
Abstract
Background
Recent studies have emphasised the important role of amino acids in cancer metabolism. Cold physical plasma is an evolving technology employed to target tumour cells by introducing reactive oxygen species (ROS). However, limited understanding is available on the role of metabolic reprogramming in tumour cells fostering or reducing plasma-induced cancer cell death.
Methods
The utilisation and impact of major metabolic substrates of fatty acid, amino acid and TCA pathways were investigated in several tumour cell lines following plasma exposure by qPCR, immunoblotting and cell death analysis.
Results
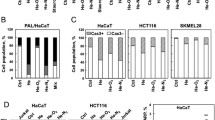
Metabolic substrates were utilised in Panc-1 and HeLa but not in OVCAR3 and SK-MEL-28 cells following plasma treatment. Among the key genes governing these pathways, ASCT2 and SLC3A2 were consistently upregulated in Panc-1, Miapaca2GR, HeLa and MeWo cells. siRNA-mediated knockdown of ASCT2, glutamine depletion and pharmacological inhibition with V9302 sensitised HeLa cells to the plasma-induced cell death. Exogenous supplementation of glutamine, valine or tyrosine led to improved metabolism and viability of tumour cells following plasma treatment.
Conclusion
These data suggest the amino acid influx driving metabolic reprogramming in tumour cells exposed to physical plasma, governing the extent of cell death. This pathway could be targeted in combination with existing anti-tumour agents.
Similar content being viewed by others

Background
Cancer is a devastating disease and the second cause of death in western societies. A particular trait of many types of cancers is their ability to evolve and become refractory to different treatment modalities, such as chemotherapy, radiotherapy and immunotherapy. To this end, novel treatment avenues are utterly needed to target tumours from multiple pathways simultaneously. Among many approaches being investigated in preclinical research, cold physical plasma has gained traction in translational research due to its inherent capacity to deposit many different ROS (reactive oxygen species) directly on the tumour tissue.1
Physical plasma is a partially ionised gas and multicomponent system. Plasmas expel ions, electrons, UV radiation, electric fields, and ROS to a different degree, depending on the type of plasma system and its settings being used.2 Plasma-derived ROS have been shown to limit tumour growth in several animal models, including, e.g. skin cancer,3,4,5 pancreatic cancer6,7,8 and colon cancer.9,5a). The integrity of the mitochondrial membrane potential was sustained. However, this effect was not seen in SK-MEL-28 cells (Fig. S4A), correlating with the findings above (Fig. 4a). The exogenous amino acid supplementation also rescued Panc-1 spheroids in 3D culture 72 h post-treatment (Fig. 5b). These results suggest that a subset of tumour cells activates the expression of genes involved in amino acid transport, thereby leading to a pro-survival phenotype following plasma exposure to attenuate ROS-mediated cytotoxicity.

Kinetic of metabolic activity and normalised area under the curve (AUC) of plasma-treated SK-MEL-28 (a), Panc-1 (b), and MeWO (c) cells with or without supplementation of exogenous glutamine, valine or tyrosine (200 μM). Data are mean ± SEM derived from three independent experiments. *p < 0.05; **p < 0.01; ***p < 0.001. Statistical analysis was done using one-way ANOVA with Tukey post-test for multiple comparisons. aa amino acid.

a representative images and quantification of viability (sytox green), cellular ROS (CellROX green) and mitochondrial membrane potential (TMRE) of plasma-treated Panc-1 cells supplemented with exogenous glutamine, valine or tyrosine (200 μM). Scale bar is 100 μm. b representative images and quantification of viability (sytox green) in Panc-1 derived spheroids supplemented with exogenous glutamine, valine or tyrosine (200 μM). Scale bar is 500 μm. Data are mean ± SEM derived from three independent experiments with *p < 0.05, **p < 0.01 and ***p < 0.001. Statistical analysis was done using one-way ANOVA with Tukey post-test for multiple comparisons.
Discussion
Cold physical plasma is an evolving technology employed to target tumour cells by delivering ROS directly to the tumour tissue.24 Since tumour cells are vulnerable to exogenous ROS, physical plasma treatment modalities provide a distinct advantage in the selective elimination of tumour cells.25 However, the ability of ROS to initiate a variety of signalling mechanisms26 and the observed tumour heterogeneity27 necessitates a more detailed understanding of how tumour cells respond to plasma.
Although glucose is the principal source of energy in cells and anti-glycolytic agents augment toxic plasma effects,28 amino acids are an important class of nutrients obligatory for cell survival. While some amino acids have specific biologic functions in metabolic processes, epigenetic regulation, and GSH synthesis,20 the primary physiologic function is to serve as building blocks for protein synthesis.29 Tumour cells employ opportunistic strategies to obtain these metabolic substrates during the exponential growth phase or to bypass therapeutic interventions. Since ROS induce gene expression and modulate protein activity, we employed a two-step screening approach to understand the metabolic signatures in tumour cell lines. The substrate utilisation in tumour cells assessed 1-h post plasma treatment could be attributed to the early post-translational modifications (e.g. phosphorylation, s-glutathionylation and nitration) of various enzymes and proteins of the TCA, glycolysis fatty acid and amino acid pathways.30 However, the mRNA expression analysis was carried out at 6 h post plasma exposure, to include specific gene expression patterns induced by redox-regulated transcription factors.19 Examining these datasets from several tumour cell lines revealed amino acid substrates, and its corresponding membrane transporters play a central role in metabolic reprogramming in tumour cells.
In our study, ASCT2 and SLC3A2 were consistently upregulated in tumour cell lines that exhibited prolonged survival following plasma treatment. The importance of these genes was further validated using a non-transformed HSMC cell line. ASCT2 belongs to the group of system ASC, transporting alanine, glutamine, serine and cysteine along with other aliphatic amino acids.31 SLC3A2 is an essential transmembrane protein critical for activating LAT1/2/3, ASC1, BAT1 and xCT involved in amino acid transport.32,33,34 Supplementation of exogenous ASCT2-specific amino acids (glutamine and valine) led to improved metabolic activity and survival in plasma-treated tumour cells in Panc-1 and MeWo but not in SK-MEL-28 cells. Glutamine is the primary amino acid that drives the TCA cycle and sustains ATP production.35 In the absence of glucose, glutamine-derived fumarate malate and citrate are increased in a glucose independent TCA cycle.36 Vice versa, the pharmacological or genetic inhibition of ASCT2 sensitised the tumour cells to plasma-induced cell death. These results validate that tumour cells are dependent on extracellular substrates for their biosynthetic machinery during stress by activation of respective transporters.
Interestingly, the LAT1-specific amino acid tyrosine also improved the metabolic activity in Panc-1 cells. The qPCR screen did not detect significant changes in LAT1 expression in Panc-1 cells, but there was a significant induction at the protein level following plasma exposure. This could be due to an observed alternate LAT1 isoform (~70 kDa) induced in these cells leading to tyrosine uptake. The increase in metabolic function due to amino acid supplementation also reduced oxidative stress and partially restored mitochondrial membrane potential and viability in Panc-1 cells. However, the comparable effect was not observed in SK-MEL-28 cells as it lacked the induction of transporter expression following plasma exposure. Our screen also identified the consumption of fatty acid and TCA substrates. It has been previously described that amino acids can supply carbon atoms for lipid biosynthesis and the TCA cycle via sustained acetyl-CoA pools.29,37 The maintenance of cellular amino acid pools is also regulated by autophagy, evoked during cellular stress, thereby providing substrates for energy production in tumour cells.38,39 It could be speculated from the current study that uptake of exogenous amino acids could be the first choice to maintain cellular amino acid pools, after which cell may commit to autophagy. The observed metabolic alterations in our study could also have a direct consequence in the tumour microenvironment (TME). Extracellular glutamine supports the differentiation of T cells into TH1 T cells.40 Targeting glutamine metabolism in tumour cells by CB-839/anti-PD-1/PD-L1 led to the depletion of glutamine in the tumour microenvironment (TME), enabling potent anti-tumour immune responses.41 However, further studies are warranted to differentiate amino acid pools derived from exogenous and de novo sources following plasma treatment and their influence in the TME. A previous study highlighted the importance of D-glutamine and D-glutamate metabolism in leukemic cells exposed to cold plasma.42 Investigating plasma-treated myeloma cells, the authors demonstrated in another study a strong enrichment of the β-alanine, propanoate, and linoleic acid metabolism, while the metabolic pathways for alanine, aspartate, glutamate, arginine, and proline were found to be enriched as well, albeit to a lesser extent.43 Our previous study determined that the cystine/glutamate antiporter xCT is induced in tumour cells following plasma exposure, leading to treatment resistance.18 In the current study, we implicate exogenous amino acids in the prolonged survival of some tumour cells. However, the involvement of other amino acid transporters (not included in this study) cannot be ruled out. Apart from this, another recent study suggested the involvement of purine metabolism and Pantothenate and CoA biosynthesis in both non-malignant and malignant cells following plasma exposure in vitro.44
Taken together, there is strong evidence that amino acids play a vital role in metabolic reprogramming in tumour cells. Due to multiple substrate specificities of the amino acid transporters,31 and rampant dosage compensation in the SLC gene family,45,46 single-transport inhibitors are unlikely to be effective against cancer therapy. Hence, combination with plasma and specific transport inhibitors could be useful in the targeting of tumour cells.
References
Privat-Maldonado, A., Schmidt, A., Lin, A., Weltmann, K. D., Wende, K., Bogaerts, A. et al. ROS from physical plasmas: redox chemistry for biomedical therapy. Oxid. Med. Cell Longev. 2019, 9062098 (2019).
Graves, D. B. Mechanisms of plasma medicine: coupling plasma physics, biochemistry, and biology. IEEE Trans. Radiat. Plasma Med. Sci. 1, 281–92 (2017).
Mizuno, K., Shirakawa, Y., Sakamoto, T., Ishizaki, H., Nishijima, Y. & Ono, R. Plasma-induced suppression of recurrent and reinoculated melanoma tumors in mice. IEEE TRPMS 2, 353–9 (2018).
Bekeschus, S., Clemen, R., Niessner, F., Sagwal, S. K., Freund, E. & Schmidt, A. Medical gas plasma jet technology targets murine melanoma in an immunogenic fashion. Adv. Sci. 7, 1903438 (2020).
Lin, A., Gorbanev, Y., De Backer, J., Van Loenhout, J., Van Boxem, W., Lemiere, F. et al. Non-thermal plasma as a unique delivery system of short-lived reactive oxygen and nitrogen species for immunogenic cell death in melanoma cells. Adv. Sci. 6, 1802062 (2019).
Brullé, L., Vandamme, M., Riès, D., Martel, E., Robert, E., Lerondel, S. et al. Effects of a Non Thermal Plasma Treatment Alone or in Combination with Gemcitabine in a MIA PaCa2-luc Orthotopic Pancreatic Carcinoma Model. PLoS ONE 7, e52653. https://doi.org/10.1371/journal.pone.0052653 (2012).
Liedtke, K. R., Bekeschus, S., Kaeding, A., Hackbarth, C., Kuehn, J. P., Heidecke, C. D. et al. Non-thermal plasma-treated solution demonstrates antitumor activity against pancreatic cancer cells in vitro and in vivo. Sci. Rep. 7, 8319 (2017).
Hattori, N., Yamada, S., Torii, K., Takeda, S., Nakamura, K., Tanaka, H. et al. Effectiveness of plasma treatment on pancreatic cancer cells. Int. J. Oncol. 47, 1655–62 (2015).
Freund, E., Liedtke, K. R., van der Linde, J., Metelmann, H. R., Heidecke, C. D., Partecke, L. I. et al. Physical plasma-treated saline promotes an immunogenic phenotype in CT26 colon cancer cells in vitro and in vivo. Sci. Rep. 9, 634 (2019).
Lin, A. G., **ang, B., Merlino, D. J., Baybutt, T. R., Sahu, J., Fridman, A. et al. Non-thermal plasma induces immunogenic cell death in vivo in murine CT26 colorectal tumors. Oncoimmunology 7, e1484978 (2018).
Sagwal, S. K., Pasqual-Melo, G., Bodnar, Y., Gandhirajan, R. K. & Bekeschus, S. Combination of chemotherapy and physical plasma elicits melanoma cell death via upregulation of SLC22A16. Cell Death Dis. 9, 1179 (2018).
Wang, K., Jiang, J., Lei, Y., Zhou, S., Wei, Y. & Huang, C. Targeting metabolic-redox circuits for cancer therapy. Trends Biochem. Sci. 44, 401–14 (2019).
Panieri, E. & Santoro, M. M. ROS homeostasis and metabolism: a dangerous liason in cancer cells. Cell Death Dis. 7, e2253 (2016).
Diebold, L. & Chandel, N. S. Mitochondrial ROS regulation of proliferating cells. Free Radic. Biol. Med. 100, 86–93 (2016).
Sabharwal, S. S. & Schumacker, P. T. Mitochondrial ROS in cancer: initiators, amplifiers or an Achilles’ heel? Nat. Rev. Cancer 14, 709–21 (2014).
Bansal, A. & Simon, M. C. Glutathione metabolism in cancer progression and treatment resistance. J. Cell Biol. 217, 2291–8 (2018).
Hill, B. G. & Bhatnagar, A. Protein S-glutathiolation: redox-sensitive regulation of protein function. J. Mol. Cell Cardiol. 52, 559–67 (2012).
Bekeschus, S., Eisenmann, S., Sagwal, S. K., Bodnar, Y., Moritz, J., Poschkamp, B. et al. xCT (SLC7A11) expression confers intrinsic resistance to physical plasma treatment in tumor cells. Redox Biol. 30, 101423 (2020).
Ye, P., Mimura, J., Okada, T., Sato, H., Liu, T., Maruyama, A. et al. Nrf2- and ATF4-dependent upregulation of xCT modulates the sensitivity of T24 bladder carcinoma cells to proteasome inhibition. Mol. Cell Biol. 34, 3421–34 (2014).
Lieu, E. L., Nguyen, T., Rhyne, S. & Kim, J. Amino acids in cancer. Exp. Mol. Med. 52, 15–30 (2020).
Bekeschus, S., Schmidt, A., Niessner, F., Gerling, T., Weltmann, K. D. & Wende, K. Basic research in plasma medicine—a throughput approach from liquids to cells. J. Vis. Exp. 129, e56331 (2017).
Reuter, S., von Woedtke, T. & Weltmann, K. D. The kINPen-a review on physics and chemistry of the atmospheric pressure plasma jet and its applications. J. Phys. D Appl. Phys. 51, 233001–52 (2018).
Gandhirajan, R. K., Rodder, K., Bodnar, Y., Pasqual-Melo, G., Emmert, S., Griguer, C. E. et al. Cytochrome C oxidase inhibition and cold plasma-derived oxidants synergize in melanoma cell death induction. Sci. Rep. 8, 12734 (2018).
Hirst, A. M., Frame, F. M., Arya, M., Maitland, N. J. & O’Connell, D. Low temperature plasmas as emerging cancer therapeutics: the state of play and thoughts for the future. Tumour Biol. 37, 7021–31 (2016).
Liedtke, K. R., Freund, E., Hermes, M., Oswald, S., Heidecke, C. D., Partecke, L. I. et al. Gas plasma-conditioned Ringer’s lactate enhances the cytotoxic activity of cisplatin and gemcitabine in pancreatic cancer in vitro and in ovo. Cancers 12, 123 (2020).
Moloney, J. N. & Cotter, T. G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 80, 50–64 (2018).
Grzywa, T. M., Paskal, W. & Wlodarski, P. K. Intratumor and intertumor heterogeneity in melanoma. Transl. Oncol. 10, 956–75 (2017).
Kaushik, N., Lee, S. J., Choi, T. G., Baik, K. Y., Uhm, H. S., Kim, C. H. et al. Non-thermal plasma with 2-deoxy-D-glucose synergistically induces cell death by targeting glycolysis in blood cancer cells. Sci. Rep. 5, 8726 (2015).
Yoneshiro, T., Wang, Q., Tajima, K., Matsushita, M., Maki, H., Igarashi, K. et al. BCAA catabolism in brown fat controls energy homeostasis through SLC25A44. Nature 572, 614 (2019).
Wang, K., Jiang, J. W., Lei, Y. L., Zhou, S. T., Wei, Y. Q. & Huang, C. H. Targeting metabolic-redox circuits for cancer therapy. Trends Biochem. Sci. 44, 401–14 (2019).
Fuchs, B. C. & Bode, B. P. Amino acid transporters ASCT2 and LAT1 in cancer: partners in crime? Semin. Cancer Biol. 15, 254–66 (2005).
Sato, H., Nomura, S., Maebara, K., Sato, K., Tamba, M. & Bannai, S. Transcriptional control of cystine/glutamate transporter gene by amino acid deprivation. Biochem. Biophys. Res. Commun. 325, 109–16 (2004).
Napolitano, L., Scalise, M., Galluccio, M., Pochini, L., Albanese, L. M. & Indiveri, C. LAT1 is the transport competent unit of the LAT1/CD98 heterodimeric amino acid transporter. Int. J. Biochem. Cell Biol. 67, 25–33 (2015).
Cano-Crespo, S., Chillaron, J., Junza, A., Fernandez-Miranda, G., Garcia, J., Polte, C. et al. CD98hc (SLC3A2) sustains amino acid and nucleotide availability for cell cycle progression. Sci. Rep. 9, 14065–84 (2019).
Hensley, C. T., Wasti, A. T. & DeBerardinis, R. J. Glutamine and cancer: cell biology, physiology, and clinical opportunities. J. Clin. Invest. 123, 3678–84 (2013).
Le, A., Lane, A. N., Hamaker, M., Bose, S., Gouw, A., Barbi, J. et al. Glucose-independent glutamine metabolism via TCA cycling for proliferation and survival in B cells. Cell Metab. 15, 110–21 (2012).
Metallo, C. M., Gameiro, P. A., Bell, E. L., Mattaini, K. R., Yang, J., Hiller, K. et al. Reductive glutamine metabolism by IDH1 mediates lipogenesis under hypoxia. Nature 481, 380–4 (2011).
Guo, J. Y., Teng, X., Laddha, S. V., Ma, S. R., Van Nostrand, S. C., Yang, Y. et al. Autophagy provides metabolic substrates to maintain energy charge and nucleotide pools in Ras-driven lung cancer cells. Genes Dev. 30, 1704–17 (2016).
Kuma, A. & Mizushima, N. Physiological role of autophagy as an intracellular recycling system: With an emphasis on nutrient metabolism. Semin. Cell Dev. Biol. 21, 683–90 (2010).
Klysz, D., Tai, X. G., Robert, P. A., Craveiro, M., Cretenet, G., Oburoglu, L. et al. Glutamine-dependent alpha-ketoglutarate production regulates the balance between T helper 1 cell and regulatory T cell generation. Sci. Signal. 8, ra97 (2015).
Gross, M., Chen, J., Engler, J., Janes, J., Leone, R., MacKinnon, A. et al. Glutaminase inhibition with CB-839 enhances anti-tumor activity of PD-1 and PD-L1 antibodies by overcoming a metabolic checkpoint blocking T cell activation. Cancer Res. 76, (2016).
Xu, D., Ning, N., Xu, Y., Wang, B., Cui, Q., Liu, Z. et al. Effect of cold atmospheric plasma treatment on the metabolites of human leukemia cells. Cancer Cell Int. 19, 135 (2019).
Xu, D., Xu, Y., Ning, N., Cui, Q., Liu, Z., Wang, X. et al. Alteration of metabolite profiling by cold atmospheric plasma treatment in human myeloma cells. Cancer Cell Int. 18, 42 (2018).
Tian, M., Xu, D., Li, B., Wang, S., Qi, M., Zhang, H. et al. Metabolome analysis of selective inactivation of human melanoma and normal cells by cold atmospheric plasma. Plasma Chem. Plasma Process. 591–605 (2021).
Cutler, M. J. & Choo, E. F. Overview of SLC22A and SLCO families of drug uptake transporters in the context of cancer treatments. Curr. Drug Metab. 12, 793–807 (2011).
Broer, A., Gauthier-Coles, G., Rahimi, F., van Geldermalsen, M., Dorsch, D., Wegener, A. et al. Ablation of the ASCT2 (SLC1 A5) gene encoding a neutral amino acid transporter reveals transporter plasticity and redundancy in cancer cells. J. Biol. Chem. 294, 4012–26 (2019).
Acknowledgements
We acknowledge the technical support of Clara Ahrend and Felix Niessner.
Author information
Authors and Affiliations
Contributions
K.D.W., T.v.W., R.K.G. and S.B. designed the study; D.M. and S.K.S. performed the experiments; K.D.W., T.v.W., R.K.G. and S.B. supervised the study; K.D.W., T.v.W., R.K.G. and S.B. interpreted the results; K.D.W., T.v.W. and S.B. provided the funding; R.K.G., D.M. and S.B. designed the figures; R.K.G. and S.B. wrote the draft; K.D.W., T.v.W. and S.B. provided the funding; all authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate and consent for publication
Human tumour cell lines: Panc-1 (CRL-1469; ATCC, Manassas, Virginia, USA), HeLa (ATCC CRM-CCL-2), MeWo (ATCC HTB-65), MaMel86a (CVCL A221), Miapaca2GR (ATCC CRM-CRL-1420), OVCAR3 (ATCC HTB-161) and SK-MEL-28 (ATCC HTB-72). Non-tumorigenic human mesenchymal stem cell line: (HMSC; PromoCell, Heidelberg, Germany).
Data availability
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Competing interests
The authors declare no competing interests.
Funding information
Funding was received by the German Federal Ministry of Education and Research (BMBF, grant numbers 03Z22DN11 and 03Z22Di1), the German Research Foundation (DFG, grant number AOBJ 669606), as well as the European Social Fund and the Ministry of Education, Science and Culture of Mecklenburg-Vorpommern, Germany (grant number ESF/14-BM-A55-0006-18). D.M. was supported by the Gerhard-Domagk-Foundation in Greifswald, Germany. Open Access funding enabled and organized by Projekt DEAL.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gandhirajan, R.K., Meyer, D., Sagwal, S.K. et al. The amino acid metabolism is essential for evading physical plasma-induced tumour cell death. Br J Cancer 124, 1854–1863 (2021). https://doi.org/10.1038/s41416-021-01335-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41416-021-01335-8
- Springer Nature Limited
This article is cited by
-
Effective combination of cold physical plasma and chemotherapy against Ewing sarcoma cells in vitro
Scientific Reports (2024)
-
Prognostic value of amino acid metabolism-related gene expression in invasive breast carcinoma
Journal of Cancer Research and Clinical Oncology (2023)