
Abstract
The goal of most invasive species suppression programs is to achieve long-term sustained reductions in population abundance, yet removal programs can be stymied by density-dependent population responses. We tested a harvest removal strategy for invasive Rusty Crayfish (Faxonius rusticus) at two nearshore native fish spawning habitats in northern Lake Michigan. Changes in average Rusty Crayfish densities were evaluated with a before-after reference-impact study design. We removed 3182 Rusty Crayfish, primarily adults (> 20 mm carapace length), at two sites over two harvest seasons, expending 17,825 trap days in effort. Generalized linear modeling results suggested a statistically significant reduction in Rusty Crayfish densities was achieved at one reef, Little Traverse Bay (LTB Crib). Reduced densities were sustained over the egg maturation period for native fish and into the following year after removal ceased. By late summer/early fall, between consecutive suppression efforts in 2018 and 2019, we observed a threefold increase in pre-removal densities. Size-frequency histograms from diver quadrat surveys showed higher abundances of juvenile (< 20 mm carapace length) size classes the following spring and summer at LTB Crib compared to its paired reference site. Stock-recruit curves fit to count data, pooled across all sites, provided further evidence of density-dependence. With a proviso that we only conducted two seasons of consecutive suppression, this study highlights an important aspect of invasive species management and raises questions about the efficacy of adult-only crayfish removal strategies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Suppression of an invasive species to reverse or lessen their impacts is a common management strategy (Hein et al. 2006; Weidel et al. 2007; Coggins et al. 2011). Mechanical removal through repeated and intense harvests can lead to sustained reductions in invasive species abundances, reducing negative effects to native species, but is often used in combination with other strategies to achieve eradication or suppression targets (Bills and Marking 1988; Hein et al. 2006; Howald et al. 2007). Most successful examples of mechanical removal have only been demonstrated in small enclosed systems, such as inland lakes, small streams, or islands (Hein et al. 2007; Glen et al. 2013; Moody et al. 2021). Few mechanical removal programs have been evaluated for large interconnected systems, but recolonization from adjacent source populations may introduce an added dimension of complexity for management (Weber et al. 2016; Dauphinais et al. 2018).
Reducing population abundance frees resources, such as food or habitat, for the remaining individuals, potentially eliciting a density dependent response in the population. Density dependence is a population-level phenomenon affecting population growth factors such as survival, individual growth rates, fecundity, and maturity rates (Colby and Nepszy 1981; Cooch et al. 1989; Coulson et al. 2000; Einum et al. 2006; De Roos et al. 2007). Populations which exhibit density dependent behavior may compensate for reductions in abundance through increased recruitment and faster growth, resulting from reduced mortality, increased survivorship, and higher consumption in the remaining individuals. By contrast, populations may experience decreases in growth at low population densities, commonly referred to as depensation in fisheries science (Myers et al. 1995; Liermann and Hilborn 1997). Depensatory population dynamics should not be confused with Allee effects, which are generally defined as a positive relationship between any component of individual fitness and conspecific densities (Stephens et al. 1999). Species with high fecundity, short juvenile stages, and low variance in survivorship are especially likely to exhibit compensatory responses to harvest (Zipkin et al. 2009). In some cases, populations can overcompensate in response to harvest leading to higher population abundances (De Roos et al. 2007; Zipkin et al. 2008). Understanding when and how populations are regulated by density dependent processes is crucial for informing invasive species management programs, lest the intended effects of harvest be negated by compensatory population growth.
Crayfish are one of the most common non-native taxa found in freshwater systems and consistently have negative overall impacts to native biota (Twardochleb et al. 2013). Management of non-native crayfish is a priority for conservation managers worldwide due to their widespread impacts, however no unified management framework exists (Hefti and Stucki 2006; Peay 2009; Freeman et al. 2010; Gherardi et al. 2011; Lodge et al. 2012). Non-native crayfish frequently replace native congeners, destroy aquatic macrophytes, and prey on or compete with other native fish and invertebrates (Hill and Lodge 1999; Claramunt et al. 2005; Olden et al. 2006; Morse et al. 2013). Under certain circumstances, non-native crayfish invasions can facilitate regime shifts in ecosystem organization, leading to alternative stable ecosystem states with permanently diminished ecosystem services (Scheffer et al. 2001; Hansen et al. 2013b). As such, substantial interest in the prevention and management of non-native crayfish has developed, especially in the last decade (Freeman et al. 2010; Gherardi et al. 2011; Manfrin et al. 2019). Rusty Crayfish (Faxonius rusticus) is the most widespread non-native crayfish species in the Laurentian Great Lakes basin (hereafter: Great Lakes) and is pervasive on nearshore spawning reef habitats for native fish such as Lake Trout (Salvelinus namaycush), Lake Whitefish (Coregonus clupeaformis), and Cisco (C. artedi) (Peters et al. 2014; Kvistad et al. 2021b). Concerns over interstitial egg predation on valuable sport and commercial fish species has motivated interest in suppressing Rusty Crayfish at reef habitats in the Great Lakes (Buckley 2016; Kvistad et al. 2021a).
Here we present multiple lines of evidence for a density dependent response to suppression in a population of invasive Rusty Crayfish on northern Lake Michigan spawning reefs. We implemented a suppression strategy on nearshore fish spawning reefs in the fall of 2018 and 2019, just prior to the arrival of native spawning fish. We used a before-after control-impact study design to determine the extent to which suppression affected Rusty Crayfish densities estimated from diver quadrat survey counts. Evidence from stock-recruitment modeling, length-frequency analysis, and ovarian egg counts suggested a potential density dependent response to suppression. We discuss these results in the larger context of invasive crayfish management.
Methods
Study sites
Rusty Crayfish suppression and monitoring efforts were implemented at two nearshore cobble spawning reefs with paired nearby reference reefs in northern Lake Michigan. Two sites in Little Traverse Bay, the Crib (hereafter, LTB Crib) and its nearby reference site (hereafter, LTB Reference) were the focus of treatment and monitoring efforts in both 2018 and 2019. Two additional sites were added in 2019 in Grand Traverse Bay near the city of Elk Rapids, MI; the Point (hereafter, ER Point) and its nearby reference site (hereafter, ER Reference). The LTB Crib (45° 25′ N, 84° 56′ W) reef is the remnants of private crib wharf (approximately 1500 m2 in shallow water < 5 m deep). Common along privately owned shorelines throughout northern Michigan, crib wharves are long rectangular wooden frames, typically filled with cobbles for weight to support a walkway (Hoxie 2009). All that remains of the wharf at LTB Crib now are the cobbles that once filled the wood structure, which provides high quality spawning habitat for Lake Trout and coregonids. The LTB Reference site is an area of naturally occurring shallow coastal cobble habitat (comparable to the cobble habitat adjacent to the core reef at the LTB Crib) about 300 m west of LTB Crib. The ER Point and ER Reference (44° 54′ N, 85° 25′ W) spawning reefs in Grand Traverse Bay are part of the Elk Rapids reef complex, a nearly contiguous matrix of glacially deposited cobble, boulder, and imbedded cobble substrate where Lake Trout, Cisco, and Lake Whitefish are known to spawn. The reef complex is approximately 2 km2. Habitat quality at the Elk Rapids complex varies, and there are certain areas (reefs) which contain higher quality spawning habitat. These reefs, like ER Point and ER Reference, are smaller areas (1500 m2), comprised of linear strips of cobble or boulder banks that sit on primarily bedrock, and run parallel to the shore.
Removal strategy
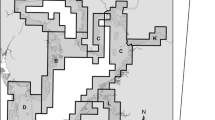
Trap** occurred at impact sites from the end of late summer (late August or early September) until mid-to-late fall (late October or early November). Observations of a rapid but short-lived increase in catch-per-unit-effort (CPUE) highlights this mid-to-late fall time period as significant in Rusty Crayfish life cycles, and presents the greatest window of opportunity for suppression (Robinson 2014; Buckley 2016; Kvistad et al. 2021b). At LTB Crib in 2018, twelve 60 m “main” traplines consisting of eleven traps spaced 5 m apart and three 30 m “core reef” traplines with eleven traps spaced 3 m apart were set perpendicular to shore covering the core reef and adjacent areas. Core reef traps were spaced closer together to increase the density of traps over the highest quality Lake Trout spawning habitat. For the main traplines, we set each trapline such that the first trap sat on sand roughly 10 m inshore from the main cobble habitat. Traplines ran parallel to the artificial crib structure, which extends from shore to approximately 100 m out into the bay. In 2018 165 traps were deployed. In 2019 at LTB Crib, we reduced the number of traplines to thirteen, which consisted of ten main traplines and three core reef traplines. We used primarily Gee’s® G-40 Minnow Trap (Tackle Factory; Fillmore, NY), approximately half of which were modified for part of a concurrent trap comparison study (Kvistad et al. 2021a). We extended the main traplines from 60 to 90 m, which increased the number of traps per line to seventeen. Extending the traplines allowed for trap** coverage on the sand habitat past the core reef. However, we reduced the number of traps to fourteen after the first month of trap** due to time constraints preventing all traps from being checked in a single day. The total number of traps deployed in 2019 at LTB Crib ranged from 200 at the start of trap** and 170 near the end of trap** (Fig. 1).

Trapline and barrier configuration over the spawning reef (boulder reef) and adjacent sand and cobble areas at LTB Crib, 2019
At ER Point in 2019, we deployed a total of ten traplines. Seventy-nine traps were deployed across seven traplines in buffer areas adjacent to the core reef. Traps were spaced 5 m apart on traplines. Forty-seven traps, across three traplines were deployed over the core reef, with traps spaced approximately 3 m apart. In total, 126 traps were deployed at ER Point at each trap** event (Fig. 2).

Trapline and barrier configuration over the spawning reef (cobble reef) and adjacent sand and cobble areas at ER Point, 2019
Soak time (number of nights fished) was variable throughout the suppression period (median soak time: 4 days; minimum soak time: 1 day; maximum soak time: 13 days), dependent on weather and sampling logistics. For all trap** events, we recorded the total numbers of Rusty Crayfish as each trap was emptied. Individuals were measured for carapace length (mm), sexed, and noted for the presence of any secondary sex characteristics (e.g., glair glands on females; copulatory stylets and ischial hooks on males). We retained all Rusty Crayfish caught to be euthanized and discarded.
Crayfish barrier
Previous evidence of rapid recolonization following removal efforts was a concern during suppression (Buckley 2016). Therefore, we deployed a metal barrier on the sand around the core reef at each of our treatment sites and used a secondary vinyl barrier based on a modified fyke wing design in the adjacent buffer areas and over uneven cobble substrate where the rigid metal barrier was ineffective. In contrast to the metal barriers, which were designed to remain in place throughout winter, vinyl barriers were only erected temporarily during Rusty Crayfish suppression. Vinyl barriers were removed during the last week of suppression efforts, prior to the last day of suppression to ensure they could be recovered before strong fall storms jeopardized safety. Rusty Crayfish catches typically decline to near zero during late fall in the Great Lakes, potentially suggesting either limited movement or offshore emigration (Robinson 2014). Therefore, it is unlikely that removing the barriers just prior to the last day of suppression would have influenced recolonization into treated areas. The metal barrier was deployed contiguous with the vinyl barrier over soft substrates at the nearshore and offshore edges of the core reef (Fig. 1). Vinyl barriers were deployed from August through December during both years of Rusty Crayfish suppression; however, metal barriers remained in place until suppression efforts terminated (2019). At ER Point, prior to the 2019 suppression efforts, we deployed vinyl barriers only around the core treatment reef and in adjacent buffer areas (Fig. 2). Barriers at ER Point were deployed from August through October 2019.
Crayfish dispersal and barrier effectiveness
A tagging study took place in August 2019 at LTB Crib, prior to suppression efforts, to assess barrier effectiveness. Divers delineated thirteen 5 × 5 m zones in the cobble habitat on either side of the boulder cobble habitat (see Fig. 1). Divers searched for adult (> 20 mm carapace length) Rusty Crayfish, measured (carapace length [mm]), sexed, and noted for secondary sex characteristics on all captured individuals. Captured Rusty Crayfish were marked with a fluorescent visual alphanumeric (VI Alpha) tag (Northwest Marine Technologies, Anacortes, WA) in the soft tissue under the third abdominal somite (Fig. 3). Tags were ascribed unique alphanumeric codes so that individuals could be identified upon recapture. Although VI Alpha tags have high retention in crayfish marking studies (Jerry et al. 2001), we gave each Rusty Crayfish a secondary mark in the form of a uropod clip. Divers returned each marked Rusty Crayfish back to the same zone from which it was captured. In total, 415 Rusty Crayfish were marked at LTB Crib in 2019. We estimated the distance traveled for each recaptured Rusty Crayfish by measuring the Euclidean distance from the center of each marking zone to the approximate location of the trap where it was recaptured.

A fluorescent visual alphanumeric (VI Alpha) tag (Northwest Marine Technologies, Anacortes, WA) being inserted into the abdomen of a Rusty Crayfish
Diver quadrat counts
Divers sampled treatment and control reefs both before and after suppression with ten randomly placed 1 m2 quadrats. Each site (LTB Crib and LTB Reference, ER Point and ER Reference) was sampled once in the summer (August or September) before Rusty Crayfish suppression started and again after suppression ended (late October or November). Additional post-suppression sampling occurred in the spring just after ice-off (late April or May). Spring quadrat sampling efforts in 2020 were precluded due to statewide restrictions imposed in response to the SARS-CoV-2 novel coronavirus pandemic. For each sampling event, divers searched the first layer of substrate in each quadrat and collected any Rusty Crayfish. Rusty Crayfish that escaped (< 10% of counts) the quadrat were noted, included in diver counts, and a visual estimate was given for approximate carapace length. All Rusty Crayfish collected were measured for carapace length (mm), sexed, and noted for the presence of any secondary sex characteristics (e.g., glair glands on females; copulatory stylets on males).
We used a before-after-reference-impact (BARI) study design coupled with a generalized linear mixed model to estimate the magnitude and statistical significance of our suppression efforts on Rusty Crayfish densities estimated from diver quadrats. Before-after-control-impact designs require simultaneous monitoring of treatment and control sites before and after treatment (Stewart-Oaten et al. 1986). The main principle behind a BARI analysis is that a disturbance inflicted on the “impact” (i.e. removal or treatment) location will result in measurable differences from before to after when compared to any natural change taking place at the reference site, which can be detected as a statistical interaction through an analysis of variance (Underwood 1992). Selection of proper controls is a critical step for BARI analyses to avoid risk of masking any statistically measurable response by variability caused by unmeasured forces (Underwood 1992).
To perform the analysis of variance, we used a Poisson family generalized linear model with a logarithmic link function. Because of differences in environmental context and geographic layouts between paired removal and reference sites, we conducted independent analyses for Little Traverse Bay and Elk Rapids site pairs. The generic model formula for the BARI design in this case was: \(Rusty\;Crayfish \sim Group + Period + \left( {Group*Period} \right)\). Where the estimated parameters included the monitoring period (before or after impact), the experimental group (treatment or reference), and the interaction between period and group \((Group*Period)\). In a BARI design, a significant interaction effect is indicative of statistical significance of the treatment overall (Underwood 1992). Statistical significance thresholds for all analyses were set at the conventional Type I error rate (α = 0.05). All statistical analysis and modeling was done in R (v4.0.0, R Core Team 2020). Generalized linear models were fit via maximum likelihood estimation using the lme4 package (Bates et al. 2015).
Female reproductive development
A random subset of female Rusty Crayfish collected semi-monthly from trap** efforts at LTB Crib were frozen and returned to the laboratory. Because females were only retained from trap removals, we did not have data on female reproductive development at reference sites. As such, these data do not adhere to the requirements of a BARI design and thus cannot be used to make inferences about differences in reproductive potential between impact and reference. However, these data are informative for examining between year differences at impact sites.
We recorded the carapace length (mm), gonad wet weight (W1), and total wet weight (W2) in grams. We calculated the gonadosomatic index (GSI) of each specimen with the formula: \(GSI=\frac{{W}_{1}}{{W}_{2}}*100\). Ovarian eggs from each female were removed and enumerated under a stereoscope. Median egg counts and the interquartile range (IQR) were calculated for each year. A regression analysis was performed to test for significant differences in ovarian egg counts between 2018 and 2019 using a Poisson distributed mixed effects model with a logarithmic l. Year, carapace length, and a year-carapace length interaction were included as fixed effects, while the month of collection was included as a random effect to account for intra-annual variability in egg production.
Stock-recruitment analysis
The stock-recruitment relationship of Rusty Crayfish was examined to evaluate evidence for density dependence on northern Lake Michigan spawning reefs. We examined evidence for density dependence by fitting three common stock-recruitment models to the relationship between adult female Rusty Crayfish density (stock) and juvenile Rusty Crayfish density (recruitment) collected during diver quadrat surveys in 2018 and 2019. The distinction between juvenile and adult Rusty Crayfish was determined through length-frequency analysis using Bhattacharya’s method (Bhattacharya 1967; Scalici et al. 2010). Using Bhattacharya’s method, we determined a reasonable size cut-off between juveniles and adults at a carapace length of 19.45 mm (Bhattacharya 1967). This size threshold agreed well with an earlier analysis by Kvistad et al. (2021b) which further subdivided Rusty Crayfish into four distinct size classes (1: > 10 mm, 2: 10–20 mm, 3: 21–30 mm, and 4: > 40 mm). Therefore, we assigned Rusty Crayfish to each of the four size classes with juveniles corresponding to size classes 1 and 2 and adults to size classes 3 and 4 for some data visualizations. We used Chi-square tests to test for equal frequencies across size classes at impact-reference site pairs.
We tested a linear model:\(R=\alpha S\), the Beverton–Holt model (Beverton and Holt 1957): \(R=\frac{\alpha S}{1+\beta S}\) and the Ricker model (Ricker 1954): \(R=\alpha S{e}^{-\beta S}\) where R is the juvenile density, S is the adult female density, \(\alpha\) represents the theoretical maximum number of offspring capable of being produced by each adult female, and \(\beta\) is the density dependence parameter. As \(\beta\) approaches 0, the effect of density dependence diminishes, and both the Beverton–Holt and the Ricker models reduce to the linear form. While the Beverton–Holt and the Ricker models both imply some form of density dependence, the interpretation of \(\beta\) differs slightly between each model. In the Beverton–Holt model, the density dependent interaction primarily occurs through competition in the juvenile stages, while the Ricker model implies that high adult densities directly impact juvenile densities (i.e., through competition or predation).
We performed model selection using Akaike’s Information Criterion corrected for small sample sizes (AICc) to determine the best fit model (Burnham and Anderson 2004). Models with \({\Delta }_{AIC}<2\) are considered to have substantial support in the data relative to the other candidate models (Burnham and Anderson 2004). Due to data sparsity issues preventing reliable parameter fits, data were pooled across all sites and years for the stock-recruitment analysis. Rusty Crayfish density values were log-transformed prior to fitting the models for analytical tractability. Upper and lower 95% confidence intervals for the non-linear least-squares parameter estimates were obtained through non-parametric bootstrap resampling.
Results
Rusty Crayfish suppression
In total, 3182 Rusty Crayfish were removed from both sites over two years (Table 1). Cumulative catch increased steadily at LTB Crib in 2018 and ER Point in 2019, while catch increased rapidly in late October at LTB Crib in 2019 before plateauing in mid-November (Fig. 4A). Catch-per-unit-effort increased at LTB Crib in 2018 and ER Point in 2019 in late October as trap** ceased while at LTB Crib in 2019, CPUE peaked in mid- to late October and declined to near zero by early November (Fig. 4B). While Rusty Crayfish densities declined at LTB Crib compared to LTB Reference following treatment in 2018 and 2019, we did not observe a similar decline at ER Point (Fig. 5). The ratio of juveniles (size classes 1 and 2) to adults (size classes 3 and 4) at LTB Crib and LTB Reference were similar before and after the first year of suppression but remained elevated at LTB Crib the following spring while it had returned to its previous level at LTB Reference (Fig. 6). Size-frequency histograms showed similar counts of size classes between LTB Crib and LTB Reference in 2018 and a higher frequency of juvenile size classes persisting over spring and fall (pre-suppression) at LTB Crib compared to LTB Reference in 2019 (Fig. 7). A Chi-squared test of the size class frequencies for 2018 determined no significant differences in size class distributions between LTB Crib and LTB Reference (χ2 = 2.83, P = 0.43). In 2019, however, the size class frequency distributions between LTB Crib and LTB Reference did differ significantly (χ2 = 21.18, P < 0.001).

Cumulative Rusty Crayfish catch (A) at each removal site and catch-per-unit-effort (B) (n trap-1 day-1; μ ± 95% CI) for Rusty Crayfish caught during removal

Rusty Crayfish density (n m2) estimates (μ ± 95% CI) from diver quadrat surveys during concurrent and post-treatment intervals (filled points) and during untreated intervals (open point) sites. The date ranges of our Rusty Crayfish suppression efforts at our treatment sites are highlighted by the shaded regions

The ratio of juvenile to adult Rusty Crayfish from diver quadrat surveys in Little Traverse Bay throughout suppression and monitoring. Shaded bars represent the suppression time window in 2018 and 2019

Size-frequency histograms for Rusty Crayfish collected from diver quadrat surveys at LTB Crib in 2018 and 2019
Rusty Crayfish densities were generally similar before suppression at impact and reference sites, and post-suppression densities varied considerably between sites (Table 2). The interaction term in the BARI analysis for the Little Traverse Bay sites was statistically significant (Z = − 4.46, P < 0.001), and the null hypothesis that suppression would have no detectable effect on Rusty Crayfish densities was rejected. However, the interaction term for the BARI analysis at Grand Traverse Bay indicated that suppression had no effect (Z = 1.67, P = 0.09). Site and year-specific means simulated from the fitted models showed differences in the expected response to suppression between site pairs in Little Traverse Bay and Grand Traverse Bay (Fig. 8).

Simulated means from site specific GLMs (closed points) with 95% prediction intervals (vertical lines) of the number of Rusty Crayfish observed in 1 m2 quadrat predicted by the model at each site before and after suppression. Empirically observed Rusty Crayfish counts from individual quadrat samples are displayed in the backgrounds of each panel (open points)
Crayfish dispersal and barrier effectiveness
We tagged 415 adult Rusty Crayfish at LTB Crib in August 2019, prior to setting traps in the water. Only 3% of the marked Rusty Crayfish (13 out of 415) were recovered during trap** and monitoring efforts at LTB Crib in 2019. However, roughly half of these (6 out of 13) were collected in a different zone from where they were tagged, suggesting that they had crossed one or more barriers over the duration of our suppression program. The average Euclidean distance traveled by marked Rusty Crayfish was 26.9 m (SD = 19.77).
Female reproductive development
A random subsample of 58 females in 2018 and 108 in 2019 were retained from LTB Crib for ovarian analysis. Mean GSI increased over the fall in both years but was higher in 2019 compared to 2018 for any given timeframe (Fig. 9A). Median ovarian egg count was 80 (IQR = 89) in 2018 compared to 174 (IQR = 108) in 2019. Median ovarian egg counts in October, when Rusty Crayfish typically engage in fall spawning on the reefs, were significantly higher (P = 0.004) in 2019 than in 2018 and increased by a factor of 1.6 (Fig. 9B). In addition, there was a significant interaction effect (P > 0.001) between year and carapace length, suggesting that the increase in median egg counts was greater for larger females.

A Mean (± SD) gonadosomatic index of female Rusty Crayfish sampled from diver quadrat surveys at LTB Crib. B Distribution (median and interquartile range) of ovarian egg counts collected from LTB Crib collected over time
Stock-recruitment analysis
The Beverton–Holt model received the most support by AICc weight (Table 3). Therefore \({\Delta }_{AIC}\) is 0 for the Beverton–Holt model, however \({\Delta }_{AIC}\) for the Ricker model was less than 2. As such, could not declare that any of the candidate models most parsimoniously explained the data. Visual comparisons of the fitted models, back transformed to the original scale, plotted against the data are displayed in Fig. 10.

Stock-recruitment curves for the three models tested to evaluate density dependence among Rusty Crayfish on northern Lake Michigan spawning reefs. Models were fit to data from diver quadrat density counts at LTB Crib and ER Point from 2018 to 2019. The Beverton–Holt model (\(\alpha =8.37, \beta =4.85)\) and the Ricker model (\(\alpha =3.24, \beta =0.70)\) tied in an AICc comparison
Discussion
In contrast with similar removal efforts in inland lake ecosystems, our study is among the first to offer evidence of a compensatory response in fecundity and juvenile recruitment in Rusty Crayfish following repeated harvest removals (Hein et al. 2006, 2007; Tetzlaff et al. 2011; Hansen et al. 2013a). But while those studies did not find evidence of density-dependence, part of their management intervention strategy was to target juveniles through increased fish predation and thus may have masked a compensatory response. In that regard, our findings are unique but not entirely unexpected. For example, Hawaiian spiny lobster (DeMartini et al. 1993) exhibit compensatory increases in fecundity in response to commercial harvest. Nonetheless, our study raises questions about the conditions under which density dependence can manifest in a population.
Numerous examples can be found in the literature of aquatic invasive species removal programs producing similar responses in the target species populations. For example, after two years of harvest removals of European Perch (Perca fluviatilis) from New Zealand ponds, young-of-the-year quickly dominated the population structure of treatment ponds compared to control ponds (Ludgate and Closs 2003). A study on Common Carp (Cyprinus carpio) removals from interconnected natural lakes in southeastern South Dakota, USA found an inverse relationship between Carp survival and removal rate (Weber et al. 2016). An eradication program targeting European Green Crab (Carcinus maenas) in a small California estuary saw a 30-fold single year increase in population abundance compared to nearby estuaries with no eradication efforts (Grosholz et al. 2021). Finally, despite removing over 450,000 Lake Trout from Yellowstone Lake, USA over a 15 year period, population growth rates remain above replacement (Syslo et al. 2011). Our study adds to the growing body of evidence that while mechanical removal programs can be effective at producing short-term reductions in population densities, alternative management strategies and/or high sustained efforts are necessary to achieve long-term suppression and the desired ecological responses as are often stated as program goals.
Cannibalism is characteristic of many age or size-structured populations and can be an important mechanism for population regulation through negative density dependence (Polis 1981; Van Buskirk 1989; Post et al. 1999). Common across decapod crustaceans, cannibalism can be a successful strategy when heterospecific prey densities are low because conspecifics are energy rich and possess the correct nutrient compositions relative to the species in question (Parsons et al. 2013). Although we did not study cannibalism in Rusty Crayfish in this study, cannibalism is frequently observed in other crayfish populations and has been discussed in the context of invasive crayfish removal or suppression (Houghton et al. 2017; He et al. 2021). Previous research has indicated that size-segregation occurs among Rusty Crayfish on Lake Michigan reefs (Kvistad et al. 2021b). Further, evidence of compensatory population dynamics from this study may indicate that cannibalism is a potentially important mechanism on these habitats. Empirical studies of cannibalism on Lake Michigan reefs are needed. However, if verified, this knowledge could have important implications for crayfish management, as it would imply that adult-only harvest removal strategies alleviate intraspecific predation on the juvenile population.
Various aspects of population biology are emphasized in invasion biology (Sakai et al. 2001), but such knowledge is rarely used to predict management outcomes prior to application in the field. Yet quantifying population growth parameters and simulating perturbations can lead to valuable insights for guiding suppression efforts (Govindarajulu et al. 2005; Day et al. 2018). Mathematical models can provide scientific knowledge on the effectiveness of various management strategies which can help decision makers direct resources appropriately (N’Guyen et al. 2018). In one example, Milligan et al. (2017) used a hybrid mathematical model of Red Swamp Crayfish (Procambarus clarkii) and California Newt (Taricha torosa) population dynamics to determine optimal crayfish trap** regimes and predict time to local newt extinction under alternative trap** scenarios. Simulation studies leveraging knowledge of population biology and carefully designed models could save considerable time and effort and improve conservation outcomes by guiding management decisions. Incorporating aspects of density-dependence in population growth models through mechanisms such as competition or cannibalism could help improve Great Lakes invasive crayfish management strategies.
Programs focused only on mechanical removal usually must maintain high levels of effort over long timeframes to achieve desired ecological responses. Rapidly maturing species with high fecundity are among the most difficult to manage through mechanical removal alone, especially if the most widely available capture technology selects only for adult size classes (Stuecheli 1991). When programs do succeed, they usually combine multiple management strategies to ensure broad coverage of all life-history stages which can be sustained without continuous human intervention, such as various forms of biological control (Hein et al. 2007; Thresher et al. 2014). Thus while mechanical control strategies used in isolation often fail to achieve desired ecological outcomes, combining mechanical removals with alternative strategies (e.g., predator manipulation) can be highly successful (Hein et al. 2007; Thresher et al. 2014; Bajer 2019).
According to the enemy-release hypothesis, successful invaders often reach abundances near carrying capacity due to lack of regulation from co-evolved predators, pathogens, and competitors (Keane 2002; Mitchell and Power 2003; Torchin et al. 2003; Colautti et al. 2004). While the reefs in this study support populations of Rusty Crayfish predators, such as Smallmouth Bass (Micropterus dolomieu) and Rock Bass (Ambloplites rupestris), little is known about their population dynamics at nearshore reefs, especially during the fall when Rusty Crayfish are most active and susceptible to predation. Due to the open nature of the Great Lakes ecosystem compared to inland lakes and streams, migration of natural enemies may play an important role in predator–prey dynamics on reefs. In Lake Michigan, Smallmouth Bass have been documented migrating great distances offshore after spawning in the summer (Kaemingk et al. 2011). Therefore, Smallmouth Bass might be absent or only present in low densities in the late summer and fall when juvenile Rusty Crayfish recruitment occurs in Lake Michigan (Kvistad et al. 2021b). As such, a spatio-temporal mismatch of predator and prey in Lake Michigan may hinder the use of natural predators as a control mechanism. The use of native predators as a form of biocontrol on Rusty Crayfish could conceivably be achieved, however, through the use of enclosures to manipulate foraging locations and behaviors (Ward-Fear et al. 2009, 2010).
Recolonization after removal is another important consideration for harvest-management programs. Both the degree of isolation and the mobility potential of the targeted species can affect the outcome of harvest-management programs. Even in loosely connected ecosystems, relatively low immigration rates occurring infrequently (e.g., after heavy flooding in a system of interconnected lakes) can lead to rapid population recovery following harvest removal efforts (Weber et al. 2016). Buckley (2016) speculated that rapid recolonization occurred on Lake Michigan reefs after a period of intense trap** due to a rapid recovery observed in relative abundance following cessation of trap**. Our observations of traps further offshore catching more Rusty Crayfish later in the season support the possibility of recolonization occurring from outside the immediate treatment area. However, spatially explicit studies focused on measuring recolonization rates would be useful for improving understanding of immigration and emigration on reef habitats. Unfortunately, our mark-recapture efforts were not sufficient to characterize movement on reef habitats due to low-recapture rates. One potential reason for our low recapture rates is that we could not monitor whether any individuals left the study area. Nonetheless, we observed individuals traveling considerable distances suggesting a high degree of mobility. Movement dynamics of invasive species, including quantifying rates of immigration and emigration following removal efforts, is an area of research in need of greater attention (Green and Grosholz 2021). Such information could help conservation practitioners develop better suppression targets by considering meta-population dynamics and information about local connectivity across habitats. For Rusty Crayfish on GL reefs, our results suggest that successful control may require either removal across a larger buffer area (to mitigate recolonization of a core treatment area) or deployment of a more effective barrier to prevent recolonization.
Conclusion
In this study we highlight the non-linear dynamics of Rusty Crayfish suppression on critical Lake Michigan fish habitats. A simple harvest removal strategy focused only on adult size classes may have resulted in a density-dependent response in population growth. Our findings are relevant for natural resource managers and conservation practitioners concerned about Rusty Crayfish impacts to native fauna in Lake Michigan. The apparent role of density-dependence in Rusty Crayfish population dynamics in Lake Michigan should inform future management strategies to account for potential compensatory responses. Simulating population responses prior to field implementation of invasive species control programs could help prioritize efforts to achieve maximum impact and would likely point to the importance of targeting removal efforts to both adults and earlier life history stages.
Invasive crayfish management is a uniquely difficult problem for which no single paradigm exists (Freeman et al. 2010; Gherardi et al. 2011; Manfrin et al. 2019). Broadly applicable and highly effective invasive crayfish management strategies remain elusive, while promising strategies based on advances in genetics (i.e., gene silencing, gene editing, transgenes) are still unproven in terms of efficacy, safety, and morality (Simberloff 2021). In the absence of any “silver bullet” technologies, better integration of key ecological information, such as population dynamics, recolonization rates, and interactions with native enemies, with invasive species control programs is essential for devising effective management strategies.
Data availability
All data generated or analyzed during this study are included in the published article and its supplementary information files.
References
Bajer P (2019) Biological control of invasive fish and aquatic invertebrates: a brief review with case studies. Manag Biol Invasions 10:227–254. https://doi.org/10.3391/mbi.2019.10.2.02
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Beverton RJH, Holt SJ (1957) On the dynamics of exploited fish populations. Chapman and Hall, London
Bhattacharya CG (1967) A simple method of resolution of a distribution into Gaussian components. Biometrics 23:115–135
Bills TD, Marking LL (1988) Control of Nuisance populations of crayfish with traps and toxicants. Prog Fish-Cult 50:103–106. https://doi.org/10.1577/1548-8640(1988)050%3c0103:CONPOC%3e2.3.CO;2
Buckley JT (2016) Understanding rusty crayfish (Orconectes rusticus) distribution patterns and removal strategies within a Great Lake System. Master’s Thesis, Central Michigan University
Burnham KP, Anderson DR (2004) Multimodel inference: understanding AIC and BIC in model selection. Sociol Methods Res 33:261–304
Claramunt RM, Jonas JL, Fitzsimons JD, Marsden JE (2005) Influences of spawning habitat characteristics and interstitial predators on Lake Trout egg deposition and mortality. Trans Am Fish Soc 134:1048–1057. https://doi.org/10.1577/T04-075.1
Coggins LG, Yard MD, Pine WE (2011) Nonnative fish control in the Colorado River in Grand Canyon, Arizona: an effective program or serendipitous timing? Trans Am Fish Soc 140:456–470. https://doi.org/10.1080/00028487.2011.572009
Colautti RI, Ricciardi A, Grigorovich IA, MacIsaac HJ (2004) Is invasion success explained by the enemy release hypothesis? Ecol Lett 7:721–733. https://doi.org/10.1111/j.1461-0248.2004.00616.x
Colby PJ, Nepszy SJ (1981) Variation among stocks of Walleye (Stizostedion vitreum vitreum): management implications. Can J Fish Aquat Sci 38:1814–1831. https://doi.org/10.1139/f81-228
Cooch EG, Rockwell RF, Cooke F (1989) Long-term decline in fecundity in a Snow Goose population: evidence for density dependence? J Anim Ecol 58:711–726
Coulson T, Milner-Gulland EJ, Clutton-Brock T (2000) The relative roles of density and climatic variation on population dynamics and fecundity rates in three contrasting ungulate species. Proc R Soc Lond B Biol Sci 267:1771–1779. https://doi.org/10.1098/rspb.2000.1209
Dauphinais JD, Miller LM, Swanson RG, Sorensen PW (2018) Source–sink dynamics explain the distribution and persistence of an invasive population of common carp across a model Midwestern watershed. Biol Invasions 20:1961–1976. https://doi.org/10.1007/s10530-018-1670-y
Day CC, Landguth EL, Bearlin A et al (2018) Using simulation modeling to inform management of invasive species: a case study of eastern brook trout suppression and eradication. Biol Conserv 221:10–22. https://doi.org/10.1016/j.biocon.2018.01.017
De Roos AM, Schellekens T, van Kooten T et al (2007) Food-dependent growth leads to overcompensation in stage-specific biomass when mortality increases: the influence of maturation versus reproduction regulation. Am Nat 170:E59–E76. https://doi.org/10.1086/520119
DeMartini EE, Ellis DM, Honda VA (1993) Comparisons of spiny lobster Panulirus marginatus fecundity, egg size, and spawning frequency before and after exploitation. Fish Bull 91:1–7
Einum S, Sundt-Hansen L, Nislow KH (2006) The partitioning of density-dependent dispersal, growth and survival throughout ontogeny in a highly fecund organism. Oikos 113:489–496
Freeman MA, Turnbull JF, Yeomans WE, Bean CW (2010) Prospects for management strategies of invasive crayfish populations with an emphasis on biological control. Aquat Conserv Mar Freshw Ecosyst 20:211–223. https://doi.org/10.1002/aqc.1065
Gherardi F, Aquiloni L, Diéguez-Uribeondo J, Tricarico E (2011) Managing invasive crayfish: is there a hope? Aquat Sci 73:185–200. https://doi.org/10.1007/s00027-011-0181-z
Glen AS, Atkinson R, Campbell KJ et al (2013) Eradicating multiple invasive species on inhabited islands: the next big step in island restoration? Biol Invasions 15:2589–2603. https://doi.org/10.1007/s10530-013-0495-y
Govindarajulu P, Altwegg R, Anholt BR (2005) Matrix model investigation of invasive species control: bullfrogrs on Vancouver Island. Ecol Appl 15:2161–2170. https://doi.org/10.1890/05-0486
Green SJ, Grosholz ED (2021) Functional eradication as a framework for invasive species control. Front Ecol Environ 19:98–107. https://doi.org/10.1002/fee.2277
Grosholz E, Ashton G, Bradley M et al (2021) Stage-specific overcompensation, the hydra effect, and the failure to eradicate an invasive predator. Proc Natl Acad Sci 118:e2003955118. https://doi.org/10.1073/pnas.2003955118
Hansen GJA, Hein CL, Roth BM et al (2013a) Food web consequences of long-term invasive crayfish control. Can J Fish Aquat Sci 70:1109–1122. https://doi.org/10.1139/cjfas-2012-0460
Hansen GJA, Ives AR, Vander Zanden MJ, Carpenter SR (2013b) Are rapid transitions between invasive and native species caused by alternative stable states, and does it matter? Ecology 94:2207–2219. https://doi.org/10.1890/13-0093.1
He M, Liu F, Wang F (2021) Quantitative analysis of density dependent resource utilization, cannibalism, and competition of the red swamp crayfish (Procambarus clarkii) in rice-crayfish cocultures without supplementary food. Aquaculture 543:736966. https://doi.org/10.1016/j.aquaculture.2021.736966
Hefti D, Stucki P (2006) Crayfish management for Swiss waters. Bull Fr Pêhe Piscic 380–81:937–950
Hein CL, Roth BM, Ives AR, Zanden MJV (2006) Fish predation and trap** for rusty crayfish (Orconectes rusticus) control: a whole-lake experiment. Can J Fish Aquat Sci 63:383–393. https://doi.org/10.1139/f05-229
Hein CL, Vander Zanden MJ, Magnuson JJ (2007) Intensive trap** and increased fish predation cause massive population decline of an invasive crayfish. Freshw Biol 52:1134–1146. https://doi.org/10.1111/j.1365-2427.2007.01741.x
Hill AM, Lodge DM (1999) Replacement of resident crayfishes by an exotic crayfish: the roles of competition and predation. Ecol Appl 9:678–690
Houghton RJ, Wood C, Lambin X (2017) Size-mediated, density-dependent cannibalism in the signal crayfish Pacifastacus leniusculus (Dana, 1852) (Decapoda, Astacidea), an invasive crayfish in Britain. Crustaceana 90:417–435. https://doi.org/10.1163/15685403-00003653
Howald G, Donlan CJ, Galván JP et al (2007) Invasive rodent eradication on islands. Conserv Biol 21:1258–1268. https://doi.org/10.1111/j.1523-1739.2007.00755.x
Hoxie BJ (2009) Crib dock permit and construction standards in the Upper Great Lakes. Doctoral Dissertation, Eastern Michigan University
Jerry DR, Stewart T, Purvis IW, Piper LR (2001) Evaluation of visual implant elastomer and alphanumeric internal tags as a method to identify juveniles of the freshwater crayfish, Cherax destructor. Aquaculture 193:149–154
Kaemingk MA, Galarowicz TL, Clevenger JA, Clapp DF (2011) Movement of smallmouth bass within the Beaver Island Archipelago, northern Lake Michigan. J Gt Lakes Res 37:625–631. https://doi.org/10.1016/j.jglr.2011.08.005
Keane R (2002) Exotic plant invasions and the enemy release hypothesis. Trends Ecol Evol 17:164–170. https://doi.org/10.1016/S0169-5347(02)02499-0
Kvistad J, Galarowicz T, Clapp D et al (2021a) An evaluation of three trap designs for invasive rusty crayfish (Faxonius rusticus) suppression on critical fish spawning habitat in northern Lake Michigan. Manag Biol Invasions 12:975–996. https://doi.org/10.3391/mbi.2021.12.4.12
Kvistad JT, Buckley JT, Robinson KM et al (2021b) Size segregation and seasonal patterns in rusty crayfish Faxonius rusticus distribution and abundance on northern Lake Michigan spawning reefs. J Gt Lakes Res 47:1050–1064. https://doi.org/10.1016/j.jglr.2021.03.009
Liermann M, Hilborn R (1997) Depensation in fish stocks: a hierarchic Bayesian meta-analysis. Can J Fish Aquat Sci 54:1976–1984. https://doi.org/10.1139/f97-105
Lodge DM, Deines A, Gherardi F et al (2012) Global introductions of crayfishes: evaluating the impact of species invasions on ecosystem services. Annu Rev Ecol Evol Syst 43:449–472. https://doi.org/10.1146/annurev-ecolsys-111511-103919
Ludgate B, Closs G (2003) Responses of fish communities to sustained removals of perch (Perca fluviatilis). Sci Conserv 210
Manfrin C, Souty-Grosset C, Anastácio PM et al (2019) Detection and control of invasive freshwater crayfish: from traditional to innovative methods. Diversity 11:5. https://doi.org/10.3390/d11010005
Milligan WR, Jones MT, Kats LB et al (2017) Predicting the effects of manual crayfish removal on California newt persistence in Santa Monica Mountain streams. Ecol Model 352:139–151. https://doi.org/10.1016/j.ecolmodel.2017.02.014
Mitchell CE, Power AG (2003) Release of invasive plants from fungal and viral pathogens. Nature 421:625–627. https://doi.org/10.1038/nature01317
Moody KN, Scherer AE, O’Connor DAJS et al (2021) Effectiveness and outcomes of invasive species removal in Hawaiian streams. Biol Invasions 23:1739–1763. https://doi.org/10.1007/s10530-021-02468-w
Morse JW, Baldridge AK, Sargent LW (2013) Invasive crayfish Orconectes rusticus (Decapoda, Cambaridae) is a more effective predator of substrate nesting fish eggs than native crayfish (O. virilis). Crustaceana 86:387–402. https://doi.org/10.1163/15685403-00003187
Myers RA, Barrowman NJ, Hutchings JA, Rosenberg AA (1995) Population dynamics of exploited fish stocks at low population levels. Science 269:1106–1108. https://doi.org/10.1126/science.269.5227.1106
N’Guyen A, Hirsch PE, Bozzuto C et al (2018) A dynamical model for invasive round goby populations reveals efficient and effective management options. J Appl Ecol 55:342–352. https://doi.org/10.1111/1365-2664.12934
Olden J, McCarthy J, Maxted J et al (2006) The rapid spread of rusty crayfish (Orconectes rusticus) with observations on native crayfish declines in Wisconsin (U.S.A.) over the past 130 years. Biol Invasions 8:1621–1628. https://doi.org/10.1007/s10530-005-7854-2
Parsons W, Zhong W, Rudolf VHW (2013) Mating status and kin recognition influence the strength of cannibalism. Anim Behav 85:365–369. https://doi.org/10.1016/j.anbehav.2012.11.006
Peay S (2009) Invasive non-indigenous crayfish species in Europe: Recommendations on managing them. Knowl Manag Aquat Ecosyst. https://doi.org/10.1051/kmae/2010009
Peters JA, Cooper MJ, Creque SM et al (2014) Historical changes and current status of crayfish diversity and distribution in the Laurentian Great Lakes. J Gt Lakes Res 40:35–46. https://doi.org/10.1016/j.jglr.2014.01.003
Polis G (1981) The evolution and dynamics of intraspecific predation. Annu Rev Ecol Syst 12:225–251
Post JR, Parkinson EA, Johnston NT (1999) Density-dependent processes in structured fish populations: interaction strengths in whole-lake experiments. Ecol Monogr 69:155–175. https://doi.org/10.1890/0012-9615(1999)069[0155:DDPISF]2.0.CO;2
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ricker WE (1954) Stock and recruitment. J Fish Res Board Can 11:559–623
Robinson KM (2014) Spatial and seasonal distribution of the invasive Round Goby (Neogobius melanostomus and Rusty Crayfish (Orconectes rusticus) on critical nearshore spawning reefs in northern Lake Michigan. Master’s Thesis, Central Michigan University
Sakai AK, Allendorf FW, Holt JS et al (2001) The population biology of invasive species. Annu Rev Ecol Syst 32:305–332. https://doi.org/10.1146/annurev.ecolsys.32.081501.114037
Scalici M, Chiesa S, Scuderi S et al (2010) Population structure and dynamics of Procambarus clarkii (Girard, 1852) in a Mediterranean brackish wetland (Central Italy). Biol Invasions 12:1415–1425. https://doi.org/10.1007/s10530-009-9557-6
Scheffer M, Carpenter S, Foley JA et al (2001) Catastrophic shifts in ecosystems. Nature 413:591–596. https://doi.org/10.1038/35098000
Simberloff D (2021) Maintenance management and eradication of established aquatic invaders. Hydrobiologia 848:2399–2420. https://doi.org/10.1007/s10750-020-04352-5
Stephens PA, Sutherland WJ, Freckleton RP (1999) What is the Allee effect? Oikos 87:185–190
Stewart-Oaten A, Murdoch WW, Parker KR (1986) Environmental impact assessment: “pseudoreplication” in time? Ecology 67:929–940. https://doi.org/10.2307/1939815
Stuecheli K (1991) Trap** bias in sampling crayfish with baited funnel traps. N Am J Fish Manag 11:236–239. https://doi.org/10.1577/1548-8675(1991)011%3c0236:TBISCW%3e2.3.CO;2
Syslo JM, Guy CS, Bigelow PE et al (2011) Response of non-native lake trout (Salvelinus namaycush) to 15 years of harvest in Yellowstone Lake, Yellowstone National Park. Can J Fish Aquat Sci 68:2132–2145. https://doi.org/10.1139/f2011-122
Tetzlaff JC, Roth BM, Weidel BC, Kitchell JF (2011) Predation by native sunfishes (Centrarchidae) on the invasive crayfish Orconectes rusticus in four northern Wisconsin lakes: native fish predation on invasive crayfish. Ecol Freshw Fish 20:133–143. https://doi.org/10.1111/j.1600-0633.2010.00469.x
Thresher RE, Hayes K, Bax NJ et al (2014) Genetic control of invasive fish: technological options and its role in integrated pest management. Biol Invasions 16:1201–1216. https://doi.org/10.1007/s10530-013-0477-0
Torchin ME, Lafferty KD, Dobson AP et al (2003) Introduced species and their missing parasites. Nature 421:628–630. https://doi.org/10.1038/nature01346
Twardochleb LA, Olden JD, Larson ER (2013) A global meta-analysis of the ecological impacts of nonnative crayfish. Freshw Sci 32:1367–1382. https://doi.org/10.1899/12-203.1
Underwood AJ (1992) Beyond BACI: the detection of environmental impacts on populations in the real, but variable, world. J Exp Mar Biol Ecol 161:145–178. https://doi.org/10.1016/0022-0981(92)90094-Q
Van Buskirk J (1989) Density-dependent cannibalism in larval dragonflies. Ecology 70:1442–1449. https://doi.org/10.2307/1938203
Ward-Fear G, Brown GP, Greenlees MJ, Shine R (2009) Maladaptive traits in invasive species: in Australia, cane toads are more vulnerable to predatory ants than are native frogs. Funct Ecol 23:559–568. https://doi.org/10.1111/j.1365-2435.2009.01556.x
Ward-Fear G, Brown GP, Shine R (2010) Using a native predator (the meat ant, Iridomyrmex reburrus ) to reduce the abundance of an invasive species (the cane toad, Bufo marinus ) in tropical Australia. J Appl Ecol 47:273–280. https://doi.org/10.1111/j.1365-2664.2010.01773.x
Weber MJ, Hennen MJ, Brown ML et al (2016) Compensatory response of invasive common carp Cyprinus carpio to harvest. Fish Res 179:168–178. https://doi.org/10.1016/j.fishres.2016.02.024
Weidel BC, Josephson DC, Kraft CE (2007) Littoral fish community response to Smallmouth Bass removal from an Adirondack Lake. Trans Am Fish Soc 136:778–789. https://doi.org/10.1577/T06-091.1
Zipkin EF, Sullivan PJ, Cooch EG et al (2008) Overcompensatory response of a smallmouth bass (Micropterus dolomieu) population to harvest: release from competition? Can J Fish Aquat Sci 65:2279–2292. https://doi.org/10.1139/F08-133
Zipkin EF, Kraft CE, Cooch EG, Sullivan PJ (2009) When can efforts to control nuisance and invasive species backfire? Ecol Appl 19:1585–1595
Acknowledgements
We are grateful to Dana Castle and John Milan for help with data collection and field work organization. We also thank Tyler Punke, Taylor Rathbun, Kyle Urban, Trevor Gronda, Erin Prior, Kaitlyn McKnight, Helen Watrous, and Makinzee Castle for field assistance. This work was supported by the Great Lakes Restoration Initiative [Grant No.: GL00E02222] and the Earth and Ecosystem Science doctoral program at Central Michigan University. Thank you to Nick Catanzaro for GIS support. We are also grateful to two anonymous reviewers whose feedback improved the quality of the manuscript.
Funding
Funding was provided by U.S. Environmental Protection Agency (Grant No. GL00E02222).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Kvistad, J.T., Galarowicz, T.L., Clapp, D.F. et al. Evidence of a compensatory response in invasive Rusty Crayfish (Faxonius rusticus) following intensive harvest removal from northern Lake Michigan fish spawning reefs. Biol Invasions 25, 2831–2847 (2023). https://doi.org/10.1007/s10530-023-03076-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-023-03076-6