
Abstract
Biodiversity loss is a main challenge for agricultural sustainability. Major drivers include local management and landscape simplification. Therefore, conservation measures aim to increase organic agriculture, reduce pesticide use, and increase the proportion of semi-natural habitats (SNH). Yet, it is important to understand the effects of such measures. We investigate how arthropod biomass, taxa richness, and community composition in Malaise trap samples are affected by organic management, pesticide use, and SNH in the landscape. The 32 studied vineyards were chosen in a crossed design of management (organic vs. conventional) and pesticide use (regular vs. reduced) along a gradient of landscape composition. Pesticide reduction by 55% was obtained by including half of the vineyards with fungus-resistant grape (FRG) varieties. Malaise trap samples were weighed and arthropods identified using metabarcoding. Surprisingly, biomass was almost one-third higher in conventionally managed vineyards compared to organic ones. Taxa richness increased by more than one third when the proportion of SNH in a radius of 1,000 m in the surrounding landscape increased from zero to 50%. Diptera richness tended to be 4% higher in conventionally managed vineyards and the richness of Hymenoptera was 9% higher in FRG varieties. Community composition changed with the proportion of SNH and differed between organic and conventional management. Overall, organic viticulture was not effective to enhance the arthropod community, which was dominated by flying insects in our study. Agricultural policies should therefore rather preserve and promote SNH in the surrounding in order to promote arthropod biodiversity in viticultural landscapes.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In the past decades, arthropods have declined strongly in many landscapes (Hochkirch 2016; Hallmann et al. 2017; Seibold et al. 2019). One of the main drivers for the loss of biomass and biodiversity is considered to be intensive agriculture, including the use of pesticides and fertilizers (Sánchez-Bayo and Wyckhuys 2019). Policy measures, such as the European Green Deal, include increasing the share of organic agriculture, reducing pesticide use, and increasing the proportion of semi-natural habitats (SNH) in agricultural landscapes (European Commission 2019). It is assumed that these measures enhance biodiversity to counteract the strong decline in recent years (Bengtsson et al. 2005). However, effects may vary between and within organism groups and between crops (Bengtsson et al. 2005; Hole et al. 2005). So it is important to understand the differential impacts of such changes in agricultural practices.
Organic farming can promote biodiversity compared to conventional farming (Bengtsson et al. 2005; Hole et al. 2005; Caprio et al. 2015; Beaumelle et al. 2023). Hence, the European Green Deal’s Farm to Fork Strategy targets, for example, to increase the share of organic farming to at least 25% by 2030 (European Commission 2019). In viticulture, at least in regions with no insecticide use, the effects of organic management appear, however, to be less pronounced compared to other crop** systems (Döring et al. 2019; Paiola et al. 2020). The occurrence of fungal diseases, particularly powdery mildew and downy mildew, causes one of the highest use of plant protection products in agriculture (Pertot et al. 2017). Their presence thus necessitates a high use of either synthetic chemicals in conventional viticulture or inorganic compounds such as copper and sulfur in organic viticulture, both of which may affect non-target organisms (Pedneault and Provost 2016; Vogelweith and Thiéry 2018).
Under the Farm to Fork Strategy, efforts are made to reduce the pesticide use in order to mitigate environmental pollution (European Commission 2019). The cultivation of fungus-resistant grape (FRG) varieties allows a reduction of pesticide use by more than 80% due to resistance traits against fungal diseases (Pedneault and Provost 2016). It was recently reported that the cultivation of FRG varieties thus has positive effects on non-target organisms (Pedneault and Provost 2016), promotes the presence of predatory mites over pest mites, and positively affects certain spider families (Pennington et al. 2017, 2019; Reiff et al. 2021a). The promotion of such beneficial species may affect trophic interactions that are important to winegrowers by improving for example natural pest control (Winkler et al. 2017; Reiff et al. 2021a).
In addition to management practices, the landscape may play an important role for species diversity and composition (Bengtsson et al. 2005). A heterogeneous landscape with high proportions of SNH in the surrounding area of agricultural land is reported to promote various organisms (Martin et al. 2019; Kolb et al. 2020; Tscharntke et al. 2021). Therefore, it is recommended and targeted, e.g. by the Biodiversity Strategy of the European Green Deal (European Commission 2019), to increase the proportion of SNH (Martin et al. 2019). Furthermore, high cover and connectivity of non-crop habitat may improve conditions for beneficial organisms such as parasitoids, predators, antagonists, and pathogens of pests in agricultural landscapes (Tscharntke et al. 2021).
Arthropod communities can be comprehensively analyzed using metabarcoding by identifying specimens based on DNA sequences in the cytochrome c oxidase I (COI) gene and clustering similar sequences into operational taxonomic units (OTUs) that can be assigned to barcode index numbers (BINs) based on reference sequences in the Barcode of Life Data System (BOLD) (Ratnasingham and Hebert 2013). In this way, metabarcoding allows the study of entire arthropod communities, including the highly diverse orders of Diptera and Hymenoptera that account for large proportion of species in, for example, Malaise trap samples (Skvarla et al. 2021). Furthermore, metabarcoding may be a valuable tool for long-term monitoring, as taxa can be identified in a time- and cost-efficient way and data effectively stored and shared for re-analysis (deWaard et al. 2019).
In this study, we aimed to distinguish the effects of three major drivers underlying agricultural biodiversity. We investigated how organic farming, reduced pesticide use, and the proportion of SNH in the landscape surrounding vineyards in southwest Germany affect the diversity of arthropods. We used Malaise traps and metabarcoding in a crossed design with FRG and classic varieties in either organically or conventionally managed vineyards along a gradient of landscape composition. In this way, our study design was suitable to assess the impact of key policies that address biodiversity loss in a crop** system with generally high pesticide use. Furthermore, the cultivation of FRG varieties allowed for the study of pesticide reduction in healthy crops under realistic crop** conditions.
We tested the following hypotheses: (H1) arthropod biomass and (H2) richness are higher in (a) organically compared to conventionally managed vineyards, (b) FRG compared to classic varieties, and (c) SNH-rich compared to vineyard-dominated landscapes; and (H3) community composition differs between (a) management types and (b) grape varieties, and (c) with the proportion of SNH in vineyard landscapes.
Materials and methods
Study area
We conducted our study in the German wine-growing region Palatinate (49.273280°N, 8.020602°E/49.147516°N, 8.175736°E; Fig. 1). The region lies in the Upper Rhine Valley east of the Palatinate Forest and is characterized by a temperate climate with an average annual temperature of 11.1 °C and a total annual precipitation of 677.7 mm, and a widespread cultivation of grapevines (Beck et al. 2018; Agrarmeteorologie Rheinland-Pfalz 2022). The mean temperature was 11.9 °C and 10.4 °C with a total precipitation of 630.4 and 814.5 mm in 2020 and 2021, respectively (Agrarmeteorologie Rheinland-Pfalz 2022).

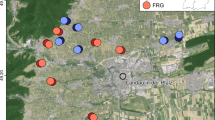
Study area with the location of the 32 vineyards. Each pair of either organically (blue) or conventionally (red) managed vineyards consisted of one fungus-resistant (FRG, brighter) and one classic (darker) grape variety (basic map data by ©GeoBasis-DE/LVermGeoRP (2022))
Design and sampling
We chose 16 pairs of vineyards along a gradient of landscape composition, varying in the proportion of SNH in their surrounding (Fig. 1, Table S1). Half of them were managed organically after the European Union regulation No 2092/91 and the other half conventionally. Each pair consisted of one vineyard with a fungus-resistant grape variety (FRG) and one with a classic variety. We conducted fieldwork from April to September in 2020 and 2021. In each vineyard, we placed a Malaise trap (first 3 months Malaise traps with a combination of black and white net, ENTO SPHINX s.r.o., Pardubice, Czech; then standard SLAM traps, MegaView Science Co., Ltd., Taichung, Taiwan) in a central inter-row for three consecutive days per month. If there were differences in tillage between adjacent inter-rows, we chose the more vegetated row. We filled the collecting bottles with 300 ml of 70% ethanol denatured with about 1% methyl ethyl ketone (EtOH MEK) and stored collected specimens in undiluted EtOH MEK.
Landscape and environmental variables
Within a radius of 1,000 m of each vineyard, the cover of SNH was calculated by using ATKIS data (Basis-DLM by ©GeoBasis-DE/BKG (2013); Table 1) with intersection of spatial data in an Oracle database 12c (Oracle 2017). We defined SNH as forests, hedges, shrubs, and grassland. Within each pair, we used the mean proportion of SNH of the two vineyards for further analyzes, resulting in a total of 16 landscapes.
We received the number of annual pesticide applications from the winegrowers. Predominantly fungicides were sprayed. Herbicides were used in three pairs of the conventional and none of the organic vineyards. To our knowledge, no insecticides were used, but mating disruption of grapevine moths (Lobesia botrana and Eupoecilia ambiguella) was done with pheromone dispensers in the whole study area. We measured vegetation cover and the number of plant species that had flowers during the survey three times each year (between first and second, third and fourth, and fifth and sixth Malaise trap sampling) in two plots per vineyard with two subplots each in adjacent inter-rows. Each subplot was 1 m² (2 m x 0.5 m) in size, covering a total of 4 m² per vineyard. For the vegetation cover, the proportion of ground covered by vegetation was measured visually in tens from 0 to 100%. For analyzes, we used the mean of the variables of all surveys and both years.
Biomass
To obtain arthropod biomass, we weighed the wet catch after placing it in a sieve and letting the liquid drip off. For each vineyard, we used the total biomass of both years for analyzes by summing all samples.
Diversity
We pooled the arthropod material of each year, resulting in two samples per vineyard. Orthopterans were sorted out beforehand and only one leg of each specimen was left in the samples for the DNA metabarcoding analysis in order to avoid the dominance of their DNA. Metabarcoding of a 313 base pairs long mini-barcode region in the CO1-5P target region and bioinformatics were conducted by AIM - Advanced Identification Methods GmbH, following the protocol and methods of Hausmann et al. (2020) and Morinière et al. (2016) and using the VSEARCH suite 2.9.2 (Rognes et al. 2016) and Cutadapt 1.18 (Martin 2011) (Online Resource 1, Supplementary methods). In the bioinformatical process, similar DNA sequences were clustered into operational taxonomic units (OTUs) and assigned to barcode index numbers (BINs) based on reference sequences in the Barcode of Life Data System (BOLD). We filtered the results table for OTUs with a Hit-%-ID value in BOLD ≥ 97% and an assignment to a BIN and condensed BINs that occurred more than once into one entry. For most analyzes, we combined the tables from the 2 years into an overall BIN list. The total number of BINs was used as a taxon richness value. Taxonomic information was used from BOLD.
Data analysis
All analyzes were conducted using R 4.1.2 (R Core Team 2021) and RStudio 2022.07.1 (RStudio Team 2022) with the R packages car (Fox and Weisberg 2019), lme4 (Bates et al. 2015), blmeco (Korner-Nievergelt 2015), MuMIn (Bartoń 2020), vegan (Oksanen et al. 2020), ggplot2 (Wickham 2016), ggpubr (Kassambara 2020), and dplyr (Wickham et al. 2022).
To investigate whether environmental variables differ between management types and grape varieties, we performed linear mixed model regressions (LMM) for spraying events and flowering plant species with management and grape variety as fixed factors and the vineyard pair as random factor. A general linear mixed model regression (GLMM) with negative binomial distribution and logarithmic link function (log link) was used for vegetation cover with the same fixed and random factors. To investigate the effects of management type, grape variety, their interaction, landscape, and environmental variables on biomass and BINs, we performed LMMs for BINs of Coleoptera, Hemiptera, and Lepidoptera and GLMMs with negative binomial distribution and log link for biomass, total BINs, and BINs of Diptera and Hymenoptera and included the vineyard pair as random factor. We rescaled and centered continuous variables. To obtain whole-numbers, we multiplied the values of biomass and vegetation cover by ten. We selected the best fitting model based on the lowest AICc value by using a backward elimination method with management, variety, and SNH as fixed factors. We used type III ANOVA to test the effects using a significance level of P < 0.05. We investigated differences in community composition between years and between management types and grape varieties by performing non-metric multidimensional scaling (NMDS) reduced to two dimensions and with the lowest stress out of 50 runs and tested effects using PERMANOVA with Binary Jaccard distances. Unlike the other analyzes, we did not use the combined BIN list of both years to investigate the differences in community composition between years.
Results
Environmental variables
With two more spraying events compared to conventional managed vineyards (seven applications, SD = ± 3), the number of annual applications tended to be higher under organic management (nine applications, SD = ± 4; Table 2; Fig. 2A). With five annual applications (SD = ± 3 applications), FRG varieties received significantly fewer sprayings than classic varieties with 11 applications per year (SD = ± 2 applications). Vegetation cover was 26% higher in conventionally managed vineyards than in organically managed ones (Fig. 2B). The plant species richness was unaffected by the studied factors (Fig. 2C).

Mean number of spraying events (A), mean proportion of ground covered by vegetation (B), and mean number of plant species (C) of both years with mean and standard deviation for fungus-resistant (FRG, brighter) and classic (darker) grape varieties in organic (green) and conventional (blue) management. Significant effects are indicated in x-axis labels. Significance codes: ***P < 0.001, **P < 0.01, ∙P < 0.1
Biomass
The cumulative wet biomass of arthropods per vineyard was on average 16.2 g (SD = ± 5.3 g), with 9.8 g (SD = ± 3.0 g) in 2020 and 6.4 g (SD = ± 3.2 g) in 2021 (Table S2). We found a significant effect of the management type (Table 3; Fig. 3A). On average, biomass was 31% higher in conventional management (18.3 g, SD = ± 5.3 g) compared to organic management (14.0 g, SD = ± 4.4 g). Biomass increased significantly with increasing number of plant species (Fig. 3C). However, we found no significant influence of the grape variety (Fig. 3A), SNH (Fig. 3B), and vegetation cover on biomass.

Summarized biomass in g of both years for fungus-resistant (FRG, brighter) and classic (darker) grape varieties in organic (green) and conventional (blue) vineyards with mean and standard deviation (A), proportion of semi-natural habitats (SNH) (B), and number of plant species (C). Shaded areas represent the 95% confidence intervals. Significant effects are indicated in x-axis labels. Significance code: **P < 0.01
Diversity
A total of 1,748 different BINs were recorded in this study (Online Resource 1, Supplementary Results; Online Resource 2, Table S1). 1,389 BINs were recorded in 2020 and 1,028 in 2021 (Table 4), with 38.3% of the BINs recorded in both years and significant differences in the community composition between the two years (F1,62 = 10.6, R² = 0.146, P = 0.001; Fig. 4A). On average, we found 200 BINs per site (SD = ± 43 BINs) in 2020 and 142 BINs per site (SD = ± 20 BINs) in 2021. 705 BINs were recorded on only one site either in 2020 or in 2021. The orders with the most BINs were Diptera (664), Hymenoptera (375), Coleoptera (262), Lepidoptera (198), Hemiptera (154), and Araneae (35). We found community composition to be significantly different between organic and conventional management (F1,30 = 1.6, R² = 0.050, P = 0.004) but no difference between grape varieties (F1,30 = 0.8, R² = 0.027, P = 0.870; Fig. 4B).

Non-metric multidimensional scaling analysis (NMDS) for arthropod communities based on the 1,748 barcode index numbers (BIN) at the 32 study sites for (A) 2020 (blue) and 2021 (pink) (2 dimensions, n = 50 runs, stress = 0.19, Binary Jaccard distance) and (B) total communities with fungus-resistant (FRG, brighter) and classic (darker) grape varieties in organic (green) and conventional (blue) management and the variables semi-natural habitats (SNH), organic management, and FRG variety (2 dimensions, n = 50 runs, stress = 0.20, Binary Jaccard distance). Dots in A are colored based on the proportion of SNH in the surrounding landscape of the vineyards. The dots of pairs of vineyards are connected by a line in B. The SNH vector in B is shown shortened by about 25 times to increase visibility of the other data points
For the total number and for most orders, we found no significant effect of the management type on the number of BINs per vineyard (Table 5; Fig. 5). Only for Diptera, richness tended to be 4% higher in conventionally managed vineyards (Fig. 5C). Similarly, we found no significant effect of the grape variety, except for a trend in Hymenoptera with 9% more BINs in FRG varieties (Fig. 5E). There was no significant interaction between management and grape variety. The cover of SNH in the surrounding landscape increased significantly total richness of BINs (Fig. 6A) as well as BINs’ richness within Diptera (Fig. 6C), Hemiptera (Fig. 6D), Hymenoptera (Fig. 6E), and Lepidoptera (Fig. 6F). For vegetation variables, total richness of arthropod BINs increased with the number of present plant species, whereas the amount of vegetation cover had no effect.

Cumulative number of barcode index numbers (BIN) of both years with mean and standard deviation for fungus-resistant (FRG, brighter) and classic (darker) grape varieties in organic (green) and conventional (blue) management for (A) total, (B) Coleoptera, (C) Diptera, (D) Hemiptera, (E) Hymenoptera, and (F) Lepidoptera. Effects with a trend are indicated in x-axis labels. Significance code: ∙ P < 0.1

Cumulative number of barcode index numbers (BIN) of both years for the proportion of semi-natural habitats (SNH) in the surrounding landscape for (A) total, (B) Coleoptera, (C) Diptera, (D) Hemiptera, (E) Hymenoptera, and (F) Lepidoptera. Colors indicate grape variety and management, with fungus-resistant (FRG, brighter) and classic (darker) grape varieties in organic (green) and conventional (blue) management. Shaded areas represent the 95% confidence intervals. Significant effects are indicated in x-axis labels. Significance codes: *** P < 0.001, ** P < 0.01
Discussion
We assessed how organic management, reduced pesticide use, and the proportion of SNH in the surrounding landscape affect arthropod biomass, richness, and community composition in viticulture. The results show that arthropods, mostly composed of flying insects in our study, are affected by both management practices within vineyards and by the surrounding landscape. Within vineyards, this is particularly reflected by reduced biomass under organic management, but also by changes in the community composition rather than species richness.
Contrary to our first hypothesis (H1a), where we expected arthropod biomass to be higher in organically managed vineyards, biomass was lower in organic vineyards than in conventional ones. Thus, an increase in the share of organic farming, as targeted by the European Green Deal (European Commission 2019), may not be efficient in enhancing arthropod biomass in viticulture in our region. We also found no difference in arthropod richness between organic and conventional viticulture, contrary to our hypothesis (H2a) that arthropod richness is higher under organic management. Only for Diptera, taxa richness tended to be 4% higher under conventional management. While a strong positive effect of organic management in agricultural landscapes has been demonstrated for other crop** systems (Bengtsson et al. 2005), the difference between management types in viticulture seems weaker with both positive and negative effects being reported (Döring et al. 2019; Kolb et al. 2020; Paiola et al. 2020). This may be due to the high use of pesticides, in particular fungicides, but a generally low use of insecticides in both management systems. However, large positive effects of organic viticulture over conventional are reported from regions where insecticide use is mandatory against Scaphoideus titanus, the vector of Flavescence dorée (Beaumelle et al. 2023). As we found an effect of management on biomass but only a trend for Diptera richness, the effects appear to vary among organism groups. Ostandie et al. (2021) observed an increased abundance of springtails and spiders in organic versus conventional viticulture, while pollinator abundance decreased and ground beetles as well as mites remained unaffected. Although such effects may also be explained by differences in tillage and plant cover, we did not find effects of vegetation cover on biomass or taxa richness. In the studied vineyards, on average, those managed organically had a lower vegetation cover compared to conventionally managed ones, but this does not necessarily mean that their overall vegetation was less diverse or structured. In fact, we surveyed the highest proportion of vegetation cover in conventionally managed vineyards with mostly dense grass covers, whereas other vineyards often had lower vegetation cover but more diverse plant communities. It is worth noting that newly tilled inter-rows may also have contributed to a temporary lower vegetation cover at the time of the survey. Arthropod biomass and richness did, however, increase with a higher number of plant species. Since there was no significant difference in the number of plant species between management types, the inter-row vegetation does not seem to explain the lower biomass and lower Diptera richness under organic management in our study. Möth et al. (2021) found pesticide toxicity for non-target organisms to be higher in organic vineyards due to the use of copper and sulfur instead of synthetic fungicides. Copper and sulfur are non-specific agents that can also affect non-target organisms (Nash et al. 2010; Pedneault and Provost 2016; Vogelweith and Thiéry 2018), which may explain the lower biomass in organic vineyards. Inorganic or synthetic products affect the abundance of diverse arthropod taxa to various extents (Nash et al. 2010; Vogelweith and Thiéry 2018), thereby favoring species that are less sensitive or less exposed to fungicides during applications over others and alter community composition and species interactions. This in turn may also enhance or reduce natural pest control (Nash et al. 2010; Reiff et al. 2021a). Along with hypothesis 3a, where we predicted community composition to differ between management types, this is reflected in our study results. Arthropod communities differed between organic and conventional management, even though there was a high overlap in community composition over the two seasons.
In contrast to the second parts of our hypotheses (H1b, H2b & H3b) that predicted higher arthropod biomass and richness in vineyards with FRG varieties and different community composition compared to classic ones, biomass and taxa richness did not increase in FRG varieties and communities did not differ compared to classic varieties. Although FRG varieties had less than half as many fungicide applications and the overall pesticide toxicity was lower in those vineyards (Pedneault and Provost 2016), it seems that there was no clear effect on the arthropod community captured by our malaise traps. Yet, the richness of Hymenoptera tended to be 9% higher compared to classic varieties. Even though no clear effects were found in our study, positive effects of FRG are reported in studies that sampled the grapevine canopy (Pennington et al. 2017, 2019; Reiff et al. 2021a) as well as on grasshoppers in the ground vegetation (M. Kaczmarek, unpublished data). Thus, it is highly likely that the cultivation of grapevine varieties with resistance traits to major fungal diseases may still be an important way to reduce the impact of pesticides on non-target organisms. In addition to the lower pesticide toxicity, differences in richness between FRG and classic grape varieties may also be caused by the lower number of pesticide applications with fewer tractor passages. To date research to this topic is sparse and limited (Pedneault and Provost 2016; Pennington et al. 2017, 2019; Reiff et al. 2021b), and consequently more studies are needed to better understand the effects of reduced pesticide use and disturbance in vineyards on biodiversity.
Similar to some earlier studies on biodiversity in viticulture, the effects of the type and the amount of pesticides, in particular fungicides, were rather weak on the arthropod community (Döring et al. 2019; Paiola et al. 2020). Besides the assumption that differences in pesticide use rather change community composition than species numbers, vineyards of our region are often small and neighboring vineyards are frequently managed differently. Species may therefore not only be affected by management practices in the focal vineyard itself, but also by pesticide drift from adjacent fields (Druart et al. 2011). This may explain the rather weak effect of pesticides in the present study. In addition, we captured mainly flying insects with our Malaise traps. The highly mobile species that are predominantly caught in these traps may be less affected because they may not be as exposed to pesticides during applications as species living only in grapevine canopies. Further, about two-thirds of captured species belonged to the highly mobile orders Diptera and Hymenoptera. Due to their mobility, effects of pesticides are likely to be less evident compared to less mobile species as species diversity and density tend to converge between differently managed vineyards. Similarly, the capturing of mostly flying species may explain why we did not find stronger effects of organic management or the local vegetation on arthropods. Earlier studies reported positive effects of organic management and local vegetation for ground-dwelling and less mobile species such as spiders, beetles, and mites (Ostandie et al. 2021; Reiff et al. 2021b; Blaise et al. 2022).
The proportion of SNH near vineyards had a strong influence on arthropod diversity. Consistent with hypothesis 2c and 3c and contrary to 1c, where we predicted that SNH-rich landscapes have higher arthropod richness and different community composition but also have higher biomass, more species were found in SNH-rich landscapes and community composition changed with changing landscape composition, while we did not find any effect on biomass. Landscape structure is a major determinant of biodiversity patterns (Tscharntke et al. 2012). SNH-rich landscapes provide shelter and habitats for overwintering, as well as resources for feeding and reproduction to numerous species that are unable to complete their life cycle in agricultural fields alone (Holland et al. 2017). Positive effects of SNH have therefore also been found in previous studies for various taxonomic groups (Martin et al. 2019; Kolb et al. 2020; Tscharntke et al. 2021). In addition, SNH-rich landscapes may favor parasitoids and predators, thereby improving natural pest control (Holland et al. 2017).
In the second year of sampling, fewer arthropods were recorded, both in terms of biomass and richness. Arthropod activity, particularly of flying insects, is influenced by meterological conditions with generally higher activity during warmer and drier weather (Goodwin et al. 2021; Kaczmarek et al. 2022). The year 2021 was overall colder and wetter than 2020 (Agrarmeteorologie Rheinland-Pfalz 2022). Therefore, differences in biomass and richness between the two sampling years can be primarily attributed to differences in species activities. Similar to Goodwin et al. (2021), we observed the largest change in Hymenoptera, where only about half as many species were recorded in 2021 compared to the previous year, while Diptera and Coleoptera were less affected. Also community composition differed strongly, with less than half of the BINs being recorded over both years. Hence, the large differences in biomass, richness, and composition between the two sampling years highlights the importance of environmental conditions on the number and community of species recorded (Williams 1961). However, the fact that less than half of the BINs are recorded in both years is also a consequence of a significant proportion of singleton species in metabarcoding studies, with high numbers of species recorded exclusively at single sites (Steinke et al. 2021; Kaczmarek et al. 2022).
Conclusions
Although targeted by agro-environmental policy, the conversion to organic farming may not be appropriate to promote arthropod diversity in viticulture in our region as it reduces their biomass. This may be the result of different types of pesticides used in organic and conventional management but could also be related to differences in tillage and ground cover management. Therefore, measures such as those under the European Green Deal should be implemented in viticulture with caution, as we have observed biodiversity effects deviating from other crop** systems and from viticultural areas with insecticide use. Arthropod species richness may rather be promoted by a more diverse landscape with a higher proportion of semi-natural habitats in the surrounding area. This result suggests that suitable Ecological Focus Areas such as hedgerows should be promoted in vineyard landscapes. We only found a small positive effect of pesticide reduction on Hymenoptera diversity. The cultivation of FRG varieties and thereby a limited use of fungicides, however, may still be an important approach, as it had positive effects on less mobile or more pesticide-exposed species in other studies (Pennington et al. 2017, 2019; Reiff 2021a). However, as management practices alter community composition, the extent to which conversion to organic farming and reduction of pesticide use promote or diminish the occurrence of species of conservation concern should be studied more closely in the future. Further, it is important to consider the role of beneficial insects and pests in natural pest control and how they may be affected by these changes.
Data Availability
All data are included in the article or supplementary material.
References
Bartoń K (2020) MuMIn: Multi-Model Inference. R package version 1.43.17. https://cran.r-project.org/web/packages/MuMIn/index.html
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67(1):1–48. https://doi.org/10.18637/jss.v067.i01
Beaumelle L, Giffard B, Tolle P, Winter S, Entling MH, Benítez E, Zaller JG, Auriol A, Bonnard O, Charbonnier Y, Fabreguettes O, Joubard B, Kolb S, Ostandie N, Reiff JM, Richart-Cervera S, Rusch A (2023) Biodiversity conservation, ecosystem services and organic viticulture: a glass half-full. Agric Ecosyst Environ 351:108474. https://doi.org/10.1016/j.agee.2023.108474
Beck HE, Zimmermann NE, McVicar TR, Vergopolan N, Berg A, Wood EF (2018) Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci Data 5:180214. https://doi.org/10.1038/sdata.2018.214
Bengtsson J, Ahnström J, Weibull A-C (2005) The effects of organic agriculture on biodiversity and abundance: a meta-analysis. J Appl Ecol 42(2):261–269. https://doi.org/10.1111/j.1365-2664.2005.01005.x
Blaise C, Mazzia C, Bischoff A, Millon A, Ponel P, Blight O (2022) Vegetation increases abundances of ground and canopy arthropods in Mediterranean vineyards. Sci Rep 12(1):3680. https://doi.org/10.1038/s41598-022-07529-1
Caprio E, Nervo B, Isaia M, Allegro G, Rolando A (2015) Organic versus conventional systems in viticulture: comparative effects on spiders and carabids in vineyards and adjacent forests. Agric Syst 136:61–69. https://doi.org/10.1016/j.agsy.2015.02.009
deWaard JR, Levesque-Beaudin V, deWaard SL, Ivanova NV, McKeown JTA, Miskie R et al (2019) Expedited assessment of terrestrial arthropod diversity by coupling malaise traps with DNA barcoding. Genome 62(3):85–95. https://doi.org/10.1139/gen-2018-0093
Döring J, Collins C, Frisch M, Kauer R (2019) Organic and biodynamic viticulture affect biodiversity and properties of vine and wine: a systematic quantitative review. Am J Enol Vitic 70(3):221–242. https://doi.org/10.5344/ajev.2019.18047
Druart C, Millet M, Scheifler R, Delhomme O, Raeppel C, de Vaufleury A (2011) Snails as indicators of pesticide drift, deposit, transfer and effects in the vineyard. Sci Total Environ 409(20):4280–4288. https://doi.org/10.1016/j.scitotenv.2011.07.006
European Commission (2019) The european Green Deal. Communication from the commission. Brussels, Belgium
Fox J, Weisberg S (2019) An R companion to applied regression. SAGE Publications, Inc, Thousand Oaks, California
Goodwin EK, Rader R, Encinas-Viso F, Saunders ME (2021) Weather conditions affect the visitation frequency, richness and detectability of insect flower visitors in the australian alpine zone. Environ Entomol 50(2):348–358. https://doi.org/10.1093/ee/nvaa180
Hallmann CA, Sorg M, Jongejans E, Siepel H, Hofland N, Schwan H et al (2017) More than 75% decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 12(10):e0185809. https://doi.org/10.1371/journal.pone.0185809
Hausmann A, Segerer AH, Greifenstein T, Knubben J, Morinière J, Bozicevic V et al (2020) Toward a standardized quantitative and qualitative insect monitoring scheme. Ecol Evol 10(9):4009–4020. https://doi.org/10.1002/ece3.6166
Hochkirch A (2016) The insect crisis we can’t ignore. Nature 539(7628):141. https://doi.org/10.1038/539141a
Hole DG, Perkins AJ, Wilson JD, Alexander IH, Grice PV, Evans AD (2005) Does organic farming benefit biodiversity? Biol Conserv 122(1):113–130. https://doi.org/10.1016/j.biocon.2004.07.018
Holland JM, Douma JC, Crowley L, James L, Kor L, Stevenson DR et al (2017) Semi-natural habitats support biological control, pollination and soil conservation in Europe. A review. Agron Sustain Dev. https://doi.org/10.1007/s13593-017-0434-x
Kaczmarek M, Entling MH, Hoffmann C (2022) Using malaise traps and metabarcoding for biodiversity assessment in vineyards: Effects of weather and trap** effort. Insects 13(6):507. https://doi.org/10.3390/insects13060507
Kassambara A (2020) ggpubr: ‘ggplot2’ based publication ready plots. R package version 0.4.0. https://cran.r-project.org/package=ggpubr
Kolb S, Uzman D, Leyer I, Reineke A, Entling MH (2020) Differential effects of semi-natural habitats and organic management on spiders in viticultural landscapes. Agric Ecosyst Environ 287:106695. https://doi.org/10.1016/j.agee.2019.106695
Korner-Nievergelt F (2015) Bayesian data analysis in ecology using linear models with R, bugs, and Stan: including comparisons to frequentist statistics. Academic Press, Amsterdam
Martin M (2011) Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J 17(1):10. https://doi.org/10.14806/ej.17.1.200
Martin EA, Dainese M, Clough Y, Báldi A, Bommarco R, Gagic V et al (2019) The interplay of landscape composition and configuration: new pathways to manage functional biodiversity and agroecosystem services across Europe. Ecol Lett 22(7):1083–1094. https://doi.org/10.1111/ele.13265
Morinière J, Cancian de Araujo B, Lam AW, Hausmann A, Balke M, Schmidt S et al (2016) Species identification in malaise trap samples by DNA barcoding based on NGS technologies and a scoring matrix. PLoS ONE 11(5):e0155497. https://doi.org/10.1371/journal.pone.0155497
Möth S, Walzer A, Redl M, Petrović B, Hoffmann C, Winter S (2021) Unexpected effects of local management and landscape composition on predatory mites and their food resources in vineyards. Insects. https://doi.org/10.3390/insects12020180
Nash MA, Hoffmann AA, Thomson LJ (2010) Identifying signature of chemical applications on indigenous and invasive nontarget arthropod communities in vineyards. Ecol Appl 20(6):1693–1703. https://doi.org/10.1890/09-1065.1
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D et al (2020) vegan: Community ecology package (Version 2.5-7). https://cran.r-project.org/package=vegan
Oracle (2017) Oracle Database 12c. https://www.oracle.com/database/index.html
Ostandie N, Giffard B, Bonnard O, Joubard B, Richart-Cervera S, Thiéry D et al (2021) Multi-community effects of organic and conventional farming practices in vineyards. Sci Rep 11(1):11979. https://doi.org/10.1038/s41598-021-91095-5
Paiola A, Assandri G, Brambilla M, Zottini M, Pedrini P, Nascimbene J (2020) Exploring the potential of vineyards for biodiversity conservation and delivery of biodiversity-mediated ecosystem services: a global-scale systematic review. Sci Total Environ 706:135839. https://doi.org/10.1016/j.scitotenv.2019.135839
Pedneault K, Provost C (2016) Fungus resistant grape varieties as a suitable alternative for organic wine production: benefits, limits, and challenges. Sci Hortic 208:57–77. https://doi.org/10.1016/j.scienta.2016.03.016
Pennington T, Kraus C, Alakina E, Entling MH, Hoffmann C (2017) Minimal pruning and reduced plant protection promote predatory mites in grapevine. Insects 8(3):86. https://doi.org/10.3390/insects8030086
Pennington T, Kolb S, Kaiser J, Hoffmann C, Entling MH (2019) Does minimal pruning and reduced fungicide use impact spiders in the grapevine canopy? J Arachnol 47(3):381. https://doi.org/10.1636/0161-8202-47.3.381
Pertot I, Caffi T, Rossi V, Mugnai L, Hoffmann C, Grando MS et al (2017) A critical review of plant protection tools for reducing pesticide use on grapevine and new perspectives for the implementation of IPM in viticulture. Crop Prot 97:70–84. https://doi.org/10.1016/j.cropro.2016.11.025
R Core Team (2021) R: a language and environment for statistical computing (Version 4.1.2). R Foundation for Statistical Computing, Vienna, Austria. https://www.r-project.org/
Ratnasingham S, Hebert PDN (2013) A DNA-based registry for all animal species: the barcode index number (BIN) system. PLoS ONE 8(7):e66213. https://doi.org/10.1371/journal.pone.0066213
Reiff JM, Ehringer M, Hoffmann C, Entling MH (2021a) Fungicide reduction favors the control of phytophagous mites under both organic and conventional viticulture. Agric Ecosyst Environ 305:107172. https://doi.org/10.1016/j.agee.2020.107172
Reiff JM, Kolb S, Entling MH, Herndl T, Möth S, Walzer A et al (2021b) Organic farming and cover-crop management reduce pest predation in austrian vineyards. Insects 12(3):220. https://doi.org/10.3390/insects12030220
Rognes T, Flouri T, Nichols B, Quince C, Mahé F (2016) VSEARCH: a versatile open source tool for metagenomics. PeerJ 4:e2584. https://doi.org/10.7717/peerj.2584
RStudio T (2022) RStudio: Integrated Development for R (Version 2022.07.1). RStudio, Inc, Boston, MA. http://www.rstudio.com/
Sánchez-Bayo F, Wyckhuys KA (2019) Worldwide decline of the entomofauna: a review of its drivers. Biol Conserv 232:8–27. https://doi.org/10.1016/j.biocon.2019.01.020
Seibold S, Gossner MM, Simons NK, Blüthgen N, Müller J, Ambarlı D et al (2019) Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 574(7780):671–674. https://doi.org/10.1038/s41586-019-1684-3
Skvarla MJ, Larson JL, Fisher JR, Dowling APG (2021) A review of terrestrial and canopy malaise traps. Ann Entomol Soc Am 114(1):27–47. https://doi.org/10.1093/aesa/saaa044
Steinke D, Braukmann TWA, Manerus L, Woodhouse AW, Elbrecht V (2021) Effects of Malaise trap spacing on species richness and composition of terrestrial arthropod bulk samples. Metabarcoding Metagenom 5:43–50
Tscharntke T, Tylianakis JM, Rand TA, Didham RK, Fahrig L, Batáry P et al (2012) Landscape moderation of biodiversity patterns and processes - eight hypotheses. Biol Rev Camb Philos Soc 87(3):661–685. https://doi.org/10.1111/j.1469-185X.2011.00216.x
Tscharntke T, Grass I, Wanger TC, Westphal C, Batáry P (2021) Beyond organic farming - harnessing biodiversity-friendly landscapes. Trends Ecol Evol 36(10):919–930. https://doi.org/10.1016/j.tree.2021.06.010
Vogelweith F, Thiéry D (2018) An assessment of the non-target effects of copper on the leaf arthropod community in a vineyard. Biol Control 127:94–100. https://doi.org/10.1016/j.biocontrol.2018.08.011
Wickham H (2016) ggplot2: elegant graphics for data analysis, 2nd edn. Springer International Publishing, Cham
Wickham H, François R, Henry L, Müller K (2022) dplyr: A grammar of data manipulation. R package version 1.0.8. https://cran.r-project.org/package=dplyr
Williams CB (1961) Studies in the effect of weather conditions on the activity and abundance of insect populations. Philosophical Trans Royal Soc Lond Ser B Biol Sci 244(713):331–378. https://doi.org/10.1098/rstb.1961.0011
Winkler KJ, Viers JH, Nicholas KA (2017) Assessing ecosystem services and multifunctionality for vineyard systems. Front Environ Sci. https://doi.org/10.3389/fenvs.2017.00015
Agrarmeteorologie Rheinland-Pfalz (2022) Weatherstation Siebeldingen: Mean values 2008–2020. https://www.wetter.rlp.de/Internet/AM/NotesAM.nsf/amweb/752dce127578072cc12572eb0020c16c?OpenDocument&TableRow=2.1.2,2.6#2.1. Accessed 1 December 2022
Acknowledgements
We thank the winegrowers for their cooperation and permission to work on their properties. We also thank our colleagues at Julius Kühn Institute and RPTU Kaiserslautern-Landau for their help in field and laboratory work and for their support during the study. We acknowledge Peter Horney for providing the landscape data and AIM - Advanced Identification Methods GmbH for performing the metabarcoding. We are grateful to the Struktur- und Genehmigungsdirektion Süd for granting an exemption for the taking of wild animals of specially protected species for scientific purposes.
Funding
Open Access funding enabled and organized by Projekt DEAL. The presented study was financed by Julius Kühn Institute funds and is part of the joint project ‘Farmland biodiversity Monitoring’ (MonViA) that has been funded by the German Federal Ministry of Food and Agriculture.
Author information
Authors and Affiliations
Contributions
MK, MHE, and CH conceived the ideas and designed methodology; MK collected and analyzed the data and led the writing of the manuscript under supervision of MHE and CH; CH administered the project and acquired funding. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Communicated by Jens Dauber.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kaczmarek, M., Entling, M.H. & Hoffmann, C. Differentiating the effects of organic management, pesticide reduction, and landscape diversification for arthropod conservation in viticulture. Biodivers Conserv 32, 2637–2653 (2023). https://doi.org/10.1007/s10531-023-02621-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-023-02621-y