
Abstract
Impaired motivational drive is a key feature of depression. Chronic stress is a known antecedent to the development of depression in humans and depressive-like states in animals. Whilst there is a clear relationship between stress and motivational drive, the mechanisms underpinning this association remain unclear. One hypothesis is that the endocrine system, via corticotropin-releasing hormone (CRH) in the paraventricular nucleus of the hypothalamus (PVN; PVNCRH), initiates a hormonal cascade resulting in glucocorticoid release, and that excessive glucocorticoids change brain circuit function to produce depression-related symptoms. Another mostly unexplored hypothesis is that the direct activity of PVNCRH neurons and their input to other stress- and reward-related brain regions drives these behaviors. To further understand the direct involvement of PVNCRH neurons in motivation, we used optogenetic stimulation to activate these neurons 1 h/day for 5 consecutive days and showed increased acute stress-related behaviors and long-lasting deficits in the motivational drive for sucrose. This was associated with increased Fos-protein expression in the lateral hypothalamus (LH). Direct stimulation of the PVNCRH inputs in the LH produced a similar pattern of effects on sucrose motivation. Together, these data suggest that PVNCRH neuronal activity may be directly responsible for changes in motivational drive and that these behavioral changes may, in part, be driven by PVNCRH synaptic projections to the LH.
Similar content being viewed by others
Introduction
Mood disorders, including depression, represent a major health and economic burden [1]. Depression has a complex etiology and encompasses a variety of symptoms such as anhedonia, despair, and reduced motivation. Unfortunately, these symptoms often impede the pursuit of and participation in treatment for depression [2]. Chronic stress is a key contributor to the onset of depression in humans and can provoke depressive-like behavior in rodents [3,4,5]. Indeed, hyperactivation of the neuroendocrine arm of the stress response, the hypothalamic-pituitary-adrenal (HPA) axis, has been reported in approximately 50% of individuals with depression [6]. Further, positive clinical outcomes after treatment with antidepressants have been associated with the normalization of stress hormone levels [7, 19]. Behavioral video analysis was undertaken by two individuals blind to treatment condition. The output from this program generated a record of all behaviors displayed over the 10-min period in Microsoft Excel.
Immunohistochemistry and microscopy
Following the completion of the experiments, mice were euthanised with sodium pentobarbitone (0.2 ml intraperitoneal, I.P, Virbac, AUS) 2 h after the commencement of their final 1-h optogenetic stimulation session and perfused with 20 ml of 0.1 M Phosphate Buffer Solution (PBS) and 50 ml of 4% paraformaldehyde (PFA). Brains were extracted and postfixed in 4% PFA at 4 °C for 2 h, then transferred to 30% sucrose in 0.1 M PBS for 48 h at 4 °C. Brains were frozen on dry ice and stored at −80 °C until sectioning. Coronal sections 40 μm thick were cut on a cryostat (Leica Biosystems CM1900) in a 1 in 4 series and stored in a 0.1 M PBS solution containing 0.1% sodium azide at 4 °C. For the detection Fos-protein in Experiment 1, tissue sections were incubated in a solution of PBS containing 1% Triton X-100, 2% normal horse serum and primary antibody (1:8000, Phospho-c-Fos, rabbit monoclonal; Cell Signaling Technology, MA, USA, CAT# 5348) for 48 h at 4 °C. Sections were then washed 3 times for 10 min in PBS before incubation with the secondary antibody (1:1000, donkey anti-rabbit IgG, Jackson ImmunoResearch, PA, USA; Product code #711-005-152) for 2 h at room temperature. After washing to remove unbound secondary antibody, sections were incubated for 1 h using a Vectastain ABC kit (Vector Laboratories, CA, USA; SKU: PK-6100) followed by incubation with diaminobenzodine in 2% filtered nickel sulfate for 15 min. Visualization of Fos-protein was achieved by the addition of glucose oxidase. This tissue was then co-labeled for orexin-A. To co-label without cross-reactivity from rabbit polyclonal antibodies, an Avidin/Biotin Blocking kit was used (Vector Laboratories, CA, USA) before incubating tissue in orexin primary antibody (1:10000, rabbit monoclonal, Phoenix Pharmaceuticals, CA, USA; CAT # H-003-30) for 48 h at 4 °C. The procedures and reagents for visualization of cytoplasmic orexin immunoreactivity, ABC kit and visualization steps are the same as for Fos-protein with the exception that 2% filtered nickel sulfate was replaced with 0.1% acetate buffer. Following completion of immunohistochemical procedures, brain sections were mounted on chrome alum-treated slides, dehydrated in a series of ethanol and xylene solutions and coverslipped.
Photomicrographs of brain sections were made using Olympus CellSens Software (version 1.3) on an Olympus BX51 microscope at 10x objective. Bilateral cell counting was undertaken using iVision (Biovision) computer program for PVN slices (Bregma -0.6 mm to -0.8 mm). Image J was used to quantify Fos-positive and orexin-positive cells in LH sections (Bregma −1.22 to −1.7). Due to limited tissue availability, Fos-positive cells were also quantified in the globus pallidus externus (GPe; Bregma –0.7 to –0.82; YFP n = 12, ChR2 n = 13) and ventral tegmental area (Bregma –2.92 to –3.08; YFP n = 4, ChR2 n = 4) using ImageJ from a subset of animals. Coronal brain sections were chosen according to Paxinos and Franklin’s Mouse Brain in Stereotaxic Coordinates 6th edition [29] and quantification was undertaken by one observer, blind to treatment.
PVN and LH sections from a second series of brain tissue were mounted onto microscope slides and coverslipped using a 0.1 M PBS and glycerol solution. Probe placement and viral expression were then visualized using an Olympus BX51 microscope. Please refer to Fig. S1 for fiber optic probe placements.
Statistical analysis
Statistical analyses were conducted using JASP V16.2. Sucrose lever pressing was analyzed using an analysis of variance (ANOVA) with the between-subjects factor of Virus (YFP, ChR2) and the within-subjects factor of Day for initial FR1 and FR3 operant training, and two within subject factors, day and stimulation period (pre vs post) for FR3 and PR operant testing, as well as 5 min time bins for PR responding on Day 3. A two-tailed t test was used to assess the effect of optogenetic PVNCRH neuron on single-measure data including recruitment on Fos-protein activity and day 3 PR. Pearson’s correlations examined the relationship between Fos activity in the LH and PVN of ChR2 animals. Stress-related behaviors were analyzed using ANOVA with the between-subjects factor of Virus (YFP, ChR2) and the within-subjects factor of Day. Tukey post-hoc comparisons were used to differentiate any significant interactions. The assumption of normality for ANOVA was analyzed using the Shapiro-Wilk test and the assumption of homogeneity of variances was assessed with Levene’s test of equality of variances. In some instances, these assumptions were violated however ANOVA is typically robust against these violations [30]. Tests of sphericity were assessed for within-subjects ANOVA comparisons, if sphericity was violated, Greenhouse-Geisser corrections were reported. Graphs show mean ± standard error of the mean (SEM).
Results
Effect of optogenetic recruitment of PVNCRH neurons on motivation for sucrose
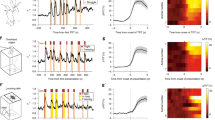
CRH-Cre transgenic mice were injected with a Cre-dependent excitatory opsin (ChR2) into the PVN (Fig. 1A, B). Additionally, a fiber optic probe was implanted immediately dorsal to the PVN to allow optogenetic activation of CRH neurons in the PVN (Fig. 1C). Mice were initially trained to lever press for sucrose by increasing fixed ratio schedules (Fig. 1C). Analysis of active lever presses during operant training revealed no significant interaction of Virus and Day during the FR1 schedule (F9,234 = 0.305, p = 0.973) or FR3 schedule of reinforcement (Fig. 1D; F7,182 = 0.731, p = 0.646). Following operant training, mice were assessed for their motivation for sucrose under both low effort (FR3) and high effort (PR) schedules of reinforcement at baseline and following 5 days of photostimulation of PVNCRH neurons (Fig. 1C; “Stim” sessions). Under FR3 conditions, there was no effect of repeated PVNCRH photostimulation on the number of active lever presses for sucrose (Fig. 1E; F1,26 = 0.315, p = 0.865). There was no effect of Virus on inactive lever presses during FR3 training days or PR sessions (Fig. S2). However, under higher effort PR conditions, there was a significant effect of repeated PVNCRH photostimulation on the number of active lever presses for sucrose (F2,52 = 14.06, p < 0.0001). There was also an effect of repeated PVNCRH stimulation on breakpoint during PR sessions (Fig. 1E; F2,52 = 14.20, p < 0.0001). Tukey post-hoc comparisons revealed significant differences in breakpoint between YFP control animals and ChR2 animals on Day 3 post-stimulation (Fig. 1F; p = 0.025). Closer inspection of 5 min time bin active lever press data on Day 3 post-stimulation revealed a significant interaction of Virus and Time bin on active lever presses (Fig. 1F; F17,442 = 5.105, p < 0.001). Interestingly, when the protocol was extended to 7 days of PR, in a second cohort of mice, the effect of repeated PVNCRH stimulation on breakpoint persisted, but began to return to control levels by Day 7 (Fig. 1G; F9,135 = 3.379, p < 0.001).
Effect of optogenetic recruitment of the PVNCRH neurons on Fos-protein activity
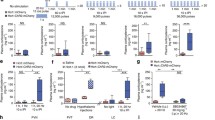
On the day after the final PR session, mice underwent a final photostimulation session and were perfused 2 h later. Immunohistochemical analysis revealed an increased number Fos-positive cells in the PVN (F1,26 = 6.011, p = 0.021) and LH (F1,26 = 5.778, p = 0.024) in ChR2 mice compared to YFP controls (Fig. 1H; 1I). There was a trend for an effect of Virus on the percentage of Fos-positive orexin cells in the LH (Fig. 1J; F1,26 = 2.942, p = 0.098). An analysis of Fos-positive neurons in extra-hypothalamic areas revealed an increased number of Fos-positive cells in the GPe of ChR2 mice compared to YFP mice (Fig. S3A; F1,23 = 6.065, p = 0.022). There was no significant difference in the number of Fos-positive cells in the VTA of ChR2 mice compared to controls (Fig. S3B; F1,6 = 0.214, p = 0.660).
Effect of optogenetic recruitment of the PVNCRH neurons on stress-related behaviors
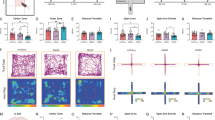
We determined the effect of PVNCRH photostimulation on stress-like behaviors across the five days during the first 10 min of 473 nm blue light exposure. Behavioral analysis during the photostimulation sessions demonstrated a significant effect of Virus and Behavior on the time spent engaging in each stress-related behavior (Fig. 3A–C; F6,189 = 18.260, p < 0.0001). Tukey post-hoc comparisons showed that ChR2 mice exhibited significantly increased digging and grooming behaviors than YFP controls across the five sessions (p’s < 0.05). Additionally, ChR2 mice exhibited reduced rearing and exploratory behaviors (walking) compared to YFP controls (p’s < 0.05).

A During the first PVNCRH photostimulation session, a shift in stress-related behaviors was observed in ChR2-mice compared to YFP controls including reduced rearing behavior, increased digging behavior and increased grooming. **p < 0.01, ***p < 0.001 post-hoc analyses. B During the third photostimulation session, ChR2 mice showed increased stress-related behaviors compared to YFP controls including reduced rearing behavior, increased digging behavior, increased grooming and reduced walking. *p < 0.05, ***p < 0.001 post-hoc analyses. C During the fifth PVNCRH photostimulation session, ChR2 mice showed increased stress-related behaviors compared to YFP controls including increased digging behavior, reduced walking, and increased grooming. ***p < 0.001 post-hoc analyses. PVN paraventricular nucleus of the hypothalamus.
Effect of optogenetic recruitment of the PVNCRH to LH pathway on motivation for sucrose
Previous work has shown that the LH is a target of PVNCRH terminals, and that activation of this pathway can elicit acute stress-related behaviors [19]. Further, in experiment 1 optogenetic photostimulation of PVNCRH cell bodies resulted in increased LH Fos. Thus, we next asked whether repeated stimulation of this circuit would suppress the motivation for sucrose in a similar manner to cell body stimulation. CRH-Cre transgenic mice were injected with DIO-ChR2 into the PVN, and fiber optic probes were implanted bilaterally immediately dorsal to the LH (Fig. 2A, B). Mice completed the same sucrose training and photostimulation sessions as conducted in Experiment 1 (Fig. 2B), but instead, targeting PVNCRH terminals in the LH. Analysis of active lever presses during operant training showed no significant effect of Virus and Day during the FR1 schedule (F9,90 = 0.713, p = 0.696) or FR3 schedule of reinforcement (Fig. 2C; F7,70 = 0.183, p = 0.988). Following operant training, mice were again assessed for their motivation for sucrose under both low effort (FR3) and high effort (PR) schedules of reinforcement at baseline and following 5 days of photostimulation of PVNCRH terminals in the LH (Fig. 2B; “Stim” sessions). Under low effort, FR3 conditions, there was no effect of repeated PVNCRH terminal photostimulation in the LH on the number of active lever presses for sucrose (Fig. 2D; F1,10 = 0.071, p = 0.795). There was also no significant effect of repeated PVNCRH terminal stimulation in the LH on the number of active lever presses for sucrose under higher effort PR conditions (F5,50 = 1.569, p = 0.186). Following PVNCRH terminal stimulation in the LH, there was a trend for altered breakpoint during PR sessions (Fig. 2D; F2,20 = 3.475, p = 0.051). Separate analysis of post-stimulation breakpoint revealed a significant Day × Virus interaction (F2,20 = 4.718, p = 0.021), suggesting a similar pattern of effects to cell body stimulation in Fig. 1F. The largest effect of PVNCRH photostimulation observed in Experiment 1 occurred on Day 3 of PR. Thus, we also decided to closely examine this time point following optogenetic recruitment of the PVNCRH to LH pathway. There was no significant effect of PVNCRH terminal stimulation in the LH on PR breakpoint when comparing pre-stimulation versus post-stimulation on Day 3 of PR testing (Fig. 2E; F1,10 = 2.781, p = 0.126). There was a trend towards a significant reduction in the number of active lever presses for sucrose in ChR2 mice compared to YFP controls on PR Day 3 post-stimulation (Fig. 2F; F1,10 = 3.517, p = 0.090). Time bin active lever press data on Day 3 post-stimulation revealed a significant effect of Virus and Time bin on active lever presses (Fig. 2G; F17,170 = 2.782, p < 0.001).
Effect of optogenetic recruitment of the PVNCRH to LH pathway on stress-related behaviors
Similar to PVNCRH stimulation experiments, optogenetic recruitment of the PVNCRH to LH pathway significantly increased stress-related behaviors compared to YFP controls. There was a significant effect of Virus and Behavior on the time spent engaging in each stress-related behavior (Fig. 4A–C; F6,70 = 8.24, p < 0.0001). Tukey post-hoc comparisons showed that ChR2 mice had significantly increased digging during the stimulation on day 1 compared to YFP controls (p < 0.05). Additionally, ChR2 mice exhibited increased grooming across the three sessions analyzed (p’s < 0.05). There was a significant correlation between PVN Fos activity and LH Fos activity in ChR2 mice, (Fig. S4; r = 0.817, p < 0.0002).

A During the first photostimulation session, ChR2 mice showed increased stress-related behaviors compared to YFP controls including increased digging behavior and increased grooming. *p < 0.05; **p < 0.01 post-hoc analyses. B During the third PVNCRH to LH pathway photostimulation session, ChR2 mice showed increased stress-related behaviors compared to YFP controls with increased grooming behavior, increased digging, reduced walking and reduced periods of inactivity. *p < 0.05; **p < 0.01 post-hoc analyses. C During the fifth photostimulation session, ChR2 mice showed increased grooming behavior and reduced walking. *p < 0.05; ***p < 0.001 post-hoc analyses. PVN paraventricular nucleus of the hypothalamus, LH lateral hypothalamus.
Discussion
Stress-induced activation of the HPA axis is strongly associated with mood disorders such as depression, however, a direct link between the activation of PVNCRH neurons that control the HPA axis and impairments in motivational drive has been lacking. Here we show that repeated photoactivation of PVNCRH neurons increased acute stress-related behaviors including grooming and digging, and is sufficient to reduce the motivation to lever press for sucrose, exclusively in a high-effort progressive ratio schedule of reinforcement. Interestingly, this effect persisted for up to 7 days following PVNCRH stimulation. Direct stimulation of PVNCRH neurons increased Fos-protein activity in the PVN and LH, with a trend towards an increase in LH orexin-expressing neurons. Given that recent literature has implicated the PVNCRH to LH pathway in stress-related behaviors and early life stress promotes LH-driven impairments in motivational drive [19, 20], we examined the direct effect of PVNCRH to LH stimulation on the effort to lever press for sucrose. Repeated activation of this pathway produced a similar pattern of changes in PR responding to PVNCRH neuronal cell body stimulation, although not as robustly as activating PVNCRH neuron cell bodies directly. Acutely, photoactivation of PVNCRH terminals in the LH also produced characteristic stress-associated behavioral responses, including increased digging and grooming. Together, these data suggest that PVNCRH neuron activity can precipitate depressive-like behaviors that may extend beyond increased HPA axis activation, as demonstrated by our finding that repeated photostimulation of PVNCRH neurons directly impacts motivational drive.
PVNCRH activation increased acute stress-related behaviors, reduced motivated responding for sucrose, and altered Fos-protein immunoreactivity
In the current study, we found that repeated activation of PVNCRH neurons selectively suppressed responding for sucrose under a high-effort progressive ratio (PR) schedule of reinforcement, but not under low-effort fixed ratio schedule (FR3). This effect of PVNCRH stimulation on motivational drive persisted for up to 7 days post-stimulation, although the effect weakened over time. This finding demonstrates that repeated activation of PVNCRH neurons has a lasting impact on the incentive motivation for sucrose, a phenotype typically observed in people with depression. Whilst we cannot discount the impact of repeated PVNCRH activation on other cognitive domains (particularly those associated with depressed states) [31], an effect of repeated stimulation on FR3 responding was not evident, suggesting relatively selective effects on high effort motivation. Additionally, prior to PVNCRH stimulation, both YFP controls and ChR2 mice showed no differences in FR1 or FR3 operant training, arguing against any potential unexpected behavioral effects of the virus. Interestingly, the impact of stimulation on sucrose responding only began to manifest from day 2 of PR testing. We have observed similar behavioral effects in our previous work assessing the impact of early life stress on the motivation for sucrose [20]. In this study, PR breakpoints were similar on day 1 of testing in control and early life stressed groups, however stressed rats showed a significant decrease in motivation from day 2 onwards. This delayed effect may reflect a loss of capacity to maintain sustained motivation over repeated sessions. This phenotype is consistent with other preclinical work demonstrating that social defeat stress reduces PR responding for saccharin and sucrose consumption up to 5 weeks following stress [32, 33].
In the current study, the motivational performance of the ChR2 group recovered after 7 days of PR testing, suggesting the possibility of compensatory re-engagement in the task. The effects also suggest that 5 days of PVNCRH photostimulation are not sufficiently intense to induce “permanent’ changes in circuit function, however a longer-term exposure may do so. Indeed, for many chronic stress paradigms, several weeks of stress are required to see long-lasting changes in behavior and physiology [34]. Analyzing protracted time points within the experiment and assessing individual susceptibility to repeated PVNCRH photostimulation may also be relevant for more direct comparisons to depression-related phenotypes. Regardless, our preclinical findings and previous research align with the human depression literature where chronic stressors have been associated with persistent depressive symptoms, frequent relapse, and poorer prognosis [35, 36]. Importantly, our findings suggest a direct role for PVNCRH neurons in these depression-related motivational impairments.
Consistent with recent work [19], we found that optogenetic activation of PVNCRH neurons produced a highly reproducible pattern of stress-related behaviors. We extend this work by demonstrating that these PVNCRH stimulation-evoked behaviors are relatively stable across five days of stimulation for certain behaviors such as grooming and digging. This pattern was characterized by increased grooming and digging, along with reduced walking and rearing behavior, although a loss of reductions in rearing was observed on Day 5. Grooming is a complex innate behavior and in the context of stress is thought to be important for de-arousal following stress exposure [37]. The increase in digging behavior we observed during activation of PVNCRH neurons has not been previously reported following a similar experimental design [19]. A potential explanation for digging in our experimental preparation is the inclusion of bedding in the test cage, which may have allowed for the expression of this behavior. Like grooming, digging is an innate rodent behavior that has potential for pathological repetition and is associated with stressful and anxiogenic situations [38]. Digging has conceptual similarity to the anxiety-associated marble burying test, which has been argued to be more appropriately considered an indicative measure of repetitive digging [39, 40]. However, it is important to note that behavioral effects following repeated photostimulation may also be reflective of dysregulated motor behavior. Changes in walking or rearing may be relevant to stress-induced exploratory behavior, or hypervigilance for threat assessment. Interestingly, the current behavioral data and that of Füzesi and colleagues [19] suggests that optogenetic stimulation of PVNCRH neurons does not fully recapitulate typical behavioral patterns following acute stress, such as increases in walking and rearing [19]. Instead, PVNCRH photostimulation may drive complex behavioral selection in which repetitive, internally focused stress behaviors predominate. Importantly, behaviors elicited by optogenetic stimulation occurred rapidly, within seconds, therefore these responses are likely driven by synaptic rather than neuroendocrine mechanisms [41]. This is supported by our similar acute behavioral observations following selective optogenetic stimulation of PVNCRH terminals in the LH. These findings indicate that PVNCRH neurons mediate fast, stress-relevant behaviors, likely independent of hormonal feedback [19].
Following repeated PVNCRH photostimulation, we observed a significant increase in the number of Fos-positive cells in the PVN and LH. These findings are consistent with work from Füzesi and colleagues [19], who showed that acute activation of PVN to LH terminals can produce a range of stress-relevant behaviors through direct, excitatory projections to the LH. However, the identity of these LH Fos-positive neurons remains to be determined. Prior evidence from our lab has shown a role of LH orexin in early life stress-induced motivational deficits [20], and in the current study repeated PVNCRH stimulation was associated with a trend towards a significant increase in the percentage of Fos-positive orexin cells. This is interesting as other work using monosynaptic retrograde rabies tracing has shown that CRH directly innervates LH orexin neurons [42]. While this effect did not reach statistical significance, there are known limitations with Fos-protein map** including sensitivity and temporal resolution. Additionally, orexin neurons are spontaneously active and sensitive to many different environmental and behavioral factors, including changes in light, social interaction and energy status [43]. Future studies using electrophysiological recordings or Ca2+ imaging might be required to detect temporally specific changes in orexin cell activity [19, 20]. Finally, we cannot rule out the possibility that other LH neuronal populations may be playing a role including melanin-concentrating hormone neurons and GABAergic neurons [44,45,46].
PVNCRH to LH pathway activation modestly reduced motivated responding for sucrose and increased stress-related behaviors
A similar, although weaker, effect on high-effort PR responding was found following repeated photostimulation of PVNCRH terminals in the LH. Photostimulation of PVNCRH terminals in the LH also produced a similar pattern of acute stress-related behaviors, including digging and grooming, as PVNCRH cell body stimulation, consistent with previous literature [19]. There is growing interest in the projections of PVNCRH neurons beyond the median eminence, with recent demonstrations of important functions outside HPA axis and autonomic activity. Indeed, PVNCRH neurons are known to project to a number of other downstream sites with well described roles in reward and stress-co** behaviors, such as the ventral tegmental area (VTA), locus coeruleus, periaqueductal gray and globus pallidus [47,48,49,50,51,52]. Thus, it may be that activation of additional terminal projections of the CRH neurons is required to produce the full PVNCRH neuronal cell body stimulation effect on motivation [53]. In support, acute stress results in CRH release in the VTA, where it has been shown to modulate reward-related behavior [54]. Thus, future research on motivational drive following activation of other reward-related projection pathways of PVNCRH neurons is required.
Methodological considerations
Our most interesting finding is that direct PVNCRH stimulation produces persistent reductions in motivational drive and an immediate onset of stress-related grooming and digging. This appears to be independent of HPA-axis activity and extends the previous work of Füzesi and colleagues [19]. One limitation of the current study is that we did not assess the involvement of circulating glucocorticoids on motivational drive. Whilst the current experiments do not have a direct read-out of neuroendocrine output, we know CRH neuron activation leads to glucocorticoid secretion, but given the reciprocal connections with the LH we cannot rule out the effects of LH terminal stimulation on circulating glucocorticoids [55, 56]. Importantly, the key outcome from this work is that activation of CRH neurons in the PVN is sufficient on its own to promote motivational disturbances, suggesting that future studies examining the links between stress and behavior need to consider both the synaptic and endocrine projections of these cells.
Another important consideration is the selectivity of AAV-ChR2 to CRH neurons within the PVN. The CRF-Cre mouse line has been validated by previous work, using an overlap of endogenous CRH-tagged td::tomato and PVN-specific Fos expression following an acute stressor [19, 25]. However, there are non-CRH expressing neurons located within the PVN, i.e. the magnocellular neurons that synthesize and release oxytocin and vasopressin [57]. The current methodologies cannot completely rule out non-selective and/or indirect ChR2 activation within these populations; however, this is a potential limitation of many optogenetic studies. Future experiments using electrophysiological patch-clamp recordings within PVN slices to study local connectivity will be important to identify possible contributions of other cell types involved. An additional consideration is the potential for neuronal damage due to the repeated nature of the optogenetic stimulation. The strength of the photostimulation in the current design (15 mW) is based on published optogenetic work [19], and the potential for tissue overheating is mitigated by the use of a pulse train and 30 s cycles of pulses followed by 30 s of no laser stimulation, as opposed to employing a continuous light signal [58]. No obvious tissue damage was observed during cFos processing of brain slices, however additional immunohistochemical markers sensitive to cell death should be considered for future experiments.
Conclusions
Here we show ChR2-mediated stimulation of PVNCRH neurons produced a long-lasting reduction in the motivation for a natural reward, distinct from a general behavioral deficit in sucrose operant responding as demonstrated by lack of effects on low effort behavior. These results are consistent with literature suggesting that repeated activation of stress responses mediated by the PVN can lead to depressive-like behaviors in rodents and humans [59, 60]. Overall, these data have important implications for the identification of novel hypothalamic circuits that govern stress-induced changes in mood and motivation. These current advances may aid in develo** more effective treatments for debilitating symptoms of neuropsychiatric diseases such as depression, through selective targeting of these PVN projection pathways.
Data availability
All data are available upon request to the corresponding authors
References
James SL, Abate D, Abate KH, Abay SM, Abbafati C, Abbasi N, et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet. 2018;392:1789–858.
Höflich A, Michenthaler P, Kasper S, Lanzenberger R. Circuit mechanisms of reward, anhedonia, and depression. Int J Neuropsychopharmacol. 2019;22:105–18.
Antoniuk S, Bijata M, Ponimaskin E, Wlodarczyk J. Chronic unpredictable mild stress for modeling depression in rodents: Meta-analysis of model reliability. Neurosci Biobehav Rev. 2019;99:101–16.
Kendler KS, Karkowski LM, Prescott CA. Causal relationship between stressful life events and the onset of major depression. Am J Psychiatry. 1999;156:837–41.
Willner P. Chronic mild stress (CMS) revisited: consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology. 2005;52:90–110.
Stetler C, Miller GE. Depression and hypothalamic-pituitary-adrenal activation: a quantitative summary of four decades of research. Psychosom Med. 2011;73:114–26.
Holsboer F. Stress, hypercortisolism and corticosteroid receptors in depression: implications for therapy. J Affect Disord. 2001;62:77–91.
Ito N, Nagai T, Yabe T, Nunome S, Hanawa T, Yamada H. Antidepressant-like activity of a Kampo (Japanese herbal) medicine, Koso-san (**ang-Su-San), and its mode of action via the hypothalamic-pituitary-adrenal axis. Phytomedicine. 2006;13:658–67.
Gourley SL, Kiraly DD, Howell JL, Olausson P, Taylor JR. Acute hippocampal brain-derived neurotrophic factor restores motivational and forced swim performance after corticosterone. Biol Psychiatry. 2008;64:884–90.
Gourley SL, Taylor JR. Recapitulation and reversal of a persistent depression-like syndrome in rodents. Curr Protoc Neurosci. 2009;Chapter 9:Unit 9.32.
Herman JP, Cullinan WE. Neurocircuitry of stress: central control of the hypothalamo–pituitary–adrenocortical axis. Trends Neurosci. 1997;20:78–84.
Sapolsky RM, Romero LM, Munck AU. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev. 2000;2:55–89.
Vale W, Rivier C, Brown MR, Spiess J, Koob G, Swanson L, et al. Chemical and biological characterization of corticotropin releasing factor. Recent Prog Horm Res. 1983;39:245–70.
Kvarta MD, Bradbrook KE, Dantrassy HM, Bailey AM, Thompson SM. Corticosterone mediates the synaptic and behavioral effects of chronic stress at rat hippocampal temporoammonic synapses. J Neurophysiol. 2015;114:1713–24.
Stanton LM, Price AJ, Manning EE Hypothalamic CRH neurons in stress induced psychopathology: revaluation of endocrine and neural mechanisms. https://doi.org/10.31234/osf.io/2z9gw (2022).
Dayas CV, Buller KM, Crane JW, Xu Y, Day TA. Stressor categorization: acute physical and psychological stressor elicit distinctive recruitment patterns in the amygdala and in medullary noradrenergic cell groups. Eur J Neurosci. 2001;14:1143–52.
Kim J, Lee S, Fang Y-Y, Shin A, Park S, Hashikawa K, et al. Rapid, biphasic CRF neuronal responses encode positive and negative valence. Nat Neurosci. 2019;22:576–85.
Yuan Y, Wu W, Chen M, Cai F, Fan C, Shen W, et al. Reward inhibits paraventricular CRH neurons to relieve stress. Curr Biol. 2019;29:1243–51.e4.
Füzesi T, Daviu N, Wamsteeker Cusulin JI, Bonin RP, Bains JS. Hypothalamic CRH neurons orchestrate complex behaviours after stress. Nat Commun. 2016;7:11937.
Campbell EJ, Mitchell CS, Adams CD, Yeoh JW, Hodgson DM, Graham BA, et al. Chemogenetic activation of the lateral hypothalamus reverses early life stress-induced deficits in motivational drive. Eur J Neurosci. 2017;46:2285–96.
Rodaros D, Caruana DA, Amir S, Stewart J. Corticotropin-releasing factor projections from limbic forebrain and paraventricular nucleus of the hypothalamus to the region of the ventral tegmental area. Neuroscience. 2007;150:8–13.
Madisen L, Zwingman TA, Sunkin SM, Oh SW, Zariwala HA, Gu H, et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat Neurosci. 2010;13:133–40.
Taniguchi H, He M, Wu P, Kim S, Paik R, Sugino K, et al. A resource of cre driver lines for genetic targeting of GABAergic neurons in cerebral cortex. Neuron. 2011;71:995–1013.
Balthasar N, Dalgaard LT, Lee CE, Yu J, Funahashi H, Williams T, et al. Divergence of melanocortin pathways in the control of food intake and energy expenditure. Cell. 2005;123:493–505.
Wamsteeker Cusulin JI, Füzesi T, Watts AG, Bains JS. Characterization of corticotropin-releasing hormone neurons in the paraventricular nucleus of the hypothalamus of Crh-IRES-Cre mutant mice. PLoS ONE. 2013;8:e64943.
Jupp B, Krivdic B, Krstew E, Lawrence AJ. The orexin(1) receptor antagonist SB-334867 dissociates the motivational properties of alcohol and sucrose in rats. Brain Res. 2011;1391:54–9.
Besheer J, Faccidomo S, Grondin JJM, Hodge CW. Regulation of Motivation to Self-Administer Ethanol by mGluR5 in Alcohol-Preferring (P) Rats. Alcohol: Clin Exp Res. 2008;32:209–21.
Bullitt E. Expression of C-fos-like protein as a marker for neuronal activity following noxious stimulation in the rat. J Comp Neurol. 1990;296:517–30.
Paxinos G, Franklin K. The mouse brain in stereotaxic coordinates. Sydney, New South Wales: Academic Press; 2001.
Howell DC. Statistical methods for psychology. 5th ed. Wadsworth; 2002.
Asken MJ, Grossman D, Christensen LW. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. Arlington, VA: American Psychiatric Pub-lishing, 2013. Archibald, Herbert C., and Read D. Tuddenham.“Persistent Stress Reac-tion after Combat: A 20-Year Follow-Up.” Archives of General Psy. Therapy. 2007;45:2317–25.
Carnevali L, Mastorci F, Graiani G, Razzoli M, Trombini M, Pico-Alfonso MA, et al. Social defeat and isolation induce clear signs of a depression-like state, but modest cardiac alterations in wild-type rats. Physiol Behav. 2012;106:142–50.
Miczek KA, Nikulina EM, Shimamoto A, Covington HE 3rd. Escalated or suppressed cocaine reward, tegmental BDNF, and accumbal dopamine caused by episodic versus continuous social stress in rats. J Neurosci. 2011;31:9848–57.
Tran I, Gellner A-K. Long-term effects of chronic stress models in adult mice. J. Neural Transmission. 2023:130;1133–51.
Lethbridge R, Allen NB. Mood induced cognitive and emotional reactivity, life stress, and the prediction of depressive relapse. Behav Res Ther. 2008;46:1142–50.
Leskelä U, Rytsälä H, Komulainen E, Melartin T, Sokero P, Lestelä-Mielonen P, et al. The influence of adversity and perceived social support on the outcome of major depressive disorder in subjects with different levels of depressive symptoms. Psychol Med. 2006;36:779–88.
van Erp AM, Kruk MR, Meelis W, Willekens-Bramer DC. Effect of environmental stressors on time course, variability and form of self-grooming in the rat: handling, social contact, defeat, novelty, restraint and fur moistening. Behav Brain Res. 1994;65:47–55.
Pond HL, Heller AT, Gural BM, McKissick OP, Wilkinson MK, Manzini MC. Digging behavior discrimination test to probe burrowing and exploratory digging in male and female mice. J Neurosci Res. 2021;99:2046–58.
Kedia S, Chattarji S. Marble burying as a test of the delayed anxiogenic effects of acute immobilisation stress in mice. J Neurosci Methods. 2014;233:150–4.
Thomas A, Burant A, Bui N, Graham D, Yuva-Paylor LA, Paylor R. Marble burying reflects a repetitive and perseverative behavior more than novelty-induced anxiety. Psychopharmacology. 2009;204:361–73.
Becker DE. Basic and clinical pharmacology of glucocorticosteroids. Anesthesia Prog. 2013;60:25–32.
Li S-B, Borniger JC, Yamaguchi H, Hédou J, Gaudilliere B, de Lecea L. Hypothalamic circuitry underlying stress-induced insomnia and peripheral immunosuppression. Sci Adv. 2020;6:eabc2590.
Azeez IA, Del Gallo F, Cristino L, Bentivoglio M. Daily fluctuation of orexin neuron activity and wiring: the challenge of “chronoconnectivity. Front Pharmacol. 2018;9:1061.
Jennings JH, Rizzi G, Stamatakis AM, Ung RL, Stuber GD. The inhibitory circuit architecture of the lateral hypothalamus orchestrates feeding. Science. 2013;341:1517–21.
Jennings JH, Ung RL, Resendez SL, Stamatakis AM, Taylor JG, Huang J, et al. Visualizing hypothalamic network dynamics for appetitive and consummatory behaviors. Cell. 2015;160:516–27.
Sharpe MJ, Marchant NJ, Whitaker LR, Richie CT, Zhang YJ, Campbell EJ, et al. Lateral hypothalamic GABAergic neurons encode reward predictions that are relayed to the ventral tegmental area to regulate learning. Curr Biol. 2017;27:2089–100.e5.
Reyes BA, Valentino RJ, Xu G, Van Bockstaele EJ. Hypothalamic projections to locus coeruleus neurons in rat brain. Eur J Neurosci. 2005;22:93–106.
Keay KA, Bandler R. Parallel circuits mediating distinct emotional co** reactions to different types of stress. Neurosci Biobehav Rev. 2001;25:669–78.
Stephenson-Jones M, Yu K, Ahrens S, Tucciarone JM, van Huijstee AN, Mejia LA, et al. A basal ganglia circuit for evaluating action outcomes. Nature. 2016;539:289–93.
Koob GF, Kelley AE, Mason ST. Locus coeruleus lesions: Learning and extinction. Physiol Behav. 1978;20:709–16.
Chen NA, Jupp B, Sztainberg Y, Lebow M, Brown RM, Kim JH, et al. Knockdown of CRF1 receptors in the ventral tegmental area attenuates cue- and acute food deprivation stress-induced cocaine seeking in mice. J Neurosci. 2014;34:11560–70.
Hunt AJ, Dasgupta R, Rajamanickam S, Jiang Z, Beierlein M, Chan CS, et al. Paraventricular hypothalamic and amygdalar CRF neurons synapse in the external globus pallidus. Brain Struct Funct. 2018;223:2685–98.
Hung LW, Neuner S, Polepalli JS, Beier KT, Wright M, Walsh JJ, et al. Gating of social reward by oxytocin in the ventral tegmental area. Science. 2017;357:1406–11.
Wang B, Shaham Y, Zitzman D, Azari S, Wise RA, You ZB. Cocaine experience establishes control of midbrain glutamate and dopamine by corticotropin-releasing factor: a role in stress-induced relapse to drug seeking. J Neurosci. 2005;25:5389–96.
Rho JH, Swanson LW. Neuroendocrine CRF motoneurons: intrahypothalamic axon terminals shown with a new retrograde-Lucifer-immuno method. Brain Res. 1987;436:143–7.
Stricker-Krongrad A, Beck B. Modulation of hypothalamic hypocretin/orexin mRNA expression by glucocorticoids. Biochem Biophys Res Commun. 2002;296:129–33.
Ferguson AV, Latchford KJ, Samson WK. The paraventricular nucleus of the hypothalamus—a potential target for integrative treatment of autonomic dysfunction. Expert Opin Ther Targets. 2008;12:717–27.
Cardozo Pinto DF, Lammel S. Hot topic in optogenetics: new implications of in vivo tissue heating. Nat Neurosci. 2019;22:1039–41.
Maghsoudi N, Ghasemi R, Ghaempanah Z, Ardekani AM, Nooshinfar E, Tahzibi A. Effect of chronic restraint stress on HPA axis activity and expression of BDNF and Trkb in the hippocampus of pregnant rats: possible contribution in depression during pregnancy and postpartum period. Basic Clin Neurosci. 2014;5:131–7.
Juruena MF. Early-life stress and HPA axis trigger recurrent adulthood depression. Epilepsy Behav. 2014;38:148–59.
Acknowledgements
The authors would like to thank Dr Toni-Lee Sterley for sharing the behavior annotator she developed for acute behavioral analysis. This work was supported by grants to CVD from the NHMRC Australia
Author information
Authors and Affiliations
Contributions
CSM, CVD, EEM, TF, JSB, GPM, SDF, BAG contributed to conceptualization and experimental design; CSM, AJP, LMS, NJB, EEM, EJC contributed to data collection; CSM, EJC, SDF contributed to data analysis; CSM, EJC, SDF, EEM, CVD contributed to manuscript preparation; and all authors reviewed and approved final manuscript draft.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mitchell, C.S., Campbell, E.J., Fisher, S.D. et al. Optogenetic recruitment of hypothalamic corticotrophin-releasing-hormone (CRH) neurons reduces motivational drive. Transl Psychiatry 14, 8 (2024). https://doi.org/10.1038/s41398-023-02710-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41398-023-02710-0
- Springer Nature Limited