
Abstract
Background
Septoria tritici blotch (STB), caused by Zymoseptoria tritici (Z. tritici), is an important biotic threat to durum wheat in the entire Mediterranean Basin. Although most durum wheat cultivars are susceptible to Z. tritici, research in STB resistance in durum wheat has been limited.
Results
In our study, we have identified resistance to a wide array of Z. tritici isolates in the Tunisian durum wheat landrace accession ‘Agili39’. Subsequently, a recombinant inbred population was developed and tested under greenhouse conditions at the seedling stage with eight Z. tritici isolates and for five years under field conditions with three Z. tritici isolates. Map** of quantitative trait loci (QTL) resulted in the identification of two major QTL on chromosome 2B designated as Qstb2B_1 and Qstb2B_2. The Qstb2B_1 QTL was mapped at the seedling and the adult plant stage (highest LOD 33.9, explained variance 61.6%), conferring an effective resistance against five Z. tritici isolates. The Qstb2B_2 conferred adult plant resistance (highest LOD 32.9, explained variance 42%) and has been effective at the field trials against two Z. tritici isolates. The physical positions of the flanking markers linked to Qstb2B_1 and Qstb2B_2 indicate that these two QTL are 5 Mb apart. In addition, we identified two minor QTL on chromosomes 1A (Qstb1A) and chromosome 7A (Qstb7A) (highest LODs 4.6 and 4.0, and explained variances of 16% and 9%, respectively) that were specific to three and one Z. tritici isolates, respectively. All identified QTL were derived from the landrace accession Agili39 that represents a valuable source for STB resistance in durum wheat.
Conclusion
This study demonstrates that Z. tritici resistance in the ‘Agili39’ landrace accession is controlled by two minor and two major QTL acting in an additive mode. We also provide evidence that the broad efficacy of the resistance to STB in ‘Agili 39’ is due to a natural pyramiding of these QTL. A sustainable use of this Z. tritici resistance source and a positive selection of the linked markers to the identified QTL will greatly support effective breeding for Z. tritici resistance in durum wheat.
Similar content being viewed by others

Background
Wheat has been, for centuries, the prime food and feed crop especially in the Mediterranean basin [1]. This staple crop supplies 20% of the human calorie intake, and is thereby a major component for global food security [2, 3]. The genus Triticum L. comprises several wheat species with various ploidy levels, but global wheat production is almost entirely based on bread wheat, T. aestivum L. em. Thell. (2n = 6x = 42, sub-genomes AABBDD), and durum wheat, T. turgidum L. var. durum (2n = 4x = 28, sub-genomes AABB), also known as pasta wheat [4]. Durum wheat accounts for about 8% to the global wheat production, and its cultivation is concentrated in latitudes ranging from 55°N to 40°S [5, 6], corresponding mostly to the Mediterranean Basin, the North American Great Plains, India and the former USSR [6]. Durum wheat is also produced in sub-Saharan Africa (SSA) where Ethiopia is a leading country and considered as one of the biggest durum wheat producers with approximately 0.6 million ha [7], and a center of diversity for tetraploid wheat [8]. Northern Africa has been also the cradle of wheat production for centuries and was the breadbasket for the Roman Empire [9, 10] with locations such as Dougga in Tunisia, as exquisite trading zones for wheat and other commodities until the late 500’s AD [11]. In Tunisia, durum wheat occupies 725 Mha approximates representing 49% of the total annual cereal area [12], with an average yield estimated at 1.7 tons per hectare between the crop** seasons 2014/2015 and 2019/2020 [13].
Alike bread wheat, durum wheat production is significantly affected by abiotic stress conditions—mostly drought—and by the emergence of more aggressive pathogens [14]. Throughout Maghreb region, the foliar blight septoria tritici blotch (STB), caused by the hemibiotroph Zymoseptoria tritici (Desm.) Quaedvlieg & Crous (formerly Mycosphaerella graminicola (Fuckel) J. Schröt. in Cohn), is among the major threats [15]. Estimated yield losses amounted up to 385 kg.ha−1 in 2008–2009, which is more than 30% in most regions [16]. Recent research increased the general understanding of the Z. tritici epidemiology in the Maghreb. Hamada [17] reported the occurrence of the teleomorph of the fungus in Tunisia, despite the arid conditions in the region, and Neddaf et al. [18] determined an equal distribution of both mating types in Algeria, indicating regular sexual reproduction, which likely contributes to the vast genetic diversity in this region. Similar results have recently been reported in Tunisia on durum wheat [19]. The use of fungicides has been more slowly adopted by durum wheat growers as compared to bread wheat producers in Europe, and the first occurrence of strobilurin resistance have been reported in Tunisia and Algeria [18, 20].
One of the best management strategies for all plant diseases is the generation of new disease resistant germplasm through plant breeding. The huge genetic diversity in wheat and its ancestors has provided new varieties for almost a century [4]. Releasing new resistant germplasm has proven its efficacy and has turned the potential havoc of re-emerging and upcoming threats into a manageable problem [21, 22], such as the stem rust caused by the Ug99 strain [23,24,25]. Before modern plant breeding, improved crops frequently resulted from farmers’ selections of outperforming genotypes in terms of yield stability. Often, such so-called landraces contained a variety of closely related lines that quenched biotic threats. During the onset of breeding, these landraces were often the starting material for targeted efforts to improve for instance disease resistance [26,27,28].
Several studies have revealed that durum wheat landraces are a valuable source of resistance alleles against fungal pathogens [8, 12, 29,30,31]. STB resistance sources on durum wheat were identified in many countries such as Tunisia [12], Ethiopia [8], Iran [32] and Spain [33]. Until now, up to 22 septoria tritici blotch (Stb) resistance genes have been identified and mapped [34, 35]. However, due to the apparent dichotomy in natural Z. tritici populations for either bread wheat or durum wheat [36,37,38], the presence of these mapped Stb genes in durum wheat cannot be determined using well characterized Z. tritici strains originating from bread wheat. Thus far, the substantial research progress is mainly based on the Z. tritici – bread wheat pathosystem [34, 39]. Therefore, and albeit the growing efforts to dissect the durum wheat – Z. tritici interactions [12, 33, 40, 41], resistance breeding to Z. tritici in durum wheat has slowly progressed over the last 25 years compared to bread wheat [34]. This affects many small growers who produce this wheat as a staple crop in an area that is severely struck by septoria tritici blotch.
In this study, we have embarked on increasing the understanding of the Z. tritici—durum wheat pathosystem. Here, we have first screened eight durum wheat Tunisian landrace accessions for Z. tritici resistance. Subsequently, we developed a map** population between the resistant landrace accession ‘Agili39’ and the susceptible modern cv. Khiar to identify the genetic basis of resistance to Z. tritici and to map the underlying genes under greenhouse and field conditions.
Results
Phenoty** of RILs, landrace accessions and modern cultivars at the seedling stage
The 20 Z. tritici isolates grew successfully under laboratory conditions enabling appropriate inoculum production and phenoty**. None of the tested durum landraces and cultivars was resistant to the entire suite of Z. tritici isolates (Table 1), but the landrace accessions showed a broader efficacy compared to the cvs. Khiar and Karim, resulting in a significant ‘line x isolate’ interaction, indicating specific gene action (Additional Table 1). Interestingly, necrosis values were high and ranged between 72 and 97% (data not shown). The parents of the developed RILs, ‘Agili39’ and cv. Khiar showed highly significantly different pycnidia values of 6% and 36%, respectively (Table 1), and henceforward, we selected a set of eight Z. tritici isolates that discriminated between ‘Agili39’ and cv. Khiar for subsequent phenoty** of the developed F6 RILs population (Table 2).
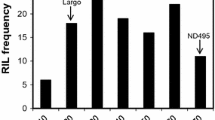
The seedling screening of the RILs with the selected eight Z. tritici isolates resulted in non-symmetric frequency distributions skewed towards the resistance phenotype for all tested isolates (Fig. 1 panel A, Additional Fig. 1). Subsequent analyses of variance of the split-plot design seedling experiment revealed that the ‘RIL’ term was highly significant for necrosis and pycnidia AUDPC scores at p = 0.0001 (Table 3). This result indicates that the observed variation in the data is accounted for the variable genetic make-up of the tested lines.

Frequency distributions of the disease severity assessed as pycnidia percentage in seedlings and adult plants of the F6-F10 recombinant inbred lines of Agili39/Khiar with three Zymoseptoria tritici under field and controlled conditions. ‘A’ and ‘K’ are referring to ‘Agili39’ and cv. Khiar parents, respectively
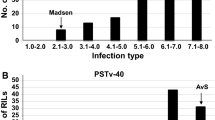
Differentiation between the isolates was observed for necrosis and pycnidia scores in the RIL population tested at the seedling stage (Table 4). The highest population mean necrosis coverage (60.60%) was registered for RILs inoculated with IPO92003 isolate (Table 4). However, the least population mean necrosis coverage (34.5%) was observed for RILs inoculated with Tun1 isolate. Pycnidia population mean scores were also variable and were high for lines inoculated with Tun6 isolate (23.9%), but relatively low for lines inoculated with IPO95052 isolate (9.4%). For all tested isolates, necrosis scores ranged between 0 and 100%, however a maximum of 90% of pycnidia score was registered for lines inoculated with IPO95052 isolate (Table 4). All seedling phenotypic traits were repeatable with the highest repeatability registered for isolates IPO92003 and Tun6 for necrosis and pycnidia scores, respectively and with an equal repeatability of 0.96 for both seedling phenotypic traits and isolates (Table 4).
Phenoty** of RILs at the adult plant stage
During all field trials, STB developed well after the inoculations, but only pycnidia coverage was assessed. The susceptible parent cv. Khiar showed high disease severities throughout the trials, whereas ‘Agili39’ remained free of disease (0 pycnidia) (Fig. 1 panel B). The three-way analysis of variance revealed that the ‘genotype’, ‘isolate’ and their interaction ‘genotype x isolate’ terms are highly significant at p = 0.0001 (Table 5). This result indicates a ‘genotype x isolate’ specificity at the adult plant. The term ‘year’ was significant at p = 0.05. However, the term ‘block’, the two-way interaction terms ‘genotype x block’ and ‘genotype x year’, and the three-way interaction term ‘genotype x year x isolate’ were not significant, indicating no variation in the micro-environment and the homogeneity of the field inoculation (Table 5).
Overall, adult plants F9 showed the highest average of pycnidia coverage compared to the F7 inoculated with the Tun1 isolate (21.5% of pycnidia for the F9 compared to 15.4% for the F7), and to the F6 and the F8 generations inoculated with the Tun6 isolate (29.9% of pycnidia for the F9 compared to 19.6% and 19.0% for the F6 and the F8, respectively) (Table 4, Fig. 1 panel B). The field disease severity on the F9 generation inoculated with Tun1 and Tun6 isolates was also variable with a higher severity for Tun6 isolate (29.9%) compared to the Tun1 isolate (21.5%), which indicates once more a specific ‘genotype x isolate’ interaction. Interestingly, average pycnidia coverage was relatively low at the F10 generation inoculated with the IIB-123 isolate (6.5% of pycnidia), with a maximum pycnidia coverage of 19.4% (Table 4).
Adult pycnidia coverage heritability (H2) was high for the Tun1 and the Tun6 isolates, with a higher heritability for the adult pycnidia coverage caused by the Tun6 isolate (H2 = 0.98) compared to the heritability of the pycnidia coverage caused by the Tun1 isolate (H2 = 0.88) (Table 4). Field data generated by the inoculation of the F10 RIL with the IIB123 isolate were also repeatable (0.97) (Table 4).
Correlations between the seedling and the adult plant assays
Low to high correlations were observed between the different traits (Fig. 2, Additional Table 2). Phenotypic scores obtained by the IPO92003 isolate were the least correlated with all phenotypic scores obtained by the tested isolates at the seedling and the adult plant stages (Fig. 2, Additional Table 2). Nonetheless, necrosis and pycnidia AUDPC scores generated by the IPO92003 isolate on the tested RILs at the seedling stage were highly correlated (r = 0.6). Z. tritici isolates IPO95052, IPO91009, Tun6, IIB-123 and IPO91004 tested at the seedling stage were correlated between each other for necrosis and pycnidia AUDPC scores (0.3 < r > 0.5) (Fig. 2, Additional Table 2). The highest correlation coefficient (r) of 0.9 was registered between pycnidia AUDPC scores generated on lines inoculated by the IPO91004 and the II-B123 Z. tritici isolates at the seedling stage. In contrast, necrosis and pycnidia scores were rather moderately correlated between IPO91018, IIB-123, IPO95052 and IP91004 isolates with a maximum r of 0.5 (Additional Table 2). Field data generated by Tun6 isolate were highly correlated across the F6 – F9 generation (r = 0.8) (Additional Table 2). Interestingly, positive correlations were also observed between adult and seedling disease scores induced by Tun6 isolate (Additional Table 2). Similarly, positive correlations were registered between field scores generated by Tun6, and II-B123 isolates, with a correlation coefficient r = 0.4 (Fig. 2, Additional Table 2).

Heatmap correlation between the Zymoseptoria tritici isolates tested under field and controlled conditions on the ‘Agili39’/Khiar map** population performed using the “Corr” function and visualized using the “corrplot” package in the R environment [42, Full size table
Finally, we selected the pairwise epistasis Qstb2B_1 / Qstb5B and Qstb2B_2 / Qstb5B showing the highest epistatic LODs for a two-way interaction test to examine pycnidia AUDPC means of the variant allele combinations linked to QTL Qstb2B_1, Qstb2B_2 and Qstb5B (Fig. 4). The first allele refers to the SNP markers linked to Qstb2B_1 or Qstb2B_2, however the second allele refers to the Qstb5B SNP marker (Fig. 4).

Epistatic interactions between QTL Qstb2B_1 and Qstb5B (Panel A), and between Qstb2B_2 and Qstb5B (Panel B). Box plots illustrate the significant effects of allele variants on pycnidia AUDPC on seedlings inoculated with the IPO91004 and the IPO95052 isolates. “R” and “S” denote the resistant and the susceptible alleles at each locus, respectively. The first allele refers to the SNP markers linked to Qstb2B_1 or Qstb2B_2, however the second allele refers to the Qstb5B SNP marker. Number of genotypes and mean values of Pycnidia AUDPC are indicated under each allele classes. The black horizontal lines in the middle of the boxes are the median values for the Pycnidia AUDPC value in the respective allele classes. The vertical size of the boxes represents the inter-quantile range. The upper and lower whiskers represent the minimum and maximum values of data
Pycnidia AUDPC means were reduced when pairing the resistant alleles (RR) linked to Qstb2B_1 /Qstb5B QTL and linked to Qstb2B_2/Qstb5B with pycnidia AUDPC scores of 7.4 and 0.8 for lines inoculated with the IPO91004 and IPO95052 Z. tritici isolates at the seedling stage, respectively (Fig. 4, panel A and B). Lines carrying the susceptible allele linked to the Qstb5B QTL (RS) showed an increase in pycnidia AUDPC scores compared to lines carrying both resistant allele (RR) (Fig. 4, panel A and B). Lines carrying solely the resistant allele linked to Qstb5B (SR) were rather susceptible showing high pycnidia AUDPC scores for both pairwise interactions Qstb2B_1/Qstb5B and Qstb2B_2/Qstb5B. This observation could be explained by the minor effect of the Qstb5B QTL in controlling pycnidia development, compared to the major effect of the Qstb2B_1 and Qstb2B_2 QTL in reducing pycnidia development.
Discussion
Zymoseptoria tritici is a major threat to European and Mediterranean bread and durum wheat production [44]. Despite the increasing efforts to elucidate the genetic basis of tetraploid wheat resistance to STB [8, 40, 45], more studies are required for an effective breeding strategy in durum wheat for STB resistance.
In our study, all data indicated and confirmed significant ‘isolate’ and ‘line x isolate’ interactions as determined in earlier studies [37, 38, 46, 47]; and recently proven in the bread wheat – Z. tritici pathosystem where both Stb6 and AvrStb6 genes were cloned [48,49,50]. Moreover, and comparably to other cereal diseases, namely to rust [51,52,53,54], we determined QTL that are detected for either seedling (Qstb1A and Qstb7A) or adult plant stage (Qstb2B_2), as well as a QTL that was detected at both stages (Qstb2B_1). Our findings confirm that specific plant physiological stage resistances are commonly observed in the Z. tritici – wheat pathosystem [34]. Specific plant physiological stage resistances were also confirmed for other fungal diseases such as the powdery mildew and the leaf rust diseases [55, 56]. In fact, some Z. tritici resistance genes are uniquely effective at the seedling stage, such as the Stb7 gene mapped in the spring wheat cultivar ST6 [57], or at the adult plant stage, such as the Stb17 gene [58, 59]. In contrast, other resistance genes have proven to be effective at both seedling and adult plant stages alike the Stb4 and Stb5 qualitative genes [60, 61].
Subsequently, we compared the identified QTL to formerly identified Z. tritici genes using the reported literature. This comparison has revealed that a putative QTL for resistance to Z. tritici was mapped on chromosome 1A at 68 cM at the adult plant stage by Kidane et al. [8] through a genome-wide association study conducted on an Ethiopian durum wheat landrace population. Two other QTL mapped on chromosome 1A were also revealed by Goudemand et al. [62] in the bread wheat Apache/Balance population, and by Risser et al. [63] named as QStb.lsa_fb-1A in the bread wheat bi-parental map** population Florett/Biscay. These QTL were mapped at the adult plant stage between 56 and 69 cM and could thus co-localize with the QTL mapped in the ‘Agili39’/Khiar population. However, and in contrast to the above-mentioned studies, the Qstb1A QTL mapped in the ‘Agili39’/Khiar population was solely detected at the seedling plant stage.
The Qstb7A QTL particularly conferred reduced necrosis values to Z. tritici isolate IIB123 and co-localizes with the major Stb3 gene that was mapped in the bread wheat cultivar Israel 493 [64, 65].
Two QTL for STB resistance were mapped on chromosome 2B in the ‘Agili39’/khiar population. Other studies have also revealed genomic regions on chromosome 2B associated with the STB resistance [8, 62, 66,67,68,69].
The Qstb2B_1 QTL identified in the ‘Agili39’/Khiar population likely co-localized with the known major gene Stb9 that was mapped in the French bread wheat cv. Courtot [66]; with the qSTB.2 QTL mapped in the Ethiopian durum wheat landrace population [8]; with the QStb.ihar-2B.2 QTL mapped in the Liwilla/Begra bread wheat doubled-haploid population [68]; and with the QStb.lfl-2B.1 mapped in the eight-founder MAGIC population of winter wheat [69]. However, the Qstb2B_1 QTL is different from the QTL mapped at the long arm of chromosome 2B in the Nimbus/Stigg bread wheat map** population [67].
The Qstb2B_2 QTL likely co-localized with the QTL identified on chromosome 2B in the map** populations Apache/Balance and FD3/Robigus [62] associated with both necrosis and pycnidia resistance in the adult plant stage. However, due to the unavailability of marker sequences, we cannot conclude that Qstb2B_2 derived from ‘Agili39’ is the same locus that was mapped in the aforementioned bread wheat map** populations.
Thus, the identified QTL in ‘Agili39’ co-localized with previously mapped QTL for STB resistance in bread and durum wheat populations, hence we cannot claim a new Stb gene in the ‘Agili39’ landrace accession. However, we clearly have identified QTL conferring resistance to a wide range of Z. tritici isolates under artificial inoculation conditions in seedlings and adult plants, known as field resistance [70].
Thus far, in durum wheat, only partial resistance to Z. tritici was reported [16, 71]. Here, we derived Qstb2B_1 from ‘Agili39’ that provides resistance to five Z. tritici isolates at the seedling stage, and to two isolates at the adult plant stage. Qstb2B_1 explains up to 61.6% of the observed phenotypic variance and was characterized by a high heritability (0.98) with a dual action at the seedling and the adult plant stages. ‘Agili39’ is also the origin of Qstb2B_2, QTL providing a major adult resistance explaining up to 54.3% of the observed phenotypic variance. Our findings confirm that Tunisian durum landraces harbor highly effective Z. tritici resistance QTL.
In fact, the initial screening of the Tunisian landrace accessions showed a remarkable genetic diversity for STB resistance, as claimed also by Ouaja et al. [12] proving that Tunisian durum wheat landraces encompass diverse and valuable sources of resistance to Z. tritici. Eight landrace accessions (Agili37; Agili38; Agili39, Sbei99; Derbessi 12, Mahmoudi101, JK85 and Azizi27) were highly resistant and one landrace showed an intermediate response (‘Agili41’). The different ‘Agili’ landrace accessions reacted differently to the deployed Z. tritici isolates, suggesting a different genetic background, which is in accord with Ferjaoui et al. [72] study hypothesizing that the tested ‘Agili’ accessions most likely carry different Stb genes combinations. Hence, and alike other durum wheat landrace populations [8], the Tunisian durum wheat landraces uncover untapped allelic diversity that is of a great value to support effective breeding strategies to enhance STB resistance in durum wheat.
Our data demonstrated that the broad efficacy of the observed STB resistance in ‘Agili39’ is due to several stacked QTL, both at seedling as well as adult plant stages, which was also commonly observed in inheritance studies in bread wheat [34, 58]. Pyramiding genes for disease resistance has been an effective strategy in preventing boom-and-bust cycles, and is now amenable through marker assisted breeding as a strategy to maintain disease resistance durability, such as for wheat stem rust where various resistance gene combinations have well controlled the disease since the mid-1950s and more recently to the devastating Ug99 race [56, 73,74,75]. A concrete illustration for Z. tritici is the effective resistance to a wide range of isolates in the bread wheat germplasm ‘KK4500’ and ‘TE11’ which is conferred by stacking several known Stb genes [76,77,78] and also in other germplasm several QTL have contributed to broad efficacy of resistance [59]. Our data also confirm that stacking QTL in durum wheat results in broad efficacy of STB resistance. This study has identified genotypes harboring diverse resistance loci entailing dual actions at the different physiological stages constituting thus potential effective sources for Z. tritici resistance and will thus support sustainable breeding approach for Z. tritici resistance in durum wheat.
Finally, we explored QTL epistasis and identified four significant pairwise interactions of the identified QTL with an epistatic QTL mapped on chromosome 5B, designated as Qstb5B. Hence, the epistasis analysis has revealed other QTL that affects the expression of Z. tritici resistance in the ‘Agili39’/Khiar population. In fact, epistatic interactions between QTL are an important factor that affects the phenotypic expression of genes and genetic variation in populations [79,80,81]. Similarly, to many other studies [82,83,84], our data demonstrated interaction between QTL having main effect (Qstb2B_1 and Qstb2B_2) that are involved in epistasis with the Qstb5B QTL affecting the same trait. The epistasis analysis showed an additive-by-additive effect between the Qstb2B_1/ Qstb5B QTL and the Qstb2B_2/Qstb5B QTL, with a major effect of QTL mapped on chromosome 2B (Qstb2B_1 and Qstb2B_2) over the Qstb5B QTL. Interestingly, the epistasis analysis showed that the Qstb2B_2 QTL, proven to control pycnidia development at the adult plant stage by QTL analysis, has also an effect in controlling pycnidia development at the seedling stage when interacting with the epistatic Qstb5B QTL. Nonetheless, the epistasis analysis did not indicate an interaction between the two major QTL Qstb2B_1 and Qstb2B_2 mapped on chromosome 2B.
Conclusion
Our study deciphered ancient broad-based resistance to Z. tritici in a durum wheat landrace accession. A positive selection for the QTL linked markers may result in new high yielding durum wheat cultivars with wide resistance to Z. tritici reminiscent of the durable resistance to STB in landraces. Given the overall high susceptibility to STB in modern durum wheat cultivars, our data shed new light on disease resistance breeding in durum wheat.
Methods
Plant materials and study layout
Eleven durum wheat accessions (Table 8) and a bi-parental recombinant inbred (RIL) population derived from a single seed descent cross between the Tunisian landrace accession ‘Agili39’ and the high yielding commercial cv. Khiar, were screened for resistance to septoria tritici blotch. The RIL population was generated by crossing the resistant parent ‘Agili39’ to the susceptible Khiar following a single seed descent approach (SSD). The F1 plants were selfed to generate the F2 seeds. One head row of each F2 plants were then randomly selected and sown in one row to produce the F2 derived F3 plants (F3). This procedure was followed for all subsequent generations up to the F10 plants. Therecombinant population (Agili39’/Khiar) is maintained and available upon request at the National Institute of Agronomy -Tunisia (INAT) and at the CIMMYT gene banks.
For STB resistance screening, we performed three experiments (Table 2). The first experiment was repeated three times and comprised the screening of the 11 Tunisian landraces at the seedling stage (Z13.3/21) with a panel of 20 Z. tritici isolates (Table 2). The first experiment was performed to understand overall resistance patterns to STB and to select potential isolates for further screening of the RILs derived from the ‘Agili39’/Khiar cross, which consisted of the second experiment (experiment 2), performed thrice at the seedling stage. Finally, in the third experiment we tested the ‘Agili39’/Khiar population under field conditions in Oued-Bejá, located in North-Western Tunisia, over a period of five years, 2011 – 2014 and 2016 with three different Z. tritici isolates.
Screening landraces and RILs population at the seedling stage for resistance to Zymoseptoria tritici
Experimental design, Plants management and growth conditions
A first experiment that consisted of the pre-screening of 11 Tunisian landrace accessions with 20 Z. tritici isolates was conducted at the seedling stage. The pre-screening assay followed a complete block design in three replicates and included the susceptible parent cv. Khiar, the susceptiblecv. Karim and the resistant parent ‘Agili39’ as checks (Table 2 and 8). This experiment enabled the selection of eight Z. tritici isolates that discriminated between the ‘Agili39’ and the cv. Khiar parents and were subsequently used to screen the ‘Agili39’/Khiar derived recombinant inbred lines (RILs) at the seedling stage.
For the seedling assay of the RIL population (experiment 2), we followed a split plot design with isolates as whole plots. Each whole plot consists of three neighbouring trays of fifty-four pots. The tested isolates were randomly allocated to the whole plots. Individual pots were the experimental units, and they were randomly arranged for each isolate/replicate combination on separate parallel greenhouse tables. ‘Replicate’, ‘isolate’, ‘line’ and the ‘isolate x line’ interaction were the fixed terms of our split plot model. However, the random term of the model consists of the ‘replicate x whole plot’ interaction. In all three replicates, eleven checks several checks were included with both parents ‘Agili39’ and cv. Khiar (Table 8).
Five seeds per pot per accessions were grown in VQB 7 × 7x8 cm plastic pots (TEKU®, Lohne, Germany), whereas 157 F6 RILs of the ‘Agili39’/Khiar map** population were planted in round peat pots (Jiffy, Moerdijk, Netherlands), also five seeds per pot, using a special mixture for growing seeds (Substraat Zaai) provided by the greenhouse facility Unifarm of Wageningen University and Research (WUR), The Netherlands.
Zymoseptoria tritici isolates and inoculation procedures
Pre-cultures of each isolate were prepared in an autoclaved 100 ml Erlenmeyer flask containing 50 ml yeast glucose (YG) liquid medium (30 g glucose, 10 g yeast per litre distilled water). The flasks were inoculated with frozen isolate samples that were selected from different durum wheat growing countries and maintained at -80 °C in a Z. tritici isolate collection at WUR. The inoculated flasks were subsequently placed in an incubated rotary shaker (Innova 4430, New Brunswick Scientific, USA) set at 125 rpm and 15 °C for 5–7 days. These pre-cultures were then used to inoculate two 1L Erlenmeyer flasks containing 500 ml YG media per isolate that were incubated under the aforementioned conditions to provide sufficient inoculum for the seedling inoculation assays at growth stage (GS) 11 [85]. Spores were subsequently collected after overnight settling in static cultures, concentrated by decanting the supernatant medium, adjusted to 107 spores ml-1 in a total volume of 40 ml for a set of 18 plastic pots or 24 Jiffy® pots and supplemented with two drops (µl/ml) of Tween 20 surfactant (MERCK®, Nottingham, UK).
Prior to the inoculation, plant development was allowed for 10 days in a greenhouse adjusted at a temperature of 18/16 °C (day/night rhythm) and relative humidity (RH) of 70%. Inoculations were conducted by spraying the inoculum over the seedlings that were placed in an inoculation cabinet on a rotary table, adjusted at 15 rpm, which is equipped with interchangeable atomizers and a water cleaning device to avoid cross- contamination. Infected plants were incubated in transparent plastic bags for 48 h under 100% RH. Post-inoculation conditions were set at a temperature of 22/ ± 2 °C and RH of ≥ 95%. Light intensity (son-T Agro 400 W lamps) and day length (16/8 h light/dark) were similar during pre- and post- inoculation conditions. Ten days after inoculation, seedlings were trimmed for the second and subsequent leaves to enable sufficient light on the inoculated primary leaves for appropriate disease development. Fertilizer (Sporumix PG®, Rotterdam, Netherlands; 0.5 g.l1) was applied to support plant growth.
Disease severity scoring in the seedling assay
In the seedling assays, disease severities were evaluated at 15, 18 and 21 days post-inoculation (dpi). These multiple observations enabled Area Under the Disease Progress Curve (AUDPC) calculations for quantitative analyses of temporal differences in disease progress. We estimated the quantitative presence of necrosis and pycnidia on the inoculated seedling leaves in percentages [36, 37, 86]. AUDPC calculations for seedling scores followed the trapezoidal method, which approximates the time variable and calculates the average disease intensity between each pair of adjacent time points [87].
Screening RILs population at the adult plant stage for resistance to Zymoseptoria tritici
Experimental design and inoculation procedures
Adult plant assays of the ‘Agili39’/Khiar population were conducted at Oued-Bejá, located in North-Western Tunisia (36° 46′ 27.516'' N, 10° 3′ 36.432'' E), for five years, 2011 – 2014 and 2016, with Tun1 – Tun6 and IIB-123 Z. tritici isolates, respectively. This region belongs to the sub-humid bioclimatic zone of Tunisia with an average rainfall ranging from 500 to 850 mm and a daily mean temperature between 10–28 °C [72].
For the field trials, we adopted an augmented randomized complete block design. Five blocks with 1.5 m width and spaced 1.5 m were linearly drilled with 30 to 35 RILs, parents and four modern durum cultivars per block. Each line was sown as one spike per row of 1.5 m length and spaced 25 cm. We randomized all RILs, the parents and four additional checks modern durum wheat cvs. Karim, Nasr, Maali and Salim, important in Tunisian breeding programs and showing different levels of susceptibility in each block. For the 2016 field trial, we used a complete random block design with three replicates with both parents ‘Agili39’ and ‘Khiar’ as checks.
Field inoculations were conducted with three isolates (Tun1, Tun6 and IIB123) across the F5:6-F9:10 RILs generations. We used Z. tritici isolate Tun6 for three years to screen the F5:6 (N = 164), the F7:8 (N = 158) and the F8:9 (N = 157) RILs in 2011, 2013 and 2014, respectively and isolate Tun1 to screen the F6:7 (N = 158) in 2012 and the F8:9 (N = 157) in 2014. In 2016, we screened the F9:10 (N = 155) with Z. tritici isolate IIB123. In all field trials and across all generations, plants were inoculated twice. The first inoculation occurred at the tillering stage (GS 21–26) in order to initiate the disease infection, and the second inoculation was applied after 25–30 days when all RILs reached approximately the elongation stage (GS37) [85]. The second inoculation was applied to ensure an increase in the disease pressure. Field inoculations were conducted with a spore suspension adjusted to 107 spores/ml of the corresponding Z. tritici isolate using a CO2-pressurized knapsack sprayer with a 1 m hand-held boom till run-off.
Disease severity scoring in the field tests
For the field evaluations, we scored pycnidial classes at 28 days post the second inoculation (GS 51) [85] that were later transformed to pycnidia percentages (0 = no pycnidia, 1 = 12%, 2 = 25%, 3 = 50%, 4 = 75% and 5 = 87%) [60, 88].
Genoty** and construction of linkage map
Genomic DNA was extracted from fresh leaves using a modified CTAB (cetyltrimethylammonium bromide) method and quantified using NanoDrop 8000 spectrophotometer V 2.1.0. Whole-genome profiling was performed using DArT-Seq™ technology by Diversity Arrays Technology Pty Ltd, Australia, as described by Kilian et al. [91]. Markers were grouped using a minimum independence LOD (logarithm of the odds) score of 10.0 and linkage groups were established at a minimum LOD score of 3.0. Markers were linearly aligned in each linkage group, converting the recombination rates into the Kosambi's map distance in centimorgans. A sequential map builds up strategy was followed to determine the best marker position [92]. The best fitting position of markers was examined based on the goodness-of-fit test (chi-square) for the resulting map. The final map included bin markers that excluded similar SNP markers.
Linkage groups were subsequently assigned to chromosomes by aligning the SNP marker sequences of the linkage groups to the reference genome of the Triticum turgidum subsp. durum Svevo.v1 and using the BLASTn function in the publicly available sequence database ‘Ensemble Plants’(https://plants.ensembl.org/Triticum_turgidum/Tools/Blast?db=core) [93].
Statistical analyses
Seedling data statistical analyses
Pycnidia AUDPC scores (P-AUDPC) of the pre-screening experiment (experiment 1) were analysed for their variance using the ‘aov’ function in the R environment [42] to test for the effect of ‘line’, the effect of ‘isolate’ and the effect of any ‘line x isolate’ interactions. The following linear model was fitted to the observed P-AUDPC scores:
where Yijk is the P-AUDPC score in the Kth replicate with isolate i and line j. μ is the overall mean. αi is the isolate i main effect. βj is the line j main effect. (αβ)ij is the interaction effect between isolate i and line j and ϵijk is the unexplained error.
For the split-plot seedling experiment performed on the RIL population (experiment 2), an analysis of variance for the necrosis and the pycnidia AUDPC scores (N-AUDPC and P-AUDPC) was performed using the sp.plot function available in the Agricolae package in the R environment [42, 94].Isolates were the whole-plot factors and RILs were the sub-plot factors. We fitted the observed N-AUDPC and P-AUDPC scores to the following linear model:
where Yijk is the N-AUDPC or P-AUDPC score in the Kth replicate of a plot with isolate i and RIL j. μ is the overall mean. αi is the fixed effect of isolate i. ηk(i) is the whole-plot error. βj is the fixed effect of RIL j. (αβ)ij is the interaction effect between isolate i and RIL j and ϵijk is the sub-plot error.
Significant differences between isolates were determined using the least significant difference (LSD) of N-AUDPC and P-AUDPC scores and using the Agricolae package in the R environment. ‘RIL x isolate’ grou** of necrosis and pycnidia AUDPC scores was defined based on the Bonferroni test at p < 0.05. Homogeneity of the seedling replicates was checked and homogeneous data across replications were subsequently averaged and used for the seedling QTL analysis [95].
Repeatability was estimated for the necrosis and pycnidia AUDPC generated at the seedling stage as follow:
where σ2G is the variance component due to genotypes, σ2GE is the variance component due to the interaction between the genotype and the isolate, E is the number of isolates which is eight in the seedling experiment, σ2ε is the variance component due to the unexplained error and r is the number of replicates which is three per isolate for all seedling assays.
Field data statistical analyses
Field pycnidia scores generated by the inoculation of adult plants with Tun1 and Tun6 Z. tritici isolates were tested for their variance to check for the effect of ‘genotype’, ‘block’, ‘year’, ‘isolate’ and for any two-way and three-way interactions between the independent variable ‘genotype’ and the independent factors ‘block’, ‘year’ and ‘isolate’. A linear model was fitted to the observed pycnidia at the adult plant stage as follow:
where Yijkt is the pycnidia coverage score of RIL j inoculated with isolate i in the Kth block at year t. μ is the overall mean. αi is the main effect of the isolate i. βj is the main effect of the RIL j. γk is the main effect of the block k. ηt is the main effect of the year t. (αβ)ij, (γβ)kj, (ηβ)tj are the two-way interaction effects of the RIL j with the isolate i, the RIL j with the block k and the RIL j with the year t, respectively. (αβη)ijt is the three-way interaction effect of RIL j with isolate i and with year t and finally, εijkt is the residual.
For the Tun1 and the Tun6 isolates tested on multiple consecutive years, a broad sense heritability (H2) was estimated as follow:
where σ2G is the variance component due to genotypes. σ2GE is the variance component due to the interaction between the genotype and the isolate. E is the number of years which is 2 and 3 for Tun1 and Tun6 isolates, respectively. σ2ε is the variance component due to the unexplained error and r is the number of blocks which is five for all adult assays.
The least-square means (Lsmeans) of pycnidia coverages were derived from the individual year trials using the SAS software [96]. Transformed field data were subsequently considered for the field QTL analysis.
Correlation between seedling and adult plant assays and frequency distribution
A heatmap correlation matrix was calculated for the generated phenotypes at the seedling and the adult plant stages using the “Corr” function and visualized using the “corrplot” package in the R environment [97] was used. Additive QTL were detected using a walk speed of 1.0 cM and the probability used in stepwise regression for additive QTLs was 0.001. The logarithm of odds (LOD) value of 3.0 was chosen to declare significant QTL, and the LOD value was calculated from 1000 permutations with type I error of 0.01. The phenotypic variance explained (PVE) and additive effect of individual QTL at the LOD peaks were also obtained. Identified QTL were plotted against their corresponding linkage groups using the MapChart © software version 2.3 [98]. Subsequently and for the identified QTL contributing to resistance, we aligned the linked SNP markers to the reference genome of the Triticum turgidum subsp. durum Svevo.v1 and using the BLASTn function (https://plants.ensembl.org/Triticum_turgidum/Tools/Blast?db=core) in the publicly available sequence database ‘Ensemble Plants’ [93].
Epistatic interactions between QTL were identified by the inclusive composite interval map** of digenic epistatic QTL (ICIM-EPI) method implemented in QTL IciMap** software v4.1 [97]. The LOD threshold was set at 5.00 to declare significant epistatic QTL and LOD value was calculated from 1000 permutations at the significance level of 0.05.
The highest epistasis interactions detected between the QTL (highest LOD) were subsequently selected for a two-way interaction test to examine pycnidia AUDPC means of the variant allele combinations linked to the epistatic QTL. Alleles linked to the detected QTL were named by ‘R’ and ‘S’ denoting the resistant and susceptible alleles, respectively. Hence, pycnidia AUDPC means of the four allele combinations ‘RR’, ‘RS’, ‘SR’ and ‘SS’ were examined and Boxplots were generated using the ‘ggplot2’ package in the R environment [99].
Availability of data and materials
The used plant materials are available at the CIMMYT gene bank upon a reasonable request to the co-author Karim Ammar < k.ammar@cgiar.org >
The sequencing raw data files generated during this study are uploaded to the CIMMYT data repository (https://hdl.handle.net/11529/10548618). The analyzed data are available as additional files to this article.
References
Arzani A, Ashraf M. Cultivated Ancient Wheats (Triticum spp): A Potential Source of Health-Beneficial Food Products. Compr Rev Food Sci Food Safety. 2017;16(3):477–88.
FAOSTAT. 2017. http://www.fao.org/faostat/en/#compare
Shiferaw B, Smale M, Braun H-J, Duveiller E, Reynolds M, Muricho G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Sec. 2013;5(3):291–317.
Charmet G. Wheat domestication: lessons for the future. CR Biol. 2011;334(3):212–20.
Palamarchuk A. Selection strategies for traits relevant for winter and facultative durum wheat. Durum wheat breeding: current approaches and future strategies. New York: Food Products Press; 2005. p. 599–644.
Royo C, Nazco R, Villegas D. The climate of the zone of origin of Mediterranean durum wheat (Triticum durum Desf) landraces affects their agronomic performance. Genet Resour Crop Evol. 2014;61(7):1345–58.
Negisho K, Shibru S, Pillen K, Ordon F, Wehner G. Genetic diversity of Ethiopian durum wheat landraces. PLoS ONE. 2021;16(2):e0247016.
Kidane YG, Hailemariam BN, Mengistu DK, Fadda C, Pè ME, Dell'Acqua M. Genome-Wide Association Study of Septoria tritici Blotch Resistance in Ethiopian Durum Wheat Landraces. Front Plant Sci. 2017;8:1586. https://doi.org/10.3389/fpls.2017.01586.
Fabricant F. Rome’s glory is now Tunisia’s. New York Times 1998. https://www.nytimes.com.
Oliveira HR, Campana MG, Jones H, Hunt HV, Leigh F, Redhouse DI, Lister DL, Jones MK. Tetraploid wheat landraces in the Mediterranean basin: taxonomy, evolution and genetic diversity. PLoS ONE. 2012;7(5):e37063.
Davis DK: Resurrecting the granary of Rome: environmental history and French colonial expansion in North Africa: Ohio University Press; 2007.
Ouaja M, Aouini L, Bahri B, Ferjaoui S, Medini M, Marcel TC, Hamza S. Identification of valuable sources of resistance to Zymoseptoria tritici in the Tunisian durum wheat landraces. Eur J Plant Pathol. 2020;156(2):647–61.
Kamel S, Cherif M: Tan spot of wheat in Northern Tunisia: distribution, prevalence, incidence and severity. Cereal Research Communications 2021.
McDonald BA, Mundt CC. How knowledge of pathogen population biology informs management of Septoria tritici blotch. Phytopathology. 2016;106(9):948–55.
Abdedayem W. M’barek S, Souissi A, Laribi M, Araar C, Kouki H, Fakhfakh M, Yahyaoui A: Septoria tritici blotch disease progression and physiological traits variation in durum wheat variety mixtures. J New Sci. 2021;80:4664–74.
Berraies S, Ammar K, Salah Gharbi M, Yahyaoui A, Rezgui S. Quantitative inheritance of resistance to Septoria tritici blotch in durum wheat in Tunisia. Chilean J Agri Res. 2014;74(1):35–40.
Hamada W. First isolation of the Mycosphaerella graminicola teleomorph stage causing Septoria leaf blotch on wheat in Tunisia. New Dis Rep. 2014;29:18–18.
Meamiche Neddaf H, Aouini L, Bouznad Z, Kema GH. Equal distribution of mating type alleles and the presence of strobilurin resistance in algerian zymoseptoria tritici field populations. Plant Dis. 2017;101(4):544–9.
Hassine M, Siah A, Hellin P, Cadalen T, Halama P, Hilbert J-L, Hamada W, Baraket M, Yahyaoui A, Legrève A. Sexual reproduction of Zymoseptoria tritici on durum wheat in Tunisia revealed by presence of airborne inoculum, fruiting bodies and high levels of genetic diversity. Fungal Biol. 2019;123(10):763–72.
Boukef S, McDonald BA, Yahyaoui A, Rezgui S, Brunner PC. Frequency of mutations associated with fungicide resistance and population structure of Mycosphaerella graminicola in Tunisia. Eur J Plant Pathol. 2012;132(1):111–22.
Singh RP, Herrera-Foessel S, Huerta-Espino J, Singh S, Bhavani S, Lan C, Basnet BR. Progress towards genetics and breeding for minor genes based resistance to Ug99 and other rusts in CIMMYT high-yielding spring wheat. J Integr Agric. 2014;13(2):255–61.
Singh R, Huerta-Espino J, Bhavani S, Herrera-Foessel S, Singh D, Singh P, Velu G, Mason R, ** Y, Njau P. Race non-specific resistance to rust diseases in CIMMYT spring wheats. Euphytica. 2011;179(1):175–86.
Saintenac C, Zhang W, Salcedo A, Rouse MN, Trick HN, Akhunov E, Dubcovsky J. Identification of wheat gene Sr35 that confers resistance to Ug99 stem rust race group. Science. 2013;341(6147):783–6.
Chen S, Rouse MN, Zhang W, ** Y, Akhunov E, Wei Y, Dubcovsky J. Fine map** and characterization of Sr21, a temperature-sensitive diploid wheat resistance gene effective against the Puccinia graminis f. sp. tritici Ug99 race group. Theor Appl Genet. 2015;128(4):645–56.
Bajgain P, Rouse M, Bulli P, Bhavani S, Gordon T, Wanyera R, Njau P, Legesse W, Anderson J, Pumphrey M. Erratum to: Association map** of North American spring wheat breeding germplasm reveals loci conferring resistance to Ug99 and other African stem rust races. BMC Plant Biol. 2016;16(1):1–9.
Kingsbury N. Hybrid: the history and science of plant breeding: University of Chicago Press; 2009. p. 512.
Mondal S, Rutkoski JE, Velu G, Singh PK, Crespo-Herrera LA, Guzmán C, Bhavani S, Lan C, He X, Singh RP. Harnessing diversity in wheat to enhance grain yield, climate resilience, disease and insect pest resistance and nutrition through conventional and modern breeding approaches. Front Plant Sci. 2016;7:991. https://doi.org/10.3389/fpls.2016.00991.
Lopes MS, El-Basyoni I, Baenziger PS, Singh S, Royo C, Ozbek K, Aktas H, Ozer E, Ozdemir F, Manickavelu A: Exploiting genetic diversity from landraces in wheat breeding for adaptation to climate change. J Exp Bot. 2015;66(12):3477–86. https://doi.org/10.1093/jxb/erv122.
Aoun M, Kolmer JA, Rouse MN, Elias EM, Breiland M, Bulbula WD, Chao S, Acevedo M. Map** of novel leaf rust and stem rust resistance genes in the Portuguese durum wheat landrace PI 192051. G3: Genes, Genomes, Genet. 2019;9(8):2535–47.
Alemu SK, Huluka AB, Tesfaye K, Haileselassie T, Uauy C. Genome-wide association map** identifies yellow rust resistance loci in Ethiopian durum wheat germplasm. PLoS ONE. 2021;16(5):e0243675.
Royo C, Ammar K, Soriano JM, Villegas D. Agronomic, physiological and genetic changes associated with evolution, migration and modern breeding in durum wheat. Front Plant Sci. 2021;12:1318.
Ghaneie A, Mehrabi R, Safaie N, Abrinbana M, Saidi A, Aghaee M. Genetic variation for resistance to septoria tritici blotch in Iranian tetraploid wheat landraces. Eur J Plant Pathol. 2012;132(2):191–202.
Porras R, Pérez-de-Luque A, Sillero J, Miguel-Rojas C. Behavior of Spanish durum wheat genotypes against Zymoseptoria tritici: resistance and susceptibility. Span J Agric Res. 2021;19(3): e1002.
Brown JK, Chartrain L, Lasserre-Zuber P, Saintenac C. Genetics of resistance to Zymoseptoria tritici and applications to wheat breeding. Fungal Genetic Biololgy. 2015;79:33–41.
Yang N, McDonald MC, Solomon PS, Milgate AW. Genetic map** of Stb19, a new resistance gene to Zymoseptoria tritici in wheat. Theor Appl Genet. 2018;131(12):2765–73.
Kema GH, van Silfhout CH. Genetic variation for virulence and resistance in the wheat-Mycosphaerella graminicola pathosystem III. Comparative seedling and adult plant experiments. Phytopathology. 1997;87(3):266–72.
Kema G, Annone JG, Sayoud R, Van Silfhout CH, Van Ginkel M, De Bree J. Genetic variation for virulence and resistance in the wheat-Mycosphaerella graminicola pathosystem I. Interactions between pathogen isolates and host cultivars. Phytopathology. 1996;86(2):200–12.
Kema G, Annone J, Sayoud R, Van Silfhout C, Van Ginkel M, De Bree J. Genetic variation for virulence and resistance in the wheat-Mycosphaerella graminicola pathosystem. I: Interactions between pathogen isolates and host cultivars. Phytopathology. 1996;86(2):200–12.
Tabib Ghaffary SM. Efficacy and map** of resistance to Mycosphaerella graminicola in wheat. PhD dissertation Wageningen University; 2011. https://edepot.wur.nl/169465.
Ballini E, Tavaud M, Ducasse A, Sanchez D, Paux E, Kitt J, Charmet G, Audigeos D, Roumet P, David J. Genome wide association map** for resistance to multiple fungal pathogens in a panel issued from a broad composite cross-population of tetraploid wheat Triticum turgidum. Euphytica. 2020;216:1–17.
El Haddad N, Kabbaj H, Zaïm M, El Hassouni K, Tidiane Sall A, Azouz M, Ortiz R, Baum M, Amri A, Gamba F. Crop wild relatives in durum wheat breeding: Drift or thrift? Crop Sci. 2021;61(1):37–54.
Team RC: R. A language and environment for statistical computing. 2013.
Wei T, Simko V, Levy M, **e Y, ** Y, Zemla J. Package ‘corrplot.’ Statistician. 2017;56:316–24.
Fones H, Gurr S. The impact of Septoria tritici Blotch disease on wheat: An EU perspective. Fungal Genet Biol. 2015;79:3–7.
Aouini L. Durum wheat and septoria tritici blotch: genes and prospects for breeding. PhD dissertation Wageningen University 2018. https://doi.org/10.18174/443719.
Arraiano L, Brading P, Brown J. A detached seedling leaf technique to study resistance to Mycosphaerella graminicola (anamorph Septoria tritici) in wheat. Plant Pathol. 2001;50(3):339–46.
Berraies S, Gharbi MS, Belzile F, Yahyaoui A, Hajlaoui MR, Trifi M, Jean M, Rezgui S. High genetic diversity of Mycospaherella graminicola (Zymoseptoria tritici) from a single wheat field in Tunisia as revealed by SSR markers. Afr J Biotechnol. 2013;12(12):1344–9.
Kema GHJ, Mirzadi Gohari A, Aouini L, Gibriel HAY, Ware SB, van den Bosch F, Manning-Smith R, Alonso-Chavez V, Helps J, Ben M’Barek S, et al. Stress and sexual reproduction affect the dynamics of the wheat pathogen effector AvrStb6 and strobilurin resistance. Nat Genet. 2018;50(3):375–80.
Zhong Z, Marcel TC, Hartmann FE, Ma X, Plissonneau C, Zala M, Ducasse A, Confais J, Compain J, Lapalu N. A small secreted protein in Zymoseptoria tritici is responsible for avirulence on wheat cultivars carrying the Stb6 resistance gene. New Phytol. 2017;214(2):619–31.
Saintenac C, Lee W-S, Cambon F, Rudd JJ, King RC, Marande W, Powers SJ, Bergès H, Phillips AL, Uauy C, et al. Wheat receptor-kinase-like protein Stb6 controls gene-for-gene resistance to fungal pathogen Zymoseptoria tritici. Nat Genet. 2018;50(3):368–74.
Lin Y, Gnanesh BN, Chong J, Chen G, Beattie AD, Mitchell Fetch JW, Kutcher HR, Eckstein PE, Menzies JG, Jackson EW. A major quantitative trait locus conferring adult plant partial resistance to crown rust in oat. BMC Plant Biol. 2014;14(1):250.
Hou L, Chen X, Wang M, See DR, Chao S, Bulli P, **g J. Map** a Large Number of QTL for Durable Resistance to Stripe Rust in Winter Wheat Druchamp Using SSR and SNP Markers. PLoS ONE. 2015;10(5):e0126794.
Liu Y, Qie Y, Li X, Wang M, Chen X. Genome-Wide Map** of Quantitative Trait Loci Conferring All-Stage and High-Temperature Adult-Plant Resistance to Stripe Rust in Spring Wheat Landrace PI 181410. Int J Mol Sci. 2020;21(2):478.
Tahir S, Zia I, Dilshad I, Fayyaz M, Noureen N, Farrakh S. Identification of stripe rust resistant genes and their validation in seedling and adult plant glass house tests. Genet Resour Crop Evol. 2020;67(4):1025–36.
Han G, Liu S, Wang J, ** Y, Zhou Y, Luo Q, Liu H, Zhao H, An D. Identification of an elite wheat-rye T1RS· 1BL translocation line conferring high resistance to powdery mildew and stripe rust. Plant Dis. 2020. https://doi.org/10.1094/PDIS-02-20-0323-RE.
Prasad P, Savadi S, Bhardwaj SC, Gupta PK. The progress of leaf rust research in wheat. Fungal Biol. 2020;124(6):537–50. https://doi.org/10.1016/j.funbio.2020.02.013.
McCartney C, Brûlé-Babel A, Lamari L, Somers D. Chromosomal location of a race-specific resistance gene to Mycosphaerella graminicola in the spring wheat ST6. Theor Appl Genet. 2003;107(7):1181–6.
Ghaffary SMT, Faris JD, Friesen TL, Visser RG, van der Lee TA, Robert O, Kema GH. New broad-spectrum resistance to septoria tritici blotch derived from synthetic hexaploid wheat. Theor Appl Genet. 2012;124(1):125–42.
Ghaffary SMT, Robert O, Laurent V, Lonnet P, Margalé E, van der Lee TA, Visser RG, Kema GH. Genetic analysis of resistance to septoria tritici blotch in the French winter wheat cultivars Balance and Apache. Theor Appl Genet. 2011;123(5):741–54.
Adhikari TB, Cavaletto JR, Dubcovsky J, Gieco JO, Schlatter AR, Goodwin SB. Molecular map** of the Stb4 gene for resistance to septoria tritici blotch in wheat. Phytopathology. 2004;94(11):1198–206.
Arraiano LS, Worland AJ, Ellerbrook C, Brown JKM. Chromosomal location of a gene for resistance to septoria tritici blotch (Mycosphaerella graminicola)in the hexaploid wheat ’Synthetic 6x’. Theor Appl Genet. 2001;103(5):758–64.
Goudemand E, Laurent V, Duchalais L, Tabib Ghaffary S, Kema G, Lonnet P, Margalé E, Robert O: Association map** and meta-analysis: two complementary approaches for the detection of reliable Septoria tritici blotch quantitative resistance in bread wheat (Triticum aestivum L.). Mol Breeding 2013, 32(3).
Risser P, Ebmeyer E, Korzun V, Hartl L, Miedaner T. Quantitative trait loci for adult-plant resistance to Mycosphaerella graminicola in two winter wheat populations. Phytopathology. 2011;101(10):1209–16.
Goodwin SB, Cavaletto JR, Hale IL, Thompson I, Xu SS, Adhikari TB, Dubcovsky J. A New Map Location of Gene for Resistance to Septoria Tritici Blotch in Wheat. Crop Sci. 2015;55(1):35–43.
Adhikari TB, Wallwork H, Goodwin SB. Microsatellite Markers Linked to the and Genes for Resistance to Septoria Tritici Blotch in Wheat. Crop Sci. 2004;44(4):1403–11.
Chartrain L, Sourdille P, Bernard M, Brown J. Identification and location of Stb9, a gene for resistance to septoria tritici blotch in wheat cultivars Courtot and Tonic. Plant Pathol. 2009;58(3):547–55.
Odilbekov F, Armoniené R, Koc A, Svensson J, Chawade A. GWAS-Assisted Genomic Prediction to Predict Resistance to Septoria Tritici Blotch in Nordic Winter Wheat at Seedling Stage. Front Genet. 2019;10:1224.
Radecka-Janusik M, Czembor PC. Genetic map** of quantitative trait loci (QTL) for resistance to Septoria tritici blotch in a winter wheat cultivar Liwilla. Euphytica. 2014;200(1):109–25.
Stadlmeier M, Hartl L, Mohler V. Usefulness of a multiparent advanced generation intercross population with a greatly reduced mating design for genetic studies in winter wheat. Front Plant Sci. 1825;2018:9.
Arraiano L, Balaam N, Fenwick P, Chapman C, Feuerhelm D, Howell P, Smith S, Widdowson J, Brown J. Contributions of disease resistance and escape to the control of Septoria tritici blotch of wheat. Plant Pathol. 2009;58(5):910–22.
Tuberosa R: Map** QTLs for Partial Resistance to Zymoseptoria tritici in Durum Wheat. In: Plant and Animal Genome XXII Conference: 2014: Plant and Animal Genome; 2014.
Ferjaoui S, M’Barek S, Bahri B, Slimane R, Hamza S. Identification of resistance sources to septoria tritici blotch in old Tunisian durum wheat germplasm appliad for the analysis of the Zymoseptoria tritici-durum wheat interaction. J Plant Pathol. 2015;97(3):471–81.
Singh RP, Hodson DP, Huerta-Espino J, ** Y, Bhavani S, Njau P, Herrera-Foessel S, Singh PK, Singh S, Govindan V. The emergence of Ug99 races of the stem rust fungus is a threat to world wheat production. Annu Rev Phytopathol. 2011;49:465–81.
Mundt CC. Durable resistance: a key to sustainable management of pathogens and pests. Infect Genet Evol. 2014;27:446–55.
Gautam T, Dhillon GS, Saripalli G, Singh VP, Prasad P, Kaur S, Chhuneja P, Sharma P, Balyan H, Gupta P. Marker-assisted pyramiding of genes/QTL for grain quality and rust resistance in wheat (Triticum aestivum L.). Mol Breeding. 2020;40:1–14.
Chartrain L, Brading P, Brown J. Presence of the Stb6 gene for resistance to Septoria tritici blotch (Mycosphaerella graminicola) in cultivars used in wheat-breeding programmes worldwide. Plant Pathol. 2005;54(2):134–43.
Chartrain L, Berry S, Brown J. Resistance of wheat line Kavkaz-K4500 L. 6. A. 4 to Septoria tritici blotch controlled by isolate-specific resistance genes. Phytopathology. 2005;95(6):664–71.
Chartrain L, Brading P, Widdowson J, Brown J. Partial resistance to Septoria tritici blotch (Mycosphaerella graminicola) in wheat cultivars Arina and Riband. Phytopathology. 2004;94(5):497–504.
Li ZK, Jiang XL, Peng T, Shi CL, Han SX, Tian B, Zhu ZL, Tian JC. Map** quantitative trait loci with additive effects and additive x additive epistatic interactions for biomass yield, grain yield, and straw yield using a doubled haploid population of wheat (Triticum aestivum L.). Genet Mol Res. 2014;13(1):1412-24. https://doi.org/10.4238/2014.
Farokhzadeh S, Fakheri BA, Nezhad NM, Tahmasebi S, Mirsoleimani A, McIntyre CL. Genetic control of some plant growth characteristics of bread wheat (Triticum aestivum L.) under aluminum stress. Genes Genom. 2020;42(3):245–61.
Li Z, Pinson S, Stansel J, Park W. Identification of quantitative trait loci (QTLs) for heading date and plant height in cultivated rice (Oryza sativa L.). Theor Appl Genet. 1995;91(2):374–81.
Lecomte L, Duffé P, Buret M, Servin B, Causse M. Marker-assisted introgression of five QTLs controlling fruit quality traits into three tomato lines revealed interactions between QTLs and genetic backgrounds. Theor Appl Genet. 2004;109(3):658–68.
Wang D, Zhu J, Li Z, Paterson A. Map** QTLs with epistatic effects and QTL× environment interactions by mixed linear model approaches. Theor Appl Genet. 1999;99(7–8):1255–64.
Lin H, Yamamoto T, Sasaki T, Yano M. Characterization and detection of epistatic interactions of 3 QTLs, Hd1, Hd2, and Hd3, controlling heading date in rice using nearly isogenic lines. Theor Appl Genet. 2000;101(7):1021–8.
Zadoks JC, Chang TT, Konzak CF. A decimal code for the growth stages of cereals. Weed Res. 1974;14(6):415–21.
Kema G, Sayoud R, Annone J, Van Silfhout C. Genetic variation for virulence and resistance in the wheat-Mycosphaerella graminicola pathosystem. II: analysis of interactions between pathogen isolates and host cultivars. Phytopathology. 1996;86(2):213–20.
Madden LV, Hughes G, Bosch F. The study of plant disease epidemics: American Phytopathological Society (APS Press); 2007. https://doi.org/10.1094/9780890545058.
Eyal Z, Brown MB. A quantitative method for estimating density of Septaria tritici pycnidia on wheat leaves. Phytopathology. 1976;66:11–4.
Kilian A, Wenzl P, Huttner E, Carling J, **a L, Blois H, Caig V, Heller-Uszynska K, Jaccoud D, Hopper C, Aschenbrenner-Kilian M, Evers M, Peng K, Cayla C, Hok P, Uszynski G. Diversity arrays technology: a generic genome profiling technology on open platforms. Methods Mol Biol. 2012;888:67–89. https://doi.org/10.1007/978-1-61779-870-2_5.
Van Ooijen J. JoinMap 4. Software for the calculation of genetic linkage maps in experimental populations Kyazma BV, Wageningen, Netherlands: Scietific research; 2006. p. 33.
Kosambi DD. The estimation of map distances from recombination values. Ann Eugenic. 1943;12(1):172–5. https://doi.org/10.1111/j.1469-1809.1943.tb02321.x.
Stam P. Construction of integrated genetic linkage maps by means of a new computer package: Join Map. Plant J. 1993;3(5):739–44.
Maccaferri M, Harris NS, Twardziok SO, Pasam RK, Gundlach H, Spannagl M, Ormanbekova D, Lux T, Prade VM, Milner SG, et al. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat Genet. 2019;51(5):885–95.
De Mendiburu F, Simon R: Agricolae-Ten years of an open source statistical tool for experiments in breeding, agriculture and biology. In.: PeerJ PrePrints; 2015.
Chu C-G, Chao S, Friesen T, Faris J, Zhong S, Xu S. Identification of novel tan spot resistance QTLs using an SSR-based linkage map of tetraploid wheat. Mol Breeding. 2010;25(2):327–38.
Institute S: Base SAS 9.4 procedures guide: SAS Institute; 2015.
Meng L, Li H, Zhang L, Wang J. QTL IciMap**: integrated software for genetic linkage map construction and quantitative trait locus map** in biparental populations. Crop J. 2015;3(3):269–83.
Voorrips R. MapChart: software for the graphical presentation of linkage maps and QTLs. J Hered. 2002;93(1):77–8.
Wickham H. Elegant graphics for data analysis. Media. 2009;35(211):10.1007.
Acknowledgements
We would like to thank Els Verstappen and all members of Bio-interaction and Plant health (Wageningen University and Research) for excellent technical support, Nicolas Letreux and Bertus Van der Laan (Unifarm, Wageningen University) for greenhouse assistance and maintenance. We, also would like to thank all members of the Genetics and Plant Breeding group at the National Institute of Agronomy, Tunisia , and staff members at the Regional Field Crops and Research Center—Beja (Tunisia) for field assistance, and CIMMYT staff members (Mexico) for genoty** support. We, finally, highly acknowledge Paul Keizer and Pieter Vereijken (Biometris, Wageningen University and Research) for providing statistical support.
Funding
The research was partly supported by the Japan International Cooperation Agency (JICA)–Japan Science and Technology Agency (JST)’s Science and Technology Research Partnership for Sustainable Development (SATREPS) under the project entitled “Valorization of Bio-resources in Semi-Arid and Arid Land for Regional Development”.
LA was financially supported by the Monsanto's Beachell-Borlaug International Scholars Program (3,340,030,501) and The ‘Bourse d’alternance’ from the Ministry of Higher Education and Scientific Research—Tunisia.
Author information
Authors and Affiliations
Contributions
GHJK and SH designed the study. SF produced the ‘Agili39’/Khiar RIL population,provided the plant materials, and generated the field phenotypic data. LA generated the seedling phenotypic data. KA and SD genotyped the ‘Agili39’/Khiar map** population. HJS, LA and RBS generated the linkage genetic map. LA performed the statistical analysis. BB and SS performed the QTL and the epistasis analyses. LA, SH and SF wrote the manuscript. All authors read and reviewed the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The used plant material comprises either registered Triticum durum cultivars and landrace accessions or experimental lines developed by the corresponding author Professor Sonia Hamza at the National Institute of Agronomy (INAT) and by Dr. Sahbi Ferjaoui at the Regional Field Crops Research Center of Beja (CRRGC). Experimental research and field studies, including the collection of plant material complies with relevant institutional, national and international guidelines and legislation.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Fig. S1.
Frequency distributions of the disease severity assessed as percentage pycnidia in seedlings of the F6 recombinant inbred lines of the ‘Agili39’/Khiar population. ‘A’ and ‘K’ are referring to the ‘Agili39’and cv. Khiar parents, respectively.
Additional file 2: Table 1.
Analysis of variance of pycnidia percent of nine Tunisian durum landraces and two modern varieties inoculated with a diverse range of twenty durum derived Zymoseptoria tritici isolates. Table 2. Pearson correlation between the different tested isolates on the 'Agili39'/khiar population RILs at the seedling and the Adult plant stages. Table 3. Linkage groups, correspondent durum wheat chromosome, the average length indicated in centimorgans (cM), number of SNPs and the average inter-loci distance (cM) in the 'Agili39'/khiar genetic linkage map. Table 4. Sequences, genetic and physical positions of the flanking markers linked to the detected QTL on the 'Agili39'/Khiar map** population.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ferjaoui, S., Aouini, L., Slimane, R.B. et al. Deciphering resistance to Zymoseptoria tritici in the Tunisian durum wheat landrace accession ‘Agili39’. BMC Genomics 23, 372 (2022). https://doi.org/10.1186/s12864-022-08560-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-022-08560-2