
Abstract
Context
Managing landscapes to increase multiple services provision in search of higher agricultural yield can be an alternative to agricultural intensification. Nonetheless, to properly guide management, we need to better understand how landscape structure affects multiple services at different scales.
Objectives
Focused on synergies and trade-offs in real-world landscapes, we investigated how and at which scale landscape features related to the supply, demand and flow of services act as a common driver of pollination and pest control in coffee plantations.
Methods
Considering landscapes in an important coffee-producing region in Brazil, we tested the effects of forest and coffee cover, distance to forest, forest-coffee edge density and coffee edge diversity at multiple scales on pollination and pest control by birds, bats and ants.
Results
Coffee edge diversity (number of land uses in contact with coffee) was an important driver of pollination and pest control, being consistently relevant at local scales (up to 300 m). However, services were also affected by other landscape features and the ‘scale of effect’ of these relationships varied. Additionally, results show the complex nature of pest control once the direction of effect revealed services and disservices.
Conclusions
Besides reinforcing the importance of known landscape effects, this study adds to previous studies by showcasing the relevance of diverse land uses around coffee crops as a common driver of pollination and pest control provision by different species. Moreover, we highlight how understanding the combined local and landscape effects may aid in offsetting disservices and tackling the variety of ‘scales of effect’ found.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Agricultural intensification (i.e., increases in agricultural inputs for higher yield) has successfully increased yield across homogenous agricultural landscapes (Winqvist et al. 2012). However, these productivity gains have been at the expense of native species and ecosystem service provision (Tscharntke et al. 2005). To face the high demand for agricultural goods and more sustainable agricultural practices, landscape management that maximizes the provision of multiple ecosystem services can be an important alternative to intensive agriculture (Qiu et al. 2021). Understanding the relative importance of landscape structure and the scale at which multiple ecosystem services respond to landscape structure is necessary to properly guide landscape management (Gagic et al. 2019; Qiu et al. 2021; Le Provost et al. 2023).
Pollination and biological pest control are two of the most important ecosystem services associated with crop yield (Dainese et al. 2019). These services are relevant for economically and culturally important agricultural goods such as coffee, one of the most traded commodities in the world (FAO 2019). Pollinators and natural pest controllers such as ants, birds and bats can increase coffee yield by up to 40% (Saturni et al. 2016; Aristizábal and Metzger 2018). Both pollination and biological pest control are ecosystem services that rely on the flow of species that connect areas of service ‘supply’ (i.e., ecosystems that support key processes or functions, such as natural habitat cover) to areas of ‘demand’ (e.g., cropland where the service is required). The influence of landscape structure on these services is well-studied (see Librán-Embid et al. 2017; Aristizábal and Metzger 2018; Gagic et al. 2019; González-Chaves et al. 2020). Both pollination and pest control tend to be positively affected by the amount and proximity to supply (e.g., native forest cover and distance to forest edge), while being negatively affected by increases in demand (e.g., homogeneous and extensive areas of coffee cover) (Saturni et al. 2016; Aristizábal and Metzger 2018; González-Chaves et al. 2020). However, potential synergies and trade-offs between those services that arise across scales and their consequences for agricultural production are still poorly understood (Chain-Guadarrama et al. 2019; Qiu et al. 2021, but see Martínez-Salinas et al. 2022). Further, more systematised and scale-standardised comparisons that account for the potential effects of different service providers (e.g., birds and ants acting as natural pest controllers) on the synergies and trade-offs relationships between those services are still lacking (Chain-Guadarrama et al. 2019).
Pollination and pest control services thus have similar spatial relationships with the three components of the ‘provision chain’ (supply, demand and flow; Metzger et al. 2021). Although pollination and pest control may be similarly linked to the provision chain, the scale at which landscape characteristics affect these services may differ. Indeed, this ‘scale of effect’ may differ with service, yield parameter and landscape attribute investigated (Martin 2018). The scale of effect is also expected to vary with characteristics of the species that mediate provision (e.g., movement range and body size), and with the ecological processes involved in service provision (e.g., species spillover and pollination efficiency). For example, services that rely on species that have large habitat requirements, such as pest control by birds and bats, may be affected by landscape composition at broad scales (Librán-Embid et al. 2017). By contrast, ecological processes that occur at more local scales and that may benefit pollination and pest control, such as local species spillover between neighbouring land uses, may be influenced by landscape components at smaller scales (Saturni et al. 2016; González-Chaves et al. 2020). Moreover, services that involve multiple species, with different biological characteristics, may be affected by multiple scales (Metzger et al. 2021). The identification of pertinent scales of effect for different services is essential to guide landscape planning for multiple service provisions, and understanding if and at which scales landscape features may be beneficial to both pollination and pest control is still a top research priority (Chain-Guadarrama et al. 2019). General management recommendations that fail to account for the complex relationships between landscape structure and multiple provisions at different scales may not succeed in creating synergies and avoiding trade-offs between services (Garibaldi et al. 2018).
Here, we combined a landscape ecological approach with exclusion experiments to investigate if and at what spatial scales landscape structure affects pollination and pest control by ants, birds and bats in coffee plantations. Specifically, we assessed whether landscape features linked to the supply, demand and flow of pollination and pest control could modulate the provision of both services and the resulting synergies and trade-offs between them. We used a standardised multiscalar approach to investigate whether important known landscape features such as the amount of forest (related to supply—%) and coffee cover (demand—%) and the distance to forest fragments (flow—m) similarly modulated the provision of pollination and pest control by three different service providers (birds, bats and ants), addressing a key gap in the literature. Also, we investigated whether novel features such as forest-coffee edge density (flow—m/pi.r2) and the diversity of land uses around coffee (supply and flow—the number of neighboring land uses to coffee within the surrounding landscape) would also play a role as a common driver of pollination and pest control across scales. With these, we hope to move further ‘distance effects’ and investigate in more detail the potential effects of forest-coffee flow, as well as aid in unravelling if and how other land uses of the landscape may affect the provision of pollination and pest control.
We predicted pollination and pest control to increase with forest cover and density of forest-coffee edges, and to decrease with increasing distance to forest and coffee cover. We expected both services to increase with the diversity of land uses in contact with coffee patches, given the greater potential of heterogeneous landscapes to maintain a diversity of species involved in pollination and pest control services (Chain-Guadarrama et al. 2019, Varah et al. 2020). Furthermore, we expected the scale of effect to vary with species and service, with pollination and pest control by ants affected by landscape structure at smaller scales, while pest control by birds and bats affected at broader scales. Finally, we also expected that landscape attributes associated with the flow of species between supply and demand patches, such as the distance to forest edges, the density of forest and coffee edges and the diversity of land uses in contact with coffee patches, would have a more local effect on both pollination and pest control provision.
Methods
Study area
Our study took place in one of the most important and traditional coffee-producing regions in Brazil, in the southeastern region of the country, between São Paulo and Minas Gerais states (Fig. 1a). This region alone is responsible for almost 25% of the country’s production, with Brazil being the main exporter and accounting on average for 30–35% of the total coffee production worldwide (FAO 2019). Coffee crops in the region are sun-grown coffee plantations (sun-exposed coffee trees with no canopy cover planted on previously cleared land), mainly Coffea arabica cultivated by family farmers (properties of approximately 20–65 ha, with traditionally 2000–8000 coffee plants/ha and managed mostly without mechanisation). The Brazilian Atlantic Forest, the original biome of the region and one of the most diverse but also threatened biomes in the world, has been drastically reduced due to generalised agricultural expansion at the beginning of the twentieth century (Carlucci et al. 2021). A subtropical climate with mean temperatures between 13.6 and 20.4 °C, dry winter and rainy summers and a hilly terrain with elevation varying between 700 to 1300 m.a.s.l., make this region ideal for coffee production. In fact, coffee expansion was one of the main drivers of deforestation of the Atlantic Forest, despite its potential in supplying pollinators and potential natural pest controllers (Saturni et al. 2016; Medeiros et al. 2019; Carlucci et al. 2021). Nowadays, landscapes in the region are composed of Atlantic Forest remnants immersed primarily in a matrix of coffee crops and pastures, and secondarily by eucalyptus and sugarcane plantations (Fig. 1).

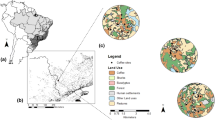
Geographical location of the ten landscapes in the Brazilian Atlantic Forest region, between São Paulo and Minas Gerais states (a). Distribution of experimental sites in one of the 3 km landscapes (b), and a zoom into pollination, pest control by ants and pest control by birds and bats sampling sites in coffee plantations (c)
Field data collection occurred between 2013 and 2015 and across 10 independent experimental landscapes with a 3-km-radius (Fig. 1a; for more details, see Saturni et al. 2016; Librán-Embid et al. 2017; Aristizábal and Metzger 2018; González-Chaves et al. 2020) as part of Projeto Interface (http://ecologia.ib.usp.br/projetointerface/en/), a large, 5-year, collaborative project that focused on assessing ecosystem service provision across forested agricultural landscapes. Landscapes were selected to encompass a gradient of landscape forest cover (13% to 48%), but controlling for soil type and altitude (for selection details, see Boesing et al. 2018a, b). Landscape centroids were at least 6 km apart from each other. Land use and land cover were mapped using high-resolution images (ArcGIS 10.3 base map imagery, DigitalGlobe satellites 2010–2011, 0.5–1 m resolution, 1:5000 visualization scale). Map** was extensively validated in the field in 2013 and 2015, and all interpretation errors or landscape changes between 2010 and 2015 were edited.
To assess the contribution of pollinators and pest controllers to pollination and biological pest control, respectively, each landscape had multiple experimental sites within coffee farms with exclusion experiments, totalling 292 experimental sites across ten 3-km landscapes (Fig. 1a). All experimental sites were separated by at least 100 m to ensure independence among samples while encompassing a wide variation of landscape attributes, such as local forest and coffee cover and distance to forest edge (for more details, see Saturni et al. 2016; Librán-Embid et al. 2017; Aristizábal and Metzger 2018; González-Chaves et al. 2020). In addition, the sub-varieties of Coffea arabica in our study were controlled, with sites only composed of varieties that are physiologically similar, such as Catuaí and Catucaí, or Catuaí and Mundo Novo (Dias and Souza 2015, but see Saturni et al. 2016).
Landscape metrics
For all ten experimental landscapes, we calculated five landscape metrics associated with one or more components of the provision chain. Forest cover and coffee edge diversity were selected as potential supply measures, acting as likely sources of pollinators and natural pest controllers (Medeiros et al. 2019), potentially affecting both the abundance and diversity of those species. However, as agricultural matrices may often offer movement resistance to species, particularly habitat-dependent ones, due to structural differences to the habitat (Boesing et al. 2018a, b; Hohlenwerger et al. 2022) and higher predation risk outside-habitat (Brown 1999), coffee edge diversity, similar to forest-coffee edge density and distance to forest edge, could also act as an indication of flow, meaning that land uses in contact with coffee patches could facilitate the flow of species between supply and demand. Finally, coffee cover was used as an indication of areas demanding the diversity of pollinators and natural pest controllers being supplied by the forest or other land uses.
We calculated four landscape metrics at multiple spatial scales using circular buffers around each of the 292 experimental sites: forest and coffee cover (%), forest-coffee edge density (m/pi.r2) and coffee edge diversity (number of land uses in contact with coffee areas). Additionally, we calculated the distance to forest edge (m) as the Euclidean distance of each experimental site to the nearest forest patch. We evaluated pollination and pest control provisions through a systematic multiscalar approach focused on the ‘scale of effect’ (the landscape extent/buffer size at which an ecological response or process is best predicted; Jackson and Fahrig 2015). This means standardizing all datasets and analyzes to allow comparison if and how these landscape attributes related to supply, demand and flow differently affect these services across different scales. Therefore, for all four scale-dependent metrics, we first considered a wide variety of service-relevant scales (30, 50, 100 up to 500 with 100 m intervals, 1000, 2000 and 3000 m) around each experimental site to encompass spatial dimensions for a variety of ecological traits of possible pollinators and pest controllers (Kremen et al 2004; Boscolo and Metzger 2009; Bichara et al. 2010; Jackson and Fahrig 2015; Boesing et al. 2018a, b, Baensch et al. 2020). Afterwards, we selected the appropriate ‘scale of effect’ of each individual landscape metric based on the Akaike Information Criterion corrected for small sample sizes (AICc) using the “multifit” function in R (version 4.2.2—R Development Core Team 2014; see Huais 2018 for methodological details) (Table S1). Considering then the appropriate ‘scale of effect’ of each scale-dependent landscape metric evaluated (Table S1), experimental sites encompassed a wide overall range of forest cover (0–61%), coffee cover (4–100%), forest-coffee edge density (0–0.00842 m/pi.r2), coffee edge diversity (one to eight neighbouring land uses) and distance to the forest (0–324 m). Not all possible variability of each scale-dependent explanatory variable (e.g., eight neighbouring land uses) will be observed in the final models (Table S3) as some were dropped in the process of model simplification. All five landscape metrics were calculated in R (R Development Core Team 2014).
Exclusion experiments
Pollination
To quantify the service of coffee pollination, we obtained data from Saturni et al. (2016) and González-Chaves et al. (2020). These data were collected through 159 pollination-exclusion experiments placed across experimental sites in nine landscapes (Fig. 1b, c). Each experiment encompassed exclusion branches (i.e. branches in which the presence of pollinators was experimentally prevented) paired with control branches (i.e. branches in which the flowers were opened and accessible to pollinators). Paired branches from both treatments were randomly selected across different coffee bushes at the experimental sites. To exclude pollinators but still allow airflow, a thin mesh bag (3 mm) was placed around each exclusion branch. Coffee plants bloom for about three days (van der Vossen 1985). In the region, coffee bloom periods may encompass two blooming seasons several weeks apart from each other. After approximately a week of this flowering period, exclusion branches were marked and had all mesh bags removed to avoid any effects on fruit development. Altogether, pollination-exclusion experiments totalled an effort of 765 branches per treatment. Finally, for each branch on both treatments, the number of flowers at the blooming period (September and October 2013, and October 2015) and the number of coffee berries at the harvest period (May 2013 and 2015) were counted (see Online Appendix S2, Saturni et al. 2016; González-Chaves et al. 2020 for detailed information on sampling design and procedures detail).
Pest control
To quantify the service of pest control, we investigated the potential of natural control of two of the main pests of coffee plantations: the Coffee Leaf Miner (CLM—Leucoptera coffeella) and the Coffee Berry Borer (CBB—Hypothenemus hampei). Here, we sourced data from Librán-Embid et al. (2017) and Aristizábal and Metzger (2018). Combined, these data were collected through natural pest control-exclusion experiments for these two coffee pests placed across experimental sites in ten landscapes (Fig. 1b).
For CLM, we considered the control by birds and bats, and used data from 24 exclusion experiments placed across eight landscapes. Exclusion experiments were composed of four coffee bushes in which birds and bats were excluded (i.e. exclusion units), paired with another four adjacent coffee bushes that were accessible to all potential pest controllers (i.e. control units), resulting in 576 branches per treatment. To exclude birds and bats while still allowing arthropods to enter, a nylon monofilament mesh was used to completely cover all four exclusion bushes. Each coffee bush, regardless of the treatment, then had randomly marked branches from which randomly selected leaves and coffee berries were marked and counted (see Online Appendix S2 and Librán-Embid et al. 2017 for detailed information on sampling design and exclusion experiments).
For CBB, we considered control by ants, and used data on 29 ant-exclusion experiments placed across 10 landscapes right before the main flowering to ensure CBB absence (see Online Appendix S2). At each of the 10 coffee bushes per experimental site, exclusion experiments encompassed one randomly selected branch in which ants were excluded paired to another randomly selected neighboring branch that was accessible to ants. Altogether, CBB control-exclusion experiments totalled an effort of 290 branches per treatment. For each experimental branch, regardless of the treatment, the number of berries produced and berries bored was counted, and all coffee berries were collected right before farmers’ harvest time. Care was taken to ensure a similar number of flowers, height, and position to all experimental branches (see Online Appendix S2 and Aristizábal and Metzger 2018 for detailed information on sampling design and procedure details).
Quantification of pollination and pest control service
We used five response variables, indicators of pollination and pest control by birds, bats and ants, to quantify how landscape structure could be affecting drivers associated with the supply, demand and flow of these services. Therefore, we used fruit set as a measure of fruit production and a proxy of pollination service. Since CLM affects leaves’ persistence in coffee bushes which in turn affects fruit production due to photosynthesis reduction (Pierre 2011), we also used fruit set, along with leaf loss, as indications of natural coffee leaf miner control by birds and bats. Finally, for CBB control by ants, we used CBB presence, CBB infestation’s level, and CBB bean damage to consider the ecological interactions between ants and CBB at the three different stages of pest control. For pollination, as well as for CLM control by birds and bats, fruit set was calculated as the proportion of berries out of the total of flowers per branch during the blooming period. Leaf loss was calculated as the proportion of fallen leaves out of the total leaves initially marked per branch. The presence of CBB was considered when there was at least one bored coffee berry on a branch, while CBB absence was considered when none of the coffee berries on a branch had CBB. CBB infestation was calculated as the proportion of coffee berries with CBB out of the total of berries produced per branch. Finally, for each bored coffee berry, CBB damage was calculated as the proportion of damage to each coffee bean out of the total size of the coffee bean inside bored berry (see Aristizábal and Metzger 2018 for damage measurement details).
Data analysis
We quantified the effects of forest cover, coffee cover, distance to forest edge, forest-coffee edge density and coffee edge diversity on all three services using generalized linear mixed models (GLMMs; Zuur et al. 2009). All three response variables for pest control by ants (CBB presence, CBB infestation and CBB damage), and the response variable for pollination (fruit set) were modeled using a binomial distribution and a logit-link function (“lme4” package; Bates et al. 2015). The two response variables for pest control by birds and bats (fruit set and leaf loss) were modeled using a beta-binomial distribution (“glmmTMB” package; Magnusson et al. 2017) to account for overdispersion (Harrison 2015). As the total number of flowers, leaves and berries differed between branches, response variables were calculated inside the models as a two-vector response variable (e.g., number of coffee berries from the total number of flowers per branch) using the “cbind” function (R Development Core Team 2014). All five landscape attributes were centred and scaled to mean zero and one standard deviation and included as fixed effects in the full model.
To account for the fact that our pollination measurement (fruit set) was collected in different years (2013 and 2014), the measurement year was included as a fixed effect in all models with this service (Bolker et al. 2013). For each service, we created ‘full models’ that included the variables: forest cover, coffee cover, distance to forest edge, forest-coffee edge density and coffee edge diversity. The full model included the interaction of the animal exclusion experiments (presence and absence of animals in branches or flowers), all landscape metrics (each at their final ‘scale of effect’) (Table S1), and random effects (details below). The scale-dependent landscapes metrics were specified at the appropriate “scale of effect” (further details in section “Landscape metrics” and in Online Appendix S1). To account for potential non-independence of multiple sampling transects within each landscape, a landscape identifier was specified as a random intercept (Zuur et al. 2009). As experimental units usually had more than one experimental coffee bush, and each bush had more than one experimental branch, both the experimental unit and bush identifications were also included as nested random effects in all models (Zuur et al. 2009). In addition, to account for overdispersion, an observation-level random effect was included in all pollination models (Harrison 2015).
For each response variable, we separately performed ‘all subsets’ model selection procedure for each full model using the “dredge” function (“MuMin” package; Barton and Barton 2015) with the full model for each service and the Akaike Information Criterion corrected for small sample sizes (AICc). Substantially supported models (ΔAICc < 2) were predicted and averaged, with final models being built using significant relationships for each service (Table S2). Model assumptions were verified by plotting residuals versus fitted values. Model validation indicated no problems. Multicollinearity was measured and adjusted using a generalized variance-inflation factor (cutoff > 4—“car” package—Fox et al. 2007). All analyses were conducted in R (R Development Core Team 2014). Results and model predictions were graphed from the minimum adequate model, with other covariates variables held at their mean and on an additive measurement scale (i.e., proportions of fruit set and leaf loss), rather than the multiplicative measurement scale of the linear predictor, as the additive scale was considered the most relevant to management (Spake et al. 2023).
Results
Mostly, pollinators and natural pest controllers increased coffee productivity by affecting the amount of fruit, leaf and pest levels on coffee plants. However, their contribution to those ecosystem services varied across landscape gradients, sometimes even changing in the direction of effect (i.e., providing service and disservice). While the presence of pollinators always resulted in service provision (higher fruit set after accounting for the difference between the presence and absence of animals, i.e., hatched areas between tendency lines in graphs), the presence of birds, bats and ants resulted in both service and disservice (i.e., contributing to both an increment and reduction in fruit set, leaf loss, and CBB infestation and damage after accounting for the difference between the presence and absence of animals). Although landscape structure exerted different effects on each of the services, the number of land uses in contact with coffee (coffee edge diversity) was the most common driver of pollination, pest control by ants, and pest control by birds and bats (Table 1). Furthermore, for both types of services, the effect of coffee edge diversity on provision was consistently local (30, 100, 200 and 300 m). Contrary to coffee edge diversity, the ‘scale of effect’ of all other scale-dependent landscape structure components (forest cover, coffee cover and forest-coffee edge density) varied greatly between services (from 100 m up to 2 km; see Table 1).
Pollination provision
The presence of bees during the pollination experiments consistently led to a higher fruit set, as observed by the positive effect of their presence compared to their absence (i.e. orange areas in Fig. 2). Pollination provision then decreased with an increase in coffee cover within 1 km and distance from the forest edge (shown by the reduction in the width of hatched areas in Figs. 2a and 3b; Table S2). However, when it came to coffee edge diversity within 100 m, the service of pollination by bees on fruit set increased as the number of land uses in contact with coffee plots increased (shown by the increment in the width of hatched areas in Fig. 2c; Table S3; Conditional R2: 54.5% and marginal R2: 15.6%).

Partial effects of the relationships between the proportion of fruit set with (orange lines with 95% confidence interval) and without (purple lines with confidence interval) bees’ contribution to pollination and a coffee cover (%), b coffee edge diversity (n) and c distance to forest edge (m). The difference between trend lines (hatched areas) represents the service or disservice provision resulting from the interaction between exclusion experiments (bees’ presence/absence), while the intensity of provision is represented by the width of the hatched area along the gradient of the predictor variable. Hatched areas with horizontal orange lines represent where bees had positive effects (i.e., service provision)

Partial effects of the relationships between the proportion of fruit set with (orange lines with confidence interval) and without (purple lines with confidence interval) the contribution of birds and bats to CLM pest control and a forest-coffee edge density (m/pi.r2) and b coffee edge diversity (n). The difference between trend lines (hatched areas) represents the service or disservice provision resulting from the interaction between exclusion experiments (birds and bats’ presence/absence), while the intensity of provision is represented by the width of the hatched area along the gradient of the predictor variable. Hatched areas with horizontal orange lines and vertical purple lines indicate where birds and bats had positive (i.e., service) and negative effects (i.e., disservice), respectively
Coffee leaf miner (CLM) control
The effects of bird and bat exclusion on fruit set were influenced by forest-coffee edge density and coffee edge diversity, measured within 300-m (Table S3; Conditional R2: 37.2% and marginal R2: 18.5%). For both landscape attributes and contrary to the patterns found for pollination provision, the presence of birds and bats did not always have a positive impact on fruit set, thus providing both service (i.e., a positive effect of the presence of birds and bats; orange areas in Fig. 3) and disservice to coffee productivity (i.e., a negative effect of the presence of birds and bats; purple areas in Fig. 3). The presence of birds and bats increased fruit set (i.e., service provision; orange areas in Fig. 3a) in landscapes with higher densities of forest-cover edges, and where there were more than three different land uses in contact with coffee patches (Fig. 3b).
Similar to the patterns observed for fruit set in CLM control experiments, birds and bats’ contribution to leaf loss switched direction across landscape gradients, thus again representing both service (i.e., a reduction in leaf loss after accounting for the difference between the presence and absence of birds and bats) and disservice (i.e., the opposite, an increment in leaf loss) (Fig. 4). The provision of leaf loss control by birds and bats responded to the greatest variability of scales and landscape metrics with forest and coffee cover (within 100 and 500-m, respectively), forest-coffee edge density (within 1-km), and distance to forest edge (see models details in Table S3; Conditional R2: 35.7% and marginal R2: 15.7%). The presence of birds and bats was associated with a reduction in leaf loss (i.e., service provision; orange areas in Fig. 4) in landscapes with high forest cover (Fig. 4a) and high forest-coffee edge densities (Fig. 4c), and with low coffee cover (Fig. 4b), and far (~ >100 m) from forest edges (Fig. 4d).

Partial effects of the relationships between the proportion of leaf loss with (orange lines with confidence interval) and without (purple lines with confidence interval) birds and bats’ contribution to CLM pest control with a forest cover (%), b coffee cover (%), c forest-coffee edge density (m/pi.r2) and d distance to forest edge (m). The difference between trend lines (hatched areas) represents the service or disservice provision resulting from the interaction between exclusion experiments (birds and bats’ presence/absence), while the intensity of provision is represented by the width of the hatched area along the gradient of the predictor variable. Hatched areas with horizontal orange lines and vertical purple lines represent where birds and bats had positive effects (i.e., service) and negative effects (i.e., disservice), respectively
Coffee berry borer (CBB) control
For CBB regulation by ants, CBB presence varied with 1-km-level coffee cover and 200-m-level coffee edge diversity (Table S3; Conditional R2: 48.2% and marginal R2: 11.5%). As coffee cover increased (Fig. 5a), the contribution of ants to the reduction of CBB presence (i.e., service provision; orange areas in Fig. 5) diminished. However, the opposite was true for coffee edge diversity, with service provision increasing as the number of land uses in contact with coffee plots increases (Fig. 5b). Therefore, the contribution of ants in the reduction of CBB presence was higher in landscapes where coffee cover does not dominate the landscape and coffee patches are surrounded by multiple land uses.

Partial effects of the relationships between the probability of CBB presence with (orange lines with confidence interval) and without (purple lines with confidence interval) ants’ contribution to CBB pest control with coffee cover (%) and coffee edge diversity (n). The difference between trend lines (hatched areas) represents the service or disservice provision resulting from the interaction between exclusion experiments (ants’ presence/absence), while the intensity of provision is represented by the width of the hatched area along the gradient of the predictor variable. Hatched areas with horizontal orange lines represent where ants had positive effects (i.e., service) and with vertical purple lines, negative effects (i.e., disservice)
The control of CBB infestation by ants was influenced by 1-km-level amount of forest-coffee edge density and 30-m-level coffee edge diversity (Table S3; Conditional R2: 39.4% and marginal R2: 12.8%). Similar to the complex relationships found for CLM control by birds and bats, ants’ contribution to CBB infestation also changed the direction of effect across landscape gradients generating both service (i.e., a positive effect of the presence of ants in reducing CBB infestation) and disservice (i.e., a negative effect of the presence of ants). However, CBB infestation, contrary to CLM by bats and birds, responded negatively to forest-coffee edge density and to coffee edge diversity (orange and purple areas in Fig. 6). Specifically, ants’ contribution to the reduction of CBB infestation (service provision; orange areas in Fig. 6) was higher in landscapes with lower forest-coffee edge density and coffee edge diversity (Fig. 6a, b, respectively).

Partial effects of the relationships between the proportion of CBB infestation with (orange lines with confidence interval) and without (purple lines with confidence interval) ants’ contribution to CBB pest control with forest-coffee edge density (m/pi.r2) and coffee edge diversity (n). The difference between trend lines (hatched areas) represents the service or disservice provision resulting from the interaction between exclusion experiments (ants’ presence/absence), while the intensity of provision is represented by the width of the hatched area along the gradient of the predictor variable. Hatched areas with horizontal orange lines and vertical purple lines represent where ants had positive effects (i.e., service) and negative effects (i.e., disservices), respectively
Once CBB was present, the damage caused by the borer (CBB damage) on coffee beans was greater in landscapes with higher forest cover at 2 km level (Table S3; Conditional R2: 44.8% and marginal R2: 13.9%). Similar to other results for pest control, ants’ contribution to CBB damage also shifted the direction of effect across different forest covers. As forest cover increases, the provision of damage control by ants reduces. Despite this reduction in the provision, up to approximately 40% of forest cover, the presence of ants effectively reduces CBB damage (service provision; orange areas in Fig. 7). After this point, the presence of ants results in a disservice to the amount of CBB damage on coffee beans (purple areas in Fig. 7).

Partial effects of the relationship between the proportion of CBB damage with (orange lines with confidence interval) and without (purple lines with confidence interval) ants’ contribution to CBB pest control with forest cover (%). The difference between trend lines (hatched areas) represents the service or disservice provision resulting from the interaction between exclusion experiments (ants’ presence/absence), while the intensity of provision is represented by the width of the hatched area along the gradient of the predictor variable. Hatched areas with horizontal orange lines and vertical purple lines represent where ants had positive effects (i.e., service) and negative effects (i.e., disservice), respectively
Discussion
Our results provide evidence that the provision of pollination, pest control by ants, and pest control by birds and bats in coffee plantations are modulated by landscape attributes associated with the supply, demand and flow of these services at multiple scales (Table 1). Most importantly, the number of land uses in contact with coffee (i.e. coffee edge diversity) was the main common driver of both pollination and pest control services, with the ‘scale of effect’ of this feature being consistently small for both services (up to 300-m). These novel results suggest that besides traditionally relevant landscape features such as forest and coffee cover, land uses surrounding coffee areas may also enhance service provision by supplying a diverse set of pollinators and pest controllers. Further, the small ‘scale of effect’ of these relationships suggests that the local flow of species (e.g., species spillover) is key to the provision of these services in coffee plantations. Interestingly, while the presence of pollinators consistently resulted in service provision (i.e., the presence of pollinators resulting in higher yield), the presence of ants, birds and bats resulted in both service and disservice depending on the landscape structure. This difference between services appeared to be consistent also for the new and relevant effects found for forest-cover edge density and coffee edge diversity. Finally, besides coffee edge diversity, pollination, pest control by ants and pest control by birds and bats were also affected by other components of landscape structure (see Table 1) and the scale of these effects varied greatly between services. This variation in other landscape structural effects was true even when comparing within pest control services. These complex results, along with the provision of pest control service and disservice, highlight and bring new insights into the challenges involved in planning for multiple service provisions.
Landscape structure effects on the supply, demand and flows of pollination and pest control
Our work shows that pollination and pest control provisions rely on landscape attributes associated with the supply (forest cover and coffee edge diversity), demand (coffee cover) and flows (forest-coffee edge density, coffee edge diversity and distance to forest edge). With regards to services' supply, our results reinforce the relevance of the amount of native forest particularly to the provision of pest control while showcasing the complexity of this service through the different effects of forest cover depending on the type of pest controller.
At local scales, higher forest cover resulted in the reduction of leaf loss mediated by birds and bats (i.e., service provision), supporting that forest patches may indeed be acting as supply areas of pest controllers. Higher local forest cover may favour connectivity between patches and thus foster species movement across the landscape (Martensen et al. 2012). In contrast, low forest cover may drastically reduce the number of edges between native habitat and matrix and increase patch isolation (Villard and Metzger 2014), which could then reduce the probability of species spilling over from habitat patches (Boesing et al. 2018b). At larger landscape scales, however, increases in forest cover reduced the provision of CBB damage control by ants. In landscapes with higher forest cover (~ > 40%), as the diversity of birds increases, ants' contribution to the service may be hindered due to higher predation pressure upon ants by birds (Librán-Embid et al. 2017; Boesing et al. 2018a). Although in this study we only investigated forest cover effects, when studies considered the interaction between the total number of ants seen in branches and forest cover it showed that more ants seen at branches are linked to higher CBB damage (Aristizábal and Metzger 2018). Intraguild predation by birds can then be limiting ants’ ability to control CBB damage, resulting in a disservice via the release of predation pressure on CBB. However, for landscapes with up to 40% of forest cover, the presence of ants was particularly important to reduce CBB damage (i.e., service provision). In these landscapes (~ < 40%), as the community of birds becomes less diverse (Boesing et al. 2018a), intraguild predation pressure on ants is expected to be relaxed, thus increasing ants' relevance as natural controllers of CBB (Aristizábal and Metzger 2018).
Regardless of the scale, our results build on previous studies by showing that higher demand (i.e., coffee cover) reduces the provision not only of pollination but also of pest control. Increases in coffee cover are known for possibly reducing service provision due to dilution effects, meaning that there is more demand for pollinators and pest controllers than they can actually meet (Vanbergen 2013). Additionally, increasing coffee cover can also reduce the diversity of pest controllers and pollinators due to the negative effects of landscape simplification and homogenization (Gámez-Virués et al. 2015; Gonzalez-Chaves et al. 2020), while facilitating pest movement and spread (Medeiros et al. 2019). Moreover, the effects of landscape homogenization at larger scales are accentuated in the absence of forest fragments and can negatively affect regional coffee productivity (González-Chaves et al. 2022). Finally, our study shows that these negative effects of large amounts of coffee cover on pollination and pest control are in accordance with the positive effects found here of higher coffee edge diversity on the provision of these services. This reinforces the relevance of higher matrix heterogeneity to pollination and pest control provision (Aguilera et al. 2020).
Besides the effects of the amount of supply (i.e., native forest areas) and demand (i.e., coffee areas), our results show that pollination and pest control provisions are also shaped by flow-related landscape attributes. As expected, as the distance to forest edge increased, pollination provision decreased, suggesting that the benefit derived from supply areas is stronger near habitat-matrix edges. This can be due to the movement resistance that anthropogenic matrices impose on the species (Brown 1999), hindering longer incursions into the matrix and thus altering the service they provide (Boesing et al. 2018b; Hohlenwerger et al. 2022). Indeed, the diversity of species of bees (mainly small-size bees and tree cavities nesting bees) tends to decrease as distance to forest increases (Gonzalez-Chaves et al. 2020). These negative distance effects then suggest that the contact between supply and demand patches should also play a role in service provision. This was the case for the service of CLM control by birds and bats, for which higher supply-demand contact (i.e., forest-coffee edge density) resulted in higher provision. Indeed, higher edge densities in the region have been reported to have positive effects on the spillover of birds probably due to the higher chance of edge encounters by habitat species (Boesing et al. 2018b). Therefore, our results move beyond distance effects suggesting that the flow of species and, therefore, service provision are not only shaped by the distance between supply and demand patches but also by the amount of contact between those patches.
However, not all services responded equally to forest-coffee edge densities and distance to forest, this being the case for the regulation of CBB infestation by ants and leaf loss by birds and bats. For these services, the provision of pest control happened in landscapes with lower forest-coffee edge density and far from the forest, respectively. Higher contribution of non-forested species or intraguild predation are two of the main processes which could be underlying these results. An increase in forest-coffee edge densities and proximity to forest could result in higher competition for resources with forest species and lower abundance of generalist species, especially near habitat-matrix edges (Boesing et al. 2018b). In landscapes with high forest-coffee edges, it is possible that the mechanisms underlying the higher contribution of birds and bats to CLM control may also be resulting in higher predation pressure on ants by birds (Martin et al. 2013), thus resulting in a reduced contribution of ants to the control of CBB infestation. Although forest-coffee edge density effects on CBB infestation are weak, the opposite effect of this variable at the same scale of effect for leaf loss by CLM control reinforces, through new evidence, the potential relevance of trophic interactions in sha** overall natural pest control effects on coffee plantations. Likewise, it is possible that near forest edges, birds and bats exert predation pressure on other invertebrates besides crop pests, including predatory arthropods such as ants and wasps (Karp and Daily 2014), thus diluting their contribution to pest control and reducing the ability of other pest controllers to provide the service themselves (Martin et al. 2013).
Landscape diversity as a common and local driver of pollination and pest control
Coffee edge diversity was the main common driver for both ecosystem services, suggesting heterogeneous landscapes may favour multiple service provision. Positive effects of the diversity of land uses surrounding coffee plots suggest that these areas may be acting as supply of multiple service-providing species or facilitating the flow of those species, contributing thus to higher service provision. Heterogeneous landscapes may increase service provision by (i) offering a greater diversity of pollinators and natural pest controllers (Aguilera et al. 2020); (ii) providing complementary or supplementary resources to forest-dependent pollinators and natural pest controllers (e.g., nesting areas and secondary food resources) (Alignier et al. 2020); and by (iii) increasing landscape connectivity (Martensen et al. 2012). Finally, higher landscape heterogeneity may hinder pest movement and thus reduce pests abundance (Medeiros et al. 2019). However, for CBB infestation, higher coffee edge diversity resulted in a disservice to CBB infestation. Different ant species may play different roles in controlling CBB colonization (i.e., defining CBB presence) and regulating CBB infestation levels (Moris and Perfecto 2016; Aristizábal and Metzger 2018). Thus, although this effect was weak, it brings new insights that possible positive effects of coffee edge diversity on CBB control through higher species richness, may not necessarily translate to higher regulation of CBB infestation. Additionally, as the efficiency in removing pests also varies among ant species (Way and Khoo 1992), an increase in coffee edge diversity may indirectly alter the diversity of species that are more efficient pest controllers once CBB is present. Therefore, our results highlight the importance of considering landscape structural effects not only on different services (e.g., pollination and pest control) but also within the same type of service (e.g., pest control by birds and bats and pest control by ants) and among different aspects of a single service (e.g., CBB presence and infestation).
Coffee edge diversity’s effects on service provision were consistently local. This small ‘scale of effect’ suggests that processes associated with the local flow of species, such as species spillover at the edges of the coffee fields, are key to the provision of both services. Our results build on previous findings which highlighted the importance of species spillover to pollination and pest control provision (Boesing et al. 2018b; Aristizábal and Metzger 2018; Gonzalez-Chaves et al. 2020), and moves further by showing that other types of land use besides forest patches can also shape service provision by determining access to supply through local flow. Furthermore, our results show that although forest cover (i.e. supply) has been reported by past studies as a relevant landscape feature alongside other variables (e.g., coffee cover and the total number of ants seen in branches) in reducing CBB presence and infestation (Aristizábal and Metzger 2018), as well as increasing fruit set under CLM control (Librán-Embid et al. 2017), this feature became less relevant when other supply and flow related landscape features (i.e., coffee edge diversity and forest-cofee edge density) were considered. Therefore, our study brings new evidence on the importance of considering different land uses when assessing the local flows and supplies of pollination and pest control. Finally, as different land uses have different degrees of usability to the species due to their structural characteristics and resource availability (Boesing et al. 2018b; Hohlenwerger et al. 2022), we suggest incorporating the effect of the degree of matrix usability (i.e., the probability of an organism to use a given matrix for resources such as nesting, foraging, shelter etc) on service-providers species and on service provision itself in future studies.
Challenges for landscape management
One of the main challenges when planning multiple service provisions is avoiding disservices and taking advantage of the potential synergies among services (Martin et al. 2013; Garibaldi et al. 2018; Martínez-Salinas et al. 2022). Our study highlights that this is particularly important when considering the provision of natural pest control, for which we had both services and disservices. Besides supporting previous findings on service and disservice provision of pest control (see Librán-Embid et al. 2017; Aristizábal and Metzger 2018), our study accentuates the complexity of natural pest control by showcasing new evidence of pest control service and disservice through the effects of forest-coffee edge density and coffee edge diversity. That is, our results highlight that even when new landscape features are considered, changes in the direction of effect seem to continue to appear for pest control provision. Our results indicate that disservices may be linked to antagonist interactions between different pest controllers (e.g., birds and ants) and their common prey. As the community of species in the landscape shifts, so do species interactions (Librán-Embid et al. 2021). Indeed, changes in the abundance of birds and bats in the landscape have been reported to influence the abundance of predatory arthropods including ants (Karp and Daily 2014), as well as intraguild predation having a negative effect on pest control (Martin et al. 2013). Therefore, we propose that landscape management practices aimed at enhancing biological pest control should incorporate these complex trophic interactions to foster landscape designs that also offset possible negative effects. We acknowledge that service provisions are a product of many interacting processes related to ecological systems and influenced by human actions (Fu et al. 2013), and suggest that future studies focus on the effect of these non-intuitive and complex processes that may also be sha** provision.
Finally, besides fostering high diversity of land uses at local scales, our results highlight the pressing need to integrate multiple scales when assessing several ecosystem services provisions in working landscapes (Spake et al. 2019; Metzger et al. 2021). Apart from coffee edge diversity, the ‘scale of effects’ of all other scale-dependent variables (forest cover, coffee cover and forest-coffee edge density) varied across services (Table 1). Specifically, from the management perspective, these results present opportunities to combine within-farm and landscape-level management actions (Tscharntke et al. 2005; Garibaldi et al. 2018). At smaller scales, farmers may manage the areas around cultivation plots seeking to primarily increase coffee edge diversity, but also proximity to forest edge and forest-coffee edge density. However, at broader-scales, landscape management actions should focus on reaching the amount of coverage needed to avoid disservices and stimulate service provision (González-Chaves et al. 2022). Such management actions may benefit from legislation and incentive programs on native forest restoration that combine both local and regional targets (d’Albertas et al. 2023). In Brazil, this could be achieved through the Brazilian environmental legislation which obliges landowners to maintain a determined amount of native vegetation within their properties (Metzger et al. 2019), and by economic incentives like payment for ecosystem services focused on promoting or ensuring long-term service provision through native vegetation protection or regeneration (Ruggiero et al. 2019). Such environmental schemes may foster service provision by promoting native vegetation conservation while still allowing landowners to better manage the arrangement of native vegetation patches and crop plantations to optimise flow-related landscape attributes. Strategies like these could balance nature conservation enforcement while maintaining enough farmers’ agency to accommodate their needs and perspectives regarding on-farm management (e.g., power to decide how to arrange productive and forested areas within-farm), two essential aspects when triggering in-field change (Hatt et al. 2018).
Conclusion
We present novel evidence on how landscape structural attributes linked to the supply, demand and flow of ecosystem services shape the provision of pollination and pest control in coffee plantations across multiple scales. Combined, our novel results on the (i) multiple landscape structural effects on pollination and pest control across different scales and also within pest control; (ii) consistent service provision of pollination in contrast to service and disservice provision of pest control; (iii) negative effects of high coffee cover not only to pollination but also to pest control; (iv) forest-coffee edge density effects on pest control service and disservice provision; and (v) common and local positive effects of high coffee edge diversity on pollination and pest control, showcase the challenges behind planning and managing landscapes for multiple service provisions. This is relevant to the current literature not only to highlight the difficulty in enhancing both pollination and pest control while avoiding trade-offs but also to stress the need to consider landscape structural effects on services and disservices both locally and regionally. We then suggest combining local and landscape management strategies to protect and restore native vegetation at multiple scales whilst allowing for different crop and habitat configurations to benefit multiple service provisions while avoiding disservices.
References
Aguilera G, Roslin T, Miller K, Tamburini G, Birkhofer K, Caballero-Lopez B, Lindström SA, Öckinger E, Rundlöf M, Rusch A, Smith HG (2020) Crop diversity benefits carabid and pollinator communities in landscapes with semi-natural habitats. J Appl Ecol 57(11):2170–2179
Alignier A, Solé-Senan XO, Robleño I, Baraibar B, Fahrig L, Giralt D, Gross N, Martin JL, Recasens J, Sirami C, Siriwardena G (2020) Configurational crop heterogeneity increases within-field plant diversity. J Appl Ecol 57(4):654–663
Aristizábal N, Metzger JP (2018) Landscape structure regulates pest control provided by ants in sun coffee farms. J Appl Ecol 56(1):21–30
Baensch S, Tscharntke T, Ratnieks FL, Haertel S, Westphal C (2020) Foraging of honey bees in agricultural landscapes with changing patterns of flower resources. Agric Ecosyst Environ 291:106792
Barton K, Barton MK (2015) Package ‘mumin.’ Version 1(18):439
Bates D, Maechler M, Bolker B, Walker S, Christensen RHB, Singmann H, Dai B, Grothendieck G, Eigen C, Rcpp L (2015) Package ‘lme4.’ Convergence 12:1–7
Bichara CC, Santos GMD, dos Santos AB, Santana-Reis VP, da Cruz JD, Gobbi N (2010) Foraging behavior of the swarm-founding Wasp Polybia (Trichothorax) sericea (Hymenoptera, Vespidae): daily resource collection activity and flight capacity. Sociobiology 55(3):899–907
Boesing AL, Nichols E, Metzger JP (2018) Biodiversity extinction thresholds are modulated by matrix type. Ecography 41(9):1520–1533
Boesing AL, Nichols E, Metzger JP (2018) Land use type, forest cover and forest edges modulate avian cross-habitat spillover. J Appl Ecol 55:1252–1264
Bolker BM, Gardner B, Maunder M, Berg CW, Brooks M, Comita L, Crone E, Cubaynes S, Davies T, de Valpine P, Ford J (2013) Strategies for fitting nonlinear ecological models in R, AD model builder, and BUGS. Methods Ecol Evol 4(6):501–512
Boscolo D, Metzger JP (2009) Is bird incidence in Atlantic forest fragments influenced by landscape patterns at multiple scales? Landsc Ecol 24:907–918
Brown JS (1999) Vigilance, patch use and habitat selection: foraging under predation risk. Evol Ecol Res 1(1):49–71
Carlucci MB, Marcilio-Silva V, Torezan JM (2021) The southern Atlantic Forest: use, degradation, and perspectives for conservation. In The Atlantic forest. Springer, Cham, pp 91–111
Chain-Guadarrama A, Martínez-Salinas A, Aristizábal N, Ricketts TH (2019) Ecosystem services by birds and bees to coffee in a changing climate: a review of coffee berry borer control and pollination. Agric Ecosyst Environ 280:53–67
d’Albertas F, Sparovek G, Pinto LFG, Hohlenwerger C, Metzger JP (2023) Yield increases mediated by pollination and carbon payments can offset restoration costs in coffee landscapes. One Earth 7(1):110–22
Dainese M, Martin EA, Aizen MA, Albrecht M, Bartomeus I, Bommarco R, Carvalheiro LG, Chaplin-Kramer R, Gagic V, Garibaldi LA, Ghazoul J (2019) A global synthesis reveals biodiversity-mediated benefits for crop production. Sci Adv 5(10):eaax0121
Dias FP, Souza CAS (2015) Caracterização de progênies do cafeeiro (Coffea arabica L.) selecionadas em Minas Gerais: II-caracteres relacionados à produção. Ceres 52:299
FAO (2019) Top 20 Commodities, Export Value by country. http://www.fao.org/faostat/en/#rankings/major_commodities_exports
Fox J, Friendly GG, Graves S, Heiberger R, Monette G, Nilsson H, Ripley B, Weisberg S, Fox MJ, Suggests MA (2007) The car package. R foundation for statistical computing, p 1109
Fu B, Wang S, Su C, Forsius M (2013) Linking ecosystem processes and ecosystem services. Curr Opin Environ Sustain 5(1):4–10
Gagic V, Marcora A, Howie L (2019) Additive and interactive effects of pollination and biological pest control on crop yield. J Appl Ecol 56(11):2528–2535
Gámez-Virués S, Perović DJ, Gossner MM, Börschig C, Blüthgen N, De Jong H, Simons NK, Klein AM, Krauss J, Maier G, Scherber C (2015) Landscape simplification filters species traits and drives biotic homogenization. Nat Commun 6(1):1–8
Garibaldi LA, Andersson GK, Requier F, Fijen TP, Hipólito J, Kleijn D, Pérez-Méndez N, Rollin O (2018) Complementarity and synergisms among ecosystem services supporting crop yield. Glob Food Secur 17:38–47
González-Chaves A, Jaffé R, Metzgerde Kleinert JPA (2020) Forest proximity rather than local forest cover affects bee diversity and coffee pollination services. Landsc Ecol 35(8):1841–1855
González-Chaves A, Carvalheiro LG, Garibaldi LA, Metzger JP (2022) Positive forest cover effects on coffee yields are consistent across regions. J Appl Ecol 59(1):330–341
Hatt S, Boeraeve F, Artru S, Dufrêne M, Francis F (2018) Spatial diversification of agroecosystems to enhance biological control and other regulating services: an agroecological perspective. Sci Total Env 621:600–611
Harrison XA (2015) A comparison of observation-level randomeffect and beta-binomial models for modelling overdispersion in binomial data in ecology & evolution. PeerJ 3:e1114
Hohlenwerger C, Tambosi LR, Metzger JP (2022) Forest cover and proximity to forest affect predation by natural enemies in pasture and coffee plantations differently. Agric Ecosyst Environ 333:107958
Huais PY (2018) multifit: an R function for multi-scale analysis in landscape ecology. Landsc Ecol 33(7):1023–1028
Jackson HB, Fahrig L (2015) Are ecologists conducting research at the optimal scale? Glob Ecol Biogeogr 24(1):52–63
Karp DS, Daily GC (2014) Cascading effects of insectivorous birds and bats in tropical coffee plantations. Ecology 95(4):1065–1074
Kremen C, Williams NM, Bugg RL, Fay JP, Thorp RW (2004) The area requirements of an ecosystem service: crop pollination by native bee communities in California. Ecol Lett 7(11):1109–1119
Le Provost G, Schenk NV, Penone C, Thiele J, Westphal C, Allan E, Ayasse M, Blüthgen N, Boeddinghaus RS, Boesing AL, Bolliger R (2023) The supply of multiple ecosystem services requires biodiversity across spatial scales. Nat Ecol Evol 7:236–249
Librán-Embid F, De Coster G, Metzger JP (2017) Effects of bird and bat exclusion on coffee pest control at multiple spatial scales. Landsc Ecol 32(9):1907–1920
Librán-Embid F, Grass I, Emer C, Ganuza C, Tscharntke T (2021) A plant–pollinator metanetwork along a habitat fragmentation gradient. Ecol Lett 24(12):2700–2712
Magnusson A, Skaug H, Nielsen A, Berg C, Kristensen K, Maechler M, van Bentham K, Bolker B, Brooks M, Brooks MM (2017) Package ‘glmmTMB’. R Package Version 0.2. 0
Martensen AC, Ribeiro MC, Banks-leite C, Prado PI, Metzger JP (2012) Associations of forest cover, fragment area, and connectivity with neotropical understory bird species richness and abundance. Conserv Biol 26:1100–1111
Martin AE (2018) The spatial scale of a species’ response to the landscape context depends on which biological response you measure. Curr Landsc Ecol Rep 3(1):23–33
Martin EA, Reineking B, Seo B, Steffan-Dewenter I (2013) Natural enemy interactions constrain pest control in complex agricultural landscapes. Proc Natl Acad Sci 110:5534–5539
Martínez-Salinas A, Chain-Guadarrama A, Aristizábal N, Vilchez-Mendoza S, Cerda R, Ricketts TH (2022) Interacting pest control and pollination services in coffee systems. Proc Natl Acad Sci 119(15):e2119959119
Medeiros HR, Martello F, Almeida EA, Mengual X, Harper KA, Grandinete YC, Metzger JP, Righi CA, Ribeiro MC (2019) Landscape structure shapes the diversity of beneficial insects in coffee producing landscapes. Biol Conserv 238:10819
Metzger JP, Bustamante MM, Ferreira J, Fernandes GW, Librán-Embid F, Pillar VD, Prist PR, Rodrigues RR, Vieira IC, Overbeck GE (2019) Why Brazil needs its legal reserves. Perspect Ecol Conserv 17(3):91–103
Metzger JP, Villarreal-Rosas J, Suárez-Castro AF, López-Cubillos S, González-Chaves A, Runting RK, Hohlenwerger C, Rhodes JR (2021) Considering landscape-level processes in ecosystem service assessments. Sci Total Environ 796:149028
Morris JR, Perfecto I (2016) Testing the potential for ant predation of immature coffee berry borer (Hypothenemus hampei) life stages. Agric Ecosyst Environ 233:224–228
Pierre LSR (2011) Níveis populacionais de Leucoptera coffeella (Lepidoptera: Lyonetiidae) e Hypothenemus hampei (Coleoptera: Scolytidae) e a ocorrência de seus parasitoides em sistemas de produção de café orgânico e convencional. PhD, Universidade de São Paulo. http://www.sbicafe.ufv.br:80/handle/123456789/9037
Qiu J, Queiroz C, Bennett EM, Cord AF, Crouzat E, Lavorel S, Maes J, Meacham M, Norström AV, Peterson GD, Seppelt R (2021) Land-use intensity mediates ecosystem service tradeoffs across regional social-ecological systems. Ecosyst People 17(1):264–278
R Development Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ruggiero PG, Metzger JP, Tambosi LR, Nichols E (2019) Payment for ecosystem services programs in the Brazilian Atlantic Forest: effective but not enough. Land Use policy 82:283–291
Saturni FT, Jaffe R, Metzger JP (2016) Landscape structure influences bee community and coffee pollination at different spatial scales. Agric Ecosyst Environ 235:1–12
Spake R, Bellamy C, Graham LJ, Watts K, Wilson T, Norton LR, Wood CM, Schmucki R, Bullock JM, Eigenbrod F (2019) An analytical framework for spatially targeted management of natural capital. Nat Sustain 2(2):90–97
Spake R, Bowler DE, Callaghan CT, Blowes SA, Doncaster CP, Antao LH, Nakagawa S, McElreath R, Chase JM (2023) Understanding ‘it depends’ in ecology: a guide to hypothesising, visualising and interpreting statistical interactions. Biol Rev. https://doi.org/10.1111/brv.12939
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C (2005) Landscape perspectives on agricultural intensification and biodiversity–ecosystem service management. Ecol Lett 8(8):857–874
Van Der Vossen HAM (1985) Coffee selection and breeding. In: Clifford MN, Wilson KC (eds) Coffee: botany, biochemistry and production of beans and beverage. Croom Herm, Westport, London, pp 48–96
Vanbergen AJ (2013) Threats to an ecosystem service: pressures on pollinators. Front Ecol Environ 11:251–259
Varah A, Jones H, Smith J, Potts SG (2020) Temperate agroforestry systems provide greater pollination service than monoculture. Agric Ecosyst Environ 301:107031
Villard MA, Metzger JP (2014) Beyond the fragmentation debate: a conceptual model to predict when habitat configuration really matters. J Appl Ecol 51(2):309–318
Way MJ, Khoo KC (1992) Role of ants in pest management. Annu Rev Entomol 37(1):479–503
Winqvist C, Ahnström J, Bengtsson J (2012) Effects of organic farming on biodiversity and ecosystem services: taking landscape complexity into account. Ann N Y Acad Sci 1249(1):191–203
Zuur A, Ieno E, Walker N, Saveliev A, Smith G (2009) Mixed-effects models and extensions in ecology with R. Springer, New York
Acknowledgements
We thank LEPAC’s great team for support with spatial analysis and Melina Leite, Danilo Muniz and Gabriel Garcia for support with statistical analysis. We also thank Elizabeth Nichols, Maria Uriarte, and Bruno Travassos-Britto for important discussions and valuable suggestions on earlier versions of the manuscript. Finally, we thank the University of Southampton for hosting C.H. during her PhD and the Student Conference on Conservation Science-Cambridge (SCCS) internships.
Funding
This work was supported by the São Paulo Research Foundation as part of the “Interface Project” (FAPESP; 2013/23457-6); the Brazilian Ministry of Education (CAPES; CAPES-DS 2014-2016 A.GC. and N.A.; F.S.; L.R.T. and C.H. Processes No. 001; 88882.327885/2019-01; 88887.309513/2018-00); the Brazilian National Council for Scientific and Technological Development (CNPQ; J.P.M. Process No. 305484/2017-6) and the Miriam Rothschild Travel Bursary Programme (known as the SCCS Internships) of the University of Cambridge who funded C.H.’s internship to the University of Southampton. R.S. and F.E. were funded by the ERC Starting Grant “SCALEFORES” (Grant ID 680176) awarded to F.E. FLE. thanks the PAEC-OEA-CGUB program (master fellowship).
Author information
Authors and Affiliations
Contributions
C.H., R.S., L.R.T., F.E. and J.P.M. contributed to the study conception and analytical design. Field study design and data collection were performed by N.A., A.GC., F.LE. and F.S., and geo data collection was performed by C.H. C.H. led final analysis and material preparation for the methodological design with contributions from N.A., A.GC., F.LE., F.S and R.S. The first draft of the manuscript was written by C.H. and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hohlenwerger, C., Spake, R., Tambosi, L.R. et al. Coffee pollination and pest control are affected by edge diversity at local scales but multiscalar approaches and disservices can not be ignored. Landsc Ecol 39, 75 (2024). https://doi.org/10.1007/s10980-024-01869-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10980-024-01869-1