
Abstract
The Water Framework Directive (WFD) requires actions from European Union Member States to reduce external nutrient loading in lakes of moderate/poor/bad ecological status and restore their good ecological status. Several poor-quality Danish lakes have been exempted from WFD criteria (and more proposed), due to suspected major imported nutrient contributions from waterbirds, without supporting evidence. Here, we estimated relative nutrient contributions from increasing numbers of roosting waterbirds compared to total nutrient loadings in four brackish lakes (43–491 ha) at an internationally important Ramsar site/EU Birds Directive Special Protection Area. Combining night- and daytime counts of roosting numbers of the most numerous species with intake and faecal output models, we estimated roosting birds (mostly geese) contributed 0.2–0.4% of all N inputs (0.3–1.2 kg N/ha/year) and 0.6–2.0% of all P inputs (0.04–0.12 kg P/ha/year) in three lakes (one of which has been proposed exempted from WFD criteria), but potentially as high as 14% and 76%, respectively, at a fourth clearwater lake without direct inflow or outlets. These results confirm the need for case-by-case assessments of avian nutrient import relative to total nutrient balance before exempting lakes from WFD conditions due to supposed natural loading from waterbirds.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Just 0.01% of the Earth’s water is fresh (< 500 ppm dissolved salts, Gleick, 1996), constituting up to 9.1% of global surface area (lakes, reservoirs, rivers and freshwater wetlands, excluding ice cover, Lehner & Döll, 2004). Freshwater ecosystems are among the most diverse globally per unit habitat volume, supporting 140,000 species (12% of all described, Reid et al., 2019; Albert et al., 2021) confirming the need for effective conservation of freshwaters to protect this valued resource, their ecological functions and the ecosystem services they provide. Despite the acknowledged and evident biodiversity crisis in visible terrestrial ecosystems, indices for freshwater organisms showed an 84% decline during 1970–2016 (Almond et al., 2020) compared to the indices for marine (36%) or terrestrial populations (38%, during 1970–2012, Collen et al., 2009; Tickner et al., 2020). Aquatic ecosystem eutrophication is recognised as one of the most widespread human cultural forces impacting global freshwater biological diversity (Harper, 1992; Carpenter, 2005; Cook et al., 2018), with anthropogenic nitrogen (N) and phosphorus (P) contributions to freshwater systems from agriculture (e.g., Mateo-Sagasta et al., 2017) and urban wastewater discharge from households and industry (Bouwman et al., 2005) adversely affecting aquatic diversity (e.g., Jeppesen et al., 2000). While P is conventionally thought to limit production in freshwaters, enhanced levels of N loadings in freshwater systems also reduces plant biodiversity and ultimately affects ecosystem species diversity (Moss et al., 2013).
In recent decades, increasing N loadings from atmospheric pollution (e.g., Bergström & Jansson, 2006) and N and P from nutrient runoff from fertiliser and human wastewater sources (e.g., Duan et al., 2012; Han & Allan, 2012) to many western European wetlands have demonstrably adversely affected aquatic biological diversity, including nesting waterbirds (Fox et al., 2019, 2020). Recent studies also suggest N may be introduced to coastal areas from buried tunnel valleys (Sandersen et al., 2023), which is known to be a source of P in at least one studied Danish lake (P.B.E. Sandersen, pers. comm., unpubl. data). Roosting bird flocks have been demonstrated to add nutrients to wetlands (e.g., Hahn et al., 2007, 2008; Dessborn et al., 2016), especially largely carnivorous gull species that forage out on surrounding habitats by day, aggregating in large numbers on water bodies at night, depositing large amounts of imported N and P in their faeces (Moss, 2017; Winton & River, 2017; Martín-Vélez et al., 2019). Large concentrations of resting herbivorous geese have also been shown to contribute up to 40% total N and 75% total P to small shallow pond systems (Kitchell et al., 1999) and herbivorous ducks, geese and swans are considered to be responsible for “most” nutrient transfer between land and freshwater systems according to Moss (2017). In this context, it is important here to distinguish between those waterbirds that remain on wetland systems throughout the diurnal cycle (such as coots and many dabbling ducks) which recycle nutrients on wetlands and those that feed out from wetlands by either day or night, but which return to aggregate for safety from predators by night or day on these wetlands, especially waterbodies, where the faeces deposited whilst present represent a net import of nutrients.
In intensively cultivated areas like Denmark (and throughout Northern Europe), many lakes have now been severely affected by N and P, resulting in highly eutrophicated waters. To combat this trend, the EU adopted the Water Framework Directive (WFD; 2000/60/EC) in 2000. The WFD classifies lakes according to ecological quality, related to nutrient concentrations and thereby the external loading of nutrients (Poikane et al., 2019). Waterbodies should be classified in five categories, i.e., “high” (best), “good”, “moderate”, “poor” and “bad” (worst), and the Directive sets the current goal of achieving at least “good” quality in all surface waters by 2027, unless they are considered to be heavily modified waters or their moderate/poor/bad status arises from some other natural explanation for not achieving “good” chemical and ecological status. In Denmark, as of 2021, only 20% of 768 lakes with known ecological quality were found to be in “high” or “good” ecological status (Miljøministeriet, 2023a). At the other 80%, managers are required to take action in the catchments and/or in the lakes elsewhere to raise their quality to achieve at least “good” ecological status.
Managing such catchments, lakes and wetlands to restore their ecological status, protect and maintain their biological diversity often involves resolving conflicts between multiple stakeholders, users and special interest groups such as farmers, anglers and conservationists, who place different values on different services provided by such ecosystems (Poikane et al., 2024). Even within areas dedicated to nature conservation protection, management interventions may benefit one species, or group of species or habitats, over others to fulfil the specific conservation objectives for the site. For example, many lakes qualify as internationally important for their nocturnally roosting geese and swans, because they offer hunting-free environments, relatively safe from predation, where these large-bodied birds gather in large numbers to sleep before dispersing elsewhere to feed during daylight (e.g., Brotherston, 1964). These species increasingly exploit agricultural grasslands and crops, which offer enhanced feeding profitability compared to the natural and semi-natural habitats used before the advent of modern intensive agriculture (Fox & Abraham, 2017; Fox et al., 2017). Many such species in Europe and North America are showing exponential population growth in recent years, exacerbating conflict with farmers, financially affected by crop damage caused by such birds (Fox & Madsen, 2017). Numbers of pink-footed Anser brachyrhynchus Baillon and greylag geese A. anser (Linnaeus) wintering in NW Europe have increased to the point where they are the subject of international species adaptive harvest management plans to limit their number within agreed population target levels (Madsen et al., 2017; Powolny et al., 2018). Barnacle geese Branta leucopsis (Bechstein) wintering in continental western Europe have increased from 20,000 birds in the 1950s to over 1.2 million individuals in 2015, simultaneously expanding their breeding and wintering ranges (Fox & Madsen, 2017). Such expansions in range and abundance of geese (Heldbjerg et al., 2024) have also resulted in increasingly large numbers aggregating at favoured roosting places, where their contributions of faecal material, accumulated during daytime feeding in agricultural habitats but to some degree deposited into their aquatic nocturnal roosts sites after flying there, add to nutrient loads contributed by precipitation, nutrient runoff, groundwater ingress and other biological and geochemical processes. In Denmark the same is the case for the numerous whooper swan Cygnus cygnus Linnaeus, which also have switched to feed on agriculture and roost on wetlands (Laubek et al., 2019). Geese and swans possess relatively inefficient digestive systems, relying on rapid transition of large amounts of grazed plant material through relatively simple alimentary canals to extract soluble carbohydrates and proteins from cell contents. This produces 130–150 faecal pellets/day in the case of barnacle geese, of which up to 25 are ejected on the roost site as the gut becomes emptied after ceasing to feed (Ebbinge et al., 1975). Up to two-thirds of all nitrogen in goose faeces is in soluble form, especially as ammonium ions and uric acid metabolised excretion products (Cargill, 1981; Fox & Kahlert, 1999). The situation is less clear for phosphorus, with some authors suggesting phosphorus salts in bird drop**s are only sparingly soluble in water as orthophosphates (e.g., Ligeza & Smal, 2003), so much organic phosphorus is deposited into sediments, while others suggest higher levels of availability (e.g., Powell et al., 1989). Individual captive gulls produced between 608 and 1,819 mg total nitrogen and 3 to 115 mg total phosphorus daily according to species, size and diet (Gould & Fletcher, 1978). Hence, deposition of such organic material harvested elsewhere, but imported by geese and other waterbirds into wetlands when they roost there by day or night, has at least the potential to contribute additionally to the nitrogen and phosphorus budgets of wetlands from this source (see above).
So, the question is: how much do diurnal and nocturnal roosting waterbirds currently contribute to waterbodies in one of Denmark’s most important wetland complexes after a period of major goose population increase, and can such results be considered representative and upscaled to a larger set of lakes?
In Denmark, 24 lakes were exempted from fulfilling the good/high water quality demands established by the WFD in the 2nd generation 2015–2021 River Basin Management Plans (RBMPs; Clausen & Bregnballe, 2017) and an additional 37 lakes have been suggested to be exempted prior to the 3rd generation 2021–2027 RBMPs [Danish Environmental Protection Agency (EPA), in litt.], including one of the lakes we study here. Exemptions to WFD requirements are made based on the assumption that naturally occurring waterbird populations provide a major source of nutrients to the lakes, which contribute significantly to nutrient loadings. In this way, without removing the influence of the naturally occurring avian populations, it is not possible for managers to make interventions or actions within the catchment or the lake to change the water quality from, for example, “moderate” to “good”. To date, Danish exemptions have been based on simple abundance estimates for great cormorant Phalacrocorax carbo (Linnaeus) at five lakes, on missing data relating to geese and duck abundance and poor data relating to foraging Eurasian coot Fulica atra Linnaeus and swans Cygnus spp. (unpubl. data, Danish EPA 2021). In all cases, it is not clear how much N and P are physically imported into these waterbodies by these birds and to what degree they feed within each site (i.e., merely recycling nutrients within the waterbody, rather than physically importing extra nutrients harvested from elsewhere). While initial studies indicate that in some nutrient-sensitive Danish lakes, waterbirds can have a significant impact on water quality (Pedersen et al., 2016; Jensen et al., 2019), there remains no direct evidence for this being the case as a basis for the exemption of the 59 lakes. There is therefore a clear need for specific case studies to examine the extent of such nutrient contributions from waterbirds to justify WFD exemptions where waterbirds may genuinely have a contributory effect.
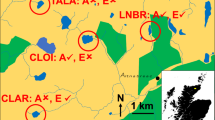
Vejlerne in northern Jutland (57° 04′ N, 9° 00′ E) is one of Denmark’s most important wetland reserves, designated as a Ramsar Site of international importance and a European Union Special Protection Area (SPA), protected for its internationally important concentrations of breeding (Lauridsen et al., 2021), as well as staging and wintering waterbirds (Clausen et al., 2019), owned and managed by a private fund. In this study, we concentrate upon the effects of waterbird-imported faecal deposition on four shallow slightly brackish lakes (Lund Fjord, Han Vejle, Selbjerg Vejle, and Glombak) and an associated major area of low-lying saltmarsh meadows (Bygholmengen), seasonally inundated by brackish water in the eastern section of the Vejler Reserve, the Eastern Vejler (Fig. 1). Three of these four lakes have shown signs of increasing eutrophication in recent years which increasingly conflict with the nature conservation objectives set for the area; Glombak, Selbjerg Vejle and Lund Fjord in particular have witnessed declines in Secchi disc depth measurements, associated with increases in total N and P (see Table 1, but summarized by He et al., 2020 and Lauridsen et al., 2021). For this reason, the owners initiated a study of the potential causes of the observed eutrophication processes. One of the lakes, Lund Fjord, that has been the most eutrophic of the lakes for decades, is indeed one of the newly proposed lakes to be exempted from the WFD.

Main map showing the geographical arrangement of the study area (Eastern Vejler) and site names (in white), while the inset map gives the location of Vejlerne within Denmark. The study area is divided into six compartments by dikes (red), and the main hydrological system in three compartments, each of which drain into Limfjorden via three independent canals and gates in the sluice Centralslusen (in turquoise), with arrows indicating the main direction of flow for water. Lund Fjord and Han Vejle drain into the Lund Fjord Kanal, which ultimately flows out via the eastern gate of Centralslusen at the southern end. There is some seepage to Selbjerg Vejle from Bygholm Vejle Nord, which itself discharges into Glombak and ultimately via Glombak Kanal through the western gate of Centralslusen. Run-off from Bygholmengen drains via the Central Kanal through the central gate of Centralslusen
In this study, we aimed to explore the degree to which these four shallow brackish lakes could have been affected by numbers of geese, as well as all the other staging and wintering waterbirds, using the site in recent years and to what extent these birds could have contributed to the varying degrees of eutrophication observed. To do so, we combine data on contemporary numbers of waterbirds using the site with estimates of the quantity and estimated N and P content of their faeces ejected into their roosting places. We also combine this information with the results of bio-tracking studies of individual pink-footed geese using the site as a roosting area, from where they forage out onto surrounding farmland to feed at night to verify (i) the precise roosting areas used by the geese and (ii) that they remain at the same roosting site throughout the night. Finally, we use recent historical N and P balance estimations to compare current inputs from avian faeces of roosting birds to see to what degree these make a significant contribution to the overall budgets of N and P in these systems.
Methods
Study area
The Eastern Vejler were formerly an inlet of the Limfjord that were embanked and drained for agriculture in the 1800s, but which were abandoned as unprofitable for farming in 1912. Since then, they have developed into one of Denmark’s pre-eminent breeding, staging and wintering waterbird resorts. The site supports large numbers of breeding greylag geese and marsh harriers Circus aeruginosus (Linnaeus), many rare nesting species such as Eurasian bittern Botaurus stellaris (Linnaeus) and black terns Chlidonias niger (Linnaeus) (Kjeldsen, 2008), and the meadows of Bygholmengen are also of national importance for their breeding wader species (Thorup, 2004). Additionally the site is regularly hosting between 40,000 and 80,000 waterbirds, with highest numbers being present in autumn and spring (Clausen et al., 2019). One of the numerically important groups are geese, for which this site is an important roost for staging and wintering greylag, pink-footed and barnacle geese that forage on farmland around the site, but which congregate on Vejlerne at night to roost. In recent years, the numbers of four species have increased dramatically, i.e., the three most numerous goose species introduced above, but also greater white-fronted geese Anser albifrons (Scopoli); only the least numerous taiga bean goose Anser fabalis (Latham) has shown declining numbers in recent years (see Fig. 2). Hence it is natural to ask if their presence in increasing numbers at the site is affecting the nutrient balance of the different lakes on site?

Long-term monitoring of nutrient concentrations and Secchi depth suggested that most of the Eastern Vejler lakes have undergone eutrophication (to varying degrees) since 2000 (Table 1, He et al., 2020). Lund Fjord has been characterised as heavily nutrient enriched, supporting dense algal growth back to the 1970s (Møller, 1980), and has become more turbid and eutrophic throughout the past two decades, with high nutrient levels and low Secchi depth (He et al., 2020). The nutrient conditions in Selbjerg Vejle were already affected by agricultural runoff in the early 1970s (Kortegaard 1974), but Glombak was always generally less affected by such nutrient sources (Degener 1994). Secchi depth decreased from > 0.9 m in 2000 to c. 0.3 m in 2017–2018 in Selbjerg Vejle and Glombak and although Han Vejle has remained macrophyte-dominated and clear since 2000, total nitrogen (TN) and phosphate (TP) concentrations have more than doubled, indicating eutrophication (Table 1, He et al., 2020). The eutrophic status of the three larger lakes are discernibly visible as a greenish–yellowish colour, compared to the more dark colour of Han Vejle (Fig. 1). The three large lakes are classified as being in “bad” or “poor” ecological status in the latest baseline assessments prior to the 3rd generation RBMPs, and although Han Vejle shows less signs of eutrophication, this is classified as “moderate” (Miljøministeriet, 2023b)—hence none of our study lakes fulfil the goal of at least “good” ecological status.
It is important at the outset to understand the complex hydrology of the Eastern Vejler system, which is summarised in Fig. 1, schematically illustrating drainage of the entire area into Limfjorden via a three-gate shallow sluice system (Centralslusen) located in the road dam, which forms the southern boundary of the area. The sluice gates can only open when the water level in Limfjorden is lower than in Vejlerne. Hence longer periods with westerly winds cause surges and elevated water levels in Limfjorden that will prevent opening of the sluice gates. Runoff from the eastern catchment area, via the Lund Fjord Kanal is far greater than that from the western hinterland via Glombak Kanal (Cowiconsult, 2000). Lund Fjord, Han Vejle and runoff from a 4,190 ha primarily agricultural catchment to the east and north all drain via Lund Fjord Kanal into its outlet to Limfjorden at the eastern gate of Centralslusen (Fig. 1). During periods with high water levels in Limfjorden water flows from Lund Fjord Kanal back into Lund Fjord, thereby flushing a subsidy of nutrients from agricultural areas east of Vejlerne into that waterbody. Since the establishment of a dam in the drain from Han Vejle in 1994, backflow from Lund Fjord Kanal has only affected Han Vejle at very high water levels. Selbjerg Vejle and Bygholm Vejle Nord drain out via Krap Kanal through Glombak, water from which flows out via Glombak Kanal in the western gate of Centralslusen (Fig. 1). Here, too, during high water levels, runoff can accumulate from the associated likewise mostly agricultural catchment area, although this contribution is much less than in Lund Fjord/Lund Fjord Kanal complex. The south central part of the system (Bygholmengen) is an area of inundated brackish meadows, ephemeral and permanent pools, largely hydrologically isolated from the rest of the wetland and runoff from here is isolated from the other elements, flowing out independently into Limfjord through the middle gate of Centralslusen (Fig. 1).
Changes in goose abundance at Vejlerne
During 1978–2020, at least ten land-based total mid-monthly counts of all wetland bird species, including geese, were carried out in January–May and August–December on the Eastern Vejler, with additional counts in June and July, following standardised protocols from fixed positions overlooking all wetlands to ensure comparability with the long time series (see Nielsen & Clausen, 2023). Additional counts were also carried out in October 2018–September 2019 to assess the true numbers of overnight roosting waterbirds using the site as overnight roosts (see below).
Estimating bird use of the different areas
Several species of geese, swans, gulls, cranes and diving ducks fly into these sites at Vejlerne to roost in safety overnight (or by daylight) from daytime (or nighttime) feeding areas in the vicinity, where they deposit faecal material imported from food derived in the surrounding landscape. They therefore make a net contribution to the nitrogen and phosphorus budgets of the lakes in a way that waterbirds using the site throughout the diurnal cycle (i.e., not feeding elsewhere) do not. To assess the numbers of waterbirds utilising these areas as a night/day-time roost in this way throughout the annual cycle, we undertook two- or three-monthly counts of geese, swans, gulls, cranes and diving ducks flying in or out at dusk and dawn from the waterbodies of Glombak, Selbjerg Vejle, Han Vejle, Lund Fjord, Bygholm Vejle Nord (and other associated lakes), and into the meadows of Bygholmengen from October 2018 until September 2019. Counts were performed either for 2 h in the morning (from 1 h before to 1 h after sunrise) or 2 h in the evening (also 1 h on each side of sunset). All outgoing (mostly morning) or incoming (mostly evening flight) flocks of swans, geese, common cranes Grus grus (Linnaeus), goldeneye Bucephala clangula (Linnaeus), red-breasted merganser Mergus serrator Linnaeus and gulls were determined to species, counted and their overnight location identified. In addition to these species that rest overnight, numbers of day-roosting tufted ducks Aythya fuligula (Linnaeus) and common pochard A. ferina (Linnaeus) were also counted on the lakes (as day-resting birds detected in daytime counts undertaken during the same period from October 2018 to September 2019 at similar times to the roost counts). These two species fly out at night to feed elsewhere. Based on these counts, we calculated the numbers of bird-days on each water body per month as the average monthly count number multiplied by the number of days in the month and summed these up to generate the numbers of birds making overnight stays per year.
Confirming that overnight roosting geese remain at their resting place all night
In total, 56 pink-footed geese (5 males and 51 females), originating from the population breeding in Svalbard and wintering/staging in Norway, western Denmark, The Netherlands and Belgium, were equipped with solar-powered GPS-GSM transmitter neckbands, type OrniTrack-N38 (Ornitela UAB, Lithuania; weight 38 g). In the Svalbard breeding areas, geese were caught in moulting family groups in a corral at two sites in Isfjorden in western Svalbard, 30 July 2018 (individuals fitted with transmitters n = 16) and 1 August 2018 (n = 19). On a spring staging site in Tyrnävä, Oulu, Finland, geese were caught by cannon-nets on 28 April 2018 (n = 10), on 27 April 2019 (n = 8) and on 1 May 2019 (n = 3; see Schreven et al., 2021 for further details). During periods with well charged battery levels, devices gave positions every 10 min, with 74% within 20 m accuracy (Schreven et al., 2021). We here primarily consider the fixes of tagged geese that fell within the confines of the boundaries of the Eastern Vejler EU Special Protection Area (national designation code No. 13), which includes all the lakes covered in the study, as well as the meadows of Bygholmengen, although for completeness and comparison, we also present goose positions within Western Vejler EU Special Protection Area (No. 20) and those foraging out on the farmland of the surrounding area (see tracking results below). For the purposes of these analyses, to filter night-time positions from those during daylight, we here defined daylight as the period from 1 h before local sunrise to 1 h after local sunset, generated using the resources at https://gml.noaa.gov/grad/solcalc/calcdetails.html.
Estimating contributions of P and N from bird use of the different areas
We used the methods of Hahn et al. (2007, 2008) to estimate the species-specific daily contribution of carnivorous (mostly gulls and diving ducks) and herbivorous waterbirds (predominantly geese and swans) to nutrient loadings in each of the waterbodies at Vejlerne. These authors used alternative physiological models to estimate the contributions of N and P from carnivorous and herbivorous birds based on food-intake and excreta-production approaches. We corrected for elevated levels of P and N in faeces when herbivorous birds fed on green plant parts which are harder to fully digest than when feeding on grains and beet/potato residues (Hahn et al., 2008). For the herbivorous goose and swan species and following Hahn et al. (2008), we generated maximum and minimum estimates for N and P deposition for each individual for each species based on the results from an (i) food-intake and (ii) drop** output model, respectively, in relation to species body mass (Fig. 3). We then used the mean value from the two models as average daily contributions, and the two model values as maximum and minimum daily contributions, respectively, where the food-intake model generally gives the highest values for herbivores, but the drop** model the highest for carnivores (Fig. 3). In the absence of literature values for common cranes and knowing that these birds are mainly herbivorous feeders in adjacent agricultural areas when their numbers are highest, we estimated their contribution based on a body mass of 4,798 g (Cramp & Simmons, 1980) and N and P contributions according to the fitted curves for herbivores in Hahn et al. (2008)—redrawn in Fig. 3. For day-resting diving ducks (classified here as carnivores), we estimated the daily contributions from tufted duck as 380 mg N and 60 mg P, respectively, based on Sørensen (1997). For the other duck species we estimated their contribution in relation to their body mass (carnivores, for tufted duck 896 g, common pochard 976 g, goldeneye 963 g and red-breasted merganser 1,091 g, based on average body mass of males and females, and on data from Denmark in Cramp & Simmons 1977). In this way, we assumed that, for instance, common pochard deposited 976/896 = 1.08 times more N and P than tufted duck. For roosting flocks of gulls (carnivores) we used daily estimates of N and P loading by non-breeding birds from Hahn et al. (2007), also redrawn in Fig. 3. In situations where mixed flocks of evening roosting gulls could not be identified to species [typically composed of black-headed Chroicocephalus ridibundus (Linnaeus), common Larus canus Linnaeus and herring gulls L. argentatus Pontoppidan] we used an average body mass for the three species and resulting estimates of N and P contributions from Fig. 3.

Estimated daily contributions of nitrogen (N, upper figure) and phosphorus (P, lower figure) from night-roosting waterbirds in relation to their body mass. The dataset for herbivores is redrawn from Hahn et al. (2008), whereas the dataset for carnivores have been compiled from the dataset Hahn et al. (2007) used to generate their Fig. 1a and b (both redrawn with permission from the authors). For each element, “a” indicates estimates based on an ‘food intake model’ and “b” based on a ‘drop** model’ (see further details in the Hahn et al., 2007, 2008)
We then scaled these up to the annual contribution to each lake complex based on the modelled numbers of bird-days for all species utilising the waterbodies for night-time roosting which fed elsewhere and combined these for each waterbody to give an annual N and P deposition for each waterbody per year.
Estimating inputs of N and P from other sources within the Eastern Vejler lakes
Regrettably, no recent studies are available to determine the current mass balances for N and P in three of the four Eastern Vejler lakes, so we can only compare the contributions of N and P from the roosting birds with historical estimates of contributions from the catchment and the atmosphere to their N and P budgets. Previous studies of N and P inputs from upstream catchment areas and atmospheric deposition are available from 1995 and 2000 for Glombak and Selbjerg Vejle, and for Lund Fjord in 1999–2000 (Cowiconsult, 2000; Jeppesen et al., 2002), which were repeated at Lund Fjord during 2017–2018 (Skovgaard et al., 2019). Since there have been no major changes in land use in the catchment upstream from our study area, we consider it legitimate to use data from these periods. Full methods can be found in these source references.
Results
Overall species contribution to abundance of roosting birds present
In the year between October 2018 and September 2019, the counts registered just under 4 million bird-days of waterfowl species roosting overnight on the combined study sites, with an additional 71,000 bird-days contributed by diving ducks using the complex as daytime resting places. Taken across the entire study area (including the meadows of Bygholmengen), geese completely dominated the total contribution of birds using the wetlands for roosting, since no other groups of bird species exceeded 5% of the calculated bird-days resting there, either by day or night in all five sub-areas studied (Fig. 4). Significant local numbers of goldeneye were also observed to overnight on Glombak and Selbjerg Vejle as well as day-resting diving ducks on Lund Fjord. However, the vast majority of the waterbirds and especially those of the two most numerous species, barnacle and pink-footed geese, spent the night on the meadows of Bygholmengen (Fig. 4).

Average monthly counts of overnight roosting swans, geese, goldeneye, common cranes and gulls as well as day-roosting diving ducks recorded in the five sub-areas of the Eastern Vejler, between October 2018 and September 2019. Note the major difference between the y-axes for the two sets of graphs
Annual cycle in roosting bird abundance
Numbers of geese overnighting in the study areas peaked during autumn and spring, as was largely the case for the other species groups; no species occurred as night- or day-roosting in significant numbers in summer between June and August on Bygholmengen (Fig. 5), but greylag geese did so in two of the lakes, i.e., Selbjerg Vejle and Lund Fjord (further details below).

Monthly average number of whooper swans, geese, goldeneye, common cranes and gulls (overnight roosting) and diving ducks (day-resting) on Bygholmengen, October 2018 to September 2019. Note the difference between the y-axes
Tracking data confirming geese remain on their overnight roost site all night
Of the 56 pink-footed geese marked with tracking devices, 22 occurred at some stage of their annual cycle within the boundaries of the two Special Protection Areas (SPAs, Fig. 6). Plotting the diurnal movements made by these individuals showed that they all foraged to a lesser extent on the meadows of Bygholmengen, but all flew out during the most part of the daylight hours to forage on arable land in the vicinity of Vejlerne. These pink-footed geese were almost entirely confined within the pentagonal area of Bygholmengen during the night hours, only on a very few occasions resorting to Selbjerg Vejle and the southern end of Bygholm Nord (Fig. 6). The telemetry data thus confirmed that pink-footed geese seen flying to and from overnight resting place remain within the area to which they resort for roosting throughout the hours of darkness. It was conspicuous that the tagged pink-footed geese remained far longer during the extended hours of daylight during spring and autumn (see Supplementary Materials Fig. S1), arriving earlier prior to sunset and remaining later after sunrise. This explains the apparent abundance of daylight fixes on the Bygholmengen (Fig. 6), but since they only made a single visit to this area during each diurnal cycle, their prolonged stay makes no difference to additional N and P contributions to the site from elsewhere. Despite the almost complete lack of use of the open waterbodies of Glombak, Selbjerg Vejle, Han Vejle, Lund Fjord, Bygholm Vejle Nord, it is interesting to note that the Western Vejler Lakes (not subject to this study) were used to a greater extent by night-resting geese (Fig. 6).

Distribution of positions recorded every 10 min of 22 pink-footed geese Anser brachyrhynchus bearing Ornitela GPS-GSM collars in the vicinity of Vejlerne from September 2018 to April 2019. The top map shows their distribution by day (here defined as from 1 h before sunrise to 1 h after sunset), the bottom shows the distribution at night (all other times). Image shows use of the Eastern Vejler Special Protection Area (SPA) No. 13 (subject of this study, outlined in yellow, where geese very rarely used the lakes), but also use of the Western Vejler SPA No. 20 (outlined in orange), where geese conspicuously used open water for night-time roosting. Note that the use of Bygholmengen during the day (upper image, bottom right corner of the map) was due to the tagged geese remaining for longer duration on the roost site (especially during autumn and spring when daylength was longer) and not due to geese returning to this area to feed or roost again during the day
Total estimated annual deposition of N and P from roosting birds to the Eastern Vejler
The distribution and phenology of the species means that the vast majority of N and P deposited onto the Eastern Vejler by birds foraging outside of the study area, but roosting there by day or night, are deposited on the meadows of Bygholmengen during the spring and autumn months (Fig. 7). The greatest source of additional nutrient is deposited on Bygholmengen, where the contribution amounted to up to 1,200 kg N and 145 kg P/month, while the lakes received between 20 and 60 kg N and 2 and 6 kg P/month. The annual inputs from day- or night-resting waterfowl were calculated to be 5.4 tonnes N and 626 kg P to Bygholmengen, and between 50 and 145 kg N and 5 and 19 kg P for the lakes (Table 2). Numbers of greylag geese roosting in the study area peaked in August, when birds flew out to feed locally on waste grain available on newly harvested fields, returning to roost and make significant contributions to Selbjerg Vejle and Lund Fjord (see Fig. 7). However, their local abundance and persistence were restricted compared to the large numbers of other geese occurring at other times of the year.

Estimated monthly contributions (mean, maximum and minimum), see “Methods” section for details of N (right) and P (left) from night and day-resting waterfowl to Bygholmengen (top) as well as the four investigated lakes in the Eastern Vejler, October 2018 to September 2019. The conspicuous spikes in August at Lund Fjord and Selbjerg Vejle are due to the contributions of roosting greylag geese feeding in stubbles and roosting here at this time of year. Note the difference in the y-axes scales, and especially that Bygholmengen (uppermost graphs) by far has the highest values for both N and P
Estimated annual deposition of N and P from sources other than roosting birds to the Eastern Vejler
Previous studies of nutrient inputs from catchment areas and atmospheric deposition estimated annual inputs of 32.5–41.1 tonnes N and 1,300–2,070 kg P to Glombak, and 31.0–50.0 tonnes N and 840–2,330 kg P to Selbjerg Vejle in 1995 and 2000, respectively, where the lower values for both lakes were from a dry year, and the higher from a wet year (Table 3). Nutrient contributions to Lund Fjord were estimated at 62 tonnes N and 2,350 kg P in 1999–2000 (wet years), whereas a more recent study found that 38 tonnes of N and 2.3 tonnes of P were added in 2017–2018 to Lund Fjord (the first year dry and the second wet)—i.e., significantly less nitrogen, but the same amount of phosphorus as 20 years ago, but in both years with major contributions from backflow of water from Lund Fjord Kanal. Nutrient contributions from buried tunnel valleys in the study site are probably negligible, given the absence of known valleys in in the Eastern Vejler area (see maps and in Sandersen et al., 2023).
The relative contributions of N and P from avian faeces compared to other sources
If we compare the birds' estimated average annual inputs in 2017–2018 with the values from other sources mentioned above, the roosting birds' contribution to these three large lakes, all of which have larger catchments, amounts to only 0.2–0.4% of all N inputs and 0.5–2.0% of all P inputs (Table 3), where the higher avian values are for years with low precipitation, hence years with less input of nutrients from the catchments. Even if we used the upper values for annual nutrient contributions from birds (Table 2), birds at most would add up to 0.6% of N and 2.1% of P to Selbjerg Vejle, the lake with the highest avian contributions of these three.
For Han Vejle (a closed system without an upstream catchment area, Fig. 1), it is not possible to establish with certainty that the birds' contribution to the nutrient load is equally low. Of the four lakes considered here, Han Vejle is the one that receives the largest area-adjusted supply of nutrients, with 1.24 kg N/ha and 0.13 kg P/ha (Table 2). If we assume that there is no backflow of water from Lund Fjord Kanal to Han Vejle, the only other source of supply to this lake is atmospheric deposition, which in 2018 for the local area amounted to 10–14 kg N/ha (Ellermann et al., 2019), hence an average atmospheric deposition of 492 (average, range 410–574) kg N to the 41 ha lake, compared to 49.4 (average, range 34.5–64.3) kg N from birds (Table 2). The combined atmospheric and avian contributions thus sum to 541.4 (range 444.5–638.3) kg N/ha, and the birds on average contribute 9.1% (range 5.7–13.6) of the total N supply to the lake, where the highest %-value is calculated from a combination of the lowest atmospheric with the highest avian estimate, and vice versa. The average atmospheric deposition of inorganic phosphorus in Denmark lies between 0.02 and 0.04 kg P/ha, with similar levels of organic P, equating to a total atmospheric P contribution of 0.04–0.08 kg P/ha (Ellermann et al., 2015), i.e., an atmospheric deposition of 2.5 (average, range 1.6–3.3) kg P to the 41 ha lake. Birds are estimated to add 5.0 (average, range 4.8–5.3) kg P. The combined atmospheric and avian contributions thus sum to 7.5 (average, range 6.4–8.6) kg P, and the birds may contribute 67.2% (average, range 59.2–76.3%) of the total P supply to the lake (range of % calculated using the same method as for N). Thus the birds definitely seem to be a major contributor of P to this lake, again assuming no contribution from water flowing into the lake from Lund Fjord Kanal, but see the discussion for a more detailed consideration of this assumption.
For the meadows of Bygholmengen (which also represent a relatively closed system without a major upstream catchment area, Fig. 1), the supply of nutrients by roosting birds per unit area was significantly higher than in any of the lakes with averages of 7.6 kg N/ha and 0.88 kg P/ha (Table 2), summing up to 8.5 (average, range 7.1–9.9) tonnes N and 626 (average, range 604–648) kg P deposited on this 710 ha wetland in the study year (Table 2). Here combined avian and atmospheric contributions equated to 13.9 (average, range 10.9–16.9) tonnes N, so the contribution from birds was 38.6% (average, range 27.6–49.4%) of total N. In the case of P, the contribution from birds and the atmosphere was 669 (average, range 632–705) kg P, so the avian contribution equated to 93.6% (average, range 91.4–95.8%) of the total P supply.
Discussion
These results show that numbers of farmland-feeding geese using Vejlerne as their overnight roost site have increased fivefold in the last 30 years and therefore the annual deposition of this source of externally derived organic N and P to the biological system must also have increased. Despite this, the combination of counts and tracking of individual pink-footed geese show that, for three out of four studied lakes, this contribution is trivial compared to other sources (while accepting the need for contemporary confirmation of these quantities) and cannot be the explanation for recent increases in eutrophication associated with enhanced P in their systems over the same period (He et al., 2020). We therefore conclude that the observed eutrophication of the three lakes Glombak, Selbjerg Vejle and Lund Fjord was not caused by import of N and P by roosting birds, hence the proposed exemption of Lund Fjord from the WFD goal of achieving at least “good” ecological status, due to avian nutrient enrichment cannot be supported. This is especially the case because these lakes have been eutrophicated for some time (Kortegaard, 1974; Møller, 1980; Jeppesen et al., 2002), well before recent increases in locally roosting waterbirds. Hence, these nutrients must originate from other sources, i.e., from upstream nutrient contributions from the agricultural catchment.
Because of the greater and increasing concentration of roosting geese in Bygholmengen, where the contributions of N and P are major in an area with almost no upstream catchment, the presence of large numbers of roosting geese has the potential to affect vegetation through nutrient enrichment and persistent goose presence in this area. Lack of N especially can be a limiting factor affecting the growth of some saltmarsh plants and affecting species composition (e.g., Kiehl et al., 1997, Rozema et al., 2002) and the recent botanical survey of Bygholmengen showed that total species richness was limited by the availability of nitrogen (Andersen et al., 2020). Total soil N content measured on Bygholmengene (13.49 g/kg, Lauridsen et al., 2021) was higher than recorded on other European saltmarshes (< 6.1 g/kg, Lauridsen et al., 2021), which might suggest roosting geese could influence soil N at this site. In contrast, levels of total P (1.42 mg/100 g) were comparatively low compared to other Danish saltmarshes (3.8 mg/100 g, Nygaard et al., 2016), but similar to saltmarshes elsewhere in Europe (Lauridsen et al., 2021). How the interaction between enhanced nutrient application and grazing by geese may affect individual plant species growth, community composition and ultimately affect biodiversity will be an important aspect to monitor in coming future years. The geese deposit faeces onto a saltmarsh system, but the area is pockmarked with salt pans, small ponds and larger waterbodies and which is inundated at times, making following and understanding nutrient flows and transport arising from waterbird origins difficult to model and predict. In this respect, it would be informative to use stable isotopes analyses to partition the relative contributions to saltmarsh plants and soil from different nutrient sources (especially N, as successfully applied by Kitchell et al., 1999; Paredes et al., 2019, 2020),
We must be prudent about our ability to compile precise N budgets for these goose species. It is known that greylag geese in late summer and early autumn feed intensively in the morning and late afternoon/evening (as do some pink-footed geese), returning to the roost in the middle of the day (e.g., Madsen, 2001), thereby doubling their contribution to a site and causing the peaks in August/September evident in Fig. 7. For most recent years, the contribution from greylag geese remains very modest compared to that from barnacle and pink-footed geese because of their lesser abundance and restricted timing on site. It is also evident that some of the tracked pink-footed geese were present on Bygholmengen during the daytime (see Fig. 7). However, these were birds that remained on the meadows after the overnight roost before departing for daytime feeding areas (especially in the late autumn and spring). They were resting and feeding in situ and so did not comprise birds returning to Bygholmengen again later during the daytime, which would have resulted in double the imported faeces and nitrogen contributions (see Supplementary Materials Fig. S1). In other words, if these birds were feeding on site, they were recycling material within Bygholmengen, not contributing external N and P from elsewhere. Hence, although we accept these methods may slightly underestimate the true contributions from goose faecal material, we are confident they provide robust estimates of the N and P contributions from this source.
There has been a fivefold increase in numbers of farmland-feeding geese night-roosting almost exclusively on Bygholmengen since the 1990s. This means that these birds potentially contribute even greater nutrient contributions in faeces from food accumulated elsewhere deposited at nocturnal roosts, which increasingly add to background nutrient loads in such a saltmarsh system, which under normal conditions is a vegetation type limited by nitrogen (Andersen et al., 2020). While we found no evidence of vegetation change associated with this increase (Lauridsen et al., 2021) and no consequent knock-on effect on the breeding waterbirds of Bygholmengen (Nielsen & Clausen, 2023), we urge continued monitoring to ensure this is the case for the future. Particular priority ought also to be given to improving our understanding of the short- and longer-term effects of contributions of N and P from roosting geese on the vegetation and biological diversity of the meadow systems of Bygholmengen which represent such a vital element of Vejlerne Nature Reserve. The magnitude of N and P from these birds is especially important in case they cause ecological change to other important nature conservation features of the site. The meadows of Bygholmengen are internationally important for their staging and nesting birds, although currently there is no evidence that vegetation change is occurring or affecting the avifauna (Nielsen & Clausen, 2023). Nevertheless, we consider it important to continue the monitoring of the vegetation height, species composition and biodiversity interests of Bygholmengen in relation to the abundance and breeding success of meadow birds nesting there to ensure nature management targets set for this area continue to be met.
We warn against generalising from our results from a few lakes, as the import of N and P in the forms of faecal deposition by roosting birds can impact the aquatic environment, as demonstrated at some other lakes in Denmark. This is because the effects of such nutrient contributions to trophic conditions are context specific. For instance, oligotrophic lakes with a clear water column [including the relatively oligotrophic Lobelia lakes in Thy National Park (Pedersen et al., 2016) and Østerø Lake near Nyborg (Jensen et al., 2019)], and particularly those nutrient poor lakes without inlets or outlets or with a small catchment, can be rapidly adversely affected by the foraging effects of geese (or other aggregations of birds) and the disproportionate contribution of their faeces to prevailing nutrient poor conditions to which much of their biota are adapted. We should also be mindful that herbivorous species deposit conspicuously less N and P per capita than piscivorous, insectivorous and more carnivorous species (such as the gulls as shown in Fig. 3), so the composition of the roosting waterbird community also has a major effect on the deposition characteristics of waterbodies. Nevertheless, the results from Glombak, Selbjerg Vejle and Lund Fjord confirmed the general absence of large numbers of overnight roosting pink-footed geese and a negligible contribution of N and P from those birds of other species (principally gulls) that did roost on these sites, confirming continued eutrophication was not linked to avian deposition, but more likely was sourced from the upland catchment.
We need improved N and P balance models for at least three of the systems presented here to monitor the continued effects of changes in all nutrient contributions to each of the systems to determine the cause of recent deterioration and eutrophication in these systems and to ultimately tackle the core reasons for these. Again, given the difference in stable isotopes from different nutrient sources (especially agricultural runoff compared to other local sources), it would be insightful to analyse their relative abundance in lake water nutrients (e.g., Kitchell et al., 1999; Paredes et al., 2019, 2020). Regrettably, no recent studies were possible to determine the current mass balances for N and P in the Eastern Vejler lakes, so we can only compare the contributions of these from the roosting birds with historical estimates from the earlier reports of their N and P budgets. In the case of the clear water lake Han Vejle, prior to 2014, major overflow of water from the Lund Fjord Kanal occurred on several occasions into the lake, due to an eroded dike east of the lake. In 2014, this dike was renovated and raised by 1.5 m, after which no dike overflows have been registered (Niels Dahlin Lisborg, personal communication). Despite this, occasional high water levels in Lund Fjord Kanal (as in March 2019, Lauridsen et al., 2021) could have caused flooding over the sluice at Han Vejle’s outlet. The degree of overflow and associated level of nutrient inputs from the nutrient-rich Lund Fjord Kanal into the lake is unknown, but would potentially make a major difference to the nutrient balance of the lake.
In conclusion, we find little evidence of avian deposition of N and P affecting the nutrient status and conservation interest of three shallow lakes (Glombak, Selbjerg Vejle and Lund Fjord), waterbodies which together form the core area of the Eastern Vejler SPA, one of the most important bird reserves in Denmark. We were unable to draw solid conclusions from a fourth lake, Han Vejle due to data insufficiencies; however, it is clear that avian nutrient deposition can potentially make a significant contribution in this lake. Thus, our study shows that lakes with large external or internal nutrient loading may not be sensitive to avian deposition, whereas other studies have shown that nutrient poor lakes or isolated lakes with little catchment can be sensitive to avian N and P deposition (Pedersen et al, 2016; Jensen et al., 2019).
The results of this study confirm the need for evidence-based assessments of avian nutrient inputs relative to the total nutrient balance of individual lakes. In Denmark, there are 24 lakes currently exempted from the WFD, and 35 additional lakes which have been suggested for exemption from the WFD because of their current moderate/poor/bad quality supposedly due to natural loadings resulting from nutrient import by waterbirds. As mentioned above, one of the four lakes studied here were among those suggested to be exempted for achieving at least “good” ecological status, the experiences reported here underline the need for WFD exemption on the basis of avian nutrient contribution needs to be established on a case-by-case basis. The results are less convincing in relation to Bygholmengen, an area of saltmarsh and wet meadows, where an estimated 39% of total N and 94% of total P is likely contributed by dense concentrations of night-roosting geese importing faecal matter. Although there is no current indication of such nutrient enrichment affecting the vegetation, or other features of its staging and breeding avifauna, in a vegetation type known to be typically limited by the supply of nitrogen, it would seem wise to monitor the situation in coming years to monitor for adverse signs of change.
Data availability
All data are available in the literature or can be made available on reasonable request.
Code availability
Not applicable.
Consent for publication
Not applicable.
References
Albert, J. S., G. Destouni, S. M. Duke-Sylvester, A. E. Magurran, T. Oberdorff, R. E. Reis, K. O. Winemiller & W. J. Ripple, 2021. Scientists’ warning to humanity on the freshwater biodiversity crisis. Ambio 50: 85–94. https://doi.org/10.1007/s13280-020-01318-8.
Almond, R. E. A., M. Grooten & T. Petersen (eds), 2020. Living Planet Report 2020 – Bending the Curve of Biodiversity Loss. WWF, Gland [available on internet at https://www.worldwildlife.org/publications/living-planet-report-2020].
Andersen, L. H., A. S. K. Skærbæk, T. B. Sørensen, J. S. Knudsen, C. Pertoldi, S. Bahrndorff & D. Bruhn, 2020. Turnover and change in plant species composition in a shielded salt marsh following variation in precipitation and temperature. Journal of Vegetation Science 31(3): 465–475. https://doi.org/10.1111/jvs.12863.
Bergström, A. K. & M. Jansson, 2006. Atmospheric nitrogen deposition has caused nitrogen enrichment and eutrophication of lakes in the northern hemisphere. Global Change Biology 12(4): 635–643. https://doi.org/10.1111/j.1365-2486.2006.01129.x.
Bouwman, A. F., G. van Drecht, J. M. Knoop, A. H. W. Beusen & C. R. Meinardi, 2005. Exploring changes in river nitrogen export to the world’s oceans. Global Biogeochemical Cycles 19(1): GB1002. https://doi.org/10.1029/2004GB002314.
Brotherston, W., 1964. The numbers and behaviour of geese in the Lothians and Berwickshire. Wildfowl 15: 57–70 [available on internet at https://wildfowl.wwt.org.uk/index.php/wildfowl/article/download/248/248].
Cappelen, J. (ed), 2021. Denmark – DMI Historical Climate Data Collection 1768–2020. Danish Meteorological Institute, Copenhagen.
Cargill, S. M., 1981. The effects of grazing by lesser snow geese on the vegetation of an arctic salt marsh. Unpublished MSc Thesis, University of Toronto.
Carpenter, S. R., 2005. Eutrophication of aquatic ecosystems: bistability and soil phosphorus. Proceedings of the National Academy of Sciences of USA 102(29): 10002–10005. https://doi.org/10.1073/pnas.0503959102.
Clausen, P. & T. Bregnballe, 2017. Metode til overvågning af fugle i søer med henblik på opgørelse af fugles eksterne næringsbelastning af søer. Technical Note 20 March 2017. Aarhus University, DCE – Danish Centre for Environment and Energy (in Danish).
Clausen, P., I. K. Petersen, T. Bregnballe & R. D. Nielsen, 2019. Numbers of Non-breeding Birds in Special Protection Areas for Birds in Denmark, 2004 to 2017. Technical Report No. 148. Aarhus University, DCE – Danish Centre for Environment and Energy (in Danish with English summary) [available on internet at http://dce2.au.dk/pub/TR148.pdf].
Collen, B., J. Loh, S. Whitmee, L. McRae, R. Amin & J. E. M. Baillie, 2009. Monitoring change in vertebrate abundance: the Living Planet Index. Conservation Biology 23: 317–327. https://doi.org/10.1111/j.1523-1739.2008.01117.x.
Cook, S. C., L. Housley, J. A. Back & R. S. King, 2018. Freshwater eutrophication drives sharp reductions in temporal beta diversity. Ecology 99: 47–56. https://doi.org/10.1111/1365-2745.13859.
Cowiconsult, 2000. Naturgenopretning, Østlige Vejlerne, indledende undersøgelser. Report to the Danish Forest and Nature Agency. Cowi Consult A/S, Kongens Lyngby (in Danish).
Cramp, S. & K. E. L. Simmons (eds), 1977. The Birds of the Western Palearctic. Vol. 1. Oxford University Press, Oxford.
Cramp, S. & K. E. L. Simmons (eds), 1980. The Birds of the Western Palearctic. Vol. 2. Oxford University Press, Oxford.
Degener, P., 1994. Naturområdet Vejlerne. Nordjyllands Amt, Viborg Amt og Skov- og Naturstyrelsen. Danish Ministry of the Environment, Copenhagen (in Danish).
Dessborn, L., R. Hessel & J. Elmberg, 2016. Geese as vectors of nitrogen and phosphorus to freshwater systems. Inland Waters 6: 111–122. https://doi.org/10.5268/IW-6.1.897.
Duan, S., S. S. Kaushal, P. M. Groffman, L. E. Band & K. T. Belt, 2012. Phosphorus export across an urban to rural gradient in the Chesapeake Bay watershed. Journal of Geophysical Research 117: G01025. https://doi.org/10.1029/2011JG001782.
Ebbinge, B., K. Canters & R. Drent, 1975. Foraging routines and estimated daily food intake in Barnacle Geese wintering in the northern Netherlands. Wildfowl 26: 5–19 [available on internet at: https://wildfowl.wwt.org.uk/index.php/wildfowl/article/view/489/489].
Ellermann, T., R. Bossi, J. Christensen, P. Løfstrøm, C. Monies, L. Grundahl & C. Geels, 2015. Atmospheric Deposition 2014. NOVANA Report 163. Danish Centre for Environment and Energy, Aarhus University, Aarhus (in Danish with English summary) [available on internet at http://dce2.au.dk/pub/SR163.pdf].
Ellermann, T., R. Bossi, J. Nygaard, J. Christensen, P. Løfstrøm, C. Monies, C. Geels, I. E. Nilesen & M. Poulsen, 2019. Atmospheric Deposition 2018. NOVANA Report 351. Danish Centre for Environment and Energy, Aarhus University, Aarhus (in Danish with English summary) [available on internet at http://dce2.au.dk/pub/SR351.pdf].
Fox, A. D. & K. F. Abraham, 2017. Why geese benefit from the transition from natural to agricultural habitats. Ambio 46(Supplement 2): 188–197. https://doi.org/10.1007/s13280-016-0879-1.
Fox, A. D. & J. Kahlert, 1999. Adjustments to nitrogen metabolism during wing moult in greylag geese. Functional Ecology 13: 661–669. https://doi.org/10.1046/j.1365-2435.1999.00356.x.
Fox, A. D. & J. Madsen, 2017. Threatened species to super-abundance: the unexpected international implications of successful goose conservation. Ambio 46(Supplement 2): 179–187. https://doi.org/10.1007/s13280-016-0878-2.
Fox, A. D., J. Elmberg, I. Tombre & R. Hessel, 2017. Agriculture and herbivorous waterfowl: a review of the scientific basis for improved management. Biological Reviews 92: 854–877. https://doi.org/10.1111/brv.12258.
Fox, A. D., T. J. S. Balsby, H. E. Jørgensen, T. L. Lauridsen, E. Jeppesen, M. Søndergaard, K. Fugl, P. Myssen & P. Clausen, 2019. Effects of lake restoration on breeding abundance of declining common pochard. Hydrobiologia 830: 33–44. https://doi.org/10.1007/s10750-018-3848-9.
Fox, A. D., H. E. Jørgensen, T. L. Lauridsen, E. Jeppesen, M. Søndergaard, K. Fugl, P. Myssen, T. J. S. Balsby & P. Clausen, 2020. Waterbird community changes as indicators of shallow nutrient-rich lake restoration to clear water status. Aquatic Conservation: Marine and Freshwater Ecosystems 30: 237–245. https://doi.org/10.1002/aqc.3260.
Gleick, P. H., 1996. Water resources. In Schneider, S. H. (ed), Encyclopedia of Climate and Weather Oxford University Press, New York: 817–823.
Gould, D. J. & M. R. Fletcher, 1978. Gull drop**s and their effects on water quality. Water Research 12(9): 665–672. https://doi.org/10.1016/0043-1354(78)90176-8.
Hahn, S., S. Bauer & M. Klaassen, 2007. Estimating the contribution of carnivorous waterbirds to nutrient loading in freshwater habitats. Freshwater Biology 52: 2421–2433. https://doi.org/10.1111/j.1365-2427.2007.01838.x.
Hahn, S., S. Bauer & M. Klaassen, 2008. Quantification of allochthonous nutrient input into freshwater bodies by herbivorous waterbirds. Freshwater Biology 53: 181–193. https://doi.org/10.1111/j.1365-2427.2007.01881.x.
Han, H. & J. D. Allan, 2012. Uneven rise in N inputs to the Lake Michigan Basin over the 20th century corresponds to agricultural and societal transitions. Biogeochemistry 109: 175–187. https://doi.org/10.1007/s10533-011-9618-7.
Harper, D. M., 1992. Eutrophication of Freshwaters, Chapman & Hall, London:
He, H., E. Jeppesen, D. Bruhn, M. Yde, J. K. Hansen, L. Spanggaard, N. Madsen, W. Liu, M. Søndergaard & T. L. Lauridsen, 2020. Decadal changes in zooplankton biomass, composition, and body mass in four shallow brackish lakes in Denmark subjected to varying degrees of eutrophication. Inland Waters 10: 186–196. https://doi.org/10.1080/20442041.2020.1732782.
Heldbjerg, H., T. Nyegaard, P. Clausen, R. D. Nielsen & A. D. Fox, 2024. Citizen science data confirm that expanding non-breeding distributions of goose and swan species correlate with their increasing abundance. Ibis. https://doi.org/10.1111/ibi.13302.
Jensen, T. K., S. Egemose, F. Ø. Andersen & K. Reitzel, 2019. Fugle som ekstern næringsstofkilde. Vand Og Jord 26: 158–161 (in Danish).
Jeppesen, E., J. P. Jensen, M. Søndergaard, T. Lauridsen & F. Landkildehus, 2000. Trophic structure, species richness and biodiversity in Danish lakes: changes along a phosphorus gradient. Freshwater Biology 45(2): 201–218. https://doi.org/10.1046/j.1365-2427.2000.00675.x.
Jeppesen, E., M. Søndergaard, S. Amsinck, J. P. Jensen, T. L. Lauridsen, L. K. Pedersen, F. Landkildehus, K. Nielsen, D. Ryves, O. Bennike, G. Krog, I. Christensen & P. Schriver, 2002. Søerne i De Østlige Vejler. Scientific Report No. 394. Danish National Environmental Research Institute, Aarhus University, Aarhus (in Danish) [available on internet at https://www2.dmu.dk/1_viden/2_publikationer/3_fagrapporter/rapporter/fr394.pdf].
Kiehl, K., P. Esselink & J. P. Bakker, 1997. Nutrient limitation and plant species composition in temperate salt marshes. Oecologia 111: 325–330. https://doi.org/10.1007/s004420050242.
Kitchell, J. F., D. E. Schindler, B. R. Herwig, D. M. Post, M. H. Olson & M. Oldham, 1999. Nutrient cycling at the landscape scale: the role of diel foraging migrations by geese at the Bosque del Apache National Wildlife Refuge, New Mexico. Limnology and Oceanography 44(3): 828–836. https://doi.org/10.4319/lo.1999.44.3_part_2.0828.
Kjeldsen, J. P., 2008. Breeding birds in Vejlerne. Dansk Ornitologisk Forenings Tidsskrift 102: 1–238 (in Danish with English summary) [available on internet at https://pub.dof.dk/artikler/162/download/doft-102-2008-1-238-ynglefugle-i-vejlerne-joergen-peter-kjeldsen].
Kortegaard, L. 1974. An ecological outline of a moulting area of Teal, Vejlerne, Denmark. Wildfowl 25: 134–142 [available on internet at https://wildfowl.wwt.org.uk/index.php/wildfowl/article/viewFile/485/485].
Laubek, B., P. Clausen, L. Nilsson, J. Wahl, M. Wieloch, W. Meissner, P. Shimmings, B. H. Larsen, M. Hornman, T. Langendoen, A. Lehikoinen, L. Luigojõe, A. Stīpniece, S. Švažas, L. Sniaukstra, V. Keller, C. Gaudard, K. Devos, Z. Musilová, N. Teufelbauer, E. Rees & A. D. Fox, 2019. Whooper Swan Cygnus cygnus January population censuses for Northwest Mainland Europe, 1995–2015. Wildfowl Special Issue 5: 102–121 [available on internet at https://wildfowl.wwt.org.uk/index.php/wildfowl/article/view/2707/pdf_175].
Lauridsen, T. L., D. Bruhn, P. Clausen, L. H. Andersen, C. Pertoldi, E. Jeppesen, M. Søndergaard, E. Levy, A. D. Fox, T. J. S. Balsby, S. Bahrndorff, H. He, C. L. Pedersen & H. H. Nielsen, 2021. Udvikling af en forvaltningsstrategi, der tilgodeser hele økosystemet i De Østlige Vejler. Scientific Report No. 428. Danish Centre for Environment and Energy, Aarhus University, Aarhus (in Danish with English summary) [available on internet at https://dce2.au.dk/pub/SR428.pdf].
Lehner, B. & P. Döll, 2004. Development and validation of a global database of lakes, reservoirs and wetlands. Journal of Hydrology 296: 1–22. https://doi.org/10.1016/j.jhydrol.2004.03.028.
Ligeza, S. & H. Smal, 2003. Accumulation of nutrients in soils affected by perennial colonies of piscivorous birds with reference to biogeochemical cycles of elements. Chemosphere 52(3): 595–602. https://doi.org/10.1016/S0045-6535(03)00241-8.
Madsen, J., 2001. Can geese adjust their clocks? Effects of diurnal regulation of goose shooting. Wildlife Biology 7: 213–222. https://doi.org/10.2981/wlb.2001.026.
Madsen, J., J. H. Williams, F. A. Johnson, I. M. Tombre, S. Dereliev & E. Kuijken, 2017. Implementation of the first adaptive management plan for a European migratory waterbird population: the case of the Svalbard pink-footed goose Anser brachyrhynchus. Ambio 46(Supplement 2): 275–289. https://doi.org/10.1007/s13280-016-0888-0.
Martín-Vélez, V., M. I. Sánchez, J. Shamoun-Baranes, C. B. Thaxter, E. W. M. Stienen, K. C. J. Camphuysen & A. J. Green, 2019. Quantifying nutrient inputs by gulls to a fluctuating lake, aided by movement ecology methods. Freshwater Biology 64(10): 1821–1832. https://doi.org/10.1111/fwb.13374.
Mateo-Sagasta, J., S. M. Zadeh, H. Turral & J. Burke, 2017. Water Pollution from Agriculture: A Global Review. Executive summary. Food and Agriculture Organisation and International Water Management Institute, Rome and Colombo [available on internet at https://www.fao.org/3/i7754e/i7754e.pdf].
Miljøministeriet, 2023a. Vandområdeplanerne 2021–2027. The Danish Ministry of Environment, Copenhagen [available on internet at https://mim.dk/media/235205/vandomraadeplanerne-2021-2027-22-9-2023.pdf].
Miljøministeriet, 2023b. MiljøGIS for offentliggørelse af vandområdeplaner 2021–2027. The Danish Ministry of Environment, Copenhagen [available on internet at https://miljoegis.mim.dk/spatialmap?profile=vandrammedirektiv3-2022].
Møller, H. S., 1980. Naturforholdene I Vejlerne. Fredningsstyrelsen, The Danish Ministry of Environment, Copenhagen (in Danish).
Moss, B., 2017. Marine reptiles, birds and mammals and nutrient transfers among the seas and the land: an appraisal of current knowledge. Journal of Experimental Marine Biology and Ecology 492: 63–80. https://doi.org/10.1016/j.jembe.2017.01.018.
Moss, B., E. Jeppesen, M. Søndergaard, T. L. Lauridsen & Z. Liu, 2013. Nitrogen, macrophytes, shallow lakes and nutrient limitation: resolution of a current controversy? Hydrobiologia 710: 3–21. https://doi.org/10.1007/s10750-012-1033-0.
Nielsen, H. H. & P. Clausen, 2023. Ynglende og rastende fugle i Vejlerne 2021. Technical Report 277. Danish Centre for the Environment and Energy, Aarhus University, Aarhus (in Danish with English summary) [available on internet at https://dce.au.dk/fileadmin/dce.au.dk/Udgivelser/Tekniske_rapporter_250-299/TR277.pdf].
Nygaard, B., C. Damgaard, K. E. Nielsen, J. Bladt & R. Ejrnæs, 2016. Terrestriske naturtyper 2004–2015, NOVANA. Danish Centre for the Environment and Energy, Aarhus University, Aarhus (in Danish) [available on internet at https://novana.au.dk/fileadmin/novana_au_dk/pic_upload/TerrestriskeNaturtyper2015.pdf].
Paredes, I., F. Ramírez, M. G. Forero & A. J. Green, 2019. Stable isotopes in helophytes reflect anthropogenic nitrogen pollution in entry streams at the Doñana World Heritage Site. Ecological Indicators 97: 130–140. https://doi.org/10.1016/j.ecolind.2018.10.009.
Paredes, I., N. Otero, A. Soler, A. J. Green & D. X. Soto, 2020. Agricultural and urban delivered nitrate pollution input to Mediterranean temporary freshwaters. Agriculture, Ecosystems and Environment 294: 106859. https://doi.org/10.1016/j.agee.2020.106859.
Pedersen, O., L. Baastrup-Spohr, M. Madsen-Østerbye, E. Kristensen, T. Kragh, M. R. Andersen, F. Ø. Andersen & K. Sand-Jensen, 2016. Lobeliesøer – trusler og restaurering. Vand Og Jord 23: 63–66 (in Danish with English summary).
Poikane, S., M. G. Kelly, F. S. Herrero, J. A. Pitt, H. P. Jarvie, U. Claussen, W. Leujak, A. L. Solheim, H. Teixeira & G. Phillips, 2019. Nutrient criteria for surface waters under the European Water Framework Directive: current state-of-the-art, challenges and future outlook. Science of the Total Environment 695: 133888. https://doi.org/10.1016/j.scitotenv.2019.133888.
Poikane, S., M. G. Kelly, G. Free, L. Carvalho, D. P. Hamilton, K. Katsanou, M. Lurling, S. Warner, B. M. Spears & K. Irvine, 2024. A global assessment of lake restoration in practice: new insights and future perspectives. Ecological Indicators 158: 111330. https://doi.org/10.1016/j.ecolind.2023.111330.
Powell, G. V., J. W. Kenworthy & J. W. Fourqurean, 1989. Experimental evidence for nutrient limitation of seagrass growth in a tropical estuary with restricted circulation. Bulletin of Marine Science 44: 324–340.
Powolny, T., G. H. Jensen, S. Nagy, A. Czajkowski, A. D. Fox, M. Lewis. & J. Madsen, 2018. AEWA International Single Species Management Plan for the Greylag Goose Anser anser Northwest/Southwest European population. African Eurasian Waterbird Agreement Secretariat, Bonn [available on internet at https://egmp.aewa.info/sites/default/files/download/population_status_reports/AEWA%20International%20Single%20Species%20Management%20Plan%20for%20the%20Greylag%20Goose_NW_SW%20European%20Population.pdf].
Reid, A. J., A. K. Carlson, I. F. Creed, E. J. Eliason, P. A. Gell, P. T. Johnson, K. A. Kidd, T. J. MacCormack, J. D. Olden, S. J. Ormerod & J. P. Smol, 2019. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biological Reviews 94: 849–873. https://doi.org/10.1111/brv.12480.
Rozema, J., P. Leendertse, J. Bakker & H. Van Wijnen, 2002. Nitrogen and vegetation dynamics in European salt marshes. In Weinstein, M. P. & D. A. Kreeger (eds), Concepts and Controversies in Tidal Marsh Ecology. Springer, Dordrecht. https://doi.org/10.1007/0-306-47534-0_21.
Sandersen, P. B. E., H. Kim, R. Jacobsen, J. B. Petersen & B. Hansen, 2023. Transport of nitrogen-containing groundwater to coastal areas through buried tunnel valleys, Denmark. GEUS Bulletin 53: 8351. https://doi.org/10.34194/geusb.v53.8351.
Schreven, K. H. T., C. Stolz, M. Madsen & B. A. Nolet, 2021. Nesting attempts and success of Arctic-breeding geese can be derived with high precision from accelerometry and GPS-tracking. Animal Biotelemetry 9: 25. https://doi.org/10.1186/s40317-021-00249-9.
Skovgaard, H., M. Larsen & H. S. Mark, 2019. Forforindsats i oplandet til Lund Fjord. Teknisk forundersøgelse [Phosphorus Input in the Catchment Area of Lund Fjord. Technical Feasibility Study]. Orbicon, Viby (in Danish).
Sørensen, B., 1997. Fugles næringsstoftilførsel til søer. Unpublished Masters Thesis, Biological Institute, Aarhus University, Aarhus (in Danish).
Thorup, O., 2004. Status of populations and management of Dunlin Calidris alpina, Ruff Philomachus pugnax and Black-tailed Godwit Limosa limosa in Denmark. Dansk Ornitologisk Forenings Tidsskrift 98: 7–20 [available on internet at https://pub.dof.dk/artikler/732/download/doft-98-2004-7-20-status-for-bestande-og-forvaltning-af-alm-ryle-brushane-og-stor-kobbersneppe-i-danmark].
Tickner, D., J. J. Opperman, R. Abell, M. Acreman, A. H. Arthington, S. E. Bunn, S. J. Cooke, J. Dalton, W. Darwall, G. Edwards, I. Harrison, K. Hughes, T. Jones, D. Leclère, A. J. Lynch, P. Leonard, M. E. McClain, D. Muruven, J. D. Olden, S. J. Ormerod, J. Robinson, R. E. Tharme, M. Thieme, K. Tockner, M. Wright & L. Young, 2020. Bending the curve of global freshwater biodiversity loss: an emergency recovery plan. BioScience 70: 330–342. https://doi.org/10.1093/biosci/biaa002.
Winton, R. S. & M. River, 2017. The biogeochemical implications of massive gull flocks at landfills. Water Research 122: 440–446. https://doi.org/10.1016/j.watres.2017.05.076.
Acknowledgements
Thanks to the Aage V. Jensens Naturfond for financial support to this study. EJ, TLL and MS were also supported by the MARS Project (Managing Aquatic ecosystems and water Resources under multiple Stress) funded under the 7th EU Framework Programme and AU Centre for Water Technology (watec.au.dk). MS was supported by funds from the Poul Due Jensen Foundation and EJ by the TÜBITAK Program BIDEB2232 (Project 118C250). GPS-tags for pink-footed geese were funded by the Netherlands Polar Programme of the Dutch Research Council (Grant No. ALWPP.2016.024) and Aarhus University. We thank Jørgen Peter Kjeldsen and Jens Frimer Andersen who contributed to the roost counts of birds and Tinna Christensen for beautifying the final figures. Thanks to two anonymous referees for their suggestions for improvements to an earlier manuscript.
Funding
Open access funding provided by Aarhus Universitet. This study was primarily funded by the Aage V. Jensen Naturfond, other sources are given in Acknowledgements.
Author information
Authors and Affiliations
Contributions
Preben Clausen developed the study conception and design. Jesper Madsen, Kees Schreven and Bart Nolet contributed the goose telemetry data. Material preparation, data collection and analysis were performed by all contributing authors. The first draft of the manuscript was written by Anthony David Fox and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest.
Ethical approval
The article contains no studies involving animals in experiments performed by any of the authors.
Informed consent
Not applicable.
Additional information
Handling editor: Sidinei M. Thomaz
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Guest editors: Priit Zingel, Helen Agasild, Maria Brigida Boveri & Erik Jeppesen / Secrets of Shallow Lakes: Insights from Research
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Clausen, P., Lauridsen, T.L., Pedersen, C.L. et al. Are increasing roosting waterbird numbers responsible for eutrophication of shallow lakes? Examples from a Danish Ramsar site. Hydrobiologia (2024). https://doi.org/10.1007/s10750-024-05475-9
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10750-024-05475-9