
Abstract
Cancer cells consistently utilize the unfolded protein response (UPR) to encounter the abnormal endoplasmic reticulum (ER) stress induced by the accumulation of misfolded proteins. Extreme activation of the UPR could also provoke maladaptive cell death. Previous reports have shown that NRF2 antioxidant signaling is activated by UPR and serves as noncanonical pathway to defense and reduce excessive ROS levels during ER stress. However, the mechanisms of regulating NRF2 signaling upon ER stress in glioblastoma have not been fully elucidated. Here we identify that SMURF1 protects against ER stress and facilitates glioblastoma cell survival by rewiring KEAP1-NRF2 pathway. We show that ER stress induces SMURF1 degradation. Knockdown of SMURF1 upregulates IRE1 and PERK signaling in the UPR pathway and prevents ER-associated protein degradation (ERAD) activity, leading to cell apoptosis. Importantly, SMURF1 overexpression activates NRF2 signaling to reduce ROS levels and alleviate UPR-mediated cell death. Mechanistically, SMURF1 interacts with and ubiquitinates KEAP1 for its degradation (NRF2 negative regulator), resulting in NRF2 nuclear import. Moreover, SMURF1 loss reduces glioblastoma cell proliferation and growth in subcutaneously implanted nude mice xenografts. Taken together, SMURF1 rewires KEAP1-NRF2 pathway to confer resistance to ER stress inducers and protect glioblastoma cell survival. ER stress and SMURF1 modulation may provide promising therapeutic targets for the treatment of glioblastoma.
Similar content being viewed by others

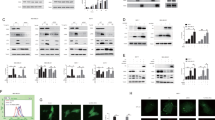
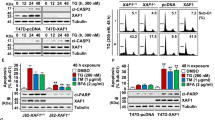
Introduction
Glioblastoma is the most malignant primary brain tumor, characterized by high heterogeneity, resistance and relapse [1, 2]. The current treatment strategies including maximal-safe surgical resection and adjuvant radiation therapy with alkylating agent temozolomide (TMZ) treatment have shown limited survival benefits, with a median survival rate about 14.6 months [3, 4]. Glioblastoma cells have a high metabolic rate and produce high levels of reactive oxide species (ROS), which further disturbs the protein folding capacity [5]. Endoplasmic reticulum (ER) stress due to the accumulation of unfolded or misfolded proteins is present highly in glioblastoma [6, 7]. The mechanisms are involved in response to ER stress by the three UPR sensors, including activating transcription factor 6 alpha (ATF6α), inositol requiring enzyme 1 (IRE1) and protein kinase R (PKR)-like ER kinase (PERK). ER chaperone BiP/GRP78 constitutively binds to the three UPR sensors, resulting in their inactivation at the basal level. Under ER stress, BiP/GRP78 dissociates from the UPR sensors to activate IRE1, PERK and ATF6α by oligomerization and trans-autophosphorylation or secretion of ATF6α [8, 9]. UPR response is an adaptive mechanism to restore ER homeostasis through multiple pathways, including attenuating transcriptional signal, alleviating the accumulation of misfolded proteins via ER-associated protein degradation (ERAD) system and recycling of misfolded proteins through the induction of autophagy [8, 10,11,12]. Accumulating evidences have indicated that a high level of basal UPR is frequently found in primary human tumors including glioblastoma, and adaptive UPR promotes cancer survival upon adverse environments [13,14,15]. However, the aberrant activation of UPR also triggers cell death under the unresolved and extreme ER stress conditions [16, 17]. For instance, arginosuccinate synthase 1 treatment suppresses tumor progression and triggers pro-apoptotic ER stress responses in hepatocellular carcinoma through the hyperactivation of PERK-eukaryotic translation initiation factor 2α (eIF2α)-activating transcription factor 4 (ATF4)-C/EBP homologous protein (CHOP) arm of the UPR [18].
Misfolded protein accumulation and aggregation induce excessive production of ROS, which can activate nuclear factor erythroid 2-related factor 2 (NRF2). Many lines of evidences have shown that the activation of NRF2 pathway suppresses ER stress-induced apoptosis by regulating antioxidant synthesis and ROS eliminating enzymes expression [19, 20]. Under normal conditions, NRF2 is sequestered in the cytoplasm and degraded by its negative factor Kelch-like ECH-associated protein 1 (KEAP1) [21]. Whereas upon oxidative stress, the canonical KEAP1-NRF2 pathway activation is mediated by the release of NRF2 from KEAP1, then NRF2 translocates into the nucleus to activate antioxidant genes such as NAD(P)H quinone dehydrogenase 1 (NQO1) and HMOX1/HO-1 heme oxygenase 1 (HO1) [14, 22]. The noncanonical KEAP1-NRF2 pathway activation is mediated by p62/SQSTM1, an autophagy receptor protein that competitively binds with and degrades KEAP1 to activate NRF2 [22,23,24]. Indeed, NRF2-p62 system and selective autophagy are vital in tolerance of tumor microenvironmental stress [25, 26]. ER stress inducing agents, such as ER Ca2+ pump inhibitor TG (Thapsigargin) and the N-glycosylation inhibitor TM (Tunicamycin), increase ROS production and trigger tumor cell apoptosis [20]. NRF2 could be phosphorylated by PERK during ER stress, further triggering NRF2 dissociation from KEAP1 and induction of protective antioxidant response [27]. In addition, PERK-dependent activation of NRF2 attenuates accumulation of ROS triggering oxidative DNA damage and contributes to redox homeostasis and cell survival [28, 29]. Importantly, studies have reported that activation of NRF2 induces several components of the transcriptional UPR target genes, including XBP1 and ATF6α, to maintain ER integrity and protein homeostasis [30]. NRF2 interacts and activates ATF4 to induce the target genes expression to survive proteotoxic stress [31, 32]. Therefore, NRF2-UPR axis serves as a bidirectional signal for maintaining ER homeostasis. Accumulating evidences have suggested that sustained activation of NRF2 also induces pro-survival genes that promote cancer cell proliferation and chemoresistance [33]. Recent studies have shown that aberrantly high expression of NRF2 signaling is found in glioblastoma and promotes tumor cell mesenchymal transition, invasion and tumorigenesis [34]. However, the mechanisms of regulating NRF2 signaling in glioblastoma have not been well defined.
The HECT-type ubiquitin ligase (E3) Smad ubiquitination regulatory factor 1 (SMURF1) belongs to the Nedd4 family and mediates multiple biological processes, including cell growth and migration, and several physiological functions in bone formation, embryonic development, and tumorigenesis [35,36,37,38,39]. Our previous studies have demonstrated that SMURF1 is hyperactivated in glioblastoma and promotes tumor growth by ubiquitination and degradation of tumor suppresser phosphatase and tensin homolog (PTEN) [ The datasets generated during the current study are available from the corresponding author on reasonable request. Ryskalin L, Gaglione A, Limanaqi F, Biagioni F, Familiari P, Frati A, et al. The autophagy status of cancer stem cells in gliobastoma multiforme: from cancer promotion to therapeutic strategies. Int J Mol Sci. 2019;20:3824. Yang K, Niu L, Bai Y, Le W. Glioblastoma: Targeting the autophagy in tumorigenesis. Brain Res Bull. 2019;153:334–40. Van Meir EG, Hadjipanayis CG, Norden AD, Shu HK, Wen PY, Olson JJ. Exciting new advances in neuro-oncology: the avenue to a cure for malignant glioma. CA Cancer J Clin. 2010;60:166–93. Stupp R, Hegi ME, Mason WP, van den Bent MJ, Taphoorn MJ, Janzer RC, et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009;10:459–66. Olivier C, Oliver L, Lalier L, Vallette FM. Drug resistance in glioblastoma: the two faces of oxidative stress. Front Mol Biosci. 2020;7:620677. Penaranda-Fajardo NM, Meijer C, Liang Y, Dijkstra BM, Aguirre-Gamboa R, den Dunnen WFA, et al. ER stress and UPR activation in glioblastoma: identification of a noncanonical PERK mechanism regulating GBM stem cells through SOX2 modulation. Cell Death Dis. 2019;10:690. Penaranda Fajardo NM, Meijer C, Kruyt FA. The endoplasmic reticulum stress/unfolded protein response in gliomagenesis, tumor progression and as a therapeutic target in glioblastoma. Biochem Pharm. 2016;118:1–8. Chevet E, Hetz C, Samali A. Endoplasmic reticulum stress-activated cell reprogramming in oncogenesis. Cancer Discov. 2015;5:586–97. Rashid HO, Yadav RK, Kim HR, Chae HJ. ER stress: Autophagy induction, inhibition and selection. Autophagy. 2015;11:1956–77. Gonzalez-Quiroz M, Urra H, Limia CM, Hetz C. Homeostatic interplay between FoxO proteins and ER proteostasis in cancer and other diseases. Semin Cancer Biol. 2018;50:42–52. Wiseman RL, Mesgarzadeh JS, Hendershot LM. Resha** endoplasmic reticulum quality control through the unfolded protein response. Mol Cell. 2022;82:1477–91. Marciniak SJ, Chambers JE, Ron D. Pharmacological targeting of endoplasmic reticulum stress in disease. Nat Rev Drug Discov. 2022;21:115–40. Chen X, Cubillos-Ruiz JR. Endoplasmic reticulum stress signals in the tumour and its microenvironment. Nat Rev Cancer. 2021;21:71–88. Clarke HJ, Chambers JE, Liniker E, Marciniak SJ. Endoplasmic reticulum stress in malignancy. Cancer Cell. 2014;25:563–73. Kang BR, Yang SH, Chung BR, Kim W, Kim Y. Cell surface GRP78 as a biomarker and target for suppressing glioma cells. Sci Rep. 2016;6:34922. Hetz C. The unfolded protein response: controlling cell fate decisions under ER stress and beyond. Nat Rev Mol Cell Biol. 2012;13:89–102. Hetz C, Zhang K, Kaufman RJ. Mechanisms, regulation and functions of the unfolded protein response. Nat Rev Mol Cell Biol. 2020;21:421–38. Kim S, Lee M, Song Y, Lee SY, Choi I, Park IS, et al. Argininosuccinate synthase 1 suppresses tumor progression through activation of PERK/eIF2alpha/ATF4/CHOP axis in hepatocellular carcinoma. J Exp Clin Canc Res. 2021;40:127. Teng Y, Zhao H, Gao L, Zhang W, Shull AY, Shay C. FGF19 protects hepatocellular carcinoma cells against endoplasmic reticulum stress via activation of FGFR4-GSK3beta-Nrf2 signaling. Cancer Res. 2017;77:6215–25. Liu Y, Tao S, Liao L, Li Y, Li H, Li Z, et al. TRIM25 promotes the cell survival and growth of hepatocellular carcinoma through targeting Keap1-Nrf2 pathway. Nat Commun. 2020;11:348. Lu K, Alcivar AL, Ma J, Foo TK, Zywea S, Mahdi A, et al. NRF2 induction supporting breast cancer cell survival Is enabled by oxidative stress-induced DPP3-KEAP1 interaction. Cancer Res. 2017;77:2881–92. Silva-Islas CA, Maldonado PD. Canonical and non-canonical mechanisms of Nrf2 activation. Pharm Res. 2018;134:92–9. Komatsu M, Kurokawa H, Waguri S, Taguchi K, Kobayashi A, Ichimura Y, et al. The selective autophagy substrate p62 activates the stress responsive transcription factor Nrf2 through inactivation of Keap1. Nat Cell Biol. 2010;12:213–23. Bae SH, Sung SH, Oh SY, Lim JM, Lee SK, Park YN, et al. Sestrins activate Nrf2 by promoting p62-dependent autophagic degradation of Keap1 and prevent oxidative liver damage. Cell Metab. 2013;17:73–84. Jena KK, Kolapalli SP, Mehto S, Nath P, Das B, Sahoo PK, et al. TRIM16 controls assembly and degradation of protein aggregates by modulating the p62-NRF2 axis and autophagy. EMBO J. 2018;37:e98358. Ichimura Y, Waguri S, Sou YS, Kageyama S, Hasegawa J, Ishimura R, et al. Phosphorylation of p62 activates the Keap1-Nrf2 pathway during selective autophagy. Mol Cell. 2013;51:618–31. Cullinan SB, Zhang D, Hannink M, Arvisais E, Kaufman RJ, Diehl JA. Nrf2 is a direct PERK substrate and effector of PERK-dependent cell survival. Mol Cell Biol. 2003;23:7198–209. Bobrovnikova-Marjon E, Grigoriadou C, Pytel D, Zhang F, Ye J, Koumenis C, et al. PERK promotes cancer cell proliferation and tumor growth by limiting oxidative DNA damage. Oncogene. 2010;29:3881–95. Cullinan SB, Diehl JA. PERK-dependent activation of Nrf2 contributes to redox homeostasis and cell survival following endoplasmic reticulum stress. J Biol Chem. 2004;279:20108–17. Glover-Cutter KM, Lin S, Blackwell TK. Integration of the unfolded protein and oxidative stress responses through SKN-1/Nrf. PLoS Genet. 2013;9:e1003701. He CH, Gong P, Hu B, Stewart D, Choi ME, Choi AM, et al. Identification of activating transcription factor 4 (ATF4) as an Nrf2-interacting protein. Implication for heme oxygenase-1 gene regulation. J Biol Chem. 2001;276:20858–65. Zhu YP, Zheng Z, Hu S, Ru X, Fan Z, Qiu L, et al. Unification of opposites between two antioxidant transcription factors Nrf1 and Nrf2 in mediating distinct cellular responses to the endoplasmic reticulum stressor tunicamycin. Antioxid (Basel, Switz). 2019;9:4. Wu S, Lu H. Nrf2 in cancers: A double-edged sword. Cancer Med. 2019;8:2252–67. Polonen P, Jawahar Deen A, Leinonen HM, Jyrkkanen HK, Kuosmanen S, Mononen M, et al. Nrf2 and SQSTM1/p62 jointly contribute to mesenchymal transition and invasion in glioblastoma. Oncogene. 2019;38:7473–90. Tao Y, Sun C, Zhang T, Song Y. SMURF1 promotes the proliferation, migration and invasion of gastric cancer cells. Oncol Rep. 2017;38:1806–14. Fu L, Cui CP, Zhang X, Zhang L. The functions and regulation of Smurfs in cancers. Semin Cancer Biol. 2020;67:102–16. Cao Y, Zhang L. A Smurf1 tale: function and regulation of an ubiquitin ligase in multiple cellular networks. Cell Mol Life Sci. 2013;70:2305–17. Chen X, Chen S, Li Y, Gao Y, Huang S, Li H, et al. SMURF1-mediated ubiquitination of ARHGAP26 promotes ovarian cancer cell invasion and migration. Exp Mol Med. 2019;51:1–12. Yang H, Yu N, Xu J, Ding X, Deng W, Wu G, et al. SMURF1 facilitates estrogen receptor a signaling in breast cancer cells. J Exp Clin Canc Res. 2018;37:24. **a Q, Zhang H, Zhang P, Li Y, Xu M, Li X, et al. Oncogenic Smurf1 promotes PTEN wild-type glioblastoma growth by mediating PTEN ubiquitylation. Oncogene 2020;39:5902–15. Dong L, Liu L, Li Y, Li W, Zhou L, **a Q. E3 ligase Smurf1 protects against misfolded SOD1 in neuronal cells by promoting its K63 ubiquitylation and aggresome formation. Hum Mol Genet. 2022;31:2035–48. Franco LH, Nair VR, Scharn CR, Xavier RJ, Torrealba JR, Shiloh MU, et al. The ubiquitin ligase Smurf1 functions in selective autophagy of mycobacterium tuberculosis and anti-tuberculous host defense. Cell Host Microbe. 2017;21:59–72. Feng X, Jia Y, Zhang Y, Ma F, Zhu Y, Hong X, et al. Ubiquitination of UVRAG by SMURF1 promotes autophagosome maturation and inhibits hepatocellular carcinoma growth. Autophagy. 2019;15:1130–49. Guo X, Shen S, Song S, He S, Cui Y, **ng G, et al. The E3 ligase Smurf1 regulates Wolfram syndrome protein stability at the endoplasmic reticulum. J Biol Chem. 2011;286:18037–47. Haynes CM, Titus EA, Cooper AA. Degradation of misfolded proteins prevents ER-derived oxidative stress and cell death. Mol Cell. 2004;15:767–76. Hauck AK, Huang Y, Hertzel AV, Bernlohr DA. Adipose oxidative stress and protein carbonylation. J Biol Chem. 2019;294:1083–8. Hwang J, Qi L. Quality control in the endoplasmic reticulum: crosstalk between ERAD and UPR pathways. Trends Biochem Sci. 2018;43:593–605. Wu J, Kaufman RJ. From acute ER stress to physiological roles of the Unfolded Protein Response. Cell Death Differ. 2006;13:374–84. Obacz J, Avril T. Endoplasmic reticulum proteostasis in glioblastoma-From molecular mechanisms to therapeutic perspectives. Sci Signal. 2017;10:eaal2323. Lei K, Davis RJ. JNK phosphorylation of Bim-related members of the Bcl2 family induces Bax-dependent apoptosis. Proc Natl Acad Sci USA. 2003;100:2432–7. Zhu Y, **e M, Meng Z, Leung LK, Chan FL, Hu X, et al. Knockdown of TM9SF4 boosts ER stress to trigger cell death of chemoresistant breast cancer cells. Oncogene. 2019;38:5778–91. Liu Q, Yang X, Long G, Hu Y, Gu Z, Boisclair YR, et al. ERAD deficiency promotes mitochondrial dysfunction and transcriptional rewiring in human hepatic cells. J Biol Chem. 2020;295:16743–53. Xu H, Liu P, Yan Y, Fang K, Liang D, Hou X, et al. FKBP9 promotes the malignant behavior of glioblastoma cells and confers resistance to endoplasmic reticulum stress inducers. J Exp Clin Canc Res. 2020;39:44. Lu Z, Zhou L, Killela P, Rasheed AB, Di C, Poe WE, et al. Glioblastoma proto-oncogene SEC61gamma is required for tumor cell survival and response to endoplasmic reticulum stress. Cancer Res. 2009;69:9105–11. Song S, Tan J, Miao Y, Zhang Q. Crosstalk of ER stress-mediated autophagy and ER-phagy: Involvement of UPR and the core autophagy machinery. J Cell Physiol. 2018;233:3867–74. Zielke S, Kardo S, Zein L, Mari M, Covarrubias-Pinto A, Kinzler MN, et al. ATF4 links ER stress with reticulophagy in glioblastoma cells. Autophagy. 2021;17:2432–48. Wang J, Qi Q, Zhou W, Feng Z, Huang B, Chen A, et al. Inhibition of glioma growth by flavokawain B is mediated through endoplasmic reticulum stress induced autophagy. Autophagy. 2018;14:2007–22. **pell E, Aragon T, Martinez-Velez N, Vera B, Idoate MA, Martinez-Irujo JJ, et al. Endoplasmic reticulum stress-inducing drugs sensitize glioma cells to temozolomide through downregulation of MGMT, MPG, and Rad51. Neuro Oncol. 2016;18:1109–19. We thank Analysis & Testing Center, Bei**g Institute of Technology and Center, School of Life Science, Bei**g Institute of Technology, Collaborative Research Fund of Chinese Institute for Brain Research, Bei**g (No: 2021-NKX-XM-06), the Shunyi District of Bei**g construction special project in from Bei**g scientific and technological achievements transformation and service platform 2021. This work was supported by grants from the National Natural Science Foundation of China (U21A20200 and 81870123), The Natural Science Foundation of Bei**g Municipality (Z190018), Bei**g Institute of Technology Research Fund Program for Young Scholars (XSQD-202110002), and the National Science Foundation for Young Scientists of China (81902545). QX conceived of the study. MCX designed and performed the experiments. LD and QX analyzed the data. LD wrote the manuscript with input from MCX. YL, WTX, HFZ and CWW contributed to revising the manuscript. The authors declare no competing interests. Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. Edited by Cristina Munoz-Pinedo Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Dong, L., Xu, M., Li, Y. et al. SMURF1 attenuates endoplasmic reticulum stress by promoting the degradation of KEAP1 to activate NRF2 antioxidant pathway.
Cell Death Dis 14, 361 (2023). https://doi.org/10.1038/s41419-023-05873-2 Received: Revised: Accepted: Published: DOI: https://doi.org/10.1038/s41419-023-05873-2 Chinese Journal of Integrative Medicine (2024)Data availability
References
Acknowledgements
Funding
Author information
Authors and Affiliations
Contributions
Corresponding author
Ethics declarations
Competing interests
Additional information
Rights and permissions
About this article
Cite this article
This article is cited by
Therapeutic Role of Chinese Medicine Targeting Nrf2/HO-1 Signaling Pathway in Myocardial Ischemia/Reperfusion Injury
