to obtain milled rice. Integrity of more than 80% was manually selected from milled rice to be head rice, and used to calculate the head rice rate.
Appearance quality: The length, width and chalkiness of rice were measured by Scanmaker i800 plus (MICROTEK, Shanghai, China). The thickness of rice grain was measured by a vernier caliper.
Cooking and eating quality: The appearance and taste of rice were determined by the rice taste meter STA1BCN (Satake chemical equipment meg., ltd, Shoukou, Osaka, Japan). Hardness and viscosity of rice were used by hardness viscometer RHS1A (Satake chemical equipment meg., ltd, Shoukou, Osaka, Japan).
Crude protein content: The nitrogen content of brown rice flour was determined by the Dumas combustion method with an NCH analyzer (Sumika Chemical Analysis Service, Tokyo, Japan). The crude protein content = nitrogen content × 5.95 [108].
The amylose starch content was determined according to the people's Republic of China standard GB/17891–1999 high quality rice.
RVA profile: Rice RVA characteristics profile was measured with an RVA (Newport Scientific Pty Ltd., Warriewood, Australia). 3 g rice flour (accounting for 14% moisture basis) was suspended in 25 mL distilled water. The measurement conditions were as follows: 1 min of heating at 50 ℃, 3.8 min of heating from 50 to 95 ℃, maintenance for 2.5 min at 95 ℃, 3.8 min of cooling from 95 ℃ to 50 ℃, and holding for 1.4 min at 50 ℃.
Protein extraction
Panicles were labeled when about one third of spikelets were flowering. Panicles from the middle and upper primary branch stalks were taken at 3, 6, 9, 12 and 15d after flowering and placed at -80 °C after rapid immersion in liquid nitrogen. The grain was ground into powder for proteomic analysis after strip** the glumes on dry ice in 2018.
Protein extraction was performed as follows: (1) 5% polyvinylpolypyrrolidone (PVPP) powder, an appropriate amount of homogenate buffer and two steel balls were added to the moderate sample. (2) The mixture was shaken with a grinder for 2 min (power = 60 Hz, time = 120 s). Take out the steel ball and add 2 times the volume of saturated phenol Trissaturated Phe, shake well for 15min. (3) Centrifuge at 25,000 g for 20 min at 4 ℃, 5 times the volume of 0.1M cold ammonium acetate/methanol and the final concentration of 10mM DTT was added to the supernatant, mix well and placed in a -20 ℃ refrigerator for 2h. (4) Centrifuge at 25,000 g for 20 min at 4 ℃, discard supernatant, and repeat steps (2), (3) and (4) twice. (5) Add 1mL of cold acetone, put in refrigerator at -20 ℃ for 30min, centrifuge at 25,000 g at 4 ℃ for 20 min to discard supernatant, and repeat this step once. (6) Air-dried precipitate, an appropriate amount of SDS was added, the final concentration of 1XCocktail containing EDTA, mix well, put on the ice for 5min, add final concentration 10mM DTT; (7) Grinder (power = 60HZ, time = 2min) was used to break and split, and the supernatant was discarded after centrifugation at 25,000 g for 15 min at 4 ℃. Add final thick DTT with a degree of 10mM was placed in a 56 ℃ water bath for 1h. (8) Add IAM with a final concentration of 55mM and placed it in a dark room for 45min. (9) Add 1mL cold acetone, place in -20 ℃ refrigerator for 2h, centrifuge at 25,000 g for 15 min at 4 ℃ to discard the supernatant. (10) Air-dried precipitate, add an appropriate amount of lysis buffer without SDS to the precipitate of each tube, using a grinder (power = 60HZ, time = 2 min) to break and crack. (11) The supernatant was proteins used for quantification which was obtained at 25,000 g for 15 min at 4 ℃.
Protein extraction quality control
Bradford assay was used to measure the concentration of protein [109]. The quality of the samples was evaluated by SDS-PAGE.
Protein enzymatic hydrolysis
(1) Each sample (100 μg) was digested with Trypsin enzyme (2.5 μg) at 37 °C for 4 h. (2) Trypsin enzyme (2.5 μg) was added again and digestion was continued for 8 h at 37 °C. (3) The enzymatic peptides were desalted and vacuumed to dryness.
High pH RP separation
Mix 10ug of all samples, and 200 ug mixture was diluted with 2 mL of mobile phase A (5% ACN pH 9.8) and injected into the Shimadzu LC-20AB HPLC system coupled with a Gemini high pH C18 column (5 um, 4.6 × 250 mm). The sample was eluted at a flow rate of 1 mL/min by gradient: 5% mobile phase B (95% CAN, pH 9.8) for 10 min, 5% to 35% mobile phase B for 40 min, 35% to 95% mobile phase B for 1 min, flow Phase B lasted 3 min and 5% mobile phase B equilibrated for 10 min. Monitor the elution peak at a wavelength of 214 nm and collected the component every minute. Components were combined into 10 fractions to freeze-dried.
DDA (Data Dependent Acquisition) and DIA (Data Independent Acquisition) analysis by nano-LC–MS/MS
The freeze-dried peptide was redissolved with mobile phase A (2% ACN, 0.1% FA), centrifuged at 20,000 g for 10 min, and the supernatant was injected into Ultimate 3000 UHPLC (Thermo Scientific, Waltham, MA, USA) to separate. Samples were first enriched and desalted in trap columns, followed by connection to a self-packing C18 column (150 um inner diameter, 1.8 um column material particle size, approximately 35 cm column length), separated by different gradients at 300 nL/min flow rate: 5% mobile phase B (98% ACN, 0.1% FA) for 0–5 min; mobile phase B increased linearly from 5 to 25% for 5–120 min; mobile phase B increased from 25 to 35% for 120–160 min; mobile phase B increased from 35 to 80% for 160–170 min; 80% mobile phase B for 170–175 min; 5% Mobile Phase B for 175–180 min. The nanoliter liquid phase separation end was directly connected to the mass spectrometer.
The Q Exactive HF mass spectrometer (Thermo Fisher Scientific, San Jose, CA, USA) was applied to DDA mode detection. Main parameters: ion source voltage was set to 1.6 kV; MS scan range was 350–1500 m/z; MS resolution was set to 120,000; Maximal injection time (MIT) was set to 50 ms; MS/MS collision type was set to higher-energy collisional dissociation (HCD), normalized collision energy (NCE) was set to 28; MS/MS resolution was set to 30,000; MIT was set to 100 ms, and dynamic exclusion time was set to 30 s. The starting m/z for MS/MS was fixed to 100; the screening conditions of the precursor for MS/MS scan were: charge 2 + to 7 + , top 30 precursors with intensity over 20,000. The automatic gain control (AGC) was set to 3e6 for MS and 1e5 for MS/MS.
For DIA analysis, the same nano-LC system and gradient were used as DDA analysis. The DIA MS parameters were set as below: MS scan range was 400–1250 m/z; MS resolution was set to 120,000; MIT was set to 50 ms; DIA isolation window was set to 17 m/z with loop count 50; MIT was set automatic; scanned at resolution 30,000; stepped NCE: 22.5, 25, 27.5; The target of AGC was 1e6.
Bioinformatic analysis
DDA data was identified within MaxQuant 1.5.3.30 integrated by Andromeda search engine [110]. Spectronaut™ was used for identification results for spectral library construction [111]. For DIA data, the deconvolution and extraction were by the constructed spectral library information, and the quality control was completed by mProphet algorithm. Differential protein screening satisfied the fold change > = 2 and P value < 0.05 and further performed GO and KEGG pathway analysis.
RT-PCR (real-time reverse transcription polymerase chain reaction) Analysis
10 key differential proteins were screened for RT-PCR experiments to validate the reliability of proteomics in 2021. Total RNAs were extracted from shelled grains by the RNAprep Pure Plant Kit (TIANGEN, Bei**g, China). Analyzing concentration and purity of total RNA and identifying high-quality RNAs were used for reverse transcription using Takara’s PrimeScriptTM RT Kit (Takara Biotechnology, Tokyo, Japan). RT-PCR analysis was conducted by the Biosystems 7300 and StepOnePlusTM real-time PCR system according to Wang et al. [52]. Actin and Ubq were used as reference genes. Primers were shown in Table S8.
Data analysis
Microsoft Excel 2019 was used for data integration (Microsoft Corporation, WA, USA). ANOVA analysis was carried out by SPSS 26 software (SPSS Inc., Chicago, IL, USA). A least significant difference (LSD) of 0.05 was considered significant differences among different treatments. Origin 8.1 (OriginLab Corporation, MA, USA) was used to draw figures.
Availability of data and materials
All data supporting the findings of this study are available within the paper and its supplementary data is published online.
Abbreviations
- 2-DE:
-
Two-dimensional electrophoresis
- HSP:
-
Heat shock proteins
- DIA:
-
Data Independent Acquisition
- PCA:
-
Principal Component Analysis
- GO:
-
Gene Ontology
- KEGG:
-
Kyoto encyclopedia of genes and genomes
- ABA:
-
Abscisic acid
- BRs:
-
Brassinosteroids
- CK:
-
Cytokinin
- SA:
-
Salicylic acid
- MAPK:
-
Mitogen-activated protein kinase
- MAPKK:
-
Mitogen-activated protein kinase kinase
- CBL:
-
Calmodulin B-like
- CIPKs:
-
CBL-Interacting Protein Kinases
- CIPKs:
-
CBL-Interacting Protein Kinases
- CIPK:
-
CBL Interacting Protein Kinase
- ROS:
-
Reactive oxygen species
- SOD:
-
Superoxide dismutase
- CAT:
-
Catalase
- UDP:
-
Uridine diphosphate
- PPDK:
-
Pyruvate phosphate dikinase
- ATP:
-
Adenosine triphosphate
- PEP:
-
Phosphoenolpyruvate
- AMP:
-
Adenosine monophosphate
- RA:
-
Rice seed allergy protein
- FATE:
-
Free-Air Temperature Enhancement
- RVA:
-
Rapid viscosity analyzer
- DDA:
-
Data Dependent Acquisition
- RT-PCR:
-
Real-time reverse transcription polymerase chain reaction
References
Masson-Delmotte V, Zhai P, Pirani A, Connors SL, Péan C, Berger S, Caud N, Chen Y, Goldfarb L, Gomis MI, Huang M, Leitzell K, Lonnoy E, Matthews JBR, Maycock TK, Waterfield T, Yelekçi O, Yu R, Zhou B. (eds.) IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, Summary for Policymakers. 2021.
Wei K, Chen W. Climatology and trends of high temperature extremes across China in summer. AOSL. 2009;2(3):153–8. https://doi.org/10.1080/16742834.2009.11446795.
Tao FL, Zhang S, Zhang Z. Changes in rice disasters across China in recent decades and the meteorological and agronomic causes. Reg Environ Change. 2013;13:743–59. https://doi.org/10.1007/s10113-012-0357-7.
Shi P, Tang L, Wang L, Sun T, Liu L, Cao W, Zhu Y. Post-Heading Heat Stress in Rice of South China during 1981–2010. PLoS ONE. 2015;10(6): e0130642. https://doi.org/10.1371/journal.pone.0130642.
Deng NY, Ling XX, Sun Y, Zhang CD, Fahad S, Peng SB, et al. Influence of temperature and solar radiation on grain yield and quality in irrigated rice system. Eur J Agron. 2015;64:37–46. https://doi.org/10.1016/j.eja.2014.12.008.
Zhang H, **g WJ, XU JJ, Ma BJ, Wang WL, Zhang WY, et al. Changes in starch quality of mid-season indica rice varieties in the lower reaches of the Yangtze River in last 80 years. J Integr Agr. 2020;19:2983–96. https://doi.org/10.1016/S2095-3119(20)63431-1.
Jagadish SVK, Murty MVR, Quick WP. Rice responses to rising temperatures – Challenges, perspectives and future directions. Plant Cell Environ. 2015;38(9):1686–98. https://doi.org/10.1111/pce.12430.
Shahid M, Nayak AK, Tripathi R, Katara JL, Bihari P, Lal B, et al. Boron application improves yield of rice cultivars under high temperature stress during vegetative and reproductive stages. Int J Biometeorol. 2018;62(8):1375–87. https://doi.org/10.1007/s00484-018-1537-z.
Balcerowicz M. Filling the grain: transcription factor OsNF-YB1 triggers auxin biosynthesis to boost rice grain size. Plant Physiol. 2021;185(3):757–8. https://doi.org/10.1093/plphys/kiaa099.
An L, Tao Y, Chen H, He MJ, **ao F, Li GH, et al. Embryo-endosperm interaction and its agronomic relevance to rice quality. Front Plant Sci. 2020;11:587641. https://doi.org/10.3389/fpls.2020.587641.
Lu S, Luh BS. Properties of the rice caryopsis. In: Luh BS, editor. Rice. New York: Springer; 1991.
Yang WB, Gao MJ, Yin X, Liu JY, Xu YH, Zeng LJ, et al. Control of rice embryo development, shoot apical meristem maintenance, and grain yield by a novel cytochrome p450. Mol Plant. 2013;6(6):1945–60. https://doi.org/10.1093/mp/sst107.
Sreenivasulu N, Butardo VM Jr, Misra G, Cuevas RP, Anacleto R, Kavi Kishor PB. Designing climate-resilient rice with ideal grain quality suited for high-temperature stress. J Exp Bot. 2015;66(7):1737–48. https://doi.org/10.1093/jxb/eru544.
Zhang GL, Liao B, Li B, Cai ZH. The effect of high temperature after anthesis on rice quality and starch granule structure of endosperm. Meteorological and Environmental Research. 2016;7(03):72–5.
Min XM, Xu HL, Huang FL, Wei YD, Lin WX, Zhang ZX. GC-MS-based metabolite profiling of key differential metabolites between superior and inferior spikelets of rice during the grain filling stage. BMC Plant Biol. 2021;21(1):439. https://doi.org/10.1186/s12870-021-03219-8.
Fu J, Xu YJ, Chen L, Yuan LM, Wang ZQ, Yang JC. Changes in enzyme activities involved in starch synthesis and hormone concentrations in superior and inferior spikelets and their association with grain filling of super rice. Rice Sci. 2013;20(02):120–8. https://doi.org/10.1016/S1672-6308(13)60116-X.
** XN, Fu ZY, Ding D, Li WH, Liu ZH, Tang JH. Proteomic identification of genes associated with maize grain-filling rate. PLoS ONE. 2013;8(3):e59353. https://doi.org/10.1371/journal.pone.0059353.
Hakata M, Kuroda M, Miyashita T, Yamaguchi T, Kojima M, Sakakibara H, et al. Suppression of α-amylase genes improves quality of rice grain ripened under high temperature. Plant Biotechnol J. 2012;10(9):1110–7. https://doi.org/10.1111/j.1467-7652.2012.00741.x.
** M, Lin ZM, Zhang XC, Liu ZH, Li GH, Wang QS. Endosperm structure of white-belly and white-core rice grains shown by scanning electron microscopy. Plant Prod Sci. 2014;17(4):285–90. https://doi.org/10.1626/pps.17.285.
Shi W, Yin X, Struik PC, Solis C, **e F, Schmidt RC, et al. High day-and night-time temperatures affect grain growth dynamics in contrasting rice genotypes. J Exp Bot. 2017;68(18):5233–45. https://doi.org/10.1093/jxb/erx344.
Nakata M, Fukamatsu Y, Miyashita T, Hakata M, Kimura R, Nakata Y, et al. High temperature-induced expression of rice α-amylases in develo** endosperm produces chalky grains. Front Plant Sci. 2017;8:2089. https://doi.org/10.3389/fpls.2017.02089.
Hatakeyama Y, Masumoto-Kubo C, Nonami H, Morita S, Hiraoka K, Onda Y, et al. Evidence for preservation of vacuolar compartments during foehn-induced chalky ring formation of Oryza sativa L. Planta. 2018;248(5):1263–75. https://doi.org/10.1007/s00425-018-2975-x.
Lin CJ, Li CY, Lin SK, Yang FH, Huang JJ, Liu YH, et al. Influence of high temperature during grain filling on the accumulation of storage proteins and grain quality in rice (Oryza sativa L.). J Agric Food Chem. 2010;58(19):10545–52. https://doi.org/10.1021/jf101575j.
Yamakawa H, Hakata M. Atlas of rice grain filling-related metabolism under high temperature: joint analysis of metabolome and transcriptome demonstrated inhibition of starch accumulation and induction of amino acid accumulation. Plant Cell Physiol. 2010;51(5):795–809. https://doi.org/10.1093/pcp/pcq034.
Thitisaksakul M, Jiménez RC, Arias MC, Beckles DM. Effects of environmental factors on cereal starch biosynthesis and composition. J Cereal Sci. 2012;56(1):67–80. https://doi.org/10.1016/j.jcs.2012.04.002.
Kong XL, Zhu P, Sui ZQ, Bao JS. Physicochemical properties of starches from diverse rice cultivars varying in apparent amylose content and gelatinisation temperature combinations. Food Chem. 2015;172:433–40. https://doi.org/10.1016/j.foodchem.2014.09.085.
Cao ZZ, Zhao Q, Pan G, Wei KS, Zhou LJ, Cheng FM. Comprehensive expression of various genes involved in storage protein synthesis in filling rice grain as affected by high temperature. Plant Growth Regul. 2017;81(3):477–88. https://doi.org/10.1007/s10725-016-0225-4.
Chen JL, Tang L, Shi PH, Yang BH, Sun T, Cao WX, Zhu Y. Effects of short-term high temperature on grain quality and starch granules of rice (Oryza sativa L.) at post-anthesis stage. Protoplasma. 2017;254(2):935–43. https://doi.org/10.1007/s00709-016-1002-y.
Yao DP, Wu J, Luo QH, Li JW, Zhuang W, **ao G, et al. Influence of high natural field temperature during grain filling stage on the morphological structure and physicochemical properties of rice (oryza sativa l.) starch. Food Chem. 2020;310:125817. https://doi.org/10.1016/j.foodchem.2019.125817.
Tang S, Chen WZ, Liu WZ, Zhou QY, Zhang HX, Wang SH. Open-field warming regulates the morphological structure, protein synthesis of grain and affects the appearance quality of rice. J Cereal Sci. 2018;84:20–9. https://doi.org/10.1016/j.jcs.2018.09.013.
Dou Z, Tang S, Chen WZ, Zhang HX, Li GH, Liu ZH. Effects of open-field warming during grain-filling stage on grain quality of two japonica rice cultivars in lower reaches of Yangtze River delta. J Cereal Sci. 2018;81:118–26. https://doi.org/10.1016/j.jcs.2018.04.004.
Rodziewicz P, Swarcewicz B, Chmielewska K, Wojakowska A, Stobiecki M. Influence of abiotic stresses on plant proteome and metabolome changes. Acta Physiol Plant. 2014;36(1):1–19. https://doi.org/10.1007/s11738-013-1402-y.
Kosová K, Vítámvás P, Prášil IT, Renaut J. Plant proteome changes under abiotic stress — contribution of proteomics studies to understanding plant stress response. J Proteomics. 2011;74(8):1301–22. https://doi.org/10.1016/j.jprot.2011.02.006.
Kosová K, Vítámvás P, Urban MO, Klíma M, Roy A, Prášil IT. Biological networks underlying abiotic stress tolerance in temperate crops—a proteomic perspective. Int J Mol Sci. 2015;16(9):20913–42. https://doi.org/10.3390/ijms160920913.
Michaletti A, Naghavi MR, Toorchi M, Zolla L, Rinalducci S. Metabolomics and proteomics reveal drought-stress responses of leaf tissues from spring-wheat. Sci Rep. 2018;8(1):5710. https://doi.org/10.1038/s41598-018-24012-y.
Wang RH, Mei Y, Xu L, Zhu XW, Wang Y, Guo J, Liu LW. Differential proteomic analysis reveals sequential heat stress-responsive regulatory network in radish (Raphanus sativus L.) taproot. Planta. 2018;247(5):1109–22. https://doi.org/10.1007/s00425-018-2846-5.
Bogeat-Triboulot MB, Brosché M, Renaut J, Jouve L, Le Thiec D, Fayyaz P, et al. Gradual soil water depletion results in reversible changes of gene expression, protein profiles, ecophysiology, and growth performance in Populus euphratica, a poplar growing in arid regions. Plant Physiol. 2007;143(2):876–92. https://doi.org/10.1104/pp.106.088708.
Wu Y, Mirzaei M, Haynes PA. Proteomics of rice-our most valuable food crop. Proteomics in Food Science. 2017. https://doi.org/10.1016/B978-0-12-804007-2.00002-3.
Hashiguchi A, Ahsan N, Komatsu S. Proteomics application of crops in context of climatic changes. J Proteomics. 2011;75(1):284–305. https://doi.org/10.1016/j.jprot.2011.07.010.
Li HX, Chen Z, Hu MX, Wang ZM, Hua H, Yin CX, Zeng HL. Different effects of night versus day high temperature on rice quality and accumulation profiling of rice grain proteins during grain filling. Plant Cell Rep. 2011;30(9):1641–59. https://doi.org/10.1007/s00299-011-1074-2.
Ji W, Cong R, Li S, Li R, Qin ZW, Li YJ. Comparative proteomic analysis of soybean leaves and roots by iTRAQ provides insights into response mechanisms to short-term salt stress. Front Plant Sci. 2016;7:573. https://doi.org/10.3389/fpls.2016.00573.
Ghatak A, Chaturvedi P, Weckwerth W. Cereal crop proteomics: Systemic analysis of crop drought stress responses towards marker-assisted selection breeding. Front Plant Sci. 2017;8:757. https://doi.org/10.3389/fpls.2017.00757.
Timabud T, Yin X, Pongdontri P, Komatsu S. Gel-free/label-free proteomic analysis of develo** rice grains under heat stress. J Proteomics. 2016;133:1–19. https://doi.org/10.1016/j.jprot.2015.12.003.
Liao JL, Zhou HW, Zhang HY, Zhong PA, Huang YJ. Comparative proteomic analysis of differentially expressed proteins in the early milky stage of rice grains during high temperature stress. J Exp Bot. 2014;65(2):655–71. https://doi.org/10.1093/jxb/ert435.
Lin SK, Chang MC, Tsai YG, Lur HS. Proteomic analysis of the expression of proteins related to rice quality during caryopsis development and the effect of high temperature on expression. Proteomics. 2005;5(8):2140–56. https://doi.org/10.1002/pmic.200401105.
Liu XL, Guo T, Wan XY, Wang HY, Zhu MZ, Li AL. Transcriptome analysis of grain-filling caryopses reveals involvement of multiple regulatory pathways in chalky grain formation in rice. BMC Genomics. 2010;11:730. https://doi.org/10.1186/1471-2164-11-730.
Nevame AYM, Emon RM, Malek MA, Hasan MM, Alam MA, Muharam FM, Aslani F, Rafii MY, et al. Relationship between High Temperature and Formation of Chalkiness and Their Effects on Quality of Rice. Biomed Res Int. 2018;2018:1653721. https://doi.org/10.1155/2018/1653721.
Kumar N, Suyal DC, Sharma IP, Verma A, Singh H. Elucidating stress proteins in rice (Oryza sativa L.) genotype under elevated temperature: a proteomic approach to understand heat stress response. 3 Biotech. 2017;7(3):205. https://doi.org/10.1007/s13205-017-0856-9.
Xu H, Li XF, Zhang H, Wang LC, Zhu ZG, Gao JP. High temperature inhibits the accumulation of storage materials by inducing alternative splicing of OsbZIP58 during filling stage in rice. Plant Cell Environ. 2020;43(8):1879–96. https://doi.org/10.1111/pce.13779.
Ren Y, Huang Z, Jiang H, Wang Z, Wu FS, **ong YF. A heat stress responsive NAC transcription factor heterodimer plays key roles in rice grain filling. J Exp Bot. 2021;72(8):2947–64. https://doi.org/10.1093/jxb/erab027.
Tang S, Zhang HX, Liu WZ, Dou Z, Zhou QY, Chen WZ, et al. Nitrogen fertilizer at heading stage effectively compensates for the deterioration of rice quality by affecting the starch-related properties under elevated temperatures. Food Chem. 2019;277:455–62. https://doi.org/10.1016/j.foodchem.2018.10.137.
Wang XQ, Wang KL, Yin TY, Zhao YF, Liu WZ, Shen YY, et al. Nitrogen Fertilizer Regulated Grain Storage Protein Synthesis and Reduced Chalkiness of Rice Under Actual Field Warming. Front Plant Sci. 2021;12:715436. https://doi.org/10.3389/fpls.2021.715436.
Liu WZ, Yin TY, Zhao YF, Wang XQ, Wang KL, Shen YY. Effects of high temperature on rice grain development and quality formation based on proteomics comparative analysis under field warming. Front Plant Sci. 2021;12:746180. https://doi.org/10.3389/fpls.2021.746180.
You J, Chan Z. ROS Regulation During Abiotic Stress Responses in Crop Plants. Front Plant Sci. 2015;6:1092. https://doi.org/10.3389/fpls.2015.01092.
Zhang HM, Zhu JH, Gong ZZ, Zhu JK. Abiotic stress responses in plants. Nat Rev Genet. 2022;23(2):104–19. https://doi.org/10.1038/s41576-021-00413-0.
Waadt R, Seller CA, Hsu PK, Takahashi Y, Munemasa S, Schroeder JI. Plant hormone regulation of abiotic stress responses. Nat Rev Mol Cell Biol. 2022;23(10):680–94. https://doi.org/10.1038/s41580-022-00479-6.
Gray WM, Ostin A, Sandberg G, Romano CP, Estelle M. High temperature promotes auxin-mediated hypocotyl elongation in Arabidopsis. Proc Natl Acad Sci U S A. 1998;95(12):7197–202. https://doi.org/10.1073/pnas.95.12.7197.
Xu ZS, Liu L, Ni ZY, Liu P, Chen M, Li LC, et al. W55a encodes a novel protein kinase that is involved in multiple stress responses. J Integr Plant Biol. 2009;51(1):58–66. https://doi.org/10.1111/j.1744-7909.2008.00776.x.
Liu P, Xu ZS, Pan-Pan L, Hu D, Chen M, Li LC, et al. A wheat PI4K gene whose product possesses threonine autophophorylation activity confers tolerance to drought and salt in Arabidopsis. J Exp Bot. 2013;64(10):2915–27. https://doi.org/10.1093/jxb/ert133.
Wang P, Hsu CC, Du Y, Zhu P, Zhao C, Fu X. Map** proteome-wide targets of protein kinases in plant stress responses. Proc Natl Acad Sci U S A. 2020;117(6):3270–80. https://doi.org/10.1073/pnas.1919901117.
Chen J, Wang LH, Yuan M. Update on the roles of rice MAPK cascades. Int J Mol Sci. 2021;22(4):1679. https://doi.org/10.3390/ijms22041679.
Liu P, Duan Y, Liu C, Xue Q, Guo J, Qi T. The calcium sensor TaCBL4 and its interacting protein TaCIPK5 are required for wheat resistance to stripe rust fungus. J Exp Bot. 2018;69(18):4443–57. https://doi.org/10.1093/jxb/ery227.
Deng JW, Yang XY, Sun WN, Miao YH, He LR, Zhang XL. The calcium sensor CBL2 and its interacting kinase CIPK6 are involved in plant sugar homeostasis via interacting with tonoplast sugar transporter TST2. Plant Physiol. 2020;183(1):236–49. https://doi.org/10.1104/pp.19.01368.
Piao HL, Xuan YH, Park SH, Je BI, Park SJ, Park SH, et al. OsCIPK31, a CBL-interacting protein kinase is involved in germination and seedling growth under abiotic stress conditions in rice plants. Mol Cells. 2010;30(1):19–27. https://doi.org/10.1007/s10059-010-0084-1.
Mittler R, Vanderauwera S, Suzuki N, Miller G, Tognetti VB, Vandepoele K, et al. ROS signaling: the new wave? Trends Plant Sci. 2011;16(6):300–9. https://doi.org/10.1016/j.tplants.2011.03.007.
Baxter A, Mittler R, Suzuki N. ROS as key players in plant stress signalling. J Exp Bot. 2014;65(5):1229–40. https://doi.org/10.1093/jxb/ert375.
Huang S, Van Aken O, Schwarzländer M, Belt K, Millar AH. The roles of mitochondrial reactive oxygen species in cellular signaling and stress response in plants. Plant Physiol. 2016;171(3):1551–9. https://doi.org/10.1104/pp.16.00166.
Nadarajah KK. ROS Homeostasis in Abiotic Stress Tolerance in Plants. Int J Mol Sci. 2020;21(15):5208. https://doi.org/10.3390/ijms21155208.
Duan J, Zhang M, Zhang H, **ong H, Liu P, Ali J, et al. OsMIOX, a myo-inositol oxygenase gene, improves drought tolerance through scavenging of reactive oxygen species in rice (Oryza sativa L.). Plant Sci. 2012;196:143–51. https://doi.org/10.1016/j.plantsci.2012.08.003.
Das K, Roychoudhury A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front Environ Sci. 2014;2:53. https://doi.org/10.3389/fenvs.2014.00053.
Gill SS, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem. 2010;48(12):909–30. https://doi.org/10.1016/j.plaphy.2010.08.016.
Lopez-Ortiz C, Dutta SK, Natarajan P, Peña-Garcia Y, Abburi V, Saminathan T, et al. Genome-wide identification and gene expression pattern of ABC transporter gene family in Capsicum spp. PLoS ONE. 2019;14(4):e0215901. https://doi.org/10.1371/journal.pone.0215901.
Nguyen V, Moon S, Jung KH. Genome-wide expression analysis of rice ABC transporter family across spatio-temporal samples and in response to abiotic stresses. J Plant Physiol. 2014;171(14):1276–88. https://doi.org/10.1016/j.jplph.2014.05.006.
Fernández-Fernández MR, Valpuesta JM. Hsp70 chaperone: a master player in protein homeostasis. F1000Res. 2018;7:F1000 Faculty Rev-1497. https://doi.org/10.12688/f1000research.15528.1.
Fragkostefanakis S, Mesihovic A, Hu Y, Schleiff E. Unfolded protein response in pollen development and heat stress tolerance. Plant Reprod. 2016;29(1–2):81–91. https://doi.org/10.1007/s00497-016-0276-8.
Müller F, Rieu I. Acclimation to high temperature during pollen development. Plant Reprod. 2016;29(1–2):107–18. https://doi.org/10.1007/s00497-016-0282-x.
Hartl FU, Bracher A, Hayer-Hartl M. Molecular chaperones in protein folding and proteostasis. Nature. 2011;475(7356):324–32. https://doi.org/10.1038/nature10317.
Ono K, Hibino T, Kohinata T, Suzuki S, Tanaka Y, Nakamura T, et al. Overexpression of DnaK from a halotolerant cyanobacterium Aphanothece halophytica enhances the high-temperatue tolerance of tobacco during germination and early growth. Plant Sci. 2001;160(3):455–61. https://doi.org/10.1016/s0168-9452(00)00412-x.
Yutaka, SatoSakiko, Yokoya. Enhanced tolerance to drought stress in transgenic rice plants overexpressing a small heat-shock protein, sHSP17.7. Plant Cell Rep. 2008;27(2):329–34. https://doi.org/10.1007/s00299-007-0470-0.
Satoh-Cruz M, Crofts AJ, Takemoto-Kuno Y, Sugino A, Washida H, Crofts N, et al. Protein disulfide isomerase like 1–1 participates in the maturation of proglutelin within the endoplasmic reticulum in rice endosperm. Plant Cell Physiol. 2010;51(9):1581–93. https://doi.org/10.1093/pcp/pcq098.
Han XH, Wang YH, Liu X, Jiang L, Ren YL, Liu F, et al. The failure to express a protein disulphide isomerase-like protein results in a floury endosperm and an endoplasmic reticulum stress response in rice. J Exp Bot. 2012;63(1):121–30. https://doi.org/10.1093/jxb/err262.
Hamzelou S, Pascovici D, Kamath KS, Amirkhani A, McKay M, Mirzaei M, et al. Proteomic responses to drought vary widely among eight diverse genotypes of rice (Oryza sativa). Int J Mol Sci. 2020;21(1):363. https://doi.org/10.3390/ijms21010363.
Suzuki N, Rivero RM, Shulaev V, Blumwald E, Mittler R. Abiotic and biotic stress combinations. New Phytol. 2014;203(1):32–43. https://doi.org/10.1111/nph.12797.
Yang Y, Li P, Liu C, Wang P, Cao P, Ye X, Li Q. Systematic analysis of the non-specific lipid transfer protein gene family in Nicotiana tabacum reveal its potential roles in stress responses. Plant Physiol Biochem. 2022;172:33–47. https://doi.org/10.1016/j.plaphy.2022.01.002.
Luo JS, Huang J, Zeng DL, Peng JS, Zhang GB, Ma HL, et al. A defensin-like protein drives cadmium efflux and allocation in rice. Nat Commun. 2018;9(1):645. https://doi.org/10.1038/s41467-018-03088-0.
Luo JS, **ao Y, Yao J, Wu Z, Yang Y, Ismail AM, et al. Overexpression of a defensin-like gene cal2 enhances cadmium accumulation in plants. Front Plant Sci. 2020;11:217. https://doi.org/10.3389/fpls.2020.00217.
Lin MY, Chai KH, Ko SS, Kuang LY, Lur HS, Charng YY. A positive feedback loop between HEAT SHOCK PROTEIN101 and HEAT STRESS-ASSOCIATED 32-KD PROTEIN modulates long-term acquired thermotolerance illustrating diverse heat stress responses in rice varieties. Plant Physiol. 2014;164(4):2045–53. https://doi.org/10.1104/pp.113.229609.
Shimada TL, Shimada T, Takahashi H, Fukao Y, Hara-Nishimura I. A novel role for oleosins in freezing tolerance of oilseeds in arabidopsis thaliana. Plant J. 2008;55(5):798–809. https://doi.org/10.1111/j.1365-313X.2008.03553.x.
Wood HG, O’Brien WEW, Micheales G. Properties of carboxytransphosphorylase; pyruvate, phosphate dikinase; pyrophosphate-phosphofructikinase and pyrophosphate-acetate kinase and their roles in the metabolism of inorganic pyrophosphate. Adv Enzymol Relat Areas Mol Biol. 1977;45:85–155. https://doi.org/10.1002/9780470122907.ch2.
Hennen-Bierwagen TA, Lin Q, Grimaud F, Planchot V, Keeling PL, James MG, et al. Proteins from multiple metabolic pathways associate with starch biosynthetic enzymes in high molecular weight complexes: a model for regulation of carbon allocation in maize amyloplasts. Plant Physiol. 2009;149(3):1541–59. https://doi.org/10.1104/pp.109.135293.
Méchin V, Thévenot C, Le Guilloux M, Prioul JL, Damerval C. Developmental analysis of maize endosperm proteome suggests a pivotal role for pyruvate orthophosphate dikinase. Plant Physiol. 2007;143(3):1203–19. https://doi.org/10.1104/pp.106.092148.
Chastain CJ, Heck JW, Colquhoun TA, Voge DG, Gu XY. Posttranslational regulation of pyruvate, orthophosphate dikinase in develo** rice (Oryza sativa) seeds. Planta. 2006;224(4):924–34. https://doi.org/10.1007/s00425-006-0259-3.
Fan CF, Wang GY, Wang YM, Zhang R, Wang YT, Feng SQ, et al. Sucrose synthase enhances hull size and grain weight by regulating cell division and starch accumulation in transgenic rice. Int J Mol Sci. 2019;20(20):4971. https://doi.org/10.3390/ijms20204971.
Baroja-Fernández E, Muñoz FJ, Montero M, Etxeberria E, Sesma MT, Ovecka M, et al. Enhancing sucrose synthase activity in transgenic potato (Solanum tuberosum L.) tubers results in increased levels of starch, ADP glucose and UDP glucose and total yield. Plant Cell Physiol. 2009;50(9):1651–62. https://doi.org/10.1093/pcp/pcp108.
Cakir B, Shiraishi S, Tuncel A, Matsusaka H, Satoh R, Singh S, et al. Analysis of the rice ADP glucose transporter (OsBT1) indicates the presence of regulatory processes in the amyloplast stroma that control ADP glucose flux into starch. Plant Physiol. 2016;170(3):1271–83. https://doi.org/10.1104/pp.15.01911.
Zhang H, Xu H, Jiang YY, Zhang H, Wang SY, Wang FL, et al. Genetic control and high temperature effects on starch biosynthesis and grain quality in rice. Front Plant Sci. 2021;12:757997. https://doi.org/10.3389/fpls.2021.757997.
Damaris RN, Lin Z, Yang P, He D. The rice alpha-amylase, conserved regulator of seed maturation and germination. Int J Mol Sci. 2019;20(2):450. https://doi.org/10.3390/ijms20020450.
Guan M, de Bang TC, Pedersen C, Schjoerring JK. Cytosolic Glutamine Synthetase Gln1;2 Is the Main Isozyme Contributing to GS1 Activity and Can Be Up-Regulated to Relieve Ammonium Toxicity. Plant Physiol. 2016;171(3):1921–33. https://doi.org/10.1104/pp.16.01195.
de la Torre F, De Santis L, Suárez MF, Crespillo R, Cánovas FM. Identification and functional analysis of a prokaryotic-type aspartate aminotransferase: implications for plant amino acid metabolism. Plant J. 2006;46(3):414–25. https://doi.org/10.1111/j.1365-313X.2006.02713.x.
Zhou W, Wang X, Zhou D, Ouyang Y, Yao JL. Overexpression of the 16-kDa α-amylase/trypsin inhibitor RAG2 improves grain yield and quality in rice. Plant Biotechnol J. 2017;15(5):568–80. https://doi.org/10.1111/pbi.12654.
Xu YF, Chu CC, Yao SG. The impact of high-temperature stress on rice: challenges and solutions. Crop J. 2021;9(5):963–76. https://doi.org/10.1016/j.cj.2021.02.011.
Waadt R, Seller CA, Hsu PK, Takahashi Y, Munemasa S, Schroeder JI. Plant hormone regulation of abiotic stress responses. Nat Rev Mol Cell Biol. 2022;23(10):680–694. https://doi.org/10.1038/s41580-022-00479-6.
Sun J, Qi L, Li Y, Chu J, Li C. PIF4-mediated activation of YUCCA8 expression integrates temperature into the auxin pathway in regulating Arabidopsis hypocotyl growth. PLoS Genet. 2012;8(3):e1002594. https://doi.org/10.1371/journal.pgen.1002594.
Wang R, Zhang Y, Kieffer M, Yu H, Kepinski S, Estelle M. HSP90 regulates temperature-dependent seedling growth in Arabidopsis by stabilizing the auxin co-receptor F-box protein TIR1. Nat Commun. 2016;7:10269. https://doi.org/10.1038/ncomms10269.
Mittler R, Zandalinas SI, Fichman Y, Van Breusegem F. Reactive oxygen species signalling in plant stress responses. Nat Rev Mol Cell Biol. 2022;23(10):663–679. https://doi.org/10.1038/s41580-022-00499-2.
Biswas MS, Fukaki H, Mori IC, Nakahara K, Mano J. Reactive oxygen species and reactive carbonyl species constitute a feed-forward loop in auxin signaling for lateral root formation. Plant J. 2019;100(3):536–548. https://doi.org/10.1111/tpj.14456.
Richards F. A flexible growth function for empirical use. J exp bot. 1959;10:290–300.
Tsukaguchi T, Taniguchi Y, Ito R. The effects of nitrogen uptake before and after heading on grain protein content and the occurrence of basal- and back-white grains in rice (Oryza sativa L.). Plant Prod Sci. 2016; 19(4):508–17. https://doi.org/10.1080/1343943X.2016.1223527.
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. https://doi.org/10.1006/abio.1976.9999.
Cox J, Mann M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat Biotechnol. 2008;26(12):1367–1372. https://doi.org/10.1038/nbt.1511.
Bruderer R, Bernhardt OM, Gandhi T, Miladinović SM, Cheng LY, Messner S, et al. Extending the limits of quantitative proteome profiling with data-independent acquisition and application to acetaminophen-treated three-dimensional liver microtissues. Mol Cell Proteomics. 2015;14(5):1400–10. https://doi.org/10.1074/mcp.M114.044305.
Acknowledgements
Not applicable.
Statement
The rice seeds used in this study were collected and preserved by Nan**g Agricultural University.
These seeds were collected and used in full compliance with the IUCN Policy Statement on Research Involving Species at Risk of Extinction.
Funding
This work was funded by the National Natural Science Foundation of China (Grant No. 32071949, 32372220&31701366). We also received support from the Collaborative Innovation Center for Modern Crop Production co-sponsored by Province and Ministry (CIC-MCP) and Chuzhou Science and Technology Program, China (Grant No. 2022ZN001).
Author information
Authors and Affiliations
Contributions
She Tang and Yanfeng Ding conceived the experiments. She Tang designed the experiments. Tongyang Yin performed the proteomics data analysis. Xuan Ran, Wenzhe Liu and Yingying Shen did the sampling and determined the rice quality. Hao Guo, Yuxuan Peng and Chen Zhang did data analysis. Yufei Zhao wrote the manuscript and She Tang revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhao, Y., Yin, T., Ran, X. et al. Stimulus-responsive proteins involved in multi-process regulation of storage substance accumulation during rice grain filling under elevated temperature. BMC Plant Biol 23, 547 (2023). https://doi.org/10.1186/s12870-023-04563-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-023-04563-7