
Abstract
Cryptogamic diversity is a reliable indicator of the state of forest ecosystems. In this study we analysed the variations in both bryophyte species richness overall and number of hemerophobic bryophyte species in Central European managed forests over a 20-year time span, based on data collected in 132 plots scattered across Poland. We tested differences in species richness among five temporal replicates, as well as among site types grouped based on elevation a.s.l., dominant tree species and stand age. The analyses revealed no significant trend in species richness across years. Meanwhile, species richness significantly increased along with elevation a.s.l., especially in broadleaved forests. No significant difference in species number between spruce and pine dominated stands emerged for mature stands, while there was a strong difference for young stands, with spruce forest hosting a much higher number of species. Species richness exhibited a slight, but not significant, increase over time in broadleaved forests, no significant variations in pine dominated stands and significant fluctuations in spruce dominated stands, yet without a significant trend. Out of the tested drivers, dominant tree species exhibited the strongest impact on species community composition. Number of hemerophobic and strongly hemerophobic species did not undergo significant variations across years either. The lack of bryophyte diversity trends highlighted in this study suggests Central European managed forests are in an equilibrium sate, maintained by the opposing effects of climate changes, on one side and of more sustainable forest management and pollutant deposition decline, from the other.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Forests of the temperate belt have undergone profound changes during the last centuries. Management, climate changes and pollutant deposition have deeply altered structure, composition and diversity of these forests (Hannah et al. 1995; Bengtsson et al. 2000; Loustau et al. 2007). In the European continent, the impact of anthropogenic disturbance has been particularly strong, because of the high density of human population and the close proximity of forested areas and inhabited ones. One straightforward way to evaluate the extent of human impact on forest ecosystems is to examine changes in diversity and composition of biological communities. This method is particularly effective when the tested organisms exhibit high responsiveness to anthropogenic disturbance, like cryptogams (Wolf 2005; Werner and Gradstein 2009).
Bryophytes are widely acknowledged to be highly sensitive to environmental changes (Newmaster and Bell 2002; Vanderpoorten and Engels 2002; Rudolphi et al. 2014) and, owing to this property, they are extensively used as indicators of the state of forest ecosystems (Frego 2007; Hofmeister et al. 2015; Mölder et al. 2015; Czerepko et al. 2021). In undisturbed conditions, forest bryoflora is rich and diversified and hosts most rare and endangered bryophyte species, whose survival is strictly linked to the preservation of these conditions (Gustafsson and Hallingback 1988; Cooper-Ellis 1998). That is because forests, by constantly supplying environmental moisture through evapotranspiration and buffering the understory from temperature extremes and water loss thanks to the shading effects of tree canopies, provide stable climatic conditions (Chen et al. 1999; Suggitt et al. 2011; von Arx et al. 2013; Frey et al. 2016), which ensure suitable habitats for poikilohydric organisms such as bryophytes. Canopy Forests also supply a variety of micro-habitats acting as microrefugia for specialized bryophytes, e.g. stumps, logs, cavities/hollows, bark furrows, uplifted root system of uprooted trees.
However, due to anthropogenic disturbance, environmental conditions in forest ecosystems have become less stable, which poses a serious threat to bryophyte diversity (Hodgetts et al. 2019). Climate changes have caused significant increase in tree evapotranspiration rates (Qu and Zhuang, 2019; Teuling et al. 2019) and the decrease in moisture content of forest soils (EEA Report 2017; Pińskwar et al. 2020). Besides, they have also extended the length of dry periods (Kundzewicz and Matczak 2012; Hänsel 2020; IPCC 2021), which threatens the less desiccation tolerant bryophyte species, such as most liverworts. Meanwhile, forest management directly affects forest structure, micro-climate and habitat heterogeneity. Although forest cover in Europe has been constantly increasing since the half of the last century (Kauppi et al. 1992; Houghton 1995), up until recently the mean age of forest stands, as well as their compositional heterogeneity, have been consistently decreasing, old-growth deciduous forests being gradually replaced by even-aged coniferous plantations, coppices or semi-natural forests (Bengtsson et al. 2000; Zerbe 2002; Paquette and Messier 2009; FAO 2011; Forest Europe 2011; West 2014). The systematic removal of large, old trees and of decaying wood determines the loss of micro-habitats for specialized bryophyte species. The paramount importance of variety and distribution of such micro-habitats in driving bryophyte diversity has been extensively described (Ross-Davis and Frego 2002; Shelley et al. 2012; Mills and MacDonlad 2004; NewMaster et al. 2005).
Pollution too contributes to bryophyte diversity decline. Since bryophytes lack a cuticle and stomata, they are more exposed to the effects of nitrogen and sulphur deposition than vascular plants (Longton 1988; Becker-Scarpitta et al. 2017). High levels of N deposition can damage bryophyte species adapted to grow under N poor conditions (Nordin et al. 2005) and facilitates the spread of synanthropic species with highly competitive strategies, which tend to quickly dominate in terms of biomass and ultimately eliminate the more sensitive and less competitive species, thus determining a decrease in bryophyte diversity (Haworth et al. 2007; Roth et al. 2013). Sulphur deposition was also shown to negatively affect sensitive bryophyte species (Farmer et al. 1991, 1992; Lee and Studholme 1992; Makipaa 1995; Sjogren 1995), although its effects are mostly hard to distinguish from those of N deposition.
In view of their sensitivity to disturbance, examination of changes in bryophyte diversity over time may thus provide relevant information about the response of forest ecosystems to anthropogenic pressure. However, not all bryophytes species exhibit high responsiveness to environmental changes. Many of them display a wide tolerance range and occur in different habitats, as well as on different substrates. Others are more demanding in their physiological requirements (Gignac 2001; Proctor et al. 2007) and are thus restricted to habitats characterized by specific micro-environmental conditions. Within this second group, those species unable to live within areas unaffected by anthropogenic disturbance are referred to as hemerophobic (Linkola 1916) or, alternatively, as primeval forest relics (Cieślinski et al. 1996) or woodland indicator species (Müller et al. 2019). Hemerophobic species are mostly associated with undisturbed habitats, shady and moist conditions and specific substrates such as logs in advanced stage of decomposition and large, old trees. These species being restricted in their occurrence to these substrates demonstrates the importance of the latter for organisms otherwise characterized by remarkable dispersal abilities such as bryophytes (Medina et al. 2011; Sundberg 2005, 2013; Szövényi et al. 2012). This is also because bryophytes occurring on the ground have to compete with vascular plants for physical space and access to resources and usually get the worst of it because of their much smaller size (Looman 1964; van Wijik et al. 2003; van der Wal et al. 2005; Walker et al. 2006; Alatalo et al. 2020), so that availability of substrates generally unsuitable for vascular plants (such as tree bark and dead-wood) represents a constraining factor for many bryophyte species, especially for liverworts and tiny mosses.
However, bryophytes also occur in managed forests, provided they can find suitable micro-environmental conditions. Czerepko et al. (2021) found that, while most bryophyte species did not exhibit a significant response to the gradient of forest naturalness expressed by different management regimes and protection time spans, red-listed or primeval forest relics showed a twice higher frequency in undisturbed, close to pristine forest habitats and nature reserves than in managed forests. By analysing patterns of diversity and dynamics of hemerophobic species richness, we may thus obtain a more reliable information about the response and adaptation of forests to anthropogenic disturbance than examining bryophyte diversity overall.
Managed forests make up 95% of the forest area in Europe (Forest Europe 2020), therefore knowledge about the dynamics of bryophyte diversity in these areas is essential to evaluate the state of bryophyte diversity in the European continent. So far, most studies about bryophyte diversity in forest ecosystems have focused on the difference in diversity patterns among areas characterized by different forest types, climate and management regimes, usually comparing two or a few sites (Humphrey et al. 2002; Hofmeister et al. 2015; Müller et al. 2019; Czerepko et al. 2020). Much less is known about temporal changes in forest bryophyte diversity across large areas. Dittrich et al. (2016) examined changes in bryophyte diversity in the German forests since the beginning of the nineteenth century, but their study involved only two temporal replicates and focused just on beech forests. Becker-Scarpitta et al. (2017), instead, followed changes in bryophyte diversity over a thirty-year time span, but with more replicates. However, these changes were monitored at just one forest site. Lastly, Baumann et al. (2021) monitored changes in ground bryophytes over three decades, but they also focused only on one forest type, i.e. spruce forests. Furthermore, none of these studies examined changes in bryophyte diversity depending on forest site features.
In this study, we aimed at highlighting changes in bryophyte species richness and number of hemerophobic species in forest sites across Poland during a 20-year-life-span, including five replicates, thus combining both spatial and temporal analyses. Unlike in Dittrich et al. (2016) and Becker-Scarpitta et al. (2017), these study sites encompass different forest types, which were tested separately. Changes in species richness and number of hemerophobic species were analysed for all sites together, as well as for different site clusters based on dominant tree species, stand age and elevation a.s.l. The study was based on data collected between 1998 and 2019 from ICP Forest level II plots.
In particular, we wanted to test the hypotheses that: (1) species richness of bryophytes overall has decreased over time due to increasing dry conditions in Polish forests and the long-term impact of forest management; (2) the highest species richness is to be found at higher elevations, in oldest forests and in broadleaved stands; (3) the number of hemerophobic species decreased over time and this decreased was stronger than the decrease of bryophyte species overall, hemerophobic species being more sensitive to environmental changes than generalists.
Materials and methods
Study site
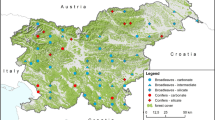
This study was based on data collected in 132 plots scattered across Poland (Fig. 1), established in the frame of the ICP Forests Monitoring Level II plots and ranging between 11 to 991 m a.s.l. The monitoring network originally included 148 study plots (Fig. 1), but areas where no monitoring was carried out following stand breakdown caused by bark beetle infestation or windstorms were excluded from the study.

Distribution of ICP Forests Level II plots in Poland
The Polish climate is predominantly continental, with an average annual temperature of 8.7 °C in the period 1991–2020 and an average annual precipitation of about 609 mm (Klimat Polski 2021). Precipitation varies greatly from region to region, especially between mountains and lowlands, ranging from over 1300 mm in the Sudetes and Carpathians to about 500 mm in the central part of the country (Wielkopolska, Mazovia) (Kluziński 2021). The forest site types encompassed by the study areas can grouped into oligotrophic (mixed-coniferous—33%, coniferous—26%) and and mesotrophic (deciduous 20%, mixed deciduous 21%). The dominant tree species on the plots was Pinus sylvestris (73% of the total surface) followed by by Picea abies (8–10%), Quercus robur and Fagus sylvatica. The mean age of stands at the beginning of monitoring (1998) was 61 years old, ranging between 45 and 96. Most sample plots had a stand age below 80 years, and only the 7% of them was older. All sampled plots were under forest management, selective thinning being carried out every 10–20 years, variations within this range depending on the main tree species, with a percentage of wood volume removed amounting to ca. 6%.
Sampling design
At each of the 132 monitoring plots, sampling was carried out in a 20 × 20 m square plot, according to the European protocol for monitoring of forest areas (Stofer et al. 2016). Bryophytes were sampled on living trees up to a height of 2 m, standing and lying dead wood, ground and rocks. Cover relative to the area of the substrate was assigned based on the protocol proposed by Solon and Wawrzoniak (1999), however, to the purposes of this work, only presence absence data were employed, as it is often the case when assemblages growing on different substrates are examined. In fact, since most recorded species occurred in the same plots on different substrates, calculating the mean cover of these species per plot based on covers recorded on different substrates would be methodologically questionable. Sampling was carried out in 1998–1999, 2003, 2008, 2013, 2019.
Out of the whole set of species recorded in the survey, hemerophobic species were identified based on Dierßen (2001) and include both ahemerophobic and oligohemerophobic species. Analyses of changes in species richness were performed both based on the whole community and on the three separate groups identified based on level of hemerophoby: non-hemerophobic (NH), hemerophobic (N) and strongly hemerophobic (SH).
Nomenclature of bryophyte species follows Hodgetts et al. (2020).
Statistical analyses
In order to evaluate the degree of overlap in the sets of species recorded per each year of the survey and thus infer the potential weight of the observer bias, we analysed the trend of the error in measuring the multivariate dispersion of the community with increasing sampling effort by using the Jaccard index. Overall differences in species richness and number of NH, H and SH species across years was tested using Generalized Linear Models (GLM) with a quasi-poisson family error (data being overdispersed). Posthoc multiple comparisons were performed using least-squares means and Holm correction using “emmeans” R package. (Lenth 2022a, b). The variation of bryophyte species richness as a function of elevation and the first-order interaction between stand age (2 levels: M—Mature, Y—Young) and stand type (3 levels: Broadleaved—B, Pine—P, and Spruce—S) was assessed using Generalized Linear Mixed Models (GLMMs) and Poisson error family. Specifically, stand age and stand type were treated as fixed effects while temporal autocorrelation among observations was accounted for using Years as random effect. Model’s R2 was computed by means of a standardized generalized variance approach using “r2glmm” R package (Jaeger 2017). Since stands dominated by Q. robur and F.sylvatica (B) were all over 60 years old, it was not possible to test the interaction between stand ages and dominant tree species within this level which was excluded from this analysis.
The Variation Partitioning approach (Borcard et al. 1992; Økland 2003; Legendre 2008) was used on each group of variables to assess the proportion of compositional variation explained by their unshared and shared effects. Patterns of species compositional variation in samples and their environmental correlates were further explored using a Redundancy Analysis (Legendre and Legendre 2012).
The trend of the error in measuring the multivariate dispersion of the community with increasing sampling effort was obtained by using the SSP R package (Anderson and Santana-Garcon 2015). GLMMs were performed using “lmerTest” and ‘lme4″ R packages (Kuznetsova et al. 2017; Bates et al. 2015), while effect plots were obtained using “effects” R package (Fox and Weisberg 2019). Variation Partitioning and RDA was performed using the vegan package (Oksanen et al. 2020). All analyses were carried out in the R software 4.1.1 (R Core Team 2021).
Results
Overall, 132 species were recorded, out of which 110 mosses and 22 liverworts (Appendix 1). The total number of species per year ranged between 84 in 1998/1999 and 98 in 2008, while the average number of species per plot ranged between 8 and 9, being the highest in 2008 and the lowest in 2003.
The observations appear relatively robust to the observer bias, no strong deviations in the set of recorded species among different years and in the relative error being observed (Appendix 2).
GLMM exhibited a good explanatory power (R2 = 0.12). No significant variation in species richness across years was highlighted by the model, neither for bryophytes overall, nor for mosses and liverworts separately (Fig. 2). The only significant difference in species richness among years was found between 2003 and 2008 (Table 1).

GLM model showing the differences in species richness among years for bryophytes overall, liverworts and mosses
Species richness stands exhibited a slight, but not significant, increase in broadleaved forests, no significant variations in pine dominated stands and significant fluctuations in spruce dominated stands, yet without a significant trend (Fig. 3). Since the first years of observations the number of species in broadleaved forest stands increased relative to the number of species in stands dominated by Pinus and Picea, until it exceeded the number of species in Picea dominated stands in 2013. Overall, the highest species richness was recorded in stands dominated by Picea abies, and the lowest in those dominated by Pinus sylvestris.

GLMM model showing the interaction between stand age and dominant tree species controlling for the year. Broadleaved forest stands were excluded from the model since they had no stands younger than 60 years. B broadleaved, P pine, S spruce
We observed a positive relationship between bryophyte species richness and elevation (Table 2) which was driven by broadleaved stands (Fig. 4). We also detected a significant interaction between dominant tree species and stand age (Table 2). Surprisingly, no significant difference in species richness between spruce and pine dominated stands emerged for mature stands, while a strong difference was observed for young stands (Table 3). While pine dominated stands the number of species was slightly higher in older than younger stands, in spruce dominated forests it was the opposite, with younger stands exhibiting a significantly higher number of species than old ones (Fig. 5). Fluctuations in the number of species per year were stronger in spruce dominated forests. Without distinction among dominant tree species, the highest number of species was recorded in older forest stands.

Relationship between species richness and elevation a.s.l for broadleaved (blue), pine (purple) and spruce (green) forest stands

Interaction between stand age and forest stand type. P pine, S spruce, M mature stands, Y young stands
Hemerophobic species amounted to 66, while strongly hemerophobic to 13 (Supplementary Materials 2) and neither group showed any significant variation over time (Fig. 6).

GLM model showing the variations over time of strongly hemerophobic (SH), hemerophobic (H) and not hemerophobic (NH) species. No significant trend emerged
Variation Partitioning explained around 10% of the total variation in community composition, both for the overall community and for its sub-groups (NH, H, SH), and showed that the main driver of the community is the dominant tree species, followed by elevation a.s.l. (Fig. 7).

Variation Partitioning of: a the whole community, b non hemerophobic species, c strongly hemerophobic species, d hemerophobic species
The RDA model explained 23% of the total variation in the community composition (Fig. 8). All axes were significant (Table 4). The main environmental predictor was elevation a.s.l., which correlated with species richness of both SH and NH. Interestingly, the latter two turned out to be strongly correlated with each other.

RDA of the bryophyte community and the set of biotic and abiotic factors. Continuous variables (elevation a.s.l. and richness of SH, H and NH) are reported in blue, while categorical variables (stand age, dominant tree species) are in green. Species are reported in red
Discussion
The results of this study show that, contrary to our first hypothesis, bryophyte species richness in central European managed forests has remained quite stable during the last twenty years. This means the environmental changes these forests have undergone during the last decades were not such dramatic as to determine significant changes in their bryoflora. The lack of a significant diversity trend is likely to be the result of the opposing effects of climate changes, from one side, and more sustainable forest management and pollutant deposition decline, from the other. In fact, while climate changes are expected to determine a decrease in species richness by affecting metabolic activity, growth and post-desiccation recovery capacity of the less desiccation tolerant species (Gignac 2001, 2011; Proctor 2011; Song et al. 2012; He et al. 2016; Hao and Chu 2021), both the recent advancements in forestry practices towards a more sustainable model and the pollutant deposition decline are likely to bring about an increase in bryophyte diversity, the first by ensuring the retention of increasing amounts of dead-wood and large trees in forests, the second by allowing restoration of suitable physiochemical conditions for the pollution-sensitive species.
Out of these three major anthropogenic drivers of cryptogamic diversity, the impact of climate change on bryophyte diversity is the less straightforward to evaluate. In fact, while studies showed a steady increase in mean and summer temperatures and the lengthening of dry periods due to decrease of precipitation in the summer period and their shift to the winter period (Kundzewicz and Matczak 2012; Ziernicka-Wojtaszek and Kopcinska 2020), Poland is considered to be less vulnerable to adverse climate change impacts than many other countries (Kundzewicz and Matczak 2012). Unlike for temperatures, precipitation patterns do not exhibit a clear trend and are highly variable across the country, so that while some areas of Poland exhibit a moderate decrease of total precipitation frequency and amount, others are experiencing an increase of both, especially in northern Poland (Ziernicka-Wojtaszek and Kopcińska 2020), which ultimately results in a lack of trend at country level. This makes it likely the expected negative impact of climate changes on bryophyte diversity has been so far just moderately strong.
Meanwhile, although managed forests are generally poorer in cryptogams than natural ones and lack the rarest and more threatened species, policy makers and foresters have been paying increasing attention to biodiversity conservation issues. Examples of improvements in forest management, based on research about the more sustainable forestry practices (Miller 1996; Noss 1996; Lindenmayer et al. 2012), are the retention of a minimum amount of dead wood on the forest floor, the reduction in the felling of trees over 100 years-old and the limitation in the use of clear-cuts and their substitution with partial and nested cuts, which lead to an increase in the complexity of the structure of stands, both in terms of age and species (Zasady, Kryteria i Wskaźniki Dobrej Gospodarki Leśnej w Polsce. 2010; Zasady hodowli lasu, 2012; Report on the European Forest Strategy—The Way Forward, 2019). Such practices are expected to have positively affected bryophyte diversity. In fact, according to the species sorting theory, “everything is everywhere, but the environment selects” (Baas-Becking 1934), which was proven to be true for most biological communities (Astorga et al. 2012; Cottenie 2005). Since bryophytes are generally very efficient disperses (Sundberg 2005; Shaw et al. 2011; Szövényi et al. 2012), if the main requirement for their occurrence, i.e. availability of suitable substrates, is ensured, euriecious and also several hemerophobic species may persist and withstand minor disturbance events with little damage. The advancements in forest practices introduced during the last 15–20 years likely played an important role in preventing bryophyte diversity loss, although they were not sufficiently strong to determine an increase of bryophyte diversity. In fact, though more dead wood is being left to natural decay in Polish managed forests, big logs required by many stenoecious saproxylic species, particularly liverworts, are still missing in these forests, since the latter altogether lack old, big trees, stand age being mostly below 80 years and just in a few cases exceeding 90 years.
The significant decrease of pollutant deposition observed during the last decades (Dirnböck et al. 2018; Vivanco et al. 2018) may have also contributed to the maintenance of bryophyte diversity, counterbalancing the negative effects of climate changes. However, as in the case of forest management practices, the pollution drop was not enough to ultimately determine an increase of bryophyte diversity. Meanwhile, Baumann et al. (2021) ascribed the observed increase in bryophyte species richness and cover in the spruce forests of eastern Germany just to the sharp decline in S deposition. These contrasting conclusions may be accounted for by the difference in the scale of observation. The study by Baumann et al. (2021) examined a limited area of just a few square kilometres, while our study encompassed the whole Poland, and the effects of the S deposition drop are expected to be less obvious at a larger geographical scale, given the increased variability of environmental factors involved in the response of biological communities to changes in disturbance regimes. So, in the end, the net effect of a moderate impact of climate changes, the improvement in forestry practices and a strong pollution decrease, as the three more likely drivers of bryophyte diversity variations, was a lack of trends. The only significant difference in species richness was observed between 2003 and 2008, when it markedly increased after having dropped between 1998/1999 and 2003. Since the observer bias turned out to be quite negligible, we must assume the highlighted increase in species number was determined by some major environmental change, whose effects yet disappeared in a few years. In fact, species richness dropped again between 2008 and 2013, thus returning to a level similar to that of the beginning of the survey.
The lack of significant variations in bryophyte species richness in Polish forests seems at first to supports the results by Dittrich et al. (2016), reporting no trends in overall species richness over a century, and to contrast with those by Becker-Scarpitta et al. (2017), who observed an increase in species richness over a 30-year time span, as well as with those by the above mentioned study by Baumann et al. (2021), also reporting a diversity increase in ground bryophytes. However, in our study we pooled and examined together both coniferous and broadleaved forests, while Dittrich et al. (2016) and Becker-Scarpitta et al. (2017) examined just broadleaved forests and Baumann et al. (2021) just coniferous ones. Maintaining that a reliable comparison of data across different studies should be drawn only when environmental conditions are comparable and considering that in our study a slight, though non-significant, increase in the number of species was found in broadleaved forests, while no significant trend was highlighted for spruce forests, our results are actually closer to those by Becker-Scarpitta et al. (2017) and contrast with both those by Dittrich et al. (2016) and by Baumann et al. (2021). A longer observation period could have clarified whether the slight increase in the number of species observed in broadleaved forests would ultimately generate a significant trend or not. Becker-Scarpitta et al., (2017) could not provide a conclusive explanation to the observed increase in species richness at his study site, except by hinting to a possible recovery from the peak of acid deposition in the 1970s. More favourable trends in species richness in broadleaved than in coniferous forests may be also explained by assuming these forests are more resistant to temperature variations than coniferous forests. Indeed, Schwaab et al. (2020) reported that broad-leaved tree species locally reduce land surface temperatures in summer compared to needle-leaved species, particularly during exceptionally warm periods.
Despite the clear lack of changes in bryophyte species richness highlighted by this study, it is worth remembering that the bryophyte sample analysed in this study, although vast, is still little representative of the overall state of Polish forests, as it is based on randomly chosen sites with a total area corresponding to around 65 ha, whereas the forest area in Poland amounts to 9435 ha, and hosts overall 132 species of bryophytes, representing only 18% of the Polish bryoflora, which amounts to 950 species (Ochyra et al. 2003; Szweykowski 2006).
As for diversity response to environmental drivers, pine dominated forests exhibited, as expected, the lowest species richness, which suggests that, in order to enhance bryophyte diversity in forest ecosystems, these forest types, mostly artificially planted, should be reconverted to broadleaved forests. Comparison of diversity trends in young and mature forest stands was possible only for pine and spruce forests, since broadleaved forests include only mature stands. Here we observed a surprising pattern. While there was no significant difference in the number of bryophyte species between pine and spruce forest stands when mature, a significantly higher number of species occurred in young spruce forests than in young pine forests. This contrasted with our expectations, since we assumed the difference in species richness between pine forests, generally bryophyte poor, and spruce forests, on average much richer, would be driven by the favourable micro-climatic conditions and availability of substrates typical of mature spruce stands, e.g. dead wood. In fact, mature spruce forests are characterized by shaded conditions and a high moisture level; in addition, spruces in Europe are periodically, massively attacked by the spruce bark-beetle and are, on account of their flat and superficial root system, more easily uprooted by strong winds. Because of that, the amount of dead-wood suitable for colonization by bryophytes in mature spruce forests is, where dead wood is not systematically removed, quite high, which favours epixylic bryophytes.
As expected, species richness increased with elevation, which had been already reported by several studies about changes in bryophyte diversity along altitudinal gradients in mountain areas (Zhang et al. 2021; Bruun et al. 2006). In our study the variations in bryophyte species richness were evaluated along an elevational gradient including the whole lowland-upland-mountain range, which provides a more complete picture of the response patterns of bryophyte diversity to altitude than assessments of changes occurring just within the mountain belt. The increase in bryophyte species richness along with altitude is most likely due to the reduced impact of temperature extremes and to the higher moisture retention in forests at higher altitudes. Under this regard, the patterns of bryophyte diversity contrast with the ones of vascular plants, who generally host more species at lower altitudes (Theurillat et al. 2003; Trigas et al. 2013; Gebrehiwot et al. 2019). This difference in the response to increasing elevation is due to the fundamental differences in the physiological requirements of these two plant groups. In fact, while vascular plants are favoured by warmer temperatures, which, besides causing a longer growing season, enhance photosynthetic activity and reproductive output (Rustad et al. 2001; Wu et al. 2011; Peñuelas et al. 2013), bryophytes, due to their poikilohydric nature and their reproductive mechanisms, thrive in moist, cool, shaded conditions (Marschall and Proctor 2004) These requirements characterize especially liverworts and hemerophobic species.
The proportion of variation in the community composition explained by the tested variables turned out to be quite small, which implies bryophyte composition is driven by other environmental factors and/or by stochastic processes. As for the first possibility, local climatic conditions and availability of micro-habitats are the more likely candidates. Forest areas characterized by the same forest type, stand age and elevation, may substantially differ in their climate depending on geographic location, distance from the sea, proximity of mountain relief and water basins. However, the analysis of the impact of local climatic conditions on biological diversity is usually hindered by the lack of a systematic network of climate monitoring systems and of long-term data. On the contrary, the crucial role of substrate availability for maintaining bryophyte diversity has been highlighted by several authors (Ross-Davis and Frego 2002; Shelley et al. 2012; Mills and MacDonald 2004; NewMaster et al. 2005). The role of stochastic processes in ecology is generally quite tricky to pinpoint. Despite frequently accounting for a large share of variability in community dynamics, stochasticity is not usually regarded as driver of such dynamics (Hart et al. 2017), being rather identified with unexplained variance, noise, or fundamental unpredictability (Shoemaker et al. 2020), namely with that inherent component of natural processes which sets an obstacle to the possibility of explaining natural dynamics and their underlying mechanisms (Boettiger 2018).
Out of the tested environmental factors, dominant tree species turned out to be the major driver of bryophyte community composition, followed by elevation, while stand age had the least influence, which was again in contrast with our hypothesis. Since mean stand age in managed forests is the result of the forest management regime, we may interpret the exiguous influence of stand age on species richness highlighted in this study as a scarce influence of management regimes on species richness. This seems to confirm the report by Müller et al. (2019), who found that species richness of four ecological bryophytes guilds (epigeic, epiphytic, epilxylic, epilythic) was independent of the management regimes and only responded to the availability of different substrates. Though this may at first appear surprising, it can be explained by assuming the managed forests examined by Muller et al. (2019) had a high degree of naturalness and retained all the substrates needed for forest bryophyte species to develop.
Surprisingly, the RDA revealed that plots characterized by a high number of hemerophobic species are independent from those characterized by high number of both strongly hemerophobic and non-hemerophobic, the latter two being correlated between them and with the elevation. The niche convergence of these two ecologically distant groups, marked differing in sensitivity to human disturbance, may be determined by similar requirements in terms of altitude-related environmental and climatic conditions. Meanwhile, it is worth highlighting that the mean number of SH species per plot is extremely low (maximum 2 species per plot), which makes this correlation incidental.
As in the case of overall species richness and again in contrast with our expectations, the number of hemerophobic species did not exhibit changes over time. This means not only generalist, euriecious species remained unaffected by environmental changes during the last 20 years, but also the more stenoecious species, strictly linked to the shaded and moist environment of close-canopy forests, which express the promptest response to environmental changes. This further confirms that Polish forests did not experience significant environmental stress over the last 20 years, since they retained all the more demanding bryophyte species. However, it is worth highlighting that many hemerophobic species do not occur at all in managed forests and are restricted to natural, old-growth forests characterized by high amounts of dead-wood and old trees and/or to areas characterized by the occurrence of disturbances such as windstorms or bark beetle outbreaks, that create new niches for the bryophyte flora associated with uprooted trees (Faliński et al. 1996; Staniaszek-Kik et al. 2021). These strongly disturbed areas were not examined in our study, since all study sites characterized by extensive tree stand breakdown following natural disturbance were excluded from the analyses. Examples of strongly hemerophobic species restricted to the above mentioned habitats are Dicranum viride, Buxbaumia viridis, Neckera pennata, Anomon viticulosus, Riccardia palmata, Anastrophyllum hellerianum (Czerepko et al. 2021). Those recorded in the frame of the monitoring of ICP Forest of II level plots are among the less sensitive among these typical forest species, which means they are able to cope to a certain degree with variations of forest environmental conditions, as long as the integrity of the forest environment overall is not compromised.
Conclusion
This study shows that both overall bryophyte species richness and number of hemerophobic species in Polish forests have not significantly changed during the last two decades. This lack of diversity trends at a time of strong anthropogenic changes may be the result of the general improvement of the state of forest ecosystems over the last decades, preventing a bryophyte diversity loss, and, at the same time, of the limited extent of such improvement, which could not generate a positive diversity trend. Thus, maintaining that bryophyte are reliable indicators of the state of forest ecosystems, we may conclude that Polish forests are in an equilibrium state where the expected negative effects from climate changes are still limited and are being counterbalance by the positive effects of a more responsible forest management and an improvement in air and water quality. Longer monitoring may reveal possible shifts from this momentary equilibrium state.
Data availability
The data that support the findings of this study are available from the corresponding author [Cecilia Cacciatori], upon reasonable request.
References
Alatalo JM, Jägerbrand AK, Erfanian MB et al (2020) Bryophyte cover and richness decline after 18 years of experimental warming in alpine Sweden. AoB Plants. https://doi.org/10.1093/aobpla/plaa061
Anderson MJ, Santana-Garcon J (2015) Measures of precision for dissimilarity-based multivariate analysis of ecological communities. Ecol Lett 18:66–73
Astorga A, Okansen J, Luoto M, Soininen J, Virtanen R, Muotka T (2012) Distance decay of similarity in freshwater communities: do macro- and microorganisms follow the same rules? Glob Ecol Biogeogr 21:365–375. https://doi.org/10.1111/j.1466-8238.2011.00681.x
Baas-Becking LGM (1934) Geobiologie of inleiding tot de milieukunde. The Hague, The Netherlands: W.P. van Stockum & Zoon
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67(1):1–48
Baumann M, Dittrich S, Körner M, von Oheimb G (2021) Temporal changes in the ground vegetation in spruce forests in the Erzgebirge (Ore Mountains): bryophytes are better indicators of the impact of liming and of sulphur and nitrogen deposition than the herb layer. Appl Veg Sci. https://doi.org/10.1111/avsc.12598
Becker-Scarpitta A, Bardat J, Lalanne A, Vellen M (2017) Long-term community change: bryophytes are more responsive than vascular plants to nitrogen deposition and warming. J Veg Sci 28:1220–1229. https://doi.org/10.1111/jvs.12579
Bengtsson J, Nilsson SG, Franc A, Menozzi P (2000) Biodiversity, disturbances, ecosystem function and management of European forests. For Ecol Manage 132:39–50. https://doi.org/10.1016/S0378-1127(00)00378-9
Boettiger C (2018) From noise to knowledge: how randomness generates novel phenomena and reveals information. Ecol Lett 21:1255–1267. https://doi.org/10.1111/ele.13085
Borcard D, Legendre P, Drapeau P (1992) Partialling out the spatial component of ecological variation. Ecology 73:1045–1055. https://doi.org/10.2307/1940179
Bruun HH, Moen J, Virtanen R, Grytnes J-A, Oksanen L, Angerbjörn A (2006) Effects of altitude and topography on species richness of vascular plants, bryophytes and lichens in alpine communities. J Veg Sci 17:37–46
Chen J, Saunders SC, Crow TR, Naiman RJ, Brosofske KD, Mroz GD, Brookshire BL, Franklin JF (1999) Microclimate in Forest Ecosystem and landscape ecology: variations in local climate can be used to monitor and compare the effects of different management regimes. Bioscience 49:288–297. https://doi.org/10.2307/1313612
Cooper-Ellis S (1998) Bryophytes in old-growth forests of western Massachusetts. J Torrey Botan Soc 125:117–132
Cottenie K (2005) Integrating environmental and spatial processes in ecological community dynamics. Ecol Lett 8:1175–1182. https://doi.org/10.1111/j.1461-0248.2005.00820.x
Czerepko J, Gawrys R, Szymczyk R, Pisarek W et al (2021) How sensitive are epiphytic and epixylic cryptogams as indicators of forest naturalness? Testing bryophyte and lichen predictive power in stands under different management regimes in the Białowieża Forest. Ecol Ind 125:107532. https://doi.org/10.1016/j.ecolind.2021.107532
Cieślinski S, Czyzewska K, Falinski JB, Klama H, Mułenko W, Zarnowiec J (1996) Relicts of the primeval (virgin) forest Relict phenomena, in: Falinski J.B., Mułenko W. (Eds.), Cryptogamous plants in the forest communities of Białowieza National Park (Project CRYPTO 3). Phytocoenosis 8 (N.S.). Archiv. Geobot. 6:197–216
Dierßen K (2001) Distribution, ecological amplitude and phytosociological characterization of European bryophytes. Bryophytorum Bibliotheca 56:1–289
Dirnböck T, Pröll G, Austnes K et al (2018) Currently legislated decreases in nitrogen deposition will yield only limited plant species recovery in European forests. Environ Res Lett 13:125010
Dittrich S, Leuschner C, Hauck M (2016) Change in the bryophyte diversity and species composition of Central European temperate broad-leaved forests since the late nineteenth century. Biodivers Conserv 25:2071–2091. https://doi.org/10.1007/s10531-016-1179-6
European Environmental Agency (EEA) Report 1/2017. Climate change, impacts and vulnerability in Europe 2016-An indicator-based report. https://doi.org/10.2800/534806
Faliński JB, Mułenko W, Żarnowiec J, Klama H, Głowacki Z, Załuski T (1996) The colonisation of fallen tree sites by green plants and fungi. In: Faliński JB, Mułenko W (eds) Cryptogamous plants in the forest communities of Białowieża National Park Phytocoenosis 8 (N.S.). Arch Geobotan 6:147–150
FAO (Food and Agriculture Organization of the United Nations) (2010) Global forest resources assessment 2010. Food and Agriculture Organization of the United Nations, Rome
Farmer AM, Bates JW, Bell JNB (1991) Seasonal variations in acidic pollutant inputs and their effects on the chemistry of stemflow, bark and epiphyte tissues in three oak woodlands in N.W. Britain New Phytologist 118:441–451
Farmer AM, Bates JW, Bell JNB (1992) Ecophysiological effects of acid rain on bryophytes and lichens. In: Bates JW, Farmer AM (eds) Bryophytes and lichens in a changing environment. Claredon Press, Oxford
Forest Europe (2020) State of Europe’s Forests 2020. Status and Trends in Sustainable Forest Management in Europe. Forest Europe, UNECE and FAO, Oslo
Forest Europe, UNECE, FAO (2011) State of Europes forests 2011. Status and trends in sustainable forest management in Europe. Forest Europe, UNECE and FAO, Oslo
Fox J (2003) Effect displays in R for generalised linear models. J Stat Softw 8:1–27
Fox J, Weisberg S (2019) An R companion to applied regression, Third edition. Sage, Thousand Oaks CA. https://socialsciences.mcmaster.ca/jfox/Books/Companion/
Frego K (2007) Bryophytes as potential indicators of forest integrity. For Ecol Manage 242(1):65–75. https://doi.org/10.1016/j.foreco.2007.01.030
Frey SJK, Hadley AS, Johnson SL, Schulze M, Jones JA, Betts MG (2016) Spatial models reveal microclimatic buffering capacity of old-growth forests. Sci Adv 2:e1501392
Gebrehiwot K, Demissew S, Woldu Z, Fekadu M, Desalegn T, Teferi E (2019) Elevational changes in vascular plants richness, diversity, and distribution pattern in Abune Yosef mountain range, Northern Ethiopia. Plant Diversity 41:220–228. https://doi.org/10.1016/j.pld.2019.06.005
Gignac LD (2001) Bryophytes as indicators of climate change. The Bryologist 104:410–420
Gignac LD (2011) Bryophytes as predictors of climate change. In: Tuba Z, Slack N, Stark LR (eds) Bryophyte ecology and climate change. Cambridge University Press, Cambridge, pp 461–482
Gustafsson L, Hallingback T (1988) Bryophyte flora and vegetation of managed and virgin coniferous forests in South-West Sweden. Biol Cons 44:283–293
Hannah L, Carr JL, Landerani A (1995) Human disturbance and natural habitat-a biome level analyses of a global data set. Biodivers Conserv 4:128–155
Hänsel S (2020) Changes in the characteristics of dry and wet periods in Europe (1851–2015). Atmosphere 11:1080. https://doi.org/10.3390/atmos11101080
Hao J, Chu LM (2021) Short-term detrimental impacts of increasing temperature and photosynthetically active radiation on the ecophysiology of selected bryophytes in Hong Kong, southern China. Global Ecology and Conservation 31:e01868. https://doi.org/10.1016/j.gecco.2021.e01868
Hart SP, Usinowicz J, Levine JM (2017) The spatial scales of species coexistence. Nature Ecol Evol 1:1066–1073. https://doi.org/10.1038/s41559-017-0230-7
Haworth BJ, Ashmore MR, Headley AD (2007) Effects of nitrogen deposition on bryophyte species composition of calcareous grasslands. In: Brimblecombe P, Hara H, Houle D, Novak M (eds) Acid rain: deposition to recovery. Springer, Dordrecht
He X, He KS, Hyvönenc J (2016) Will bryophytes survive in a warming world? Systematics 19:49–60. https://doi.org/10.1016/j.ppees.2016.02.005
Hodgetts NG, Calix M, Englefield E, Fettes N et al (2019) A miniature world in decline European Red List of Mosses Liverworts and Hornworts. https://doi.org/10.2305/IUCN.CH.2019.ERL.2.en
Hodgetts NG, Söderström L, Blockeel TL, Caspari S et al (2020) An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J Bryol 42:1–116. https://doi.org/10.1080/03736687.2019.1694329
Hofmeister J, Hosek J, Brabec M, Dvorak D, Beran M, Deckerova H et al (2015) Value of old forest attributes related to cryptogam species richness in temperate forests: a quantitative assessment. Ecol Ind 57:497–504. https://doi.org/10.1007/s10531-015-0863-2
Houghton RA (1995) Changes in the storage of terrestrial carbon since 1850. In: Lal R, Kimble J, Levine E, Stewart BA (eds) Soils and global change. CRC Press Inc, Boca Raton
Humphrey JW, Davey S, Peace AJ, Ferris R, Harding K (2002) Lichens and bryophyte communities of planted and semi-natural forests in Britain: the influence of site type, stand structure and deadwood. Biol Cons 107:165–180
IPCC (2021) Summary for policymakers. In: Climate change 2021: the physical science basis. Contribution of Working Group I to the sixth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge
Jaeger B (2017) r2glmm: computes R squared for mixed (multilevel) models. R package version 0.1.2. https://CRAN.R-project.org/package=r2glmm
Kauppi P, Mielikainen K, Kuusela K (1992) Biomass and ¨carbon budget of European forests, 1971 to 1990. Science 256:70–74
Klimat P (2021) Klimat Polski 2020. Instytut Meteorologii i GospodarkiWodnej – Państwowy Instytut Badawczy, Warszawa.
Kluziński L (2021) Warunki pogodowe w 2020 roku na wybranych powierzchniach SPO MI. In: P Lech (ed) Stan zdrowotny lasów w Polsce w 2020 roku na podstawie badań monitoringowych. Instytut Badawczy Leśnictwa, Sękocin Stary, pp 71–75. https://www.gios.gov.pl/monlas/raporty/raport_SUL_2020.pdf
Kundzewicz ZW, Matczak P (2012) Climate change regional review: Poland. Wires Clim Change 3:297–311. https://doi.org/10.1002/wcc.175
Kuznetsova A, Brockhoff PB, Christensen RHB (2017) lmerTest package: tests in linear mixed effects models. J Stat Softw 82(13):1–26
Lee JA, Studholme CJ (1992) Responses of Sphagnum species to polluted environments. In: Bates JW, Farmer AM (eds) Bryophytes and lichens in a changing environment. Clarendon Press, Oxford, pp 314–332
Legendre P (2008) Studying beta diversity: ecological variation partitioning by multiple regression and canonical analysis. J Plant Ecol 1:3–8. https://doi.org/10.1093/jpe/rtm001
Legendre P, Legendre L (2012) Numerical Ecology. Elsevier, 990 pp
Lenth RV (2022a) emmeans: Estimated marginal means, aka least-squares means. R package version 1.7.2. https://CRAN.R-project.org/package=emmeans
Lindenmayer DB, Franklin JF, Lohmus A, Baker SC, Bauhus J, Beese W et al (2012) A major shift to the retention approach for forestry can help resolve some global forest sustainability issues. Conserv Lett 5:421–431. https://doi.org/10.1111/j.1755-263X.2012.00257.x
Linkola K (1916) Studien über den Einfluss der Kultur aufdie Flora in den Gegenden nördlich vom Ladogasee. I. Acta Soc Flora Fauna Fennica 45:1–432
Longton RE (1988) Biology of polar bryophytes and lichens. Cambridge University Press, Cambridge
Looman J (1964) Ecology of lichen and bryophyte communities in Saskatchewan. Ecology 45:481–491
Loustau D, Ogée J, Dufrêne E, Déqué M, Dupouey JL, Badeau V, Viovy N, Ciais P, Desprez-Loustau ML, Roques A, Chuine I, Mouillot F (2007) Impacts of climate change on temperate forests and interaction with management. In: Freer-Smith PH, Broadmeadow MSJ, Lynch JM (eds) Forestry and climate change. CAB International, Wallingford, pp 143–150
Makipaa R (1995) Sensitivity of forest-floor mosses in boreal forests to nitrogen and sulphur deposition. Water Air Soil Pollut 85:1239–1244
Marschall M, Proctor MCF (2004) Are bryophytes shade plants? Photosynthetic light responses and proportions of chlorophyll a, chlorophyll b and total carotenoids. Ann Bot 94:593–603. https://doi.org/10.1093/aob/mch178
Medina NG, Draper I, Lara F (2011) Biogeography of mosses and allies: does size matter? In: Fontaneto D (ed) Biogeography of microscopic organisms. Is everything small everywhere? Cambridge University Press, Cambridge, pp 209–233
Mills SE, MacDonald SE (2004) Predictors of moss and liverwort species diversity of microsites in conifer-dominated boreal forest. J Veg Sci 15:189–198. https://doi.org/10.1658/1100-9233(2004)015
Mölder A, Schmidt M, Engel F, Schonfelder E, Schulz F (2015) Bryophytes as indicators of ancient woodlands in Schleswig-Holstein (Northern Germany). Ecol Ind 54:12–30
Miller KR (1996) Conserving biodiversity in managed landscapes. In: Szaro RC, Johnston DW (eds) Biodiversity in managed landscapes. Oxford University Press, New York, pp 425–441
Müller J, Boch S, Prati D, Socher SA, Pommer U, Hessenmöller D, Schall P, Schulze ED, Fischer M (2019) Effects of forest management on bryophyte species richness in Central European forests. For Ecol Manage 432:850–859
Newmaster SG, Bell FW (2002) The effects of silvicultural disturbances on cryptogam diversity in the boreal-mixed wood forests. Can J for Res 32:38–51
Newmaster SG, Vitt DH, Belland RJ, Arsenault A (2005) The ones we left behind: comparing plot sampling and floristic habitat sampling for estimating biodiversity. Divers Distrib 11:57–72. https://doi.org/10.1111/j.1366-9516.2005.00123.x
Nordin A, Strengbom J, Witzell J, Näsholm T, Ericson L (2005) Nitrogen deposition and the biodiversity of boreal forests: implications for the nitrogen critical load. Ambio 34:20–24
Noss RF (1996) Conservation of biodiversity at the landscape scale. In: Szaro RC, Johnston DW (eds) Biodiversity in managed landscapes. Oxford University Press, Oxford, pp 574–592
Ochyra R, Zarnowiec J, Bednarek-Ochyra H (2003) Census catalogue of polish mosses. Biodiversity of Poland, vol 3. IB PAN, Krakow.
Økland RH (2003) Partitioning the variation in a plotby-species data matrix that is related to n sets of explanatory variables. J Veg Sci 14:693–700. https://doi.org/10.1111/j.1654-1103.2003.tb02201.x
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchinm PR, O’Hara RB, Simpson GL, Solymos P, Stevens MH, Eduard Szoecs E, Wagner H (2020) vegan: Community ecology package. R package version 2.5–7. https://CRAN.R-project.org/package=vegan.
Paquette A, Messier C (2009) The role of plantations in managing the world’s forests in the Anthropocene. Front Ecol Environ 8:27–34
Peñuelas J, Sardans J, Estiarte M, Ogaya R, Carnicer J, Coll M et al (2013) Evidence of current impact of climate change on life: a walk from genes to the biosphere. Glob Change Biol 19:2303–2338
Pińskwar I, Choryński A, Kundzewicz ZW (2020) Severe drought in the spring of 2020 in Poland—more of the same? Agronomy 10:1646. https://doi.org/10.3390/agronomy10111646
Proctor MCF, Oliver MJ, Wood AJ, Alpert P, Stark LR, Cleavitt NL, Mishler BD (2007) Desiccation-tolerance in bryophytes: a review. Bryologist 110:595–621
Proctor MCF (2011) Climatic responses and limits of bryophytes: comparison and contrast with vascular plants. In: Tuba Z, Slack N, Stark LR (eds) Bryophyte ecology and climate change. Cambridge University Press, Cambridge, UK, pp 35–54
Qu Y, Zhuang Q (2019) Evapotranspiration in North America: implications for water resources in a changing climate. Mitig Adapt Strat Glob Change 25:205–220. https://doi.org/10.1007/s11027-019-09865-6
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
Report on the European Forest Strategy - the way forward (2019/2157(INI)), 2020.
Ross-Davis AL, Frego KA (2002) Comparison of plantations and naturally regenerated clearcuts in the Acadian Forest: forest floor bryophyte community and habitat features. Can J Bot 80:21–33
Roth T, Kohli L, Rihm B, Achermann B (2013) Nitrogen deposition is negatively related to species richness and species composition of vascular plants and bryophytes in Swiss mountain grassland. Agr Ecosyst Environ 178:121–126
Rudolphi J, Jönssonm T, Gustafsson L (2014) Biological legacies buffer local species extinction after logging. J Appl Ecol 51:53–62. https://doi.org/10.1111/1365-2664.12187
Russell VL (2022b) emmeans: estimated marginal means, aka Least-Squares Means. R package version 1.7.2. https://CRAN.R-project.org/package=emmeans
Rustad LE, Campbell JL, Marion GM, Norby RJ, Mitchell MJ, Hartley AE et al (2001) A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 126:543–562
Schwaab J, Davin EL, Bebi P et al (2020) Increasing the broad-leaved tree fraction in European forests mitigates hot temperature extremes. Sci Rep 10:14153. https://doi.org/10.1038/s41598-020-71055-1
Shaw J, Szövényi P, Shaw B (2011) Bryophyte diversity and evolution: windows into the early evolution of land plants. Am J Bot 98:352–369. https://doi.org/10.3732/ajb.1000316
Shelley AE, Halpern CB, McKenzie D (2012) The contributions of forest structure and substrate to bryophyte diversity and abundance in mature coniferous forests of the Pacific Northwest. Bryologist 115:278–294
Shoemaker LJ, Sullivan LL, Donohue I, Cabral JS, Williams RJ et al (2020) Integrating the underlying structure of stochasticity into community ecology. Ecol 101:e02922. https://doi.org/10.1002/ecy.2922
Sjogren E (1995) Changes in the epilithic and epiphytic moss cover in two deciduous forest areas on the island of Oland (Sweden)—a comparison between 1958–1962 and 1988–1990. Stud Plant Ecol 19:1–108
Solon J, Wawrzoniak J (1999) Monitoring runa i odnowienia naturalnego w systemie monitoring lasu, In Prace IBL, Seria A, pp 877–108
Song L, Liu W-Y, Nadkarni NM (2012) Response of non-vascular epiphytes to simulated climate change in a montane moist evergreen broad-leaved forest in southwest China. Biol Conserv 152:127–135. https://doi.org/10.1016/j.biocon.2012.04.002
Staniaszek-Kik M, Żarnowiec J, Stefańska-Krzaczek E (2021) Diversity and composition of moss guilds on uprooted trees in Central European mountain forests: effects of uprooting components and environmental variables. Ann Sci 78:45
Stehn SE, Webster CR, Glime J, Jenkins MA (2010) Elevational gradients of bryophyte diversity, life forms, and community assemblage in the southern Appalachian Mountains. Can J Res 40:2164–2174. https://doi.org/10.1139/X10-156
Stofer S, Calatayud V, Giordani P, Neville P (2016) Part VII.2: assessment of epiphytic lichen diversity. In: UNECE ICP Forests Programme Co-ordinating Centre (ed.): Manual on methods and criteria for harmonized sampling, assessment, monitoring and analysis of the effects of air pollution on forests. Thünen Institute of Forest Ecosystems, Eberswalde, Germany. http://www.icp-forests.org/manual.htm
Suggitt AJ, Gillingham PK, Hill JK, Huntley B, Kunin WE, Roy DB, Thomas CD (2011) Habitat microclimates drive fine-scale variation in extreme temperatures. Oikos 120:1–8. https://doi.org/10.1111/j.1600-0706.2010.18270.x
Sundberg S (2005) Larger capsules enhance short-range spore dispersal in Sphagnum, but what happens further away? Oikos 108:115–124. https://doi.org/10.1111/j.0030-1299.2005.12916.x
Sundberg S (2013) Spore rain in relation to regional sources and beyond. Ecography 36:364–373. https://doi.org/10.1111/j.1600-0587.2012.07664.x
Szövényi P, Sundberg S, Shaw AJ (2012) Long-distance dispersal and genetic structure of natural populations: an assessment of the inverse isolation hypothesis in peat mosses. Mol Ecol 21:5461–5472. https://doi.org/10.1111/mec.12055
Szweykowski J (2006) An annotated checklist of Polish liverworts and hornworts. IB PAN, Kraków
Teuling AJ, de Badts EAG, Jansen FA, Fuchs R et al (2019) Climate change, reforestation/afforestation, and urbanization impacts on evapotranspiration and streamflow in Europe. Hydrol Earth Syst Sci 23:3631–3652. https://doi.org/10.5194/hess-23-3631-2019
Theurillat JP, Schlüssel A, Geissler P, Guisan A, Velluti C, Wiget L (2003) Vascular plant and bryophyte diversity along elevation gradients in the alps. In: Nagy L, Grabherr G, Körner C, Thompson DBA (eds) Alpine biodiversity in Europe. Ecological studies (analysis and synthesis), vol 167. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-18967-8_8
Trigas P, Panitsa M, Tsiftsis S (2013) Elevational gradient of vascular plant species richness and endemism in Crete—the effect of post-isolation mountain uplift on a continental island system. PLoS ONE 8(3):e59425. https://doi.org/10.1371/journal.pone.0059425
Vanderpoorten A, Engels P (2002) The effects of environmental variation on bryophytes at a regional scale. Ecography 25:513–522
van der Wal R, Imogen SKP, Brooker RW (2005) Mosses and the struggle for light in a nitrogen-polluted world. Oecologia 142:159–168. https://doi.org/10.1007/s00442-004-1706-0
van Wijk MT, Clemmensen KE, Shaver GR, Williams M, Callaghan TV et al (2003) Long-term ecosystem level experiments at Toolik Lake, Alaska, and at Abisko, Northern Sweden: generalizations and differences in ecosystem and plant type responses to global change. Glob Change Biol 10:105–123
Vivanco MG, Theobald MR, García-Gómez H et al (2018) Modeled deposition of nitrogen and sulfur in Europe estimated by 14 air quality model systems: evaluation, effects of changes in emissions and implications for habitat protection. Atmos Chem Phys 18(10199–10218):2018. https://doi.org/10.5194/acp-18-10199-2018
von Arx G, Pannatier EG, Thimonier A, Rebetez M (2013) Microclimate in forests with varying leaf area index and soil moisture: potential implications for seedling establishment in a changing climate. J Ecol 101:1201–1213
Walker MD, Wahren CH et al (2006) Plant community responses to experimental warming across the tundra biome. Proc Natl Aced Sci USA 103:1342–1346
Werner FA, Gradstein SR (2009) Diversity of dry forest epiphytes along a gradientof human disturbance in the tropical Andes. J Veg Sci 20:59–68. https://doi.org/10.1111/j.1654-1103.2009.05286.x
West PW (2014) Growing plantation forests. Springer, Dordrecht, Heidelberg, London, New York
Wolf JHD (2005) The response of epiphytes to anthropogenicdisturbance of pine-oak forests in the highlands of Chiapas, Mexico. For Ecol Manage 212:376–393
Wu Z, Dijkstra P, Koch GW, Peñuelas J, Hungate BA (2011) Responses of terrestrial ecosystems to temperature and precipitation change: a meta-analysis of experimental manipulation. Glob Change Biol 17:927–942
Zasady, Kryteria i Wskaźniki Dobrej Gospodarki Leśnej w Polsce (2010) Forest Stewardship Council, Poland.
Zasady hodowli lasu (2012) CILP, Warszawa
Zerbe S (2002) Restoration of natural broad-leaved woodland in Central Europe on sites with coniferous forest plantations. For Ecol Manage 167:27–42
Zhang R, Zhang Z, Shang K, Zhao M, Kong J, et al (2021) A taxonomic and phylogenetic perspective on plant community assembly along an elevational gradient in subtropical forests. J Plant Ecol 14:702–716. https://doi.org/10.1093/jpe/rtab026
Ziernicka-Wojtaszek A, Kopcińska J (2020) Variation in atmospheric precipitation in Poland in the Years 2001–2018. Atmosphere 11:794. https://doi.org/10.3390/atmos11080794
Acknowledgements
We owe special thanks to Dr. Enrico Tordoni for suggestions and help with statistical analyses. We thank Robert Hildebrand and Jadwiga Małachowska for providing archival data and preparing the plot distribution map, and the Bureau of Forest Management and Geodesy staff who participated in the collection of field data.
Funding
This work was supported by a grant from the Polish Institute of Forest Research Institute. This work was supported by a scholarship fund from the Forest Research Institute (IBL), Poland. The collection of raw data was supported by numerous projects of the Ministry of Environment, the General Inspectorate of Environmental Protection and the General Directorate of State Forests and co-financed by the National Fund for Environmental Protection and Water Management.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Field data were provided by PL and JC. Material preparation, data collection and analysis were performed by CC. The manuscript was written by CC. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Communicated by T.G. Allan Green.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article belongs to the Topical Collection: Forest and plantation biodiversity.
Appendices
Appendix 1
List of bryophyte species occuring in the study areas. NH-non hemerophobic; H-hemerophobic; SH-strongly hemerophobic.
Species | Hemerophoby status |
|---|---|
Amblystegium serpens (Hedw.) Schimp | NH |
Atrichum undulatum (Hedw.) P.Beauv | NH |
Aulacomnium androgynum (Hedw.) Schwägr | H |
Aulacomnium palustre (Hedw.) Schwägr | H |
Bazzania trilobata (L.) S. Gray | H |
Blepharostoma trichophyllum (L.) Dumort | H |
Brachytheciastrum velutinum (Hedw.) Ignatov & Huttunen | NH |
Brachythecium albicans (Hedw.) Schimp | NH |
Brachythecium campestre (Müll. Hal.) Schimp | NH |
Brachythecium laetum (Bridel) Schimper | NH |
Brachythecium mildaeanum (Schimp.) Schimp | NH |
Brachythecium rivulare Schimp | H |
Brachythecium rutabulum (Hedw.) Schimp | NH |
Brachythecium salebrosum (Hoffm. ex F.Weber et D.Mohr) Schimp | H |
Bryum capillare Hedw | NH |
Bryum subneodamense Kindb | H |
Buxbaumia aphylla Hedw | H |
Callicladium haldanianum (Grev.) H.A.Crum | H |
Calliergon stramineum (Dicks. ex Brid.) Hedenäs | NH |
Calypogeia azurea Stotler & Crotz | NH |
Calypogeia integristipula Steph | H |
Calypogeia muelleriana (Schiffn.) Müll.Frib | H |
Cephalozia bicuspidata (L.) Dumort | NH |
Cephaloziella starkei (Sm.) Schiffn | NH |
Ceratodon purpuraeus (Hedw.) Brid | NH |
Cirriphyllum crassinervium (Taylor ex Wilson) Loeske & M.Fleisch | SH |
Cirriphyllum piliferum (Hedw.) Grout | NH |
Climacium dendroides (Hedw.) Web. & Mohr | NH |
Dicranella cerviculata (Hedwig) Schimp | NH |
Dicranella heteromalla (Hedw.) Schimp | NH |
Dicranodontium denudatum (Brid.) E.Britton | SH |
Dicranoweisia cirrata (Hedw.) Lindb | NH |
Dicranum majus Turn | H |
Dicranum polysetum Sw | H |
Dicranum scoparium Hedw | H |
Dicranum spurium Hedw | NH |
Dicranum tauricum Sapjegin | NH |
Dicranum undulatum Schrad. ex Brid | SH |
Diplophyllum albicans (L.) Dumort | NH |
Ditrichum heteromallum (Hedwig) E. Britton | NH |
Encalypta streptocarpa Hedw | H |
Eurhynchium anguistirete (Broth.) T.J. Kop | H |
Eurhynchium schleicheri (R. Hedw.) Milde | NH |
Eurhynchium striatum (Schreb. ex Hedw.) Schimp | H |
Fissidens adianthoides Hedw | H |
Fissidens bryoides Hedw | NH |
Fissidens osmundoides Hedw | SH |
Geocalyx graveolens (Schrad.) Nees | SH |
Grimmia pulvinata (Hedw.) Sm | NH |
Herzogiella seligeri (Brid.) Z.Iwats | H |
Homalia trichomanoides (Hedw.) Schimp | H |
Homalothecium sericeum (Hedw.) Bruch, Schimp. & W.Guembel | H |
Hylocomium splendens (Hedw.) Schimp | H |
Hypnum andoi A.J.E.Sm | NH |
Calliergonella lindbergii (Mitt.) Hedenäs | NH |
Hypnum cupressiforme Hedw | NH |
Hypnum imponens Hedw | NH |
Hypnum jutlandicum Holmen & E.Warncke | NH |
Hypnum pallescens (Hedw.) P.Beauv | SH |
Isothecium alopecuroides (Lam. ex Dubois) Isov | H |
Isothecium myosuroides Brid | H |
Lepidozia reptans (L.) Dumort | H |
Leucobryum glaucum (Hedw.) Ångstr | H |
Leucobryum juniperoideum (Brid.) Müll.Hal | NH |
Leucodon sciuroides (Hedw.) Schwägr | NH |
Lophocolea bidentata (L.) Dumort | NH |
Lophocolea heterophylla (Schrad.) Dumort | NH |
Lophozia ventricosa (Dicks.) Dumort | H |
Metzgeria furcata (L.) Dumort | H |
Mnium hornum Hedw | H |
Nowellia curvifolia (Dicks.) Mitt | H |
Orthodicranum montanum (Hedw.) Loeske | NH |
Orthotrichum affine Schrad. ex Brid | NH |
Oxyrrhynchium hians (Hedw.) Loeske | NH |
Paraleucobryum longifolium (Ehrh. ex Hedw.) Loeske | SH |
Pellia epiphylla (L.) Corda | H |
Plagiochila asplenioides (L. emend. Taylor) Dumort | H |
Plagiochila porelloides (Torrey ex Nees) Lindenb | H |
Plagiomnium affine (Blandow ex Funck) T.J.Kop | NH |
Plagiomnium cuspidatum (Hedw.) T.J.Kop | H |
Plagiomnium rostratum (Schrad.) T.J.Kop | H |
Plagiomnium undulatum (Hedw.) T.J. Kop | NH |
Plagiothecium cavifolium (Brid.) Z.Iwats | H |
Plagiothecium curvifolium Schlieph. ex Limpr | H |
Plagiothecium denticulatum (Hedw.) Schimp | H |
Plagiothecium laetum Schimp | SH |
Plagiothecium nemorale (Mitt.) A.Jaeger | NH |
Plagiothecium succulentum (Wilson) Lindb | NH |
Plagiothecium undulatum (Hedw.) Schimp | H |
Platygyrium repens (Brid.) Schimp | NH |
Pleurozium schreiberi (Willd. ex Brid.) Mitt | H |
Pohlia nutans (Hedw.) Lindb | NH |
Polytrichastrum formosum (Hedw.) G. L. Sm | H |
Polytrichastrum longisetum (Brid.) G.L.Sm | NH |
Polytrichum commune Hedw | H |
Polytrichum juniperinum Hedw | NH |
Polytrichum piliferum Hedw | NH |
Polytrichum strictum Brid | H |
Pseudoscleropodium purum (Limpr) M. Fleisch. ex Broth | NH |
Pseudotaxyphillum elegans Z. Iwats | H |
Pterigynandrum filiforme Hedw | SH |
Ptilidium ciliare (L.) Hampe | H |
Ptilidium pulcherrimum (Weber) Vain | H |
Ptilium crista-castrensis (Hedw.) De Not | NH |
Pylaisia polyantha (Hedw.) Schimp | H |
Racomitrium heterostichum (Hedw.) Brid | H |
Racomitrium sudeticum (Funck) Bruch & Schimp | SH |
Radula complanata (L.) Dumort | SH |
Rhizomnium punctatum (Hedw.) T.J. Kop | H |
Rhodobryum roseum (Hedw.) Limpr | H |
Rhytidiadelphus loreus (Hedw.) Warnst | SH |
Rhytidiadelphus squarrosus (Hedw.) Warnst | NH |
Rhytidiadelphus subpinnatus (Lindb.) T.J.Kop | H |
Rhytidiadelphus triquetrus (Hedw.) Warnst | H |
Sanionia uncinata (Hedw.) Loeske | NH |
Scapania nemorea (L.) Grolle | H |
Sciuro-hypnum oedipodium (Mitt.) Ignatov et Huttunen | NH |
Sciuro-hypnum plumosum (Hedw.) Ignatov et Huttunen | H |
Sciuro-hypnum reflexum (Starke) Ignatov et Huttunen | H |
Sciuro-hypnum starkei (Brid.) Ignatov et Huttunen | H |
Sphagnum capillifolium (Ehrh.) Hedw | H |
Sphagnum cuspidatum Ehrh. ex Hoffm | H |
Sphagnum fallax (Klinggr.) Klinggr | NH |
Sphagnum girgensohnii Russow | H |
Sphagnum palustre L | H |
Tetraphis pellucida Hedw | H |
Thuidium philibertii Limpr | H |
Thuidium recognitum (Hedw.) Lindb | H |
Thuidium tamariscinum (Hedw.) Schimp | H |
Ulota bruchii Hornsch | H |
Ulota crispa (Hedw.) Brid | H |
Ulota phyllantha (Brid.) Sawicki, Plášek & Ochyra | SH |
Appendix 2
Trend of the error in measuring the multivariate dispersion of the community as the sampling effort increases calculated by applying Jaccard dissimilarity.

Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cacciatori, C., Czerepko, J. & Lech, P. Long-term changes in bryophyte diversity of central European managed forests depending on site environmental features. Biodivers Conserv 31, 2657–2681 (2022). https://doi.org/10.1007/s10531-022-02449-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-022-02449-y