
Abstract
Background
C-reactive protein (CRP) is an acute phase protein that can activate various immune cells and bind to certain Fcγ receptors. The latter may compete with the binding of IgG antibodies to these receptors and could thereby interfere with the antigen-specific immune response. Polymorphisms in the promoter region of the CRP gene have been strongly associated with the plasma concentration of CRP. The known lower susceptibility to malaria in the Fulani ethnic group, as compared to their sympatric neighbours in Africa, has been linked to different genetic backgrounds. The present study was performed to investigate if polymorphisms in the CRP gene could contribute to the lower susceptibility to malaria seen in the Fulani ethnic group.
Methods
The CRP -717 T>C, -286 C>T>A, and +1444 C>T polymorphisms were analysed in asymptomatic Fulani and non-Fulani individuals from Mali and Sudan using Pyrosequencing T and TaqMan r MGB probes.
Results
The rare -286 A allele, previously shown to be associated with increased CRP expression and plasma levels, was shown to be more frequent in the non-Fulani ethnic groups as compared to the sympatric Fulani ethnic group both in Mali and Sudan. The common -717 T allele was more prevalent in the non-Fulani ethnic group compared to the sympatric Fulani ethnic group, but only in Mali. The parasite prevalence was increased for the -286 A allele, but not for the -717 T allele. No differences regarding genotype frequency or parasite prevalence were seen for +1444 C>T.
Conclusion
This study indicate that CRP may play an important role in the immune responses to malaria, and that the -286 C/T/A CRP polymorphism may be a contributing factor to the lower susceptibility to malaria seen in the Fulani.
Similar content being viewed by others


Background
Malaria, caused by parasites from the Plasmodium genus, is a prevalent infectious disease worldwide. The Fulani ethnic group in Africa has shown a lower susceptibility to malaria as compared to their sympatric neighbours, despite being similarly exposed to malaria and having no obvious differences in socio-cultural circumstances. The Fulani have lower parasite rates and parasitaemia, higher anti-malarial immune responses and higher spleen rates than other sympatric groups [1–3]. It has been previously shown that this relative resistance to malaria in the Fulani is pathogen related and not caused by a higher general reactivity of the immune system [4]. A number of studies have reported differences between Fulani and non-Fulani groups in polymorphic immune-related genes [5, 6]. Recently, such an inter-ethnic difference in the allele frequencies of a polymorphism in the gene encoding Fca receptor (R) IIa, resulting in an arginine (R)/histidine (H)-131 receptor diversity was demonstrated [7, 8].
C-reactive protein (CRP) is an acute phase protein, the levels of which increase rapidly in the circulation during infection and/or inflammation. CRP has the ability to activate various immune cells and has also shown a possible capacity to bind to certain FcγR [9]. Fc receptors are widely expressed on leukocytes and they are important players in the immune response, the binding of antibodies to these receptors leading to activation and onset of many important biological events [10]. Interestingly, CRP shows a sequence homology to the regions in IgG that are important for FcγR binding [9], indicating that the two proteins may bind to a common site on these receptors. The binding of CRP may, therefore, compete with the binding of IgG antibodies to these receptors, and could thereby interfere with the antigen-specific immune response. The FcγRIIa has been shown to have a higher affinity for CRP if the R131 allele of the receptor is present [11], the same allele that in some studies has been related to protection against malaria [12]. Previous studies on falciparum malaria have related high circulating CRP levels with parasite density and severity of the malaria infection [13–15]. Interestingly, in individuals having an asymptomatic Plasmodium falciparum infection, the plasma levels of CRP are low [16, 17], suggesting that the increased CRP levels found in P. falciparum symptomatic individuals are not merely an acute phase response. The role of CRP in P. falciparum infections is not clear. CRP has been associated with complement mediated haemolysis of infected erythrocytes and subsequent anaemia [18], but CRP has also been implicated in the defence against pre-erythrocytic stages of malaria [19]. Importantly, CRP induces the anti-inflammatory interleukin (IL)-10 [20], which could affect the early immune response seen in malaria infections. The influence of genetic variations on the levels of circulating CRP is estimated to be 40–60% [21], and single nucleotide polymorphisms (SNPs) in different cytokine genes [22, 23] and the CRP gene [24] have been suggested to be associated with the circulating levels of CRP. The strongest association with the circulating concentrations of CRP has been shown for the tri-allelic -286 SNP [25, 26], and it has been suggested that this is a truly functional polymorphism in the CRP gene [24]. Previous studies of the -286 C>T>A CRP SNP have revealed that the A allele is more common in African American than in Caucasian populations, and that this difference in genotypes is associated with higher CRP levels in the former population [27]. This suggests that CRP could have been selected for in the African ancestors, as a factor beneficial for survival from infectious diseases.
In order to investigate the possible involvement of CRP in innate immunity against malaria, and to further define potential immunological factors contributing to the ethnic differences in malaria susceptibility, the -717 T>C (rs2794521), -286 C>T>A (rs3091244,) +1444 C>T (rs1130864) CRP genotypes and malariometric indices were analysed in individuals of sympatric ethnic groups from two countries, Mali and Sudan, with markedly different malaria endemicity.
Methods
Study areas
The study area in Mali is located in the Mopti area about 850 km Northeast of Bamako, the capital of Mali. Four villages, Mantéourou, Naye, Binédama, and Anakédié were identified for the study. Malaria transmission is mesoendemic in the area, with P. falciparum as the main parasite species. The entomological inoculation rate is similar in both ethnic groups [3]. In this area, the dry season extends from October to May and the rainy season from July to October.
The study area in Sudan is located in the Daraweesh village in the Gedaref State in eastern Sudan, 450 km from Khartoum and 16 km from the Gedaref town. Malaria transmission is markedly seasonal and unstable, and annual peak parasite prevalence ranges from 1 to 40% in different years, and P. falciparum is responsible for >96% of the malaria cases [28].
Human samples
In Mali, blood was collected during the rainy season in September 2005 from 166 Fulani (age range 1–60 years, median age 15 years, 51% males) and 608 non-Fulani represented by the Dogon (98%) and Rimaibé (2%) ethnic groups (age range 7 month – 61 years, median age 8 years, 44% males). All individuals were asymptomatic at the time of collection, and parasite prevalence was detected by microscopy. For PCR, finger prick blood was collected on filter papers. Spleen enlargement was assessed by palpation and graded according to the Hackett score (0–5). The proportion of individuals that had enlarged spleens was denoted spleen rate. Informed consent was obtained from all participants or their guardians and the Ethical committee of the Faculty of Medicine and Pharmacy, University of Bamako, in Mali and the National Ethics Committee in Sweden approved the study.
In Sudan, blood was collected, as part of a longitudinal study, in May before the rainy seasons between 2004 and 2006, from 225 Fulani (age range 4–75 years, median age 15 years, 36% males) and 100 non-Fulani represented by the Four (22%), Hausa (28%) and Masaleit (50%) ethnic groups (age range 2–55 years, median age 17 years, 41% males). All the individuals were asymptomatic and parasite negative by microscopy at the time of collection. Three ml of peripheral blood were collected from all individuals into vacuum EDTA tubes. The study received ethical approval from the ethical Committee of University of Khartoum and national clearance from the Sudanese Ministry of Health.
DNA preparation
DNA was extracted from filter papers and venous blood with Chelex-100 as described previously [5], and then stored at -20°C. In brief, discs from filter paper or 25 μl from peripheral blood were incubated overnight in 1 ml of 0.5% saponin in PBS at 4°C, and were then washed 15–30 minutes in 1 ml PBS at 4°C. The discs or the pellets, were then boiled in 200 μl of 5% Chelex-100 in water for 15 minutes, and subsequently DNA was collected in the supernatants after centrifugation at 8,000 × g for three minutes.
CRP genoty**
The -286 C>T>A CRP polymorphism (rs3091244) was analysed using the pyrosequencing method. PCR primers were designed to amplify the -286 C>T>A polymorphism, using the forward primer 5'-TGT TTT CCT CAT TTC CCA GTC T-3' and the reverse primer 5'-biotinylated TGG CTA TCT ATC CTG CGA AAA T-3' (MWG-Biotech AG, Ebersberg, Germany). The PCR amplification was performed in a 40 μl reaction using 8 μl of genomic DNA template, 20 μl iProof T High-Fidelity Master Mix (BIO-RAD Laboratories, Hercules, CA), and 1.5 μl of each primer (10 pmol/μl). The PCR was carried out in an Eppendorf Mastercyclerr (Eppendorf AG, Hamburg, Germany) using a 30 sec denaturation at 98°C, followed by 35 cycles with 98°C for 10 sec, 59°C for 20 sec and 72°C for 15 sec. The final extension was at 72°C for 5 min. The sequencing primer was 5'-GTG CAC CCA GAT GGC-3' (MWG-Biotech AG), and the nucleotide dispensation order was GCA CGT ACA GT. Sequencing was carried using the PSQ™ 96MA pyrosequencing apparatus (Pyrosequencing, Biotage AB, Uppsala, Sweden) according to the instructions from the manufacturer.
The -717 T>C (rs2794521) and +1444 C>T (rs1130864) CRP polymorphisms were analysed with Taqmanr MGB Probes from Applied Biosystems according to the manufacturers protocol.
Statistical analysis
All statistical analyses were performed in StatView version 5.0.1 unless stated otherwise. Differences in genotype frequencies and allele frequencies between the ethnic groups were tested for statistical significances using χ2 tests. Analyses of the estimated haplotype frequencies and associations of alleles were performed in Unphased (version 2.403) [29]. Linkage analysis was performed in FStat version 2.9.3.2. A P-value of 0.05 or less was considered as statistically significant.
Results
CRP genotype and allele frequencies
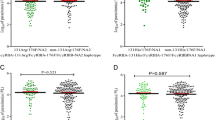
The genotype frequency of the -286 C>T>A polymorphism was similar in both countries (Table 1) and was found to be in Hardy-Weinberg equilibrium. The allele frequencies were significantly different between the two ethnic groups in Mali and Sudan (P < 0.0001) (Table 1), the A allele being at significantly higher frequency in the non-Fulani groups than in the Fulani groups (Mali: OR = 2.79, P = 1.4 × 10-8 and Sudan: OR = 2.51, P = 8.2 × 10-7, respectively). While the C allele was found to be at a lower frequency in the Malian non-Fulani as compared to the Fulani (OR = 0.36, P = 3 × 10-10), the T allele frequency was lower in the Sudanese non-Fulani group (OR = 0.28, P = 4 × 10-7).
Further, the possible influence of the A allele on some malariometric data collected for the Mali study group were analysed, and the results revealed that individuals being parasite positive were more likely to be A-allele carriers than parasite negative individuals (p = 0.04) (Table 2), but no differences were seen for genotypes or when separated based on ethnicity. No differences were seen in the frequency of splenomegaly (A-allele carriers with enlarged spleen: 25%, non A-allele carriers with enlarged spleen: 21%, p = 0.2) or haemoglobin levels (median (range): A-allele carriers: 11 (5.4–17.5), non A-allele carriers: 11 (3.3–17.7), p = 0.8) between A- and non-A allele carriers.
The genotype frequency of the -717 T>C polymorphism was similar in both countries (Table 3), and was found to be in Hardy-Weinberg equilibrium. Both the genotype and the allele frequencies of the -717 T>C polymorphism showed a significant difference in Mali, but not in Sudan. No differences in parasite positive individuals, splenomegaly or haemoglobin levels were found between the different genotypes (data not shown).
The genotype frequency of the +1444 C>T polymorphism was similar in both countries, and was found to be in Hardy-Weinberg equilibrium. No differences in either genotype, allele frequencies (Table 4) or malariometric indexes (data not shown) in neither Mali nor Sudan were detected for the +1444 C>T CRP polymorphism
Haplotype associations between CRP -717 T/C, -286 C/T/A, +1444C/T and FcγRIIa 131 R/H
Both the CRP and FcγRIIa genes are located on chromosome 1q21–1q23. Previous investigations of the two allotypes of the FcγR131H (rs1801274) polymorphism showed a difference in allele frequency between the Fulani and non-Fulani groups in both Mali [7] and Sudan [30]. These findings, together with the results in this study regarding the -286 CRP polymorphism, led us to investigate if the haplotype patterns in the FcγRIIa R131H and CRP -286, -717 and +1444 polymorphisms differ between the different ethnic groups. The polymorphisms were not in linkage disequilibrium. A clear pattern was seen with regards to haplotype patterns, haplotypes containing the A allele of the -286 CRP polymorphism being exclusively associated with the non-Fulani group in both Mali and Sudan (Table 5). This association was regardless of the FcγRIIa allotypes or the -717 and +1444 CRP polymorphisms.
Discussion
The present study demonstrates striking differences regarding CRP single nucleotide polymorphisms between sympatric ethnic groups, which may contribute to the differences in their susceptibility to malaria. In particular, the CRP SNP -286 C/T/A showed a marked difference in genotype frequencies between Fulani and non-Fulani individuals in two independent cohorts from Mali and Sudan. Lower frequencies of the A-allele of this SNP, previously associated with higher circulating CRP concentrations, were demonstrated in the two independent Fulani groups as compared to other sympatric ethnic groups. Moreover, a higher parasite prevalence was detected in individuals with the A allele, supporting the cause-relationship. The other two analysed polymorphisms did not show any consistent differences between Mali and Sudan, and in the haplotype analysis it was clearly the -286 A-allele that showed a consistent association with one ethnic group in both countries. The results suggest that the -286 CRP polymorphism may be a contributing factor in the lower susceptibility to malaria seen in the Fulani group.
However, the hypothesis that the -286 A-allele is associated with protection against infectious diseases due to a selection in African populations was refuted. The reason for this is not clear, but the A-allele might be associated with a lower risk of other diseases, as discussed below.
So far, inconclusive results, suggesting both a beneficial and harmful role of CRP in malaria [18, 19], have made it difficult to conclude about its role in the disease. However, a recent study showed that binding of CRP to infected red blood cells (RBCs) increased the removal of damaged RBCs from the circulation [31], which could lead to a more pronounced anaemia. Further, the control of P. falciparum parasitaemia is dependent on a pro-inflammatory response, and an uncontrolled inflammation is suggested to cause severe symptoms [32]. Higher levels of circulating CRP could influence the IL-10 levels and thereby affect the delicate balance between pro- and anti-inflammatory responses, leading to a reduced control of parasitaemia. Support for this notion comes from previous studies showing high circulating CRP levels in individuals with high parasitaemia [13, 14], indicating an effect of CRP on parasite clearance. IL-10 is also involved in the generation of peripheral regulatory T cells (Treg) [33]. The up-regulation of Tregs in a malaria infection has been shown to increase the parasite growth [34], and a recent study demonstrated that the Fulani present a functionally impaired Treg repertoire as compared to their sympatric neighbours [35]. Moreover, CRP may show direct effects on dendritic cell differentiation, maturation and function [36], and neutrophil chemotaxis and signalling [37]. Since dendritic cells are important in initiating and regulating immune responses, and neutrophils have shown a protective effect against malaria in vitro [38, 39], such influences may seriously hamper an effective immune response.
Low levels of CRP have been shown for a number of autoimmune diseases [40], and with the predicted low levels of circulating CRP in the Fulani groups, it could be assumed that the Fulani may be more susceptible to autoimmune diseases, and this is true for some disorders, e.g. diabetes [41]. The results of the present study may suggest, that a high level of circulating CRP can be detrimental for malaria protection, although by a yet unknown mechanism. The supporting promising finding of a higher frequency of parasite positive individuals among -286 A-allele carriers than among the non-A-allele carriers further strengthens this hypothesis. One limitation of the present study is that no reliable data on plasma levels of CRP could be presented due to lack of plasma in Mali and to lack of standardisation of blood sampling in Sudan. In future studies there is a need to study both unstimulated and plasma levels stimulated by malaria in relation to CRP genotypes in Africa.
The FcγRIIa 131 H/R polymorphism has been associated with malaria susceptibility in several studies [12, 42, 43]. CRP binds with a higher affinity to the receptor expressed by the R allele [11], which might have a competitive effect on the binding of the previously shown malaria protective immunoglobulins (Ig) G1 and IgG3 [44], thereby interfering with the protection against malaria. Since the CRP gene and the FcγRIIa gene are located in the same region on chromosome 1, the possibility of a susceptibility locus on this chromosome was investigated. Although the linkage analysis did not show any linkage between these two polymorphisms, associations were found between haplotypes containing the -286 A-allele, regardless of the FcγRIIa allotype or the -717 or +1444 CRP polymorphisms, and the non-Fulani groups. This finding suggests that the -286 CRP polymorphism could have a stronger impact on malaria susceptibility than the other investigated variants. The finding that there are more parasite positive individuals among the A-allele carriers than among the non-A allele carriers, further strengthens this suggestion. The sample size in this study was probably too small for making any firm conclusions, but the results strongly indicate a need for further studies on the impact of CRP in relation to susceptibility to malaria.
Conclusion
In conclusion, this study has demonstrated a marked difference in CRP genotype frequencies in two independent samples of Africans with low susceptibility to malaria as compared to sympatric ethnic groups. The -286 CRP polymorphism that was analysed has been shown to be functional in vitro, and associated with in vivo CRP levels in the circulation. This study further shows that the high producing -286 A-allele may be associated with parasitaemia, and it can, therefore, be speculated that CRP play a role in the early immune response to malaria infection. This may possibly occur by inhibiting dendritic cells and/or neutrophils, by competition with the binding of malaria specific IgG antibodies to Fcγ receptors, by enhance the clearance of infected RBCs, leading to more severe anaemia, or a deviation towards an anti-inflammatory cytokine response induced by CRP. Further research on the effect of CRP on malaria susceptibility is warranted, since recent studies on new therapeutic agents, that lower the circulating CRP levels [45], might open up new adjunct treatment options against malaria.
References
Greenwood BM, Groenendaal F, Bradley AK, Greenwood AM, Shenton F, Tulloch S, Hayes R: Ethnic differences in the prevalence of splenomegaly and malaria in The Gambia. Ann Trop Med Parasitol. 1987, 81: 345-354.
Modiano D, V P, Sirima BS, Nebie I, Diallo D, Esposito F, Coluzzi M: Different response to Plasmodium falciparum malaria in west African sympatric ethnic groups. Proc Natl Acad Sci USA. 1996, 93: 13206-13211. 10.1073/pnas.93.23.13206.
Dolo A, Modiano D, Maiga B, Daou M, Dolo G, Guindo H, Ba M, Maiga H, Coulibaly D, Perlman H, Blomberg MT, Toure YT, Coluzzi M, Doumbo O: Difference in susceptibility to malaria between two sympatric ethnic groups in Mali. Am J Trop Med Hyg. 2005, 72: 243-248.
Bolad A, Farouk SE, Israelsson E, Dolo A, Doumbo OK, Nebie I, Maiga B, Kouriba B, Luoni G, Sirima BS, Modiano D, Berzins K, Troye-Blomberg M: Distinct interethnic differences in immunoglobulin G class/subclass and immunoglobulin M antibody responses to malaria antigens but not in immunoglobulin G responses to nonmalarial antigens in sympatric tribes living in West Africa. Scand J Immunol. 2005, 61: 380-386. 10.1111/j.1365-3083.2005.01587.x.
Vafa M, Maiga B, Berzins K, Hayano M, Bereczky S, Dolo A, Daou M, Arama C, Kouriba B, F„rnert A, Doumbo OK, Troye-Blomberg M: Associations between the IL-4 -590 T allele and Plasmodium falciparum infection prevalence in asymptomatic Fulani of Mali. Microbes Infect. 2007, 9: 1043-1048. 10.1016/j.micinf.2007.04.011.
Luoni G, Verra F, Arca B, Sirima BS, Troye-Blomberg M, Coluzzi M, Kwiatkowski D, Modiano D: Antimalarial antibody levels and IL4 polymorphism in the Fulani of West Africa. Genes Immun. 2001, 2: 411-414. 10.1038/sj.gene.6363797.
Israelsson E, Vafa M, Maiga B, Lysen A, Iriemenam NC, Dolo A, Doumbo OK, Troye-Blomberg M, Berzins K: Differences in Fcγ receptor IIa genotypes and IgG subclass pattern of anti-malarial antibodies between sympatric ethnic groups in Mali. Malar J. 2008, 7: 175-10.1186/1475-2875-7-175.
Nasr A, Elghazali G, Giha H, Troye-Blomberg M, Berzins K: Interethnic differences in carriage of haemoglobin AS and Fcγ receptor IIa (CD32) genotypes in children living in eastern Sudan. Acta Trop. 2008, 105: 191-195. 10.1016/j.actatropica.2007.10.003.
Bang R, Marnell L, Mold C, Stein MP, Clos KT, Chivington-Buck C, Clos TW: Analysis of binding sites in human C-reactive protein for FcγRI, FcγRIIA, and C1q by site-directed mutagenesis. J Biol Chem. 2005, 280: 25095-25102. 10.1074/jbc.M504782200.
Daeron M: Fc receptor biology. Annu Rev Immunol. 1997, 15: 203-234. 10.1146/annurev.immunol.15.1.203.
Stein MP, Edberg JC, Kimberly RP, Mangan EK, Bharadwaj D, Mold C, Du Clos TW: C-reactive protein binding to FcγRIIa on human monocytes and neutrophils is allele-specific. J Clin Invest. 2000, 105: 369-376. 10.1172/JCI7817.
Braga ÉM, Scopel KKG, Komatsu NT, da Silva-Nunes M, Ferreira MU: Polymorphism in the Fcγ receptor IIA and malaria morbidity. J Mol Gen Med. 2005, 1: 5-10.
Abrams ET, Kwiek JJ, V M, Kamwendo DD, Tadesse E, V L, Molyneux ME, Rogerson SJ, Meshnick SR: Malaria during pregnancy and foetal haematological status in Blantyre, Malawi. Malar J. 2005, 4: 39-10.1186/1475-2875-4-39.
Hurt N, Smith T, Tanner M, Mwankusye S, Bordmann G, Weiss NA, Teuscher T: Evaluation of C-reactive protein and haptoglobin as malaria episode markers in an area of high transmission in Africa. Trans R Soc Trop Med Hyg. 1994, 88: 182-6. 10.1016/0035-9203(94)90287-9.
Nahrevanian H, Gholizadeh J, Farahmand M, Assmar M: Patterns of co-association of C-reactive protein and nitric oxide in malaria in endemic areas of Iran. Mem Inst Oswaldo Cruz. 2008, 103: 39-44. 10.1590/S0074-02762008000100006.
Imrie H, Fowkes FJ, Michon P, Tavul L, Reeder JC, Day KP: Low prevalence of an acute phase response in asymptomatic children from a malaria-endemic area of Papua New Guinea. Am J Trop Med Hyg. 2007, 76: 280-284.
Rohner F, Zimmermann MB, Wegmueller R, Tschannen AB, Hurrell RF: Mild riboflavin deficiency is highly prevalent in school-age children but does not increase risk for anaemia in Cote d'Ivoire. Br J Nutr. 2007, 97: 970-976. 10.1017/S0007114507665180.
Ansar W, Bandyopadhyay SM, Chowdhury S, Habib SH, Mandal C: Role of C-reactive protein in complement-mediated hemolysis in malaria. Glycoconj J. 2006, 23: 233-240. 10.1007/s10719-006-7928-0.
Pied S, Nussler A, Pontent M, Miltgen F, Matile H, Lambert PH, Mazier D: C-reactive protein protects against preerythrocytic stages of malaria. Infect Immun. 1989, 57: 278-82.
Marnell L, Mold C, Du Clos TW: C-reactive protein: ligands, receptors and role in inflammation. Clin Immunol. 2005, 117: 104-11. 10.1016/j.clim.2005.08.004.
MacGregor AJ, Gallimore JR, Spector TD, Pepys MB: Genetic effects on baseline values of C-reactive protein and serum amyloid a protein: a comparison of monozygotic and dizygotic twins. Clin Chem. 2004, 50: 130-134. 10.1373/clinchem.2003.028258.
Sainz J, Perez E, Gomez-Lopera S, Lopez-Fernandez E, Moratalla L, Oyonarte S, Jurado M: Genetic variants of IL6 gene promoter influence on C-reactive protein levels but are not associated with susceptibility to invasive pulmonary aspergillosis in haematological patients. Cytokine. 2008, 41: 268-278. 10.1016/j.cyto.2007.11.018.
Rogus J, Beck JD, Offenbacher S, Huttner K, Iacoviello L, Latella MC, de Gaetano M, Wang HY, Kornman KS, Duff GW: IL1B gene promoter haplotype pairs predict clinical levels of interleukin-1beta and C-reactive protein. Hum Genet. 2008, 123: 387-398. 10.1007/s00439-008-0488-6.
Hage FG, Szalai AJ: C-reactive protein gene polymorphisms, C-reactive protein blood levels, and cardiovascular disease risk. J Am Coll Cardiol. 2007, 50: 1115-1122. 10.1016/j.jacc.2007.06.012.
Kovacs A, Green F, Hansson LO, Lundman P, Samnegard A, Boquist S, Ericsson CG, Watkins H, Hamsten A, Tornvall P: A novel common single nucleotide polymorphism in the promoter region of the C-reactive protein gene associated with the plasma concentration of C-reactive protein. Atherosclerosis. 2005, 178: 193-198. 10.1016/j.atherosclerosis.2004.08.018.
Yanbaeva DG, Dentener MA, Spruit MA, Houwing-Duistermaat JJ, Kotz D, Passos VL, Wouters EF: IL6 and CRP haplotypes are associated with COPD risk and systemic inflammation: a case-control study. BMC Med Genet. 2009, 10: 23-10.1186/1471-2350-10-23.
Carlson CS, Aldred SF, Lee PK, Tracy RP, Schwartz SM, Rieder M, Liu K, Williams OD, Iribarren C, Lewis EC, Fornage M, Boerwinkle E, Gross M, Jaquish C, Nickerson DA, Myers RM, Siscovick DS, Reiner AP: Polymorphisms within the C-reactive protein (CRP) promoter region are associated with plasma CRP levels. Am J Hum Genet. 2005, 77: 64-77. 10.1086/431366.
Giha HA, Rosthoj S, Dodoo D, Hviid L, Satti GM, Scheike T, Arnot DE, Theander TG: The epidemiology of febrile malaria episodes in an area of unstable and seasonal transmission. Trans R Soc Trop Med Hyg. 2000, 94: 645-651. 10.1016/S0035-9203(00)90218-9.
Dudbridge F: Pedigree disequilibrium tests for multilocus haplotypes. Genet Epidemiol. 2003, 25: 115-121. 10.1002/gepi.10252.
Nasr A, Iriemenam NC, Giha HA, Balogun HA, Anders RF, Troye-Blomberg M, ElGhazali G, Berzins K: FcγRIIa (CD32) polymorphism and antimalarial IgG subclass pattern among Fulani and sympatric etnic groups living in eastern Sudan. Malar J. 2009, 8: 43-10.1186/1475-2875-8-43.
Ansar W, Habib SK, Roy S, Mandal C, Mandal C: Unraveling the C-reactive protein complement-cascade in destruction of red blood cells: potential pathological implications in Plasmodium falciparum malaria. Cell Physiol Biochem. 2009, 23: 175-190. 10.1159/000204106.
Malaguarnera L, Musumeci S: The immune response to Plasmodium falciparum malaria. Lancet Infect Dis. 2002, 2: 472-478. 10.1016/S1473-3099(02)00344-4.
Wilczynski JR, Radwan M, Kalinka J: The characterization and role of regulatory T cells in immune reactions. Front Biosci. 2008, 13: 2266-2274. 10.2741/2840.
Walther M, Tongren JE, Andrews L, Korbel D, King E, Fletcher H, Andersen RF, Bejon P, Thompson F, Dunachie SJ, Edele F, de Souza JB, Sinden RE, Gilbert SC, Riley EM, Hill AV: Upregulation of TGF-beta, FOXP3, and CD4+CD25+ regulatory T cells correlates with more rapid parasite growth in human malaria infection. Immunity. 2005, 23: 287-296. 10.1016/j.immuni.2005.08.006.
Torcia MG, Santarlasci V, Cosmi L, Clemente A, Maggi L, Mangano VD, Verra F, Bancone G, Nebie I, Sirima BS, Liotta F, Frosali F, Angeli R, Severini C, Sannella AR, Bonini P, Lucibello M, Maggi E, Garaci E, Coluzzi M, Cozzolino F, Annunziato F, Romagnani S, Modiano D: Functional deficit of T regulatory cells in Fulani, an ethnic group with low susceptibility to Plasmodium falciparum malaria. Proc Natl Acad Sci USA. 2008, 105: 646-651. 10.1073/pnas.0709969105.
Zhang R, Becnel L, Li M, Chen C, Yao Q: C-reactive protein impairs human CD14+ monocyte-derived dendritic cell differentiation, maturation and function. Eur J Immunol. 2006, 36: 2993-3006. 10.1002/eji.200635207.
Zhong W, Zen Q, Tebo J, Schlottmann K, Coggeshall M, Mortensen RF: Effect of human C-reactive protein on chemokine and chemotactic factor-induced neutrophil chemotaxis and signaling. J Immunol. 1998, 161: 2533-2540.
Brown J, Smalley ME: Inhibition of the in vitro growth of Plasmodium falciparum by human polymorphonuclear neutrophil leucocytes. Clin Exp Immunol. 1981, 46: 106-109.
Pierrot C, Adam E, Hot D, Lafitte S, Capron M, George JD, Khalife J: Contribution of T cells and neutrophils in protection of young susceptible rats from fatal experimental malaria. J Immunol. 2007, 178: 1713-1722.
Kravitz MS, Pitashny M, Shoenfeld Y: Protective molecules – C-reactive protein (CRP), serum amyloid P (SAP), pentraxin3 (PTX3), mannose-binding lectin (MBL), and apolipoprotein A1 (Apo A1), and their autoantibodies: prevalence and clinical significance in autoimmunity. J Clin Immunol. 2005, 25: 582-591. 10.1007/s10875-005-7828-2.
Fisch A, Pichard E, Prazuck T, Leblanc H, Sidibe Y, Brucker G: Prevalence and risk factors of diabetes mellitus in the rural region of Mali (West Africa): a practical approach. Diabetologia. 1987, 30: 859-862.
Nasr A, Iriemenam NC, Troye-Blomberg M, Giha HA, Balogun HA, Osman OF, Montgomery SM, ElGhazali G, Berzins K: Fcγ receptor IIa (CD32) polymorphism and antibody responses to asexual blood-stage antigens of Plasmodium falciparum malaria in Sudanese patients. Scand J Immunol. 2007, 66: 87-96. 10.1111/j.1365-3083.2007.01947.x.
Sinha S, Mishra SK, Sharma S, Patibandla PK, Mallick PK, Sharma SK, Mohanty S, Pati SS, Mishra SK, Ramteke BK, Bhatt RM, Joshi H, Dash AP, Ahuja RC, Awasthi S, Consortium IG, Venkatesh V, Habib S: Polymorphisms of TNF-enhancer and gene for FcγRIIa correlate with the severity of falciparum malaria in the ethnically diverse Indian population. Malar J. 2008, 7: 13-10.1186/1475-2875-7-13.
Aribot G, Rogier C, Sarthou JL, Trape JF, Balde AT, Druilhe P, Roussilhon C: Pattern of immunoglobulin isotype response to Plasmodium falciparum blood-stage antigens in individuals living in a holoendemic area of Senegal (Dielmo, west Africa). Am J Trop Med Hyg. 1996, 54: 449-57.
Pepys MB, Hirschfield GM, Tennent GA, Gallimore JR, Kahan MC, Bellotti V, Hawkins PN, Myers RM, Smith MD, Polara A, Cobb AJ, Ley SV, Aquilina JA, Robinson CV, Sharif I, Gray GA, Sabin CA, Jenvey MC, Kolstoe SE, Thompson D, Wood SP: Targeting C-reactive protein for the treatment of cardiovascular disease. Nature. 2006, 440: 1217-1221. 10.1038/nature04672.
Acknowledgements
We are grateful to the blood donors that participated in this study, and therefore made it possible. We thank Jenny Link for assistance in the statistical analysis and Selim Sengül for assistance in the pyrosequencing analysis.
This work was supported by grants to MTB and KB from the Swedish Agency for Research Development with Develo** Countries (SIDA, SAREC), the Swedish Medical Research Council (VR) as well as grants within the BioMalPar European Network of Excellence (LSMP-CT-2004-503578), from the Priority 1 "Life Sciences, Genomics and Biotechnology for Health" in the 6th Framework Programme. PT received grants from Hj„rt-Lungfonden.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
EI participated in the design and conception of the study, collection of samples, performed the CRP genoty**s and the CRP measurements, carried out the statistical analyses and drafted the manuscript. ME participated in the design and conception of the study and helped to draft the manuscript. AN was responsible for design of sample collection in Sudan and helped to draft the manuscript. AD was responsible for design of sample collection in Mali and helped to draft the manuscript. SK helped in the design of TaqMan analysies and performed the CRP genoty** and helped to draft the manuscript. GA participated in the Pyrosequencing analysis and the CRP measurements and helped to draft the manuscript. MV helped in the collection of samples and helped to draft the manuscript. BM examined all participants from Mali and participated in design and collection of samples and helped to draft the manuscript. OKD was responsible for design of sample collection in Mali and helped to draft the manuscript. GE was responsible for design of sample collection in Sudan and helped to draft the manuscript. HAG was responsible for design of sample collection in Sudan and helped to draft the manuscript. MTB participated in the design and the conception of the study and helped to draft the manuscript. KB participated in the design and the conception of the study and drafted the manuscript. PT participated in the design and the conception of the study, performed the CRP genoty** and helped to draft the manuscript. All authors read and approved the final manuscript
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Israelsson, E., Ekström, M., Nasr, A. et al. Marked differences in CRP genotype frequencies between the Fulani and sympatric ethnic groups in Africa. Malar J 8, 136 (2009). https://doi.org/10.1186/1475-2875-8-136
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2875-8-136