
Abstract
Hydrocarbon-degrading bacteria, which can be found living with eukaryotic phytoplankton, play a pivotal role in the fate of oil spillage to the marine environment. Considering the susceptibility of calcium carbonate-bearing phytoplankton under future ocean acidification conditions and their oil-degrading communities to oil exposure under such conditions, we investigated the response of non-axenic E. huxleyi to crude oil under ambient versus elevated CO2 concentrations. Under elevated CO2 conditions, exposure to crude oil resulted in the immediate decline of E. huxleyi, with concomitant shifts in the relative abundance of Alphaproteobacteria and Gammaproteobacteria. Survival of E. huxleyi under ambient conditions following oil enrichment was likely facilitated by enrichment of oil-degraders Methylobacterium and Sphingomonas, while the increase in relative abundance of Marinobacter and unclassified Gammaproteobacteria may have increased competitive pressure with E. huxleyi for micronutrient acquisition. Biodegradation of the oil was not affected by elevated CO2 despite a shift in relative abundance of known and putative hydrocarbon degraders. While ocean acidification does not appear to affect microbial degradation of crude oil, elevated mortality responses of E. huxleyi and shifts in the bacterial community illustrates the complexity of microalgal-bacterial interactions and highlights the need to factor these into future ecosystem recovery projections.
Similar content being viewed by others
Introduction
Marine eukaryotic phytoplankton (microalgae) contribute significantly to some key global processes, including approximately half of global carbon fixation1 and approximately half of the oxygen in the atmosphere2. As key members at the base of the food chain, they play a fundamental role in the ecology of marine ecosystems. Together with their bacterial symbionts, which are found associated with their microalgal hosts at the cell surface (i.e. the phycosphere), these algal–bacterial communities are a major source and recycler of organic and inorganic carbon and nutrients3,4,5. Bacteria associated with microalgae have been posited to utilise algal exudates as carbon and energy sources6,7, whereas the algal hosts can benefit from bacterial-mediated trace metal/nutrient bioavailability8. A mutual sharing of iron and fixed carbon was shown to occur between several species of microalgae and bacteria9. Another study demonstrated that the supply of bacterial-produced vitamin B12 to the microalgal host occurred in exchange for fixed carbon10. It is also noteworthy that very few microalgal species can be maintained or sub-cultured in the laboratory for long periods in the absence of their bacterial symbionts (i.e. in an axenic state), testament to the pivotal role that the associated bacterial community plays in their overall success. However, studies aimed at assessing the impact of future ocean conditions on microalgae have largely employed the use of axenic species, or non-axenic individual species or consortia where the focus was on either the microalgae or the bacteria, rather than both as a collective holobiont (e.g. Joint et al.11).
The microbiology of the ocean, likely at all levels of habitation, are and will continue to be subject to anthropogenic-induced global change, with temperature and CO2 increases12,13. Of particular interest, the impact of future ocean conditions on coccolithophores has been extensively studied in recent years due to the potential sensitivity of these organisms to ocean acidification (OA)14,15. Despite OA affecting calcification of coccoliths, field studies have demonstrated an increase in abundance of coccolithophores in the subtropical region in recent years due to CO2 enrichment16,17. Laboratory studies of the coccolithophore E. huxleyi reported inconsistencies in response to OA due to strain specific responses as a result of pan-genomic variability16,18,19, which likely accounts for their global distribution and abundance during phytoplankton blooms.
E. huxleyi produces organic carbon compounds such as alkenone lipids, long-chained alkenes, long-chained alkenones, phospholipids, and glycolipids that all bare similarities to hydrocarbons that are constituents of crude oil20,21. As such, hydrocarbon-degrading bacteria have commonly been found living associated with E. huxleyi9,22, as well as with other species of eukaryotic phytoplankton23,24,25,26,27. Coccolithophores such as E. huxleyi, have also been found to harbour a bacterial community that is more diverse than that of dinoflagellates and diatoms, possibly due to the biochemically and biophysically more complex environment associated with the cell surface of actively growing and calcifying coccolithophores22. As E. huxleyi thrives in oligotrophic expanses of the open ocean, they could be a significant seed source of hydrocarbon-degrading bacteria in low-nutrient surface waters, as well as from the subsurface to the seafloor if these bacteria remain attached to dead coccolith cells and survive vertical sedimentation to the seafloor as marine snow.
The total quantity of underground oil reserves around the world is finite, so the rate of its production, which also accounts for the rate of discovering new underground oil reserves, has been in a steady state of decline for decades now. Consequently, the oil and gas industry has intensified its interests to exploring for oil in more challenging environments that include the Arctic, the subarctic, and in ever deeper waters. All these environments carry risk of disaster and the Deepwater Horizon oil spill is testament to this, which resulted in approximately 700,000 tonnes (4.9 million barrels) of Louisiana light sweet crude oil discharged into the Gulf of Mexico from a blown-out wellhead at a depth of ~ 1500 m28,29. Crude oil is a heterogenous mixture of thousands of chemical compounds grouped as saturates, aromatics, resins, and asphaltenes that can cause acute or chronic toxicity to a wide variety of sea life. Crude oil toxicity to microalgae can include impairment of cell permeability, loss of cell nuclei and cell mobility, CO2 absorption capacity, alterations to protein content, shrinkage of chloroplasts and pyrenoids, disruption of nuclei acid synthesis, and damage to DNA and RNA from oxidative stress30,31,32,33,34,35.
In the absence of national and international government intervention to prevent the oil and gas industry from exploring for oil in deeper waters and polar/subpolar environments, the likelihood of oil spills occurring in oligotrophic environments will inevitably be much higher than it has ever been. It is therefore imperative to gain foresight on how microbial communities might respond to crude oil spillage under future OA conditions. In this study, we investigated the growth and physicochemical changes in E. huxleyi, as well as shifts to its associated bacterial community to crude oil under future OA conditions. Our results provide new understanding on the future risk of crude oil spillage in the open ocean, where the synergistic or additive responses of these communities will be an underpinning factor in the resilience of the ocean to recovering from oil pollution in a future climate.
Results
E. huxleyi response to oil enrichment in elevated CO2 conditions
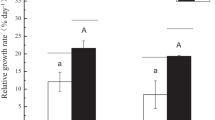
Changes in Chl a were used to determine the dynamics of E. huxleyi in response to crude oil under ambient and elevated CO2 conditions (Fig. 1). Distinct differences in the growth dynamic of the alga occurred between these conditions. Initially, Chl a concentrations (proxy for E. huxleyi growth) in elevated CO2 (750 ppm) microcosms were not significantly different to those in ambient CO2-treated microcosms on day 2 or 3 (p = 0.994, and p = 0.9644). On days 4 and 5, Chl a concentrations were significantly higher in elevated CO2-treated microcosms (p = 0.0002, p < 0.0001). In oil-amended microcosms, Chl a concentrations under ambient CO2 conditions continued to increase exponentially for an additional 24 h after oil enrichment at day 5, whereas in the elevated CO2 microcosms Chl a concentrations ceased increasing following oil enrichment. Chl a concentrations in the ambient microcosms reached significantly higher levels at days 12 and 15 (p < 0.0001 and p < 0.0001), and remained as such until the termination of these experiments. In contrast, Chl a concentrations after oil enrichment in the elevated CO2 microcosms decreased to below detection levels after day 12.

Growth response of E. huxleyi, measured by monitoring Chl a concentrations, in microcosms treated with ambient (400 ppm) and elevated (750 ppm) CO2. Red line indicates the timepoint (day 5) at which crude oil was added to the oil-amended microcosms. Values are averages of triplicate incubations ± standard error.
Bacterial community dynamics in response to crude oil under ambient and elevated CO2 conditions
MiSeq sequencing of all samples (including triplicates) returned a total of 4,463,167 individual sequence reads. After pre-processing (merging, trimming, short-read and quality filtering), demultiplexing, denoising/clustering, chimera filtration and applying an OTU clustering cut-off of 97% for sequence similarity, 1,138 good-quality OTUs were obtained. This included 7 putatively novel taxa belonging to the classes Gammaproteobacteria and Alphaproteobacteria, and the families Rhodobacteraceae, Halomonadaceae, Alteromonadaceae and Veillonellaceae, and to an unclassified bacterial taxon. Species richness of the bacterial communities across treatments showed similar evenness in Shannon diversity index (p > 0.05; Supplementary Fig. 1). A non-metric multidimensional scaling (nMDS) ordination plot showed at least 60% similarity of the communities across all treatments (Supplementary Fig. 2). A heatmap showing the bacterial community at family level (Fig. 2) shows similarity of taxa distribution were not clustered depending on CO2 levels or oil exposure. ANOSIM pairwise test identified no statistical differences (p > 0.05) in the beta diversity of the bacterial communities, based on the Bray–Curtis dissimilarity matrix, between all treatments, thus supporting the results in Supplementary Fig. 2 by nMDS indicating that OA conditions (with reference to the elevated CO2 treatments) and exposure to crude oil does not significantly affect the diversity and composition of the bacterial community associated with E. huxleyi.

Heatmap of the bacterial OTUs that became enriched in incubations of E. huxleyi treated to ambient or elevated CO2 concentrations with or without exposure to crude oil at timepoints t0, t1 and t3 (described in main text). Uninoculated treatments represent those that were not inoculated with E. huxleyi (uninoculated controls). OTUs were considered enriched if there was a mean increase of at least 1% relative abundance (e.g., shift from 1 to 2%) in at least one time point for replicate incubations (i, ii, iii), and if the difference was statistically significant (p < 0.05). Colour key indicates square-root normalized relative abundance (%).
Further analysis by pairwise comparison (log2 median ratio) of bacterial taxa between treatments displayed as heat trees (Fig. 3) showed significant shifts in relative abundance of taxa within the community between treatments. Under ambient CO2 conditions, the bacterial community associated with E. huxleyi were dominated by members belonging to the genus Marinobacter (29% relative abundance), followed by unclassified members of the class Gammaproteobacteria (11%), the genus Methylobacterium (8%), the class Alphaproteobacteria (6%) and to unclassified bacteria (6%) (Supplementary Table 1). Prior to oil enrichment, Marinobacter and Methylobacterium were significantly higher in relative abundance in ambient CO2 microcosms compared to in microcosms with elevated CO2, while no taxa were found enriched under elevated CO2 conditions compared to ambient treated microcosms prior to oil enrichment (Fig. 3). In comparison, the bacterial community in the elevated CO2 treatments was dominated by Marinobacter (15%), Methylobacterium (11%), Sphingomonas (8%) and unclassified bacteria (13%) (Supplementary Table 1). A decrease of 6% relative abundance of Marinobacter was observed to have occurred in the ambient CO2 microcosms (reaching 15% abundance) after 7 days of oil enrichment. However, an increase of 9% was observed in the elevated CO2 treated microcosms (reaching 24%), while Methylobacterium decreased by 1% to 3% in the elevated CO2 microcosms (Supplementary Table 1).

Heat-tree plot of bacterial community associated with E. huxleyi in different CO2 and oil enrichment treatments. Larger grey tree represents taxon labelled key of the smaller heat trees showing pairwise comparison between treatments. Diameter of nodes shows qualitative number of OTU of each taxon. Colour intensity of smaller trees (green or brown) corresponds to significantly abundant taxons between treatments determined with Wilcox rank-sum test followed by Benjamini Hochberg (FDR) correction for multiple comparisons. Treatments correspond to CO2 acclimation at 400 ppm (ambient) or 750 ppm (elevated) at different timepoints. Timepoints: t0 represents day 5 since the start of the incubations and just prior to oil enrichment, t1 represent day 12 since the start of the incubations or 7 days after oil enrichment, and t2 represents day 19 since the start of the incubations or 14 days after oil enrichment.
After just one day since the addition of crude oil to the ambient CO2 treatments, major increases in relative abundances were observed for members of unclassified Bacteria (by 12%) and unclassified Halomonadaceae (by 5% from undetectable levels), whereas minor increases occurred with Methylobacterium and unclassified Rhodobacteraceae (3% each), Methylophaga, Corynebacterium and unclassified Alphaproteobacteria (2% each), and Sphingomonas, Stappia (from undetectable levels) and Halomonas (1% each) (Supplementary Table 1). Within this same short time period since the addition of the oil to the elevated CO2 treatments, major increases in relative abundances were observed for Marinobacter (9%), Prevotella (7%), Streptococcus (6%) and Veillonella (5% from undetectable levels), with minor increases by Gracilimonas (4% from undetectable levels), Stappia (from undetectable levels), unclassified Veillonellaceae (from undetectable levels), Sulfitobacter and unclassified Gammaproteobacteria (3% each), and Campylobacter (from undetectable levels) and Fusobacterium (2% each) (Supplementary Table 1). At the end of these microcosm incubations that represented 14 days after the addition of the oil (t2), notable dynamics of specific taxa were observed for Methylobacterium which maintained a higher relative abundance in the ambient CO2 incubations without oil, whereas Marinobacter maintained a high relative abundance (15–29%) in both the ambient (without oil) and the elevated (with oil) CO2 treatments.
Degradation of crude oil under ambient versus elevated CO2 conditions
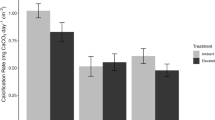
Of 35 n-alkanes analysed (nC8–nC43), thirteen (nC15–nC28) were biodegraded by the bacterial community associated with E. huxleyi, with significant differences detected for nC17/pristane and nC18/phytane between killed controls and inoculated samples (p < 0.0001) at the termination of the experiment (Fig. 4; Supplementary Fig. 3), thus indicating biodegradation of crude oil by the bacterial community associated with E. huxleyi. However, no significant differences (p < 0.05) of these aliphatic biomarkers was found in neither the ambient nor elevated CO2 treatments, suggesting that biodegradation of aliphatic hydrocarbons in crude oil is not affected by elevated CO2 conditions. Lower molecular weight n-alkanes, which includes nC8–nC10, were not detected in acid-killed controls and E. huxleyi cultures likely due to their evaporation over the course of the experiment.

Differences in hydrocarbon ratios comparing live treatments of E. huxleyi with its associated bacterial community (solid bars; Ehux microbiota) to their respective acidified controls (open bars; killed controls) for parameters indicative of biodegradation: nC17/pristine, nC18/phytane, naphthalene/2-methylnaphthalene (N:2-MN), 2-methylnaphthalene/1-methylnaphthalene (2-MN;1-MN) 2-ethylnaphthalene/2,6 + 2,7-dimethylnapthalene (2-ET:(2,6 + 2,7)-DMN), phenanthrene/9-methylphenanthrene (P:9-MP), 3-methylphenanthrene + 2methylphenanthrene/9-methylphenanthrene + 1-methylphenanthrene (3-MP + 2-MP:9-MP + 1-MP), 3-methylphenanthrene/9-methylphenanthrene (3-MP:9-MP). Values are averages of triplicate incubations with standard error bars.
Reductions of aromatic compounds, which include naphthalene/2-methylnaphthalene (N:2-MN), 2-methylnaphthalene/1-methylnaphthalene (2-MN;1-MN) 2-ethylnaphthalene/2,6 + 2,7-dimethylnapthalene (2-ET:(2,6 + 2,7)-DMN), phenanthrene/9-methylphenanthrene (P:9-MP), 3-methylphenanthrene + 2methylphenanthrene/9-methylphenanthrene + 1-methylphenanthrene (3-MP + 2-MP:9-MP + 1-MP), 3-methylphenanthrene/9-methylphenanthrene (3-MP:9-MP), were not detected after 15 days of exposure with E. huxleyi and its associated bacterial community (Fig. 4). Although the bacterial community associated with E. huxleyi had only reduced the low-molecular-weight fraction of n-alkanes such as pristane and phytane, but not aromatics within 15 days of crude oil enrichment, elevated CO2 conditions did not significantly affect the n-alkanes degradation of crude oil in inoculated samples.
Discussion
This study shows, for the first time, the effects of crude oil on E. huxleyi and its associated bacterial community under future OA (elevated CO2) conditions. Initially, we show that OA enhanced the growth of E. huxleyi, as elevated CO2 resulted in higher Chl a concentrations (a proxy for growth of microalgae) and this is consistent with some previous studies36,37. However, crude oil induced a negative response, and together with elevated CO2, both conditions appeared to have a synergistic affect upon the growth of E. huxleyi. CO2 is a limiting factor in photosynthesis, therefore higher concentrations can be utilised to compensate for the metabolic stress of maintaining intracellular pH and especially as cells divide and replicate. Whilst the cells may grow in more abundance under elevated CO2, we posit that their coccoliths might be less calcified due to the relatively lower pH. If this were the case, it could explain their higher susceptibility to crude oil toxicity. E. huxleyi was reported to adopt the ‘Cheshire Cat’ escape strategy where the cells exist predominantly in a haploid stage that lack in mineralised scales, allowing evasion of viral predation in addition to physical and chemical stress38,39,40. Further work would be needed to ascertain whether calcification provides a physical defence against crude oil toxicity, or if the converse (i.e. a lack of mineralised scales) puts the organism at risk of being "killed" when challenged with crude oil. Hydrocarbon adsorption on calcite surfaces has been reported41. As such, it may be conjectured that thicker, healthier calcite scales would prevent, or might at least reduce the absorption of toxic hydrocarbon chemicals through and into the cell membrane and intracellular region of the cells where it would inevitably cause impairment of cell functioning and ultimately death. As discussed below, the associated bacterial community, and more specifically the oil-degrading community will play a protective role from crude oil toxicity.
Community analysis of the various treatments showed that bacterial species diversity associated with E. huxleyi was not significantly different neither under elevated compared to ambient CO2 conditions, nor with/without exposure to crude oil. However, certain shifts in the communities, as effected by certain taxa, were observed that delineated these treatments, as observed with the transient enrichment of Marinobacter, Methylobacterium and unclassified Bacteria in the ambient CO2 treatments after exposure to crude oil. Marinobacter is a genus known to utilize aliphatic and polycyclic aromatic hydrocarbons as a sole source of carbon and energy42,43,44. Whilst Marinobacter are commonly associated with the degradation of aliphatic and aromatic hydrocarbons in the ocean, they are in fact also capable of utilizing various non-hydrocarbon substrates, such as organic nitrogen-containing compounds (e.g. amino acids, carboxylic acids etc.)45. A decrease in their relative abundance (0–19%) after exposure to the oil was likely due to increased competition for bioavailable macro- and/or micronutrients. Although primary production increases production of organic carbon, photosynthesis utilises micronutrients such as iron that are co-limiting growth factors for heterotrophic bacteria3,46,47. Not unexpectedly, Marinobacter increased in relative abundance after oil enrichment and primarily under elevated CO2 conditions which has not been previously reported. We posit that under future OA conditions, these ubiquitous oil-degrading organisms may not be expected to be affected in their natural response to oil spillage.
Unlike Marinobacter, under elevated CO2 conditions Methylophaga and Methylobacterium became negatively, albeit marginally, impacted following exposure to crude oil, whereas these organisms thrived under the ambient CO2 treatments after exposure to the oil. Whilst some methylotrophs have been shown to become enriched by crude oil or its petrochemical refined products [42 and references therein], hitherto this has not been reported for members of the genus Methylobacterium. This genus belongs to a group of organisms known for methylotrophy, in that they rely exclusively on single carbon compounds, such as methanol, methylamine and dimethylsulfide, as a sole source of carbon and energy48. However, genes have been identified within Methylobacterium genomes that are associated with the degradation of hydrocarbons49, and the genome annotation for Methylobacterium extorquens PA1 in the KEGG (http://www.genome.jp/kegg-bin/show_pathway?mex01220) has been shown to possess anaerobic benzene degradation genes. Some methylotrophs, particularly of the genus Methylophaga, are also capable of utilising fructose50 or even hydrocarbons51,52 as sole growth substrates. Furthermore, the presence of hydrocarbon-degrading methylotrophs has been shown amongst communities of bacteria associated with eukaryotic phytoplankton27,53. This might be because eukaryotic phytoplankton can adsorb hydrocarbons from the seawater environment or because they synthesise alkenones and other hydrocarbon-like compounds [54 and references therein] which hydrocarbon-degrading methylotrophs could potentially feed on, although this hydrocarbon-mediated algal–bacterial symbiosis remains unproven. Whilst it remains to be substantiated, we posit that the enrichment of Methylobacterium in the presence of crude oil under ambient CO2 levels may be due to their potential to utilise hydrocarbons as a source of carbon and energy. An alternative explanation for their enrichment may be because these organisms were able to acquire their carbon and energy requirements from extracellular particulate and/or dissolved organic carbon (DOC) released by E. huxleyi; such exudates can offer a rich source of methylated sugars that can be utilised by methylotrophs55,56. It is also noteworthy to mention the release of organic matter exudates by E. huxleyi has been reported to be affected by changes in dissolved CO2 levels57. Whilst we did not analyse for exudate production in our experiments, others showed that elevated CO2 conditions significantly increased the release of transparent exopolymer particles (TEP) and particulate combined carbohydrates (pCCHO) by E. huxleyi57. Although the lability of this carbon source to be utilised by methylotrophs, like Methylobacterium, is unknown. At the very least, Methylobacterium was able to survive when challenged with the crude oil, but only under ambient CO2 conditions. As explained earlier, the coccolith scales of E. huxleyi would be in a weakened state, or less robust, to deal with oil exposure under elevated CO2 conditions, due to the detrimental effects caused by a reduced pH to the structure of their calcium carbonate armour, potentially permitting the intracellular entry of toxic hydrocarbons.
Sphingomonas are metabolically diverse and capable of utilising hydrocarbons as growth substrates58, but these organisms were not enriched after oil exposure under ambient CO2 conditions, and their relative abundance had noticeably decreased after oil exposure in the elevated CO2 treatment. This genus belongs to the class Alphaproteobacteria, and their decrease in relative abundance under elevated CO2 supports studies that found that OA conditions (from elevated CO2 levels) can negatively impact members belonging to this class59,60. Other genera comprising members with reported hydrocarbon-degrading abilities, such as Halomonas and Alteromonas, and which have been reported associated with E. huxleyi, such as Marivita, Hoeflea, Balneola, Arenibacter, Marinoscillum and Thalassospira22, were also detected but not affected by either CO2 treatment and nor exposure to the oil. We suspect that these organisms found associated with E. huxleyi are incapable of utilising hydrocarbons, which might explain why they were not enriched by the oil in either of the CO2 treatments. On the other hand, members of the genera Prevotella, Streptococcus and Veillonella which have been reported with hydrocarbon-degrading ability61,62,63,64, had increased in relative abundance following oil enrichment, but only under the elevated CO2 treatment, but then declined to < 1% abundance by the termination of these experiments. It is unknown to us at present what may have caused this post-bloom decline of these taxa and why these changes only occurred in the elevated CO2 treatment. It is possible their decline unfolded as a consequence of an essential nutrient becoming limited, accumulation of a toxic byproduct(s) in these microcosms, or other factor(s) that could be explored in future studies.
With atmospheric CO2 concentrations continuing to unabatedly rise and drive OA, our results show that the structuring of bacterial communities living in association with coccolithophores will be quite different in a future ocean, including potential extinctions of some oil-degrading taxa from these communities. Consequently, it is expected that this could have quite profound effects on the fate of spilled crude oil in the ocean, whether from natural seepage or anthropogenic inputs, considering that oil-degrading bacteria are crucial in the biodegradation and ultimate purging of petrochemical pollutants in the ocean. Assessing this by comparing nC17/pristane and nC18/phytane ratios of acid-killed incubations with the same from live incubations, we found that the biodegradation of the aliphatic fraction occurred under both ambient and elevated CO2 conditions. Despite shifts in microbial population dynamics due to elevated CO2 conditions, aliphatic hydrocarbon degradation was unaffected in both ambient and elevated CO2 conditions tested. We note that nC8, nC9, and nC10 were not detected in all treatments, likely due to their loss by volatilisation as expected for these low-molecular-weight hydrocarbons. On the other hand, the aromatic fraction in the crude oil, as measured by our analysis of naphthalene, methylnaphthalene, dimethylnaphthalene, ethylnaphthalene, C3-alkylnaphthalene, phenanthrene, methylphenanthrene, dimethylphenanthrene, ethylphenanthrene and C3-alkylphenanthrene, was not significantly degraded in neither the ambient or elevated CO2 conditions. It is possible that the oil-degrading microbial community associated with E. huxleyi was, for some reason, incapable of inducing the degradation of aromatic hydrocarbons. The metabolic capability to degrade aromatic hydrocarbons, however, would likely have been served by Marinobacter65,66,67,68, as members of this genus have been reported to utilise polycyclic aromatic hydrocarbons as sole growth substrates. Our experiments were run for 22 days, which was likely too short to begin to detect the initial stages in the biodegradation of the aromatic fraction because, as has been reported in many marine oil spill studies, the biodegradation of the aromatic fraction in crude oil does not often commence until the more labile aliphatics have become almost depleted (e.g. Head et al.69). Whilst we cannot conclude on what effects OA conditions might have on the biodegradation of aromatic hydrocarbons, based on our analysis of the aliphatic fraction, our results suggest that the response and biodegradation activities of oil-degrading communities associated with coccoliphores, such as E. huxleyi, would be largely unaltered when challenged with crude oil in future OA conditions.
Conclusion
The study presents evidence that in the event of an oil spill under future OA conditions, E. huxleyi becomes highly vulnerable, whereas its associated bacterial community, with the exception of a few taxa, are resilient. Marinobacter was the dominant bacteria associated with E. huxleyi prior to oil-enrichment in both ambient and elevated CO2 conditions, and in elevated CO2/oil-enriched conditions up to 16 days after oil exposure. Only under ambient/oil-enriched conditions were Marinobacter overtaken by unclassified bacteria as the dominant taxa. Significant changes in population dynamics after oil enrichment between CO2 treatments where relative abundance of Sphingomonas and Methylobacterium from the class Alphaproteobacteria were reduced, did not significantly affect degradation of pristane and phytane from crude oil. Changes in bacterial population, though small, can be crucial to host survival. Recovery of an ecosystem from oil pollution under projected future ocean conditions depends not just on the potential for hydrocarbon degradation, but also on the microalgal host survival and primary production. Therefore, in a natural community of phytoplankton, where multiple species of microalgae and their microbiota co-exist and interact with each other, it is important to assess how primary producers and their bacterial community respond to crude oil enrichment under projected OA conditions since they are the foundation of an ecosystem.
Methods
Organism, maintenance and preparation of CO2-acclimatised inocula
A non-axenic Emiliania huxleyi, strain 920/8, was obtained from the Culture Collection of Algae and Protozoa (CCAP; Oban, Scotland). The strain was maintained in f/2 algal medium70 and in a temperature-controlled illuminated incubator as per the guidelines of the CCAP. The strain was originally isolated in 1992 from a Bergen mesocosm in Norway and in subsequent work it was found to harbour members of bacteria belonging to genera of obligate (e.g. Alcanivorax, Marinobacter) and generalist oil-degrading genera (e.g. Arenibacter, Sulfitobacter, Thalassospira)22.
E. huxleyi acclimatised to either ambient or elevated CO2 levels were prepared for use as inocula. Cultures were pre-conditioned at 14 °C with ambient light (50 μmol m−2 s−1) at a 16:8 h light:dark cycle. Bubbling of CO2 at ambient or elevated CO2 directly into the cultures was used to achieve soluble CO2 concentrations of 400 or 750 ppm, respectively. A 750 ppm concentration is the expected moderate projection for atmospheric CO2 by 2100, a value midway between SRES scenario A1B and A271, and has been used in numerous algal-based ocean acidification studies. Ambient air was supplied from an air pump connected to outside air, while elevated CO2 was mixed using mass flow controllers to control the flow rate of ambient air or of compressed CO2. CO2 levels determined for elevated conditions were measured and monitored constantly using a COZIR ambient sensor (Gas Sensing Solutions Ltd., Glasgow, UK) calibrated with premixed 750 ppm CO2 (CalGaz Ltd., Newcastle, UK). CO2 readings were recorded per second and daily averages were calculated using R software. E. huxleyi was acclimatised for 100 generations under the two CO2 conditions (400 and 750 ppm) prior to use in experiments.
Supplementary Figure 4 shows the daily average CO2 concentrations during 20-day incubations supplemented with elevated atmospheric CO2, producing a CO2 concentration linear regression (standard curve) of 740 ppm. Two-way ANOVA analysis of pH in microcosms inoculated with or without E. huxleyi (Supplementary Fig. 5) shows significant differences between the treatments (p < 0.05), and between time and treatment (p < 0.005). A Tukey’s multiple comparison test showed that just prior to oil addition on day 5, pH levels were significantly different in ambient and elevated CO2 treated microcosms inoculated with E. huxleyi (p = 0.0107) and on day 20 (p = 0.0174; Two-Way ANOVA).
Microcosm setup
Microcosms were set up using 500 ml Duran Youtility bottles with GL-45 4-inlet screw caps (Supplementary Fig. 6). Two of the 4-inlets were used for channelling air in and out of the bottles. Each microcosm was fitted with a 0.22 μm inline air filter attached to the inlet and outlet of the GL-45 bottle cap to prevent contamination. A glass Pasteur pipette connected to the inside of the GL-45 cap of each microcosm was used to aerate the system and also provide agitation by gentle bubbling. All materials that were in direct contact with the aqueous system inside the microcosms were made of glass in order to prevent the adsorption of hydrocarbons when crude oil was used.
Exponentially-growing cells of pre-acclimatised E. huxleyi were counted using a haemocytometer in order to standardise the inocula so that all microcosms received approximately the same cell concentration. A series of the sterilised Duran bottles, each containing 300 ml of sterile f/2 medium, were inoculated with standardised cell suspensions that were acclimatised to either ambient or elevated CO2 levels—final cell concentrations achieved across all microcosms were 8 × 103 cells/ml. Microcosm treatments (each in triplicate) included (i) an oil-amended treatment inoculated with E. huxleyi for biological sampling, (ii) an oil-amended treatment inoculated with E. huxleyi for hydrocarbon analysis, (iii) a non-oil-amended treatment inoculated with E. huxleyi for biological sampling, and (iv) an acid-killed oil-amended control inoculated with E. huxleyi to account for loss of hydrocarbons due to abiotic factors, for which phosphoric acid was added to a final pH of ~ 1. The treatments were conducted under ambient and elevated CO2 conditions, and the complete setup totalled 24 individual microcosms. For the microcosm treatments designated for amendment with oil, the oil was added at day 5 to 1% (v/v) and Schiehallion crude oil (API 25°; sourced from BP) was used. All microcosms were incubated in a temperature-controlled incubator maintained at 14 °C.
The bottles were arranged equidistant from a central, vertical light source to ensure each microcosm received equal light intensity at a 16:8 h light:dark cycle. This arrangement avoids self-shadowing of microalgal cells, especially during their exponential growth which can lead to reduction of light intensity with increasing path length, as per Beer’s law, and shadowing from bottle caps and airline connections. AquaWhite flexi-LED (Aqualease Ltd., Blackburn, UK) strips were used to construct the light cylinders by wrap** them around a plastic tube for provision of a centralized vertical light source for each microcosm setup. Neutral density filters were used to reduce the light intensity to 58 µmol m−2 s−1.
Samples for chlorophyl a analysis and DNA extraction for monitoring the dynamics of E. huxleyi and the bacterial community by amplicon sequencing analysis were taken periodically from all microcosms, with the exception of the oil-amended incubations which were maintained unsampled until the end of the experiment, when they were sacrificed for hydrocarbon analysis (described below).
Quantification of E. huxleyi dynamics by Chl a analysis
To assess the dynamics of E. huxleyi in response to crude oil exposure, changes in chlorophyll a (Chl a) were determined using a modified version of EPA 44572. For this, samples (2 ml) were centrifuged (13,000 xg; 10 min) and the cell pellets carefully transferred to a clean tube containing 1 ml of 90% acetone and stored in the dark at − 20 °C overnight. For samples derived from oil-amended microcosms, transferring the pellets to a clean tube helped eliminate the possibility of any carry-over from residual oil or solubilised hydrocarbons. After overnight storage, the samples were centrifuged and the supernatant fractions transferred into clean glass vials for fluorometric analysis at an excitation wavelength of 485 nm and emission of 685 nm using a Turner Trilogy Fluorometer (Turner Designs, CA, USA). Chl a concentrations were calculated from a standard curve constructed from serial dilutions of known Chl a concentrations (Turner Designs, CA, USA), as per the method of Welschmeyer73.
DNA extraction, amplification and Illumina MiSeq sequencing
The diversity and response of the algal (E. huxleyi)-associated bacterial community to crude oil was assessed for each of the ambient and elevated CO2 treatments using Illumina MiSeq sequencing. This was performed at t0 (representing day 5 since the start of the incubations and just prior to oil enrichment), t1 (representing day 12 since the start of the incubations and 7 days after oil enrichment), and t2 (representing day 19 since the start of the incubations and 14 days after oil enrichment). Cell biomass from samples (5 ml) were taken at these three time points for extraction of whole genomic DNA. The samples were filtered using a vacuum filtration manifold (Millipore Sigma, UK) with 47 mm polycarbonate membrane filters (0.22 μm pore size) and the filters stored at − 20 °C. For extraction of DNA, the filters were crushed in liquid nitrogen to a fine powder and the nitrogen then allowed to evaporate off completely. DNA was extracted using the potassium ethyl xanthogenate method based on that by Tillett and Neilan74. Extracted DNA was confirmed using a Nanodrop 3300 fluorescence spectrometer (ThermoFisher Scientific, UK) and further confirmed by gel electrophoresis. PCR amplification of the V4 16S rRNA gene fragment was performed using Platinum Hot Start PCR Mastermix (ThermoFisher Scientific, UK), as described by the manufacturer’s protocol. Thermocycler conditions were 94 °C for 3 min, 35 cycles of 94 °C for 30 s, 50 °C for 60 s, and 70 °C for 90 s, and then a final extension step at 72 °C for 10 min.
Two to three replicates from each sample were selected and underwent amplicon sequence library preparation using Golay barcoded 515FB forward primers (5’-GTGYCAGCMGCCGCGGTAA-3’) and a complementary 806RB reverse primer (5’-GGACTACNVGGGTWTCTAAT-3’), according to the Earth Microbiome Project75,76. Libraries were pooled and sequenced at Edinburgh Genomics (Edinburgh, UK), using an Illumina Miseq sequencing platform employing the V2 (2 × 250) reagent kit. De-multiplexed and primer trimmed data files were returned for downstream processing, as described below.
Targeted sequencing of the modified V4 region using this pair of 515FB and 806RB improves detection of clade SAR11 in marine samples without affecting detection of taxa previously identified in region 515f/926r75.
Sequencing data analysis
Libraries of bacterial community 16S rRNA gene sequences were constructed for OTU-based analysis using mothur version 1.39.577. Contiguous sequences from paired-end sample reads were constructed, and homopolymers more than 8 bases long were removed. The sequences were then aligned against the SILVA database. Chimeras were removed and sequences that were left were classified and taxonomically referenced to the RDP database (version 16), and similarities below 80% were removed. Variation in species abundance and composition of bacterial communities associated with E. huxleyi in different treatments were analysed with non-metric multidimensional scaling (NMDS) and ANOSIM analysis. Shannon-Weiner diversity was calculated to determine the alpha diversity of treatments and a heat map was generated to visualise similarities of bacterial families in the different treatments. Dominant microbial taxa between treatments were visualised by heat tree plots using metacodeR78. Pairwise comparisons of the heat tree plots were generated by calculating taxon abundance with Wilcox rank-sum test, and Benjamini Hochberg (FDR) correction for multiple comparisons was used to determine significant differences of taxa between treatments.
Hydrocarbon analysis
Microcosms used for hydrocarbon analysis were not sampled for biological analysis and remained untouched until the end of experiment. Each of these microcosms was sacrificed at day 20 for extraction of TPH using HPLC-grade dichloromethane (DCM) at an oil/water mix to DCM ratio of 2:1. The DCM fraction was removed and the oil/water mix re-extracted an additional 3 times. DCM fractions from each microcosm were pooled and further treated as previously described27 to obtain gravimetric data to calculate the original sample weight and the weight of oil remaining, and to prepare the TPH extracts for analysis of aliphatic and aromatic hydrocarbons. Briefly, GC-FID was used for the analysis of aliphatic hydrocarbons, with ratios of nC17/pristine and nC18/phytane used for differentiating biological degradation from weathering79. Similarly, aromatic ratios for biodegradation were compared for naphthalene/2-methylnaphthalene (N/2-MN), 2-methylnaphthalene/1-methylnaphthalene (2MN/1-MN), 2-ethylnaphthalene/2,6 + 2,7-dimethylnaphthalene (2-ET/(2,6 + 2,7)-DMN), 2-methylnaphthalene/2,6 + 2,7-dimethylnaphthalene, phenanthrene/9-methylphenanthrene (P/9-MP), 3 + 2-methylphenanthrene/9 + 1-methylphenanthrene (3-MP + 2-MP/9-MP + 1-MP), and 3-methylphenanthrene/9-methylphenanthrene (3-MP/9MP). These methods were followed as previously described27. One-way ANOVA was carried out on ratios to determine significant differences of hydrocarbon degradation between treatments.
Statistical analysis
The two-way ANOVA test was used to analyse the differences in Chl a concentration of E. huxleyi as well as pH observed in E. huxleyi cultures across the CO2 treatments (i.e. ambient vs elevated CO2), oil-enriched treatments and treatment-time factors using PRISM 9 statistical software (Graphstats Technologies, Karnataka, India). Similarly, aliphatic and aromatic hydrocarbon ratios were also compared using Tukey’s multiple comparison test after two-way ANOVA analysis. To examine bacterial community abundance between treatments, Wilcox rank-sum test followed by Benjamini-Hochberg (FDR) correction for multiple testing was applied to median proportion of OTU reads.
Data availability
All data used and/or analyzed during the current study are presented in the article. For additional data requests, Tony Gutierrez (tony.gutierrez@hw.ac.uk) or Sebastian Hennige (s.hennige@ed.ac.uk) can be contacted.
References
Ducklow, H. W., Steinberg, D. K. & Buesseler, K. O. Upper ocean carbon export and the biological pump. Oceanography 14, 50–58 (2001).
Field, C. B., Behrenfield, M. J., Randerson, J. T. & Falkowski, P. Primary production of the biosphere: Integrating terrestrial oceanic components. Science 281, 237–240 (1998).
Amin, S. A., Parker, M. S. & Ambrust, E. V. Interactions between diatoms and bacteria. Microbiol. Mol. Biol. Rev. 76, 667–684 (2012).
Amin, S. A. et al. Interactions and signalling between a cosmopolitan phytoplankton and associated bacteria. Nature 522, 98–101 (2015).
Buchan, A., LeCleir, G. R., Gulvik, C. A. & Gonzalez, J. M. Master recyclers: Features and functions of bacteria associated with phytoplankton blooms. Nat. Rev. Microbiol. 12, 686–698 (2014).
Bell, W. & Mitchell, R. Chemotactic and growth responses of marine bacteria to algal extracellular products. Biol. Bull. 143, 265–277 (1972).
Myklestad, S. M. Release of extracellular products by phytoplankton with special emphasis on polysaccharides. Sci. Total Environ. 165, 155–164 (1995).
McGenity, T. J., Folwell, B. D., McKew, B. A. & Sanni, G. O. Marine crude-oil biodegradation: A central role for interspecies interactions. Aquat. Biosyst. 8, 10 (2012).
Amin, S. A. et al. Photolysis of ironsiderophore chelates promotes bacterial-algal mutualism. Proc. Natl. Acad. Sci. U. S. A. 106, 17071–17076 (2009).
Kazamia, E. et al. Mutualistic interactions between vitamin B12-dependent algae and heterotrophic bacteria exhibit regulation. Environ. Microbiol. 14, 1466–1476 (2012).
Joint, I., Doney, S. C. & Karl, D. M. Will ocean acidification affect marine microbes?. ISME J. 5, 1–7 (2011).
Coelho, F. J. R. C. et al. Multitaxon activity profiling reveals differential microbial response to reduced seawater pH and oil pollution. Mol. Ecol. 25, 4645–4659 (2016).
O’brien, C. J., Vogt, M. & Gruber, N. Global coccolithophore diversity: Drivers and future change. Prog. Oceanogr. 140, 27–42 (2016).
Riebesell, U. et al. Reduced calcification of marine plankton in response to increased atmospheric CO2. Nature 407, 364–367 (2000).
Wolf-Gladrow, D. A., Riebesell, U., Burkhardt, S. & Buma, J. Direct effects of CO2 concentration on growth and isotopic composition of marine plankton. Tellus B Chem. Phys. Meteorol. 51, 461–476 (1999).
Iglesias-Rodriguez, M. D. et al. Phytoplankton calcification in a high-CO2 world. Science 320, 336–340 (2008).
Krumhardt, K. M., Lovenduski, N. S., Freeman, N. M. & Bates, N. R. Apparent increase in coccolithophore abundance in the subtropical North Atlantic from 1990 to 2014. Biogeosciences 13, 1163–1177 (2016).
Read, B. A. et al. Pan genome of the phytoplankton Emiliania underpins its global distribution. Nature 499, 209–213 (2013).
Riebesell, U. et al. Comment on “Phytoplankton calcification in a high-CO2 world”. Science 322, 1466 (2008).
Marlowe, I. T. et al. Long chain (n-C37–C39) alkenones in the Prymnesiophyceae. Distribution of alkenones and other lipids and their taxonomic significance. Br. Phycol. J. 19, 203–216 (1984).
Yamane, K. et al. Pyrolytic generation of petroleum crude oils from the marine phytomicroalgal coccolithophore Emiliania huxleyi (Haptophyta) and preparation of n-alkane-rich biofuel. Energy Fuels. 27, 7470–7479 (2013).
Green, D. H., Echavarri-Bravo, V., Brennan, D. & Hart, M. C. Bacterial diversity associated with the coccolithophorid algae Emiliania huxleyi and Coccolithus pelagicus f. braarudii. BioMed. Res. Int. 2015, 194540 (2015).
Gutierrez, T. et al. Algiphilus aromaticivorans gen. nov., sp. nov., an aromatic hydrocarbon-degrading bacterium isolated from a culture of the marine dinoflagellate Lingulodinium polyedrum, and proposal of Algiphilaceae fam. Nov.. Int. J. Syst. Evol. Microbiol. 62, 2743–2749 (2012).
Gutierrez, T., Nichols, P. D., Whitman, W. B. & Aitken, M. D. Porticoccus hydrocarbonoclasticus sp. nov., an aromatic hydrocarbon-degrading bacterium identified in laboratory cultures of marine phytoplankton. Appl. Environ. Microbiol. 78, 628–637 (2012).
Gutierrez, T. et al. PAH degradation of phytoplankton-associated Arenibacter and description of Arenibacter algicola sp. nov., an aromatic hydrocarbon-degrading bacterium. Appl. Environ. Microbiol. 80, 618–628 (2014).
Gutierrez, T. et al. Polycyclovorans algicola gen. nov., sp. nov., an aromatic-hydrocarbon-degrading marine bacterium found associated with laboratory cultures of marine phytoplankton. Appl. Environ. Microbiol. 79, 205–214 (2013).
Thompson, H., Angelova, A., Bowler, B., Jones, M. & Gutierrez, T. Enhanced crude oil biodegradative potential of natural phytoplankton-associated hydrocarbonoclastic bacteria. Environ. Microbiol. 19, 2843–2861 (2017).
Crone, T. J. & Tolstoy, M. Magnitude of the 2010 Gulf of Mexico oil leak. Science 330, 634 (2010).
McNutt, M. K., Camilli, R., Crone, T. J. & Shaffer, F. Review of flow rate estimates of the Deepwater Horizon oil spill. Proc. Natl. Acad. Sci. U. S. A. 109, 20260–20267 (2011).
Sikkema, J., De Bont, J. A. M. & Poolman, B. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Rev. 59, 201–222 (1995).
Tukaj, Z., Bohdanowicz, J. & Aksmann, A. A morphometric and stereological analysis of ultrastructural changes in two Scenedesmus (Chlorococcales, Chlorophyta) strains subjected to diesel fuel oil pollution. Phycologia 37, 388–393 (1998).
Soto, C., Hellebust, J. A., Hutchinson, T. C. & Sawa, T. Effect of naphthalene and aqueous crude oil extracts on the green flagellate Chlamydomonas angulosa. I. Growth. Can. J. Bot. 53, 109–117 (1975).
Gonzalez, J. et al. Effect of a simulated oil spill on natural assemblages of marine phytoplankton enclosed in microcosms. Estuar. Coast. Shelf Sci. 83, 265–276 (2009).
Koshikawa, H. et al. Effect of the water-soluble fraction of diesel oil on bacterial and primary production and the trophic transfer to mesozooplankton through a microbial food web in Yangtze estuary, China. Estuar. Coast. Shelf Sci. 71, 68–80 (2007).
Parab, S. R. et al. Effect of Bombay high crude oil and its water-soluble fraction on growth and metabolism of diatom Thalassiosira sp. Ind. J. Mar. Sci. 37, 251–255 (2008).
Riebesell, U. et al. Arctic ocean acidification: pelagic ecosystem and biogeochemical responses during a mesocosm study. Biogeosciences 10, 5619–5626 (2013).
Liu, X. et al. Effects of elevated CO2 on phytoplankton during a mesocosm experiment in the southern eutrophicated coastal water of China. Sci. Rep. 7, 6868 (2017).
Frada, M., Robert, I., Allen, M. J. & de Vargas, C. The, “Cheshire Cat” escape strategy of the coccolithophore Emiliania huxleyi in response to viral infection. Proc. Nat. Acad. Sci. U. S. A. 105, 15944–15949 (2008).
Thornber, C. S. Functional properties of the isomorphic biphasic algal life cycle. Integr. Compar. Biol. 46, 605–614 (2006).
Valero, M., Richerd, S., Perrot, V. & Destombe, C. Evolution of Alternation of haploid and diploid phases in life cycles. Trends Ecol. Evol. 7, 25–29 (1992).
Bevilaqua, R. C. A., Rigo, V. A., Verissimo-Alves, M. & Miranda, C. R. NMR characterization of hydrocarbon adsorption on calcite surfaces: A first principles study. J. Chem. Phys. 141, 204705 (2014).
Vila, J., Nieto, J. M., Mertens, J., Springael, D. & Grifoll, M. Microbial community structure of a heavy fuel oil-degrading marine consortium: Linking microbial dynamics with polycyclic aromatic hydrocarbon utilization. FEMS Microbiol. Ecol. 73, 349–362 (2010).
Gutierrez, T. et al. Hydrocarbon-degrading bacteria enriched by the Deepwater Horizon oil spill identified by cultivation and DNA-SIP. ISME J. 7, 2091–2104 (2013).
Kappell, A. D. et al. The polycyclic aromatic hydrocarbon degradation potential of Gulf of Mexico native coastal microbial communities after the Deepwater Horizon oil spill. Front. Microbiol. 5, 205 (2014).
Evans, M. V. et al. Members of Marinobacter and Arcobacter influence system biogeochemistry during early production of hydraulically fractured natural gas wells in the Appalachian Basin. Front. Microbiol. 9, 2646 (2018).
Schoffman, H., Lis, H., Shaked, Y. & Keren, N. Iron-nutrient interactions within phytoplankton. Front. Plant Sci. 7, 1223 (2016).
Tortell, P. D., Maldonado, M. T., Granger, J. & Price, N. M. Marine bacteria and biogeochemical cycling of iron in the oceans. FEMS Microbiol. Ecol. 29, 1–11 (1999).
Green, P. N. Methylobacterium. In The Prokaryotes (eds Dworkin, M. et al.) 257–326 (Springer, New York, 2006).
Mukherjee, A. et al. Bioinformatic approaches including predictive metagenomic profiling reveal characteristics of bacterial response to petroleum hydrocarbon contamination in diverse environments. Sci. Rep. 7, 1108 (2017).
Janvier, M. & Grimont, P. A. D. The genus Methylophaga, a new line of descent within phylogenetic branch gamma of Proteobacteria. Res. Microbiol. 146, 543–550 (1995).
Gutierrez, T. & Aitken, M. D. Role of methylotrophs in the degradation of hydrocarbons during the Deepwater Horizon oil spill. ISME J. 8, 2543–2545 (2014).
Mishamandani, S., Gutierrez, T. & Aitken, M. D. DNA-based stable isotope probing coupled with cultivation methods implicates Methylophaga in hydrocarbon degradation. Front. Microbiol. 5, 76 (2014).
Mishamandani, T., Gutierrez, T., Berry, D. & Aitken, M. Response of the bacterial community associated with a cosmopolitan marine diatom to crude oil shows a preference for the biodegradation of aromatic hydrocarbons. Environ. Microbiol. 18, 1817–1833 (2016).
Gutierrez, T. Occurrence and roles of the obligate hydrocarbonoclastic bacteria in the ocean when there is no obvious hydrocarbon contamination. In Handbook of Hydrocarbon and Lipid Microbiology Vol. 6 (eds McGenity, T. J. & Prince, R.) (Springer, Berlin, 2018).
McCarren, J. et al. Microbial community transcriptomes reveal microbes and metabolic pathways associated with dissolved organic matter turnover in the sea. Proc. Natl. Acad. Sci. U. S. A. 107, 16420–16427 (2010).
Sosa, O. A., Gifford, S. M., Repeta, D. J. & DeLong, E. F. High molecular weight dissolved organic matter enrichment selects for methylotrophs in dilution to extinction cultures. ISME J. 9, 2725–2739 (2015).
Borchard, C. & Engel, A. Organic matter exudation by Emiliania huxleyi under simulated future ocean conditions. Biogeosciences 9, 3405–3423 (2012).
Balkwill, D. L., Fredrickson, J. K. & Romine, M. F. Sphingomonas and related genera. In The Prokaryotes (eds Dworkin, M. et al.) 605–629 (Springer, New York, 2006).
Hassenrück, C. et al. Quantification of the effects of ocean acidification on sediment microbial communities in the environment: The importance of ecosystem approaches. FEMS Microbiol. Ecol. 92, fiw027 (2016).
Witt, V., Wild, C., Anthony, K. R. N., Diaz-Pulido, G. & Uthicke, S. Effects of ocean acidification on microbial community composition of, and oxygen fluxes through, biofilms from the Great Barrier Reef. Environ. Microbiol. 13, 2976–2989 (2011).
Chaerun, S. K., Tazaki, K., Asada, R. & Kogure, K. Bioremediation of coastal areas 5 years after the Nakhodka oil spill in the Sea of Japan: Isolation and characterization of hydrocarbon-degrading bacteria. Environ. Int. 30, 911–922 (2004).
Lin, Y., Lay, J.-J. & Shieh, W. K. Diesel degradation in soil catalyzed by the addition of a bioagent. Int. J. Environ. Sci. Technol. 13, 551–560 (2016).
Moussa, H. A., Wasfi, R., Abdeltawab, N. F. & Megahed, S. A. High counts and anthracene degradation ability of Streptococcus mutans and Veillonella parvula isolated from the oral cavity of cigarette smokers and non-smokers. Front. Microbiol. 12, 661509 (2021).
Xu, X. et al. Petroleum hydrocarbon-degrading bacteria for the remediation of oil pollution under aerobic conditions: A perspective analysis. Front. Microbiol. 9, 2885 (2018).
Brzeszcz, J. & Kaszycki, P. Aerobic bacteria degrading both n-alkanes and aromatic hydrocarbons: An undervalued strategy for metabolic diversity and flexibility. Biodegradation 29, 359–407 (2018).
Cui, Z. et al. Marinobacter aromaticivorans sp. nov., a polycyclic aromatic hydrocarbon-degrading bacterium isolated from sea sediment. Int. J. Syst. Evol. Microbiol. 66, 353–359 (2016).
Duran, R. Marinobacter. In Handbook of Hydrocarbon and Lipid Microbiology (ed. Timmis, K. N.) (Springer, Berlin, 2010).
Gao, W. et al. Marinobacter nanhaiticus sp. nov., polycyclic aromatic hydrocarbon-degrading bacterium isolated from the sediment of the South China Sea. Antonie Van Leeuwenhoek 103, 485–491 (2013).
Head, I. M., Jones, D. M. & Röling, W. F. M. Marine microorganisms make a meal of oil. Nat. Rev. Microbiol. 4, 173–182 (2006).
Guillard, R. R. & Ryther, J. H. Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt, and Detonula confervacea (cleve) Gran. Can. J. Microbiol. 8, 229–239 (1962).
Riebesell, U., Fabry, V. J., Hansson, L. & Gattuso, J.-P. (eds.) Guide to best practices for ocean acidification research and data reporting. Office for Official Publications of the European Communities, Luxembourg. (European Union, Belgium, 2011). https://doi.org/10.2777/66906.
Arar, E. J. & Collins, G. B. Method 445.0 In vitro determination of chlorophyll a and pheophytin ain marine and freshwater algae by fluorescence. US Environmental Protection Agency, Washington, DC. https://cfpub.epa.gov/si/si_public_record_report.cfm?dirEntryId=309417 (1997).
Welschmeyer, N. A. Fluorometric analysis of chlorophyll a in the presence of chlorophyll b and pheopigments. Limnol. Oceanogr. 39, 1985–1992 (1994).
Tillett, D. & Neilan, B. A. Xanthogenate nucleic acid isolation from cultured and environmental cyanobacteria. J. Phycol. 36, 251–258 (2000).
Apprill, A., McNally, S., Parsons, R. & Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 75, 129–137 (2015).
Caporaso, J. G. et al. EMP 16S Illumina amplicon protocol. https://www.protocols.io/view/emp-16s-illumina-amplicon-protocol-nuudeww (2018).
Schloss, P. D. et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541 (2009).
Foster, Z. S. L., Sharpton, T. J. & Grünwald, N. J. Metacoder: An R package for visualization and manipulation of community taxonomic diversity data. PLoS Comput. Biol. 13, e1005404 (2017).
Olson, G. M., Gao, H., Meyer, B. M., Miles, M. S. & Overton, E. B. Effect of Corexit 9500A on Mississippi Canyon crude oil weathering patterns using artificial and natural seawater. Heliyon 3, e00269 (2017).
Acknowledgements
This manuscript contains work conducted during a PhD study sponsored by a studentship to AMF from the University of Malaysia, Terengganu (UMT) and whose support is gratefully acknowledged. Support was also provided through two Natural Environment Research Council grants (NERC, NE/K009028/1 and NE/K009028/2) to SH. We would also like to thank BP for providing the Schiehallion crude oil, and the two anonymous reviewers for their valuable comments during the preparation of the manuscript. The authors declare no conflict of interest.
Author information
Authors and Affiliations
Contributions
S.H. and T.G. contributed to the conception and design of this study with contributions from A.M.F. A.M.F., S.S., B.B. and M.J. conducted laboratory work and/or data analysis with guidance from S.H. and T.G. T.G. wrote the manuscript with contributions from A.M.F. and S.H. All authors approved the final manuscript submission.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fahmi, A.M., Summers, S., Jones, M. et al. Effect of ocean acidification on the growth, response and hydrocarbon degradation of coccolithophore-bacterial communities exposed to crude oil. Sci Rep 13, 5013 (2023). https://doi.org/10.1038/s41598-023-31784-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-31784-5
- Springer Nature Limited