
Abstract
Septins as GTPases in the cytoskeleton, are linked to a broad spectrum of cellular functions, including cell migration and the progression of hepatocellular carcinoma (HCC). However, roles of SEPT11, the new member of septin, have been hardly understood in HCC. In the study, the clinical significance and biological function of SEPT11 in HCC was explored. SEPT11 was screened out by combining ATAC-seq with mRNA-seq. Role of SEPT11 in HCC was further investigated by using overexpression, shRNA and CRISPR/Cas9-mediated SEPT11-knockout cells or in vivo models. We found RNA-seq and ATAC-seq highlights LncRNA AY927503 (AY) induced SEPT11 transcription, resulting in Rho GTPase activation and cytoskeleton actin aggregation. The GTP-binding protein SEPT11 is thus considered, as a downstream factor of AY, highly expressed in various tumors, including HCC, and associated with poor prognosis of the patients. In vitro, SEPT11 overexpression promotes the migration and invasion of HCC cells, while SEPT11-knockout inhibits migration and invasion. In vivo, SEPT11-overexpressed HCC cells show high metastasis incidents but don’t significantly affect proliferation. Meanwhile, we found SEPT11 targets RhoA, thereby regulating cytoskeleton rearrangement and abnormal cell adhesion through ROCK1/cofilin and FAK/paxillin signaling pathways, promoting invasion and migration of HCC. Further, we found SEPT11 facilitates the binding of GEF-H1 to RhoA, which enhances the activity of RhoA. Overall, our study confirmed function of SEPT11 in promoting metastasis in HCC, and preliminarily explored its related molecular mechanism. SEPT11 acts as an oncogene in HCC, also draws further interest regarding its clinical application as a potential therapeutic target.
Similar content being viewed by others


Introduction
Hepatocellular carcinoma (HCC) accounts for 90% of the cases of primary liver cancer and is the leading cause of cancer-related death worldwide [1]. At present, most HCC patients are often accompanied by regional spread and metastasis, and the prognosis of patients is poor [2, 3]. Therefore, there is an urgent need to investigate the cellular biology and molecular mechanisms of HCC metastasis, providing opportunities for improving clinical outcomes and develo** better treatment options.
Long non-coding RNAs (LncRNAs) are closely associated with the progression of cancer. Hundreds of HCC-related LncRNAs regulate gene expression through different patterns, thereby exerting the function as oncogenes or tumor suppressor genes in HCC [4]. Our previous study has explored the function of AY as an oncogene in the progression of HCC [5], but the downstream regulatory signals of its function were unclear.
The development of tumor cells that acquire metastatic potential into a more motile and invasive phenotype is a critical step in tumor metastasis, and these processes require cytoskeleton rearrangement and abnormal cell adhesion [6, 7]. Many evidences suggest that some Rho GTPases, especially RhoA, Rac1 and Cdc42, are key regulators of the process [8, 9]. Rho GTPases are mainly involved in cell morphological changes, cell adhesion and regulation of cytoskeleton reorganization, which are essential for invasion and migration of tumor cells [10]. According to reports, LncRNAs can regulate the cytoskeleton directly or may influence the cytoskeleton via Rho/ROCK signaling during tumor migration [11].
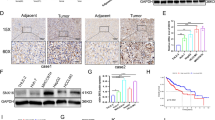
Septins are highly conserved cytoskeletal GTPases, closely associated with actin, microtubule and associated motion. Recently, septins have been identified as the fourth major component of the cytoskeleton, involved in major membrane motility processes such as cytokinesis, vesicle trafficking and exocytosis [12, 13]. Various septins have been reported to play a role in cancer progression [14,15,SEPT11 promotes HCC progression by regulating of actin cytoskeleton through activated RhoA The analysis of the sequencing results above suggested that the function of SEPT11 may be related to regulation of actin cytoskeleton and Rho protein signal transduction. In next analyses, GSEA results indicated that in the high SEPT11 samples were found to be enriched in gene sets related to regulation of actin cytoskeleton (p < 0.001) (Fig. 4A), focal adhesion and FC-gamma-r-mediated-phagocytosis (p < 0.001) (Supplementary Fig. 4A, B), three pathways all contain Rho proteins, which played an important role in cell migration and invasion. Meanwhile, the close connection between Septins and Rho proteins has been extensively reported [30, 32, 33], Therefore, we make a reasonable hypothesis that SEPT11 regulates the cytoskeleton to promote the progress of HCC, and this function is related to Rho protein signal. A GSEA analysis of high expression of SEPT11 in TCGA database affects the enrichment of genes in regulatory of actin cytoskeleton pathway; B WB experiment detects the effect of SEPT11 expression on actin dynamics in two types of cells; C Cell immunofluorescence was used to detect the effect of SEPT11 on actin content and arrangement; D The effect of KO-SEPT11 on the content of several major Rho proteins; E Rho pulldown activity assay kit was used to quantify the activities of RhoA, CDC42, and RAC1 in SEPT11 overexpressing cells; F The effect of AY expression on the activity of RhoA; G The co-localization of RhoA and SEPT11 in cells was observed by immunofluorescence combined with confocal, the arrows indicate puncta of SEPT11 co-localization with RhoA; H WB experiment detects the effect of RhoA inhibitor Rhosin on actin dynamics; I The effect of RhoA inhibitor Rhosin on the content and arrangement of actin was detected by cellular immunofluorescence. E The effect of RhoA inhibitor Rhosin on the content and arrangement of actin was detected by cellular immunofluorescence. Following the hypothesis, we examined the effect of SEPT11 on the dynamic regulation of actin in HepG2 and Huh7 cells. The remodeling of actin filament (F-actin) is an important part of cytoskeletal rearrangement, which provides the impetus for cell invasion and migration [34]. The ratio of F-actin to G-actin can reflect the balance between actin polymerization and depolymerization. The results showed that SEPT11 increased the ratio of F/G-actin, while KO-SEPT11 decreased it (Fig. 4B). At the same time, cellular immunofluorescence staining showed that overexpression of SEPT11 significantly increased actin fibers, thickened and ordered the fiber structure compared with the control, while the actin fibers decreased and disarranged in the KO-SEPT11 group. Overexpression of AY, the upstream regulator of SEPT11, significantly increased the number of actin fibers, but in the AY + KO-SEPT11 group, actin fibers did not become thickened and orderly due to the knockout of SEPT11. FCF, an inhibitor of SEPT11, reduced actin fibers and even affected cell morphology, and significantly inhibited actin fiber thickening caused by overexpression of SEPT11 (Fig. 4C). These results suggest that SEPT11 increased HCC progression by regulating of actin cytoskeleton. Then, in KO-SEPT11 cells, we detected Rho proteins. The results showed that the contents of RhoA, CDC42 and RAC1 decreased after KO-SEPT11, the decrease of RhoA was the most obvious, and there was no significant change in RhoB and RhoC (Fig. 4D). Considering that Rho protein is activated to regulate its downstream signaling [35]. We therefore used Rho pulldown activation assay kit to quantify RhoA, CDC42 and RAC1 activity in SEPT11 overexpression cell. The results showed that SEPT11 promoted the activation of RhoA, but had little effect on CDC42 and RAC1 activity (Fig. 4E). Meanwhile, we found that AY could promote the activation of RhoA, but not in KO-SEPT11 cells (Fig. 4F). Cellular immunofluorescence observation with confocal microscopy showed that SEPT11 and RhoA were obviously co-localization in the two kinds of cells (Fig. 4G), but Co-IP did not find endogenous binding of SEPT11 to RHOA (Supplementary Fig. 4C). Next, we conducted Co-IP assays with exogenous tag plasmid to further confirm whether septin11 and RhoA have an interaction. Through the structural analysis of SEPT11, SEPT11 is consisted of three domains, including NTE (N-terminal extension), GTP binding (GTP-binding domain), and CCD (Coiled coil domain) (Supplementary Fig. 4D). We have constructed a full-length and multifunctional Flag-SEPT11 plasmid for Co-IP assay. It was found that SEPT11 did not form a firm complex with RhoA to influence its activity (Supplementary Fig. 4E). This may be due to the binding between them short-lived and relatively weak, or it works through intermediate factors. We further treated the cells with Rhosin, a specific inhibitor of RhoA, and found that it could significantly reduce the ratio of F/G-actin and abolish the increased ratio caused by overexpression of SEPT11 (Fig. 4H). At the same time, Rhosin treatment reduced the number of actin fibers and disarranged them, and abolished the phenomenon of structural thickening and ordering of actin fibers caused by overexpression of SEPT11 (Fig. 4I). These results suggest that SEPT11 increased HCC progression through activated RhoA. Owing to the mechanism of SEPT11 regulating cytoskeleton is through activation of RhoA, we explored the effect of SEPT11 on the downstream factors of the RhoA pathway. We found that SEPT11 affected the expression of ROCK1, but not ROCK2 and IQGAP (Fig. 5A). It is known that the phosphorylation of LIMK and cofilin, the downstream factors of ROCK1, regulates the dynamic changes of actin. We further explore the effect of SEPT11 on ROCK1 and its downstream signals. The results showed that the expression of p-LIMK and p-cofilin was significantly inhibited by KO-SEPT11, but the overexpression of RhoA rescued the inhibition of ROCK1 and its downstream signal caused by KO-SEPT11 (Fig. 5B). Next, we treated SEPT11-overexpressing cells with Y27632, a specific inhibitor of ROCK1, and found that Y27632 could significantly inhibit the LIMK/cofilin pathway activated by SEPT11 overexpression (Fig. 5C). ROCK1 is known to be a direct downstream factor of RhoA, so these results suggest that SEPT11 regulates the ROCK1/LIMK/cofilin pathway through RhoA. At the same time, Y27632 significantly inhibited the increase of actin fibers, fiber structure thickening and orderly arrangement induced by overexpression of SEPT11 without affecting the expression of SEPT11 and RhoA (Fig. 5D). Scratch experiments showed that overexpression of RhoA rescued the inhibition of cell migration induced by KO-SEPT11 (Fig. 5E). Migration experiments showed that Y27632 significantly inhibited cell migration promoted by overexpression of SEPT11 (Fig. 5F). Thus, SEPT11 promotes activation of the ROCK1/LIMK/cofilin pathway, regulating cytoskeleton and migration in HCC cells. A WB was used to detected the effect of SEPT11 expression on several key factors downstream of RhoA; B The effect of RhoA on SEPT11-activated LIMK/cofilin pathway was measured by WB; C The effect of ROCK1 inhibitor Y27632 on SEPT11-activated LIMK/cofilin pathway was detected by WB; D The effect of Y27632 on SEPT11-regulated actin arrangement and increase was tested by cellular immunofluorescence; E Scratch assay was used to detect the effect of RhoA on cell migration by inhibited by KO-SEPT11; F Transwell assay was used to detect the effect of Y27632 on cell migration regulated by SEPT11. RhoA not only regulates cytoskeleton reorganization, but is closely related to FAK signaling [36, 37]. FAK is associated with the dynamic assembly of focal adhesions (FAs) and the FAK/Src/paxillin signaling cascade is known to be involved in cell migration and adhesion [29]. This study has demonstrated that SEPT11 activates RhoA to function, and GSEA analysis also showed that SEPT11 regulates FAs (Supplementary Fig. 4A). Given these available results, we explored the effect of SEPT11 on FAs. We found that overexpression of SEPT11-activated FAK/Src/paxillin pathway, but knockdown of SEPT11 inhibited this signaling pathway (Fig. 6A). These results suggest that SEPT11 can mediate the activation of the FAK/Src/paxillin pathway. What is the upstream and downstream relationships between FAK signaling and RhoA/ROCK1 signaling in SEPT11-mediated HCC cell migration? We found that Rhosin could significantly inhibit the FAK/Src/paxillin pathway and the activation of this signaling pathway by SEPT11 (Fig. 6B). Cells were treated with FAK inhibitor PF-573228 or ROCK1 inhibitor Y27632, and found that PF-573228 inhibited FAK signaling but not affect RhoA/LIMK/cofilin signaling, while Y27632 inhibited both pathways (Fig. 6C). Meanwhile, PF-573228 also did not affect the number and orderly arrangement of actin fibers (Fig. 6D). So it is likely that RhoA-ROCK1 play a role upstream of FAK/Src/ paxillin signaling to enhance HCC cell adhesion and migration. Transwell migration and invasion experiments found that PF-573228 significantly inhibited cell migration and invasion promoted by SEPT11 overexpression (Fig. 6E; Supplementary Fig. 4F). The rescue experiments in vivo revealed that Rhosin nearly abolished SEPT11-promoted HCC metastasis (Fig. 6F). Thus, SEPT11 targets RhoA, thereby regulating cytoskeleton rearrangement and abnormal cell adhesion through ROCK1/cofilin and FAK/paxillin signaling pathways, promoting invasion and migration of HCC. A WB detection of the effect of SEPT11 expression on FAK/Src/paxillin signaling; B WB detection of the effect of RhoA on SEPT11-activated FAK/Src/paxillin signaling; C WB detection of Y27632 and FAK inhibitor PF573228 on FAK/Src/paxillin and LIMK/cofilin pathways; D The effect of PF573228 on actin arrangement and thickness detected by cellular immunofluorescence. E Detection of the effect of PF573228 on SEPT11-promoted cell migration; F Huh7-luciferase cells were injected into the tail vein of nude mice, and two groups of mice were treated with Rhosin (30 mg/kg) by intraperitoneal injection, then observed and photographed on the 28th day. It is well known that Rho GTPases are activated by guanine nucleotide exchange factors (GEFs), and multiple guanine nucleotide exchange factors have been shown to activate Rho GTPase, including GEF-H1, ARHGEF3, NET1 and so on [38]. GEF-H1 was of focus, because it was considered as RhoA-specific GEF and its catalytic activity toward RhoA is changed through microtubule dynamics influenced by Septins [39,40,41]. Therefore, we explored that whether SEPT11 affects the activity of RhoA through GEF-H1. Firstly, we explored whether SEPT11 affects the expression of GEF-H1. We found that SEPT11 did not significantly affect the transcriptional expression and protein content of GEF-H1(Fig. 7A, B). However, we found SEPT11 can promote the binding of GEF-H1 to RhoA (Fig. 7C). It is well known that GEF-H1 binds to RhoA can increase the GTP-RhoA form and activate RhoA [42]. Thus, SEPT11 may affects RhoA activity through GEF-H1. Co-IP was used to detect the combination between SEPT11 and GEF-H1, but we did not find the interaction between them (Fig. 7D). Phosphorylation of GEF-H1 Ser886 can promote the binding of GEF-H1 to RhoA, and activate RhoA [43]. We found that SEPT11 can promote the level of Phospho-GEF-H1 Ser886 (Fig. 7E). We further explored the effect of each structural domain of SEPT11 on Phospho-GEF-H1. Then we found that the phosphorylation effect of GEF-H1 Ser886 promoted by SEPT11 was abolished after removal of the GTP-binding domain (Fig. 7F; Supplementary Fig. 4D). GTP-binding domain of SEPT11 is also critical for activation of RhoA (Fig. 7G). Further, we knocked down GEF-H1 though siRNA, which inhibited the activation of RhoA by SEPT11 (Fig. 7H). Furthermore, in the ΔGTP-binding domain of SEPT11 fragment treatment groups, the GEF-H1 binding to RhoA less than the other SEPT11 fragment treatment groups (Supplementary Fig. 4E). Therefore, SEPT11 facilitates the binding of GEF-H1 to RhoA by promoting Phospho-GEF-H1 Ser886, which enhances the activity of RhoA, and the GTP-binding domain is crucial to play this role. A, B QPCR and WB were used to detect the transcriptional expression and protein content of GEF-H1; C CO-IP and WB were used to detect the effect of SEPT11 overexpression on GEF-H1 and RhoA binding; D CO-IP and WB were used to detect the binding of SEPT11 and GEF-H1 in HepG2 and Huh7 cells of SEPT11-overexpressed; E WB was used to detect the influence of SEPT11 on phosphorylation of GEF-H1 Ser886; F WB was used to detect the effect of each structural domain of SEPT11 on Phospho-GEF-H1; G The activation of RhoA promoted by SEPT11 was abolished after removal of the GTP-binding domain; H GEF-H1 can influence the activation of RhoA by SEPT11; I The diagram shows that SEPT11 promotes the activation of RhoA through GEF-H1, then RhoA regulates cytoskeletal organization and FA dynamics by activating LIMK/cofilin and FAK/Src pathways, thereby promoting the invasion and migration of HCC.
SEPT11 promotes activation of the ROCK1/LIMK/cofilin pathway, regulating cytoskeleton and migration in HCC cells

SEPT11 activates FAK/Src/paxillin signaling, promoting HCC cell adhesion, and migration

SEPT11 activates RhoA by promoting the binding of GEF-H1 to RhoA

Discussion
Central to the poor prognosis of HCC is regional spread and metastasis [1, 2]. Septins are an important part of the cytoskeleton, involved in cell invasion and migration, and regulate tumor development and progression [44, 45]. It has been reported that SEPT9 promotes the upregulation of MMPs expression near FAs and secretion of MMPs, thereby enhancing the migration and invasion of breast cancer cells [46]. Septin 7 plays a downstream effect in ERK3-induced migration of cancer cells, and its deletion abolishes the ability of ERK3 to promote lung cancer cell migration and invasion [47]. CDK2 interacts with SEPT2 to stabilize SEPT2 in HCC cells, and HCC with high expression of both CDK2 and SEPT2 is more prone to recurrence and may be more aggressive [48]. As a new member of the septin family, we reported the function of SEPT11 in HCC progression for the first time. In this study, we demonstrated the function of SEPT11 in promoting the metastasis of HCC, and preliminarily explored the related molecular mechanism of its promoting HCC metastasis.
Our previously results found that high expression of LncRNA AY is closely associated with poor prognosis and metastasis of patients with HCC [5]. In the process of exploring the function of AY in promoting HCC, we accidentally discovered a tumor-promoting gene SEPT11. We found that the expression of SEPT11 in HCC is regulated by AY. SEPT11 is an important target gene of AY to promote HCC progression. SEPT11 is widely expressed in various human tissues. We found that SEPT11 upregulation in HCC was significantly correlated with tumor differentiation, vascular invasion and predicted a poor prognosis. Furthermore, SEPT11 promoted invasion and migration in HCC cells in vitro and tumor metastasis in vivo, which is consistent with the functions of other septins in tumors. It has been reported that the elevated expression of SEPT9 and SEPT2 in glioma tissues and cell lines not only promote cell invasion but also enhance cell proliferation [49]. Overexpression of SEPT6 significantly promoted the proliferation, cell cycle transition, migration and invasion of HCC cells, and SEPT6 gene knockout has a significant inhibitory effect on the survival of HCC cell lines [50].
However, our study found that SEPT11 had no significant effect on tumor cell proliferation either in vivo or in vitro, and the high expression or knockout of SEPT11 also had little effect on cell proliferation-related factors. The result is different from other reported septins to a certain extent, which suggests that there are some differences in the function of diverse septins, but these results also confirm the ability of septins to regulate cell migration and invasion.
Given that migratory capability in SEPT11 has been found in vitro and in vivo, the mechanism of SEPT11 in HCC cells had important clinical values. As a cytoskeleton GTPase, the classical function of septins is to regulate the reorganization of cytoskeleton, thereby regulating cell motility [9, 10]. Actin (microfilaments), the main component of cytoskeleton, which abnormal rearrangement and polymerization were important to the migration of cancer cells. Our experiments demonstrate that SEPT11 can indeed regulate actin dynamics in HCC cells. Further GSEA analysis found that most of the signaling pathways regulated by SEPT11 were related to cell migration and invasion, and the core effectors in multiple pathways included Rho proteins. RNA-seq and ATAC-seq suggested that AY-regulated genes were enrichment in Rho GTPase activity and regulation of actin cytoskeletal. SEPT11 acts as an important downstream factor for the function of AY, which also implies that SEPT11 may be related to Rho GTPase signaling. Meanwhile, several studies have demonstrated that septins induce abnormal migration of cancer cells by regulating actin cytoskeleton signaling through Rho GTPase [29, 30, 32, 33]. Furthermore, our results are similar to those previously reports that SEPT11 indeed regulates the dynamic changes of actin by promoting the activation of RhoA, and play a role in promoting migration in HCC.
Increased migration ability was always related to high cytoskeletal tension, ordered arrangement of actin and abnormal cell adhesion [26]. RhoA and its downstream effectors are key regulators of adhesion and actin organization [35]. The study explored the effect of SEPT11 on downstream factors of RhoA. ROCK1 is the main target of RhoA. Through combining with LIM domains ROCK1 activates LIMK and its direct downstream effector cofilin, drives the turnover and spatial reorganization of F-actin [51]. Actin reorganization was reregulated by cofilin, as actin-depolymerizing factor of actin filaments, the phosphorylation level of cofilin regulates the cytoskeleton dynamics [26]. Prior work has shown that ROCK/LIMK/cofilin axis is important to promote migration of cancer [52]. In this study, we found that SEPT11 regulates the ROCK1/LIMK/cofilin pathway through RhoA, modulates actin cytoskeletal reorganization through phosphorylating and inactivating cofilin, and ultimately induces cell migration. The ROCK1-specific inhibitor Y27632 significantly suppressed the ROCK1/LIMK/cofilin pathway activated by SEPT11, inhibited polymerization of F-actin, decreased cell migration ability. These results were consistent with previous findings that LIMK and cofilin were downstream effectors of RhoA/ROCK pathway to regulate actin organization.
Cell adhesion and migration involve FA remodeling that is related to cytoskeletal organization and associated intracellular signals [53]. Cell adhesion is mediated by two core effectors FAK and Src within cells [54]. FAK is a protein tyrosine kinase that is activated by Rho protein, integrin and other signals, thus performing a variety of functions, such as attachment, migration, invasion and so on [55]. FAK is also a scaffolding protein that binds to Src, then can form FAK-Src complex and regulate downstream FA proteins such as paxillin. After phosphorylation of Paxlin Y118 regulated by FAK-Src complex, other adhesion molecules were recruited to mediate cell adhesion [56]. Our bioinformatics analysis showed that SEPT11 regulates the focal adhesion pathway. SEPT11 activates FAK/Src/paxillin pathway and thus promotes tumor migration, but both Rhosin and Y27632 can significantly inhibit the activation of this signal pathway by SEPT11. Meanwhile, Rhosin also nearly abolished SEPT11-promoted HCC metastasis. FAK signaling is closely related to Rho GTPase, RhoA is critical for modulating the actin-associated adhesion during cell migration [57]. Cell treatment with inhibitors of contractility significantly reduces cytoskeletal tension, then suppress FAK signaling [58]. FA dynamics and cytoskeletal organization make concerted efforts to drive cell migration [59]. Our work links SEPT11, F-actin and FAs to tumor cell motility, migration and metastasis for the first time.
SEPT11 did not form a firm complex with RhoA to influence its activity. Thus, “How SEPT11 affects RhoA activity”. GEFs specifically catalyze the exchange of GDP for GTP, which activates Rho GTPases. GEF-H1 is considered as RhoA-specific GEF, and can stimulate guanine nucleotide exchange of RhoA but is weak or inactive toward RAC, Cdc42, TC10 or Ras [60, 61]. The catalytic activity of GEF-H1 toward RhoA is changed through microtubule dynamics [39, 40], while septins (including SEPT11) associate with microtubule bundles and promote as well as guide microtubule growth in living cells, to influence their dynamics [62, 63]. Therefore, we hypothesized that SEPT11 affects the activity of RhoA through GEF-H1. We found that SEPT11 did not significantly affect the transcriptional expression and protein content of GEF-H1, but it could promote the phosphorylation of GEF-H1 Ser886 and the binding of GEF-H1 to RhoA. Further, the knocked down of GEF-H1 can inhibite the activation of RhoA by SEPT11. Phosphorylation of GEF-H1 Ser886 is important for GEF-H1-driven activation of RhoA [43]. Thus, SEPT11 affects RhoA activity through GEF-H1. We further found that GTP-binding domain of SEPT11 was critical for phosphorylation of GEF-H1 Ser886 and activation of RhoA. In sum, our study showed that SEPT11 affects RhoA activity through EGF-H1, and the GTP-binding domain is crucial to play this role.
In conclusion, SEPT11 is an important downstream factor of AY in promoting HCC. SEPT11 promotes the activation of RhoA through GEF-H1, then RhoA regulates cytoskeletal organization and FA dynamics by activating LIMK/cofilin and FAK/Src pathways, thereby promoting the invasion and migration of HCC (Fig. 7I). These data provide evidence for the involvement of SEPT11 in tumor progression and suggest that SEPT11 may serve as a novel prognostic marker and therapeutic target for HCC.
Data availability
The data needed to evaluate the conclusions in the paper are present in the paper and/or the supplementary materials, including full and uncropped western blots. Raw datasets generated and/or analyzed of this study are available from the corresponding author, upon reasonable request.
References
Forner A, Llovet JM, Bruix J. Hepatocellular carcinoma. Lancet. 2012;379:1245–55.
Llovet JM. Liver cancer: time to evolve trial design after everolimus failure. Nat Rev Clin Oncol. 2014;11:506–7.
McGlynn KA, London WT. The global epidemiology of hepatocellular carcinoma: present and future. Clin Liver Dis. 2011;15:223–43.
Li D, Fan X, Li Y, Yang J, Lin H. The paradoxical functions of long noncoding RNAs in hepatocellular carcinoma: Implications in therapeutic opportunities and precision medicine. Genes Dis. 2022;9:358–69.
Kang CL, Qi B, Cai QQ, Fu LS, Yang Y, Tang C, et al. LncRNA AY promotes hepatocellular carcinoma metastasis by stimulating ITGAV transcription. Theranostics. 2019;9:4421–36.
Stuelten CH, Parent CA, Montell DJ. Cell motility in cancer invasion and metastasis: insights from simple model organisms. Nat Rev Cancer. 2018;18:296–312.
Marx V. Tracking metastasis and tricking cancer. Nature. 2013;494:133–6.
van der Bijl I, Nawaz K, Kazlauskaite U, van Stalborch AM, Tol S, Jimenez Orgaz A, et al. Reciprocal integrin/integrin antagonism through kindlin-2 and Rho GTPases regulates cell cohesion and collective migration. Matrix Biol. 2020;93:60–78.
Mayor R, Etienne-Manneville S. The front and rear of collective cell migration. Nat Rev Mol Cell Biol. 2016;17:97–109.
Etienne-Manneville S, Hall A. Rho GTPases in cell biology. Nature. 2002;420:629–35.
Tang Y, He Y, Zhang P, Wang J, Fan C, Yang L, et al. LncRNAs regulate the cytoskeleton and related Rho/ROCK signaling in cancer metastasis. Mol Cancer. 2018;17:77.
Mostowy S, Cossart P. Septins: the fourth component of the cytoskeleton. Nat Rev Mol Cell Biol. 2012;13:183–94.
Bartsch I, Bläser S, Röseler S, Sandrock K, Busse A, Huber M, et al. Human endothelial and platelet septin SEPT11: cloning of novel variants and characterisation of interaction partners. Thromb Haemost. 2010;104:1201–10.
Li M, Li X, Zhang Y, Wu H, Zhou H, Ding X, et al. Micropeptide MIAC inhibits HNSCC progression by interacting with aquaporin 2. J Am Chem Soc. 2020;142:6708–16.
Jeon TW, Yang H, Lee CG, Oh ST, Seo D, Baik IH, et al. Electro-hyperthermia up-regulates tumour suppressor Septin 4 to induce apoptotic cell death in hepatocellular carcinoma. Int J Hyperth. 2016;32:648–56.
Yue X, Cao D, Lan F, Pan Q, **a T, Yu H. MiR-301a is activated by the Wnt/β-catenin pathway and promotes glioma cell invasion by suppressing SEPT7. Neuro Oncol. 2016;18:1288–96.
Hanai N, Nagata K, Kawajiri A, Shiromizu T, Saitoh N, Hasegawa Y, et al. Biochemical and cell biological characterization of a mammalian septin, Sept11. FEBS Lett. 2004;568:83–88.
Son HY, Hwangbo Y, Yoo SK, Im SW, Yang SD, Kwak SJ, et al. Genome-wide association and expression quantitative trait loci studies identify multiple susceptibility loci for thyroid cancer. Nat Commun. 2017;8:15966.
Stevens SJ, Meers LE, Albrechts JC, Mebis-Verhees K, Bos GM, Engelen JJ, et al. A translocation in acute lymphoblastic leukemia that cytogenetically mimics the recurrent MLL-AFF1 translocation and fuses SEPT11 to MLL. Cancer Genet Cytogenet. 2010;201:48–51.
Giampazolias E, Zunino B, Dhayade S, Bock F, Cloix C, Cao K, et al. Mitochondrial permeabilization engages NF-κB-dependent anti-tumour activity under caspase deficiency. Nat Cell Biol. 2017;19:1116–29.
Bramswig NC, Everett LJ, Schug J, Dorrell C, Liu C, Luo Y, et al. Epigenomic plasticity enables human pancreatic α to β cell reprogramming. J Clin Invest. 2013;123:1275–84.
Lewis JJ, Reed RD. Genome-wide regulatory adaptation shapes population-level genomic landscapes in Heliconius. Mol Biol Evol. 2019;36:159–73.
Weinstein JN, Collisson EA, Mills GB, Shaw KR, Ozenberger BA, Ellrott K, et al. The Cancer Genome Atlas Pan-Cancer analysis project. Nat Genet. 2013;45:1113–20.
Fu L, Wang X, Zhai J, Qi W, **g L, Ge Y, et al. Changes in apoptosis, proliferation and T lymphocyte subtype on thymic cells of SPF chickens infected with reticuloendotheliosis virus. Mol Immunol. 2019;111:87–94.
Huang W, Zhu PJ, Zhang S, Zhou H, Stoica L, Galiano M, et al. mTORC2 controls actin polymerization required for consolidation of long-term memory. Nat Neurosci. 2013;16:441–8.
Li X, Cheng Y, Wang Z, Zhou J, Jia Y, He X, et al. Calcium and TRPV4 promote metastasis by regulating cytoskeleton through the RhoA/ROCK1 pathway in endometrial cancer. Cell Death Dis. 2020;11:1009.
Chen MH, Fu LS, Zhang F, Yang Y, Wu XZ. LncAY controls BMI1 expression and activates BMI1/Wnt/β-catenin signaling axis in hepatocellular carcinoma. Life Sci. 2021;280:119748.
Parsons JT, Horwitz AR, Schwartz MA. Cell adhesion: integrating cytoskeletal dynamics and cellular tension. Nat Rev Mol Cell Biol. 2010;11:633–43.
Zeng Y, Cao Y, Liu L, Zhao J, Zhang T, **ao L, et al. SEPT9_i1 regulates human breast cancer cell motility through cytoskeletal and RhoA/FAK signaling pathway regulation. Cell Death Dis. 2019;10:720.
Ito H, Iwamoto I, Morishita R, Nozawa Y, Narumiya S, Asano T, et al. Possible role of Rho/Rhotekin signaling in mammalian septin organization. Oncogene. 2005;24:7064–72.
Hu Q, Nelson WJ, Spiliotis ET. Forchlorfenuron alters mammalian septin assembly, organization, and dynamics. J Biol Chem. 2008;283:29563–71.
Hickson GR, O’Farrell PH. Rho-dependent control of anillin behavior during cytokinesis. J Cell Biol. 2008;180:285–94.
Piatti S. Cytokinesis: An Anillin-RhoGEF Module Sets the Stage for Septin Double Ring Assembly. Curr Biol. 2020;30:R347–49.
Huang D, Cao L, **ao L, Song JX, Zhang YJ, Zheng P, et al. Hypoxia induces actin cytoskeleton remodeling by regulating the binding of CAPZA1 to F-actin via PIP2 to drive EMT in hepatocellular carcinoma. Cancer Lett. 2019;448:117–27.
Tuntithavornwat S, Shea DJ, Wong BS, Guardia T, Lee SJ, Yankaskas CL, et al. Giant obscurin regulates migration and metastasis via RhoA-dependent cytoskeletal remodeling in pancreatic cancer. Cancer Lett. 2022;526:155–67.
Wu GS, Song YL, Yin ZQ, Guo JJ, Wang SP, Zhao WW, et al. Ganoderiol A-enriched extract suppresses migration and adhesion of MDA-MB-231 cells by inhibiting FAK-SRC-paxillin cascade pathway. PLoS ONE. 2013;8:e76620.
Chatterjee T, Zhang S, Posey TA, Jacob J, Wu L, Yu W, et al. Anti-GPR56 monoclonal antibody potentiates GPR56-mediated Src-Fak signaling to modulate cell adhesion. J Biol Chem. 2021;296:100261.
Lawson CD, Ridley AJ. Rho GTPase signaling complexes in cell migration and invasion. J Cell Biol. 2018;217:447–57.
Krendel M, Zenke FT, Bokoch GM. Nucleotide exchange factor GEF-H1 mediates cross-talk between microtubules and the actin cytoskeleton. Nat Cell Biol. 2002;4:294–301.
Seetharaman S, Vianay B, Roca V, Farrugia AJ, De Pascalis C, Boëda B, et al. Microtubules tune mechanosensitive cell responses. Nat Mater. 2022;21:366–77.
Bowen JR, Hwang D, Bai X, Roy D, Spiliotis ET. Septin GTPases spatially guide microtubule organization and plus end dynamics in polarizing epithelia. J Cell Biol. 2011;194:187–97.
Pan M, Chew TW, Wong DCP, **ao J, Ong HT, Chin JFL, et al. BNIP-2 retards breast cancer cell migration by coupling microtubule-mediated GEF-H1 and RhoA activation. Sci Adv. 2020;6:eaaz1534.
Voloshin T, Schneiderman RS, Volodin A, Shamir RR, Kaynan N, Zeevi E, et al. Tumor treating fields (TTFields) hinder cancer cell motility through regulation of microtubule and acting dynamics. Cancers (Basel). 2020;12:3016.
Poüs C, Klipfel L, Baillet A. Cancer-related functions and subcellular localizations of septins. Front Cell Dev Biol. 2016;4:126.
Liu M, Shen S, Chen F, Yu W, Yu L. Linking the septin expression with carcinogenesis. Mol Biol Rep. 2010;37:3601–8.
Marcus J, Bejerano-Sagie M, Patterson N, Bagchi S, Verkhusha VV, Connolly D, et al. Septin 9 isoforms promote tumorigenesis in mammary epithelial cells by increasing migration and ECM degradation through metalloproteinase secretion at focal adhesions. Oncogene. 2019;38:5839–59.
Elkhadragy L, Alsaran H, Long W. The C-terminus tail regulates ERK3 kinase activity and its ability in promoting cancer cell migration and invasion. Int J Mol Sci. 2020;21:4044.
Xu C, Zhang W, Zhang X, Zhou D, Qu L, Liu J, et al. Coupling function of cyclin-dependent kinase 2 and Septin2 in the promotion of hepatocellular carcinoma. Cancer Sci. 2019;110:540–9.
Xu D, Liu A, Wang X, Chen Y, Shen Y, Tan Z, et al. Repression of Septin9 and Septin2 suppresses tumor growth of human glioblastoma cells. Cell Death Dis. 2018;9:514.
Fan Y, Du Z, Ding Q, Zhang J, Op Den Winkel M, Gerbes AL, et al. SEPT6 drives hepatocellular carcinoma cell proliferation, migration and invasion via the Hippo/YAP signaling pathway. Int J Oncol. 2021;58:25.
McCall PM, MacKintosh FC, Kovar DR, Gardel ML. Cofilin drives rapid turnover and fluidization of entangled F-actin. Proc Natl Acad Sci USA. 2019;116:12629–37.
Lee HJ, Diaz MF, Price KM, Ozuna JA, Zhang S, Sevick-Muraca EM, et al. Fluid shear stress activates YAP1 to promote cancer cell motility. Nat Commun. 2017;8:14122.
Rondas D, Tomas A, Halban PA. Focal adhesion remodeling is crucial for glucose-stimulated insulin secretion and involves activation of focal adhesion kinase and paxillin. Diabetes. 2011;60:1146–57.
Li R, Wang Y, Zhang X, Feng M, Ma J, Li J, et al. Exosome-mediated secretion of LOXL4 promotes hepatocellular carcinoma cell invasion and metastasis. Mol Cancer. 2019;18:18.
van Nimwegen MJ, van de Water B. Focal adhesion kinase: a potential target in cancer therapy. Biochem Pharm. 2007;73:597–609.
Pasapera AM, Schneider IC, Rericha E, Schlaepfer DD, Waterman CM. Myosin II activity regulates vinculin recruitment to focal adhesions through FAK-mediated paxillin phosphorylation. J Cell Biol. 2010;188:877–90.
Zhang W, Huang Y, Gunst SJ. The small GTPase RhoA regulates the contraction of smooth muscle tissues by catalyzing the assembly of cytoskeletal signaling complexes at membrane adhesion sites. J Biol Chem. 2012;287:33996–4008.
Zhou DW, Fernández-Yagüe MA, Holland EN, García AF, Castro NS, O’Neill EB, et al. Force-FAK signaling coupling at individual focal adhesions coordinates mechanosensing and microtissue repair. Nat Commun. 2021;12:2359.
Dedden D, Schumacher S, Kelley CF, Zacharias M, Biertümpfel C, Fässler R, et al. The architecture of Talin1 reveals an autoinhibition mechanism. Cell. 2019;179:120–131.e113.
Ren Y, Li R, Zheng Y, Busch H. Cloning and characterization of GEF-H1, a microtubule-associated guanine nucleotide exchange factor for Rac and Rho GTPases. J Biol Chem. 1998;273:34954–60.
Gebregiworgis T, Marshall CB, Nishikawa T, Radulovich N, Sandí MJ, Fang Z, et al. Multiplexed real-time NMR GTPase assay for simultaneous monitoring of multiple guanine nucleotide exchange factor activities from human cancer cells and organoids. J Am Chem Soc. 2018;140:4473–6.
Woods BL, Gladfelter AS. The state of the septin cytoskeleton from assembly to function. Curr Opin Cell Biol. 2021;68:105–12.
Targa B, Klipfel L, Cantaloube I, Salameh J, Benoit B, Poüs C, et al. Septin filament coalignment with microtubules depends on SEPT9_i1 and tubulin polyglutamylation, and is an early feature of acquired cell resistance to paclitaxel. Cell Death Dis. 2019;10:54.
Acknowledgements
This line of research was supported by NHC key laboratory of glycoconjugates research (Fudan University), Tumor Hospital affiliated to Fudan University, cardiology of Huadong Hospital affiliated to Fudan University. Thanks to Shushu Song postdoctoral of Liver Cancer Institute of Zhongshan Hospital affiliated to Fudan University for donating Huh7-luciferase cells. This study was supported by the National Natural Science Foundation of China (31570800, 82003101).
Author information
Authors and Affiliations
Contributions
LSF: acquisition, analysis and interpretation of data, manuscript preparation; XYW: acquisition and analysis data; QQC: interpretation of data and supervised the project; YY, MHC, AK, YWD, HJL, and YXX: technical support and data acquisition; MZM and XZW: study design, analysis of data and manuscript editing. The approval of the final manuscript was received by all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Edited by Professor Stephen Tait
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fu, L., Wang, X., Yang, Y. et al. Septin11 promotes hepatocellular carcinoma cell motility by activating RhoA to regulate cytoskeleton and cell adhesion. Cell Death Dis 14, 280 (2023). https://doi.org/10.1038/s41419-023-05726-y
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41419-023-05726-y
- Springer Nature Limited