
Abstract
In recent years, the immunoderivative (IMiD) agents have been extensively used for the treatment of multiple myeloma (MM). IMiDs and their newer derivatives CRBN E3 ligase modulator bind the E3 ligase substrate recognition adapter protein cereblon (CRBN), which has been recognized as one of the IMiDs’ direct target proteins, and it is essential for the therapeutic effect of these agents.
High expression of CRBN was associated with improved clinical response in patients with MM treated with IMiDs, further confirming that the expression of IMiDs’ direct target protein CRBN is required for the anti-MM activity. CRBN’s central role as a target of IMiDs suggests potential utility as a predictive biomarker of response or resistance to IMiDs therapy. Additionally, the presence of alternatively spliced variants of CRBN in MM cells, especially those lacking the drug-binding domain for IMiDs, raise questions concerning their potential biological function, making difficult the transcript measurement, which leads to inaccurate overestimation of full-length CRBN transcripts. In sight of this, in the present study, we evaluated the CRBN expression, both full-length and spliced isoforms, by using real-time assay data from 87 patients and RNA sequencing data from 50 patients (n = 137 newly diagnosed MM patients), aiming at defining CRBN’s role as a predictive biomarker for response to IMiDs-based induction therapy. We found that the expression level of the spliced isoform tends to be higher in not-responding patients, confirming that the presence of a more CRBN spliced transcript predicts for lack of IMiDs response.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The use of immunomodulatory drugs (IMiDs) have brought many advantages in the treatment of hematological disease particularly in patients with multiple myeloma (MM) [1]. Despite the teratogenic effect [2], the IMiDs’ molecular mechanism of action proves an important therapeutic outcome on MM, since IMiDs both interfere with the growth and the development of neoplastic cells and stimulate the bone marrow environment to produce factors with anti-angiogenic, pro-apoptotic, anti-proliferative, and anti-inflammatory properties.
A major breakthrough in deciphering the molecular mechanism of IMiDs came using zebrafish and chicken embryo modeling, with the discovery that thalidomide bounds directly to cereblon (CRBN) and that this interaction was necessary for its teratogenic effects [2]. Soon after, it was shown that CRBN expression was required for the antimyeloma activity of IMiDs [3]. CRBN is part of a multi-proteic complex called CRL4CRBN, an E3 ubiquitin ligase, which includes damage DNA binding protein 1 (DDB1), Cullin 4 (Cul-4), and regulator of Cullin1 (ROC1) [4, 5]. In physiological conditions, this complex exerts both an auto-ubiquitination activity as well as the ubiquitination of intracellular substrates, which direct them toward a proteasome-mediated degradation [6]. Targeting CRBN by IMiDs modifies its substrate specificity toward non-physiological proteins which are subsequently ubiquitinated and degraded by the proteasome [5, 7,8,9,10]. Specifically, in the presence of IMiDs, the complex becomes unable to auto-ubiquitinate and redirects its ubiquitination activity toward novel targets, among which are Ikaros and Aiolos (IKZF1 and IKZF3, respectively), two transcription factors mainly involved in the lymphopoiesis [11, 12]. One of the downstream effects of IKZF1 and IKZF3 degradation is the inhibition of IRF-4, which finally results cytotoxic for MM cells [13]. CRBN, acting as the CRL4CRBN complex’ receptor, is essential for the anti-tumoral activity of the IMiDs [3]. Moreover, IMiD drug-induced degradation of Ikaros and Aiolos also occurs in T lymphocytes and is involved in T-cell co-stimulation via down-regulation of interleukin-2 transcription [10]. CSNK1A1 (CK1α) was also identified as a lenalidomide-specific neosubstrate with clinical relevance to del(5q) myelodysplastic syndrome [14, 15]. Additionally, GSPT1 is a CC-885-specific neosubstrate that mediates the broad-spectrum anticancer proprieties of CC-885 [16]. Notably, genomic defects in CRBN, including mutations, copy number loss, transcriptomic aberrations (epigenetic, RNA splicing/stability), and specific exon10-deleted splice transcript variant, increase in IMiD-resistant relapsed and refractory MM (RRMM) patients. Indeed, almost one-third of MM patients have genetic alterations in CRBN by the time they are refractory to IMiDs (both lenalidomide and pomalidomide), and these genetic changes are associated with inferior outcomes [2, 17,18,19,20,21]. The CRBN gene contains 11 exons encoding a protein comprising 442 amino acids residues with its C-terminal portion (encoded in part by exon10) containing the drug-biding domain. CRBN spliced variant has been associated with lenalidomide refractoriness, and unlike mutation or copy loss, CRBN exon10 splice isoform was observed in newly diagnosed MM (NDMM) patients at ~ 2.9% prevalence which increased to almost 30% in IMiD-resistant RRMM and was a prognostic marker for poor outcome in both disease setting [18]. Disruption of CRBN activity is the best understood mechanism of IMiD resistance in MM. As CRBN is necessary for IMiD activity [3], either quantitative or qualitative CRBN defects may contribute to IMiD ineffectiveness or resistance. It has been shown that the efficacy of a thalidomide analog may be influenced by both the level of CRBN expression and the levels of alternative substrates [7, 22, 23]. Substrate competition is a novel mediator of resistance to this class of drugs, whereby increased expression of a secondary protein drug target unrelated to canonical drug activity can lead to resistance [24]. Indeed, ARID2 it has been proposed as a pomalidomide-dependent CRL4CRBN substrate in MM cells, and its degradation is necessary for the anti-MM activity of pomalidomide. Several lines of evidence indicated that pomalidomide-induced ARID2 degradation is mediated by BRD7, suggesting a new mechanism of substrate recognition by CRL4CRBN [25]. The authors shown that pomalidomide can induce formation of the CRBN–BRD7–ARID2 trimeric complex even if CRBN–BRD7 and BRD7–ARID2 interactions are essentially constitutive. BRD7 exists either in a free form or an ARID2-bound form at an unknown ratio. In the absence of pomalidomide, the two forms bind equally to CRBN: in the presence of pomalidomide, however, the ARID2-bound form preferentially binds to CRBN [25]. Aberrations to any or all of these factors could cause IMiD resistance.
Nevertheless, the biological basis for the prognostic and/or predictive role of CRBN (e.g., as a biomarker of resistance to IMiDs) is still unknown.
Here, we analyzed a cohort of NDMM patients who received an IMiDs induction therapy and we evaluated the total CRBN expression level at baseline, to establish its role, already at diagnosis, as biomarker to predict the response to IMiDs therapy. Particular attention has been paid to the presence of different CRBN isoforms, especially those missing the IMiDs binding domain (i.e., CRBN lacking the exon 10), which might alter the quantification results, but also allegedly impact on patients’ outcome. Gene expression profiles of patients, either responding to IMiDs or not based therapy, were also determined to identify putative pathways involved in IMiDs resistance.
Materials and methods
Patients
An overall number of 137 of diagnostic samples from newly diagnosed MM (NDMM) patients, belonging to two different datasets, were enrolled in this study. Briefly, Bologna cohort (MM-BO) includes a total of 87 NDMM, homogeneously treated with IMiDs-based induction therapy (Thalidomide–Dexamethasone (TD, n = 86) or Bortezomib plus TD (VTD, n = 1) regimens), and then exposed to consolidation therapy after autologous peripheral blood stem cell transplantation (ASCT). The remaining patients (n = 50) were enrolled in Multiple Myeloma Research Foundation (MMRF) CoMMpass trial [26], and samples were selected based on IMiDs first-line therapy (Elotuzumab–Lenalidomide–Dexamethasone, n = 2, Lenalidomide, n = 4, Lenalidomide–BIAXIN–Dexamethasone, n = 1, Lenalidomide–Dexamethasone, n = 40, Lenalidomide–Methylprednisolone, n = 1, Lenalidomide–Prednisone, n = 1, Thalidomide–Melphalan–Prednisone, n = 1). Three patients treated with Lenalidomide–Dexamethasone induction therapy were then exposed to ASCT. The MMRF CoMMpass study is a prospective observational clinical trial (NCT01454297) with comprehensive genomic and transcriptomic characterization of NDMM patients, funded and managed by the MMRF. The study is ongoing, with data released regularly for research use via the MMRF research gateway, https://research.themmrf.org. In this study, we used Interim Analysis (IA) 18. Response to therapy was evaluated according to the International Myeloma Working Group criteria (IMWG) [27]. Patients’ baseline clinical and cytogenetic characteristics are summarized in Table 1.
Cells isolation
Mononuclear cells from diagnostic bone marrow (BM) aspirates were isolated by density gradient centrifugation over Ficoll-Paque Plus (Amersham Biosciences, Piscataway, NJ, USA). BM-CD138+ cells isolation and purity evaluation were performed as previously published [28,29,30]. Briefly, the purity of MM cells was confirmed by flow cytometry, and only cases enriched for more than 85% were included in the study. Patients’ samples were collected after informed consent.
RNA isolation and quantitative real-time PCR (qPCR)
Total RNA was isolated using RNeasy total RNA isolation kit (QIAGEN, Valencia, CA) with an automated RNA extraction method according to the manufacturer’s instructions (QIAcube, QIAGEN, Valencia, CA). 100 ng of total RNA was reverse transcribed using SuperScript™ III First-Strand Synthesis System and random hexamers (Inivtrogen Life Technologies). All reactions were performed in triplicate using the LightCycler 480 Instrument (Roche, Applied science) in a total volume of 20 μl. An absolute quantification was performed using probe assays (PrimeTime® qPCR 5′ Nuclease Assay, IDT) both for CRBN full-length (FL), using a standard pre-designed assay with the probe localized between exons 9 and 11, and for CRBN isoform, lacking exons 8 and 10 (henceforth called CRBN exon10-spliced), with the probe localized between exons junction 7–9. GAPDH was used as endogenous control to normalize the level expression of CRBN. CRBN available for IMiDs binding was expressed for each patient as ratio between CRBN exon10-spliced and CRBN-FL isoforms. Analysis of relative transcript levels was calculated using the delta-delta Ct method (ΔΔCT). For CoMMpass dataset, CRBN expression analysis was performed by using RNA sequencing (RNAseq) online data of diagnostic samples.
CoMMpass gene expression data analysis
Together with MM-BO results, RNAseq expression data about the two CRBN isoforms from CoMMpass project were integrated. Specifically, we selected the transcript-level counts of the isoform ENST00000231948 (CRBN-FL) and ENST00000424814 (CRBN exon10-spliced) from baseline CD138 + samples of 50 patients having first-line IMID-based therapy. Instead, the gene-level count matrices were used to perform a differential expression gene (DEG) analysis with the R package DEseq2 [31], pre-filtering only genes that have at least 5 reads total. Following, to study the pathways of the biological processes that were mainly expressed or not in the compared patient groups, a gene set enrichment analysis (GSEA) was done with R package clusterProfiler [32].
Clinical and statistical analysis
Ratios CRBN exon10-spliced/CRBN-FL (mRNA) were correlated to response to therapy after front-line IMiDs-based induction treatment. CRBN ratios were calculated based on the CRBN gene expression data derived from qPCR (MM-BO dataset) and RNA sequencing (RNAseq for CoMMpass samples) data. Patients were clustered according to their response to induction therapy (first response), and patients achieving at least Very Good Partial Response (VGPR) were compared to patients failing this objective. Ratios of the two populations were compared through Wilcoxon rank sum test. Cox regression model was used to assess the relationship between risk factors and patient’s survival times. Survival curves of each model were visualized with the Kaplan–Meier method and compared with long-rank test. Fisher’s test and generalized linear model (GLM) were chosen for univariate and multivariate analyses, respectively. All statistical analyses were performed with R software with survival, survminer, gmodels, and stats packages.
Results
Baseline CRBN isoforms expression level in newly diagnosed MM patients
We first determined the baseline expression of CRBN isoforms by analyzing quantitative real-time PCR (qPCR) data from 87 and RNA sequencing (RNASeq) data from 50 NDMM patients (MM-BO and CoMMpass cohort, respectively) who received an IMiD-based induction therapy (summary patients characteristics are listed in Supplementary Table 1). A comparison of the patients’ baseline characteristics revealed no significant differences between the IMiD-based treatment groups, except for higher rates of male/female patients (60% vs. 40% and 56% vs. 44% in MM-BO and CoMMpass dataset, respectively) (Table 1).
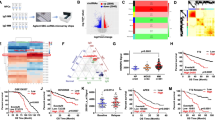
Overall, CRBN expression levels of both CRBN-FL and CRBN exon10-spliced isoforms were lower in our dataset compared to CoMMpass subgroup (Supplementary Table 1), differences mainly ascribed to the methods used for measuring CRBN transcript. In particular, the analysis of the CRBN exon10-spliced isoform mRNA level in the two datasets highlighted a wide range of deleted isoform expression values (for MM-BO dataset range 0.0000260–0.004549, and for CoMMpass dataset range 0.104776–3.790971, respectively). We then determined the optimum sample cut-off for both CRBN isoforms by using the medians value in each dataset to establish the high and low-expression range (MM-BO: CRBN exon10-spliced 0.000494 and CRBN-FL 0.021798, respectively; CoMMpass: CRBN exon10-spliced 0.774215 and CRBN-FL 4.944441, respectively). According to the established cut-offs, we compared the distribution of CRBN isoforms between samples in each dataset, and we found that CRBN exon10-spliced expression did not significantly vary between cases with high vs. low CRBN-FL isoform in the two cohorts (Fig. 1).

Baseline expression levels of CRBN isoforms in newly diagnosed MM patients. Violin plots show the level of CRBN-FL and CRBN exon10-spliced variants in MM-BO (n = 87) and CoMMpass (n = 50) datasets. CRBN isoforms are expressed as absolute mRNA transcript level. Overall, CRBN exon10-spliced variant does not change with respect to the level of full-length isoform in the two subgroups. High FL = high CRBN-FL, low FL = low CRBN-FL
The ratio between CRBN exon10-spliced and CRBN-FL transcripts correlate with response to therapy
According to the response rate evaluated referring to the IMWG criteria [27], all patients (n = 137) were stratified in “responders” (R) (i.e., achieving at least a Very Good Partial Response, VGPR) and “non-responders” (NR) (i.e., achieving a suboptimal response, < VGPR).
Overall, 54 out of 137 patients (i.e., 39.42%) were defined as R (10 achieved Complete Response, CR, 13 achieved near Complete Response, nCR, and 31 achieved VGPR). The remaining 83 patients (i.e., 60.58%) were included in the NR group (27 in Stable Disease, SD, 48 achieved Partial Response, PR, and 8 achieved Progressive Disease, PD). Most of the patients received an IMiDs-based induction therapy (n = 115, with Lenalidomide n = 29 or Thalidomide n = 86), whereas the remaining patients received a combined Bortezomib-IMiDs-based first-line therapy (n = 22) (patients characteristic summarized in Supplementary Table 1). We compared the CRBN isoforms expression level between subgroups (R vs. NR), and we found that 57% (31 out of 54) of R patients showed lower expression of CRBN exon10-spliced isoform compared to NR (45%, 37 out of 83), whereas 59% (32 out of 54) showed a higher CRBN-FL isoform level respect to NR patients (45%, 37 out of 83) (Fig. 2A). Conversely, NR patients showed lower CRBN-FL isoform level (55%, 46 out of 83) compared to R patients (41%, 22 out of 54). Although we did not observe significant differences in terms of transcript level between the two patients’ subgroups (Fisher’s exact test, Low vs High CRBN exon10-spliced p = 0.1640 and Low vs High CRBN-FL p = 0.1161), these results suggested that a higher percentage of deleted CRBN predicted for lack of IMiDs response, even though some overlaps still persist between the expression levels of the deleted CRBN isoform among the two subgroups of patients.

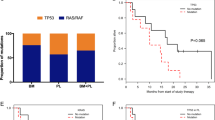
Distribution of CRBN isoforms and correlation with response to IMiDs first-line induction therapy. A distribution of CRBN-FL and CRBN exon10-spliced in MM patients stratified, according to the response rate evaluated referring to the IMWG criteria13 in responder and non-responder. The definition of high- or low-expression CRBN mRNA level of both variants was defined based on the median value calculated on all MM samples (n = 137). B Change in ratio of CRBN exon10-spliced/CRBN-FL transcripts between non-responder and responder patients. Significant differences detected by Wilcoxon signed-rank test (nonparametric)
Considering that the ratio of CRBN exon10-spliced (ENST00000424814.5)/CRBN-FL (ENST00000231948.8) has previously been proposed to correlate with outcome after IMiD-based regimens [18], we evaluated the response to induction therapy of these patients based on the ratio of transcript levels of spliced CRBN to that of CRBN-FL. We used the median value of the ratio calculated on all 137 MM samples to determine the optimum cut-off to distinguish patients with high vs low ratio. As shown in Fig. 2B, using a cut-off ratio of 0.0595, we found that the ratio was significantly lower in R patients as compared to NR patients (Wilcoxon test, p = 0.0084), suggesting a tendency drives by CRBN-FL isoform-carrying patients to respond to IMiDs-based induction therapy. To verified this hypothesis, we performed a subgroup analysis by stratifying R and NR patients in different categories based both on their specific response to treatment (i.e., CR, nCR, VGPR and SD, PR, and PD, respectively), and the CRBN ratio level (i.e., high vs low ratio). We observed an increasing number of patients with a high CRBN ratio as the response to therapy become suboptimal (data not shown, p = 0.0097), confirming the need of a high amount of CRBN-FL isoform to achieve an optimal response. Subsequently, to further validate this observation, we assessed whether the distribution of CRBN transcripts, both spliced and full-length, defined as high or low level (i.e., high CRBN-FL and/or CRBN exon10-spliced and low CRBN-FL and/or CRBN exon10-spliced), was different between R vs NR. As shown in Fig. 3, most of the patients achieving at least a VGPR or better response have a higher percentage of CRBN-FL isoform compared to those achieving a suboptimal response, regardless of CRBN ratio level. Indeed, the expression level of the spliced isoform tends to be higher in NR patients, confirming that the higher presence of CRBN exon10-spliced transcript predicts for lack of IMiDs response.

Correlation between CRBN-FL and exon10-spliced transcript levels respect to ratio level, and response to IMiDs first-line induction therapy. Histograms show the distribution of both CRBN transcript variants, defined as low- and high-level, low- and high-ratio level and response to induction therapy. Groups: High CRBN-FL-high CRBN exon10-spliced-dark red, high CRBN-FL-low CRBN exon10-spliced-light red, low CRBN-FL-high CRBN exon10-spliced-dark blue, low CRBN-FL-low CRBN exon10-spliced-light blue. CR = complete response, nCR = near complete response, VGPR = very good partial response, PD = partial disease, PR = progression disease, SD = stable disease
Unfortunately, we observed that patients treated with PIs-IMID are mainly NR (81%, Fisher’s exact test 95% CI 1.041–14.82, p = 0.031), which prevents analysis in a homogeneous clinical context. However, no particular distribution of isoforms is observed between the two patient groups distinguished by treatment. Furthermore, in a multivariate model with treatment and ratio, the independent effect of the therapy was not confirmed (p = 0.124).
Correlation between CRBN isoforms expression and clinical outcomes
We examined the association between CRBN’s transcript levels and patients’ outcome. Firstly, we analyzed the clinical outcomes of patients based on CRBN-FL isoform levels, and we observed that progression free survival (PFS) was significantly longer in patients expressing high CRBN-FL (PFS: HR 0.6481 [95% CI 1.063–2.24], p = 0.021) but not in patients with high CRBN exon10-spliced variant (PFS: HR 1.1341 [95% CI 0.6096–1.275], p = 0.5), confirming the strong association of CRBN-FL isoform with higher response to therapy. Indeed, the baseline deleted CRBN expression level does not affect patient’s outcome, since both the PFS and OS of patients expressing higher levels of CRBN exon10-spliced isoform were not significantly different from those of patients expressing low levels (PFS: HR 0.887 [95% CI 0.6096–1.275], p = 0.5; OS: HR 0.9012 [95% CI 0.554–1.466], p = 0.675) (Fig. 4A and B). Remarkably, when patients were stratified according to CRBN ratio level in high vs low ratio, the presence of a high-ratio level was significantly related with worse patients’ outcome, whereas patients expressing a low ratio have a better outcome (PFS: HR 0.588 [95% CI 0.4024–0.8594], p = 0.0055; OS: HR 0.8407 [95% CI 0.5142–1.375], p = 0,49) (Fig. 5A). In a multivariate model, we evaluated the independent effects of CRBN-FL isoform levels and response to therapy on PFS, showing that response has a significant impact on relapse (NR: HR 1.86 [95% CI 0.36–0.81], p = 0.00262; CRBN-FL: HR 1.3297 [95% CI 0.91–1.95], p = 0.14635). However, it was observed that the combination of these variables allows further stratification of NR patients, as those with low CRBN-FL level and no response to therapy have a higher risk of relapse (HR 2.5 [95% CI 1.52–4.12] p = 0.000324, Fig. 5B), while NR patients with high CRBN-FL level have a lower risk (HR 1.93 [95% CI 1.13–3.31], p = 0.016109, Fig. 5B).

Correlation between CRBN isoforms expression and clinical outcome to IMiDs induction therapy in ND myeloma. A PFS and OS of non-responder versus responder patients. (PFS: HR 0.4999 [95% CI 0.3371–0.7415] p = 0.00042, OS: HR 0.6713 [95% CI 0.4027–1.119] p = 0.12). B PFS and OS of high CRBN-FL versus low CRBN-FL expressing patients. (PFS: HR 1.543 [95% CI 1.063–2.24] p = 0.021, OS: HR 1.4084 [95% CI 0.8631–2.298] p = 0.17). C PFS and OS of high CRBN exon10-spliced versus low CRBN exon10-spliced expressing patients. (PFS: HR 0.8817 [95% CI 0.6096–1.275] p = 0.5, OS: HR 0.9012 [95% CI 0.554–1.466] p = 0.67)

Correlation between CRBN ratio level and clinical outcome to IMiDs induction therapy in ND myeloma. A PFS and OS of high CRBN ratio vs low CRBN ratio. (PFS: HR 0.588 [95% CI 0.4024–0.8594] p = 0.0055, OS: HR 0.8407 [95% CI 0.5142–1.375] p = 0.49). B PFS curve of the patient groups determined by iteration between CRBN-FL levels and clinical response (p = 0.002). The risk of each group was calculated compared to the high CRBN-FL-R patient group (High CRBN-FL + NR, HR: 1.93 [95% CI = 1.13–3.31], p = 0.016109; Low CRBN-FL + R, HR: 1.4 [95% CI = 0.73–2.7], p = 0.309500; Low CRBN-FL + NR, HR: 2.5 [95% CI 1.52–4.12] p = 0.000324)
Additionally, to explore the prognostic impact of CRBN-FL levels, we performed a logistic regression model with those factors known to have an impact on response in MM patients and were available for at least 50% of patients. After assessing the correlation of the variables with the response in univariate, the only significant variable was first-line transplantation. As expected, in the multivariate analysis, the probability of first-line response is mainly determined by the use of transplantation in the treatment regimen (OR 3.62; p = 0.00135; CI 1.67943–8.16039), and by the presence of high levels of CRBN-FL in CD138 + cells (OR 2.58; p = 0.01347; CI 1.22746–5.55781) (Supplementary Table 2).
Taken together, these results demonstrated a positive correlation with outcome and CRBN-FL expression level, with low percentage of CRBN-FL transcript associated with higher probability to unfavorable response to induction therapy.
CRBN-FL low-expressing patients down-regulated pathways involved in immune response
Based on the results described above, we decided to perform an exploratory analysis by comparing the gene expression profile of patients (CoMMpass dataset, n = 50) expressing high levels of CRBN-FL to those with a low-expression level, to identify differentially expressed pathways and genes in CRBN-FL expressing patients, potentially related to response to therapy and eventually to IMiDs resistance.
The baseline clinical and cytogenetic characteristics of the two subgroups of patients were homogeneous, thus suggesting that only the different expression levels of CRBN might account for the differentially expressed gene profile (Table 1).
Both down- and up-regulated probe-sets were analyzed as described in Material and Methods section, to identify the biological processes, as well as the molecular functions which might be altered in patients expressing low levels of CRBN-FL. To this end, to identify genes set significantly deregulated in high CRBN-FL-expressing patients as compared to low-expressing ones, the gene-based expression estimation was used to perform a differential expression gene analysis (DEG) between the two subgroups of patients. Overall, 326 probe-sets resulted significantly differentially down-regulated in patients with lower versus those with higher amount of CRBN-FL (Fold Change, FC > 2 and p ≤ 0.05). As shown in the volcano plot (Fig. 6A) among the down-regulated genes, the most represented were involved in antigen binding activity and immunoglobulin receptor binding activity as well as activation of immune response, accounted mainly for the down-regulation of IGHV2-70D (FC = − 5.3772, p = 2.86e-07), IGHV3-21 (FC = − 39.556, p = 6.20e-10), and IGHV3 (FC = − 2.78443, p = 4.19e-9). Moreover, by gene set enrichment analysis (GSEA), a significant down-modulation of several pathways was observed, including multiple immune system-related signaling pathways, such as neutrophil activation involved in immune response (Enrichment Score, ES = − 0.43007, p = 1e-10), neutrophil-mediated immunity (ES = − 0.43447, p = 1e-10), positive regulation of leukocyte activation (ES = − 0.44334, p = 1e-10), granulocyte activation (ES = − 0.43266, p = 1e-10), and neutrophil activation (ES = − 0.43505, p = 1e-10) which were the most represented (Fig. 6B and C). Overall, these data suggest that plasma cells expressing low levels of CRBN-FL might have impaired the competence to correctly interact with other immune cells in the marrow, thus eventually implying a breakage of the MM cells and microenvironment cross-talk in these patients. To confirm this observation, we next compared the transcriptomic profile of low CRBN-FL NR patients with high CRBN-FL R patients, and we found the up-regulation of several genes involved in positive regulation of acute inflammatory response, (C2CD4A, FC = 4.2823, p = 0.00148) and involved in cytotoxic lymphocyte-mediated immunity (FGFBP2, FC = 37.915, p = 0.00858) (Fig. 7A). Notably, we observed that among the up-regulated pathways, predominantly, the immune system-related signaling pathways were significantly enriched in CRBN-FL R patients (e.g., leukocyte-mediated immunity ES = 0.41539, p = 4.89e-8, leukocyte-mediated cytotoxicity ES = 0.52510, p = 1.55e-6, lymphocyte-mediated immunity ES = 0.41791, p = 1.27e-6, B cell activation ES = 0.40423, p = 8.92e-6 and regulation of inflammatory response ES = 0.40030, p = 3.17e-6) (Fig. 7B and C).

Gene expression and gene set enrichment analysis of low CRBN-FL versus high CRBN-FL expressing patients. A Volcano plot of differential expression gene analysis (DESeq2). Labeled genes have a p-value < 0.001. B Transcriptional network of the five significantly regulated pathways. C Gene set enrichment analysis of the five significantly down-regulated pathways

Gene expression and gene set enrichment analysis of low CRBN-FL and NR versus high CRBN-FL R patients. A Volcano plot of differential expression gene analysis (DESeq2). Labeled genes have a p-value < 0.001. B Transcriptional network of the five significantly regulated pathways. C Gene set enrichment analysis of the five significantly up-regulated pathways
Overall, these data strongly support the connection between CRBN and immune system pathway suggesting a potential link with IMiD resistance.
Discussion
CRBN is the direct binding target of the IMiDs, one of the main classes of drugs in use in different MM phases. There is an increasing amount of evidence indicating that IMiD resistance arises, at least in part, from the acquisition and selection of genetic changes carried by CRBN. In this contest, the accurate quantification of CRBN mRNA level represents an issue, since CRBN transcript might undergo a complex sequence of alternative splicing, which results in the translation of several isoforms, frequently co-expressed in the same individual. This should be considered, when performing the analysis of the CRBN expression, since diverse isoforms have been described, which lack one or more of the CRBN protein functional domains. For instance, the CRBN binding domain of IMiDs, located in the C-terminal region (i.e., Thalidomide-Binding Domain, TBD), [2] is encoded by the CRBN exon 10, a region that might be alternatively spliced in MM patients [33, 34].
In this study, we demonstrated that CRBN transcript levels, both full-length and spliced isoforms, correlate with IMiDs response already at diagnosis, and this observation was in line with previously published data [19, 35]. The potential deletion of the drug-binding pocket may lead to a direct loss of drug-binding function of CRBN. However, it is unclear if the relative amount of CRBN-FL isoform present in the MM cells may be sufficient to confer drug sensitivity, and/or whether the presence of a high CRBN exon10-spliced variant in untreated patients may have a “dominant negative” impact. Indeed, results have been frequently conflicting, and so far, no standard and consistent assays have been set-up, to assess the exact CRBN expression level by quantitative PCR, principally for diagnostic purposes. Moreover, these methods actually allow for the quantification of CRBN-FL, but do not consider the possible presence of different isoforms [33, 36]. To the best of our knowledge, this study is the first attempt to quantify the absolute expression levels of both CRBN-FL and the CRBN exon10-spliced variant by analyzing the transcript data of diagnostic samples from NDMM patients who received an IMiDs-based induction therapy. We found that the level of CRBN exon10-spliced isoform was associated with inferior IMiD-based treatment response. Notably, patients with high CRBN-FL were more likely to improve depth of response compared to those with low level. Additionally, when comparing the ratio between CRBN exon10-spliced/CRBN-FL transcripts, we demonstrated that this ratio was inversely correlated with response to therapy. Remarkably, we observed that the ratio was significantly lower in R patients rather than NR patients, suggesting the need of high amount of CRBN-FL isoform to respond to IMiDs-based induction therapy. This speculation is further endorsed by the finding that patients achieving at least a VGPR or better response have a higher percentage of CRBN-FL isoform compared to those achieving a suboptimal response, regardless of CRBN ratio level. Moreover, both high CRBN-FL-expressing patients and low ratio patients were sufficiently represented at diagnosis, to enabling us to assess correlation, and they were associated with significantly improved PFS. These results strongly support the notion that the accurate expression of CRBN could potentially predict the clinical course of MM patients and identify, upfront, patients with a high probability of responding to IMiD-based regimens.
Newer derivatives of IMiDs, CELMoDs, bind target protein CRBN with greater affinity, leading to faster substrate degradation and therefore more potent anti-proliferative effects. Recent data suggest CELMoDs iberdomide (CC-220) and mezigdomide (CC92480) may have efficacy in lenalidomide- and pomalidomide-resistant patients in ongoing clinical trials [37,38,39]. The association of CELMoD compounds to the TBD has been shown to be both necessary and sufficient to trigger CRBN allosteric rearrangements, leading to the “closure” of the non-canonical “open” conformation [40]. Indeed, CELMoD agents bind to a conserved hydrophobic pocket in CRBN to create a molecular surface, able to recruit downstream target proteins [40, 41]. Importantly, just one of the most critical direct downstream CRBN’s targets (i.e., Ikaros) stably associates with the closed CRBN conformation, highlighting the importance of allostery for CELMoD compound efficacy. In this regard, the allosteric perturbations possibly introduced by mutations and/or aberrations of CRBN gene (i.e., presence of high amount of spliced isoform) should be considered as possible mechanism of drug resistance, in order to improve CELMoD’s clinical activity [40, 41].
Notably, the gene expression profile of CRBN-FL-carrying patients showed that among up-regulated pathways, the multiple immune system-related signaling pathways were significantly enriched in patients expressing high levels of CRBN-FL variant. These results highlighted an unexpected correlation between CRBN-FL level and immune pathway genes within MM cells of untreated patients. However, we cannot exclude that other factor, including transcriptional and/or post-transcriptional mechanisms, CRBN mutations and/or mutations in genes involved in CRBN’s pathways (i.e., in IKZF1 or IKZF3), reflecting the presence of sub-clonal mutations in genes encoding the splicing machinery [19, 21], might be also present and implicated in the acquisition of immunomodulatory drug resistance and subsequent clinical relapse; these additional factors should be further explored.
Moreover, we also analyzed the clinical outcomes of patients based on response to induction therapy (i.e., R vs NR), and, as expected, we observed a significant longer PFS in R subgroup compared to NR (PFS: HR 2 [95% CI 0.3371–0.7415]) p = 0.00042; conversely, we did not see a significant impact on overall survival (OS) (OS: HR 1.49 [95% CI 0.4027–1.119], p = 0.12) (data not shown). Finally, we showed a clear positive association between CRBN-FL expression and clinical outcome, highlighting the expression of high levels of CRBN exon10-spliced variant in NR patients, even though we did not explore the underlying mechanisms that cause such CRBN aberrations and result in drug resistance. The identification of a clear biological connection between the occurrence of CRBN aberrations and drug resistance was outside the aim of this investigation, and will be the subject of future analyses. Moreover, we acknowledge that the cut-off employed to identify patients with high CRBN exon10-spliced isoform might be more precisely defined on larger cohorts of patients. We also recognize that one of the pitfalls of this investigation is that patients included in this study have been treated with regimens that are no longer considered as up-to date, and that further analyses employing patients’ samples who will receive new drugs combinations therapy might strengthen the data.
Conclusions
Our results highlight the importance to define the amount of deleted CRBN isoforms, in order to employ CRBN as a biomarker of IMiDs response already at the diagnosis. To this end, it becomes clear that the precise quantification of CRBN variants levels is necessary to correctly determine both the “penetrance” and/or the optimal cut-off aim at defining upfront patients who will lack an IMiD response. What is the right balance between the two CRBN isoforms inside tumor cell needed for achieving a good response? This is still an open question that must be answered to have a complete overview and understanding of CRBN-related IMiD resistance mechanisms. In this scenario, it becomes clear that our data must be confirmed employing a large cohort of patients, to introduce a reliable approach in the routine clinical setting; in fact, the importance of a predictive biomarker in a complex genomic disease as MM is crucial, since it may lead to the establishment of personalized-treatment therapy.
Data availability
Bioinformatics datasets presented in this study can be found in online repositories, and the datasets used and/or analyzed during experiments are available from the corresponding author on reasonable request.
References
Chang X, Zhu Y, Shi C, Stewart AK. Mechanism of immunomodulatory drugs’ action in the treatment of multiple myeloma. Acta Biochim Biophys Sin. 2014;46(3):240–53.
Ito T, Ando H, Suzuki T, et al. Identification of a primary target of thalidomide teratogenicity. Science. 2010;327(5971):1345–50.
Zhu YX, Braggio E, Shi CX, et al. Cereblon expression is required for the antimyeloma activity of lenalidomide and pomalidomide. Blood. 2011;118(18):4771–9.
Chamberlain PP, Lopez-Girona A, Miller K, et al. Structure of the human Cereblon-DDB1-lenalidomide complex reveals basis for responsiveness to thalidomide analogs. Nat Struct Mol Biol. 2014;21(9):803–9.
Fischer ES, Bohm K, Lydeard JR, et al. Structure of the DDB1-CRBN E3 ubiquitin ligase in complex with thalidomide. Nature. 2014;512(7512):49–53.
Chang XB, Stewart AK. What is the functional role of the thalidomide binding protein cereblon? Int J Biochem Mol Biol. 2011;2(3):287–94.
Zhu YX, Braggio E, Shi CX, et al. Identification of cereblon-binding proteins and relationship with response and survival after IMiDs in multiple myeloma. Blood. 2014;124(4):536–45.
Lu G, Middleton RE, Sun H, et al. The myeloma drug lenalidomide promotes the cereblon-dependent destruction of Ikaros proteins. Science. 2014;343(6168):305–9.
Kronke J, Udeshi ND, Narla A, et al. Lenalidomide causes selective degradation of IKZF1 and IKZF3 in multiple myeloma cells. Science. 2014;343(6168):301–5.
Gandhi AK, Kang J, Havens CG, et al. Immunomodulatory agents lenalidomide and pomalidomide co-stimulate T cells by inducing degradation of T cell repressors Ikaros and aiolos via modulation of the E3 ubiquitin ligase complex CRL4(CRBN). Br J Haematol. 2014;164(6):811–21.
Yoshida T, Georgopoulos K. Ikaros fingers on lymphocyte differentiation. Int J Hematol. 2014;100(3):220–9.
Schwickert TA, Tagoh H, Gultekin S, et al. Stage-specific control of early B cell development by the transcription factor Ikaros. Nat Immunol. 2014;15(3):283–93.
Bjorklund CC, Lu L, Kang J, et al. Rate of CRL4(CRBN) substrate Ikaros and aiolos degradation underlies differential activity of lenalidomide and pomalidomide in multiple myeloma cells by regulation of c-Myc and IRF4. Blood Cancer J. 2015;5:e354.
Kronke J, Fink EC, Hollenbach PW, et al. Lenalidomide induces ubiquitination and degradation of CK1alpha in del(5q) MDS. Nature. 2015;523(7559):183–8.
Petzold G, Fischer ES, Thoma NH. Structural basis of lenalidomide-induced CK1alpha degradation by the CRL4(CRBN) ubiquitin ligase. Nature. 2016;532(7597):127–30.
Matyskiela ME, Lu G, Ito T, et al. A novel cereblon modulator recruits GSPT1 to the CRL4(CRBN) ubiquitin ligase. Nature. 2016;535(7611):252–7.
Franssen LE, Nijhof IS, Couto S, et al. Cereblon loss and up-regulation of c-Myc are associated with lenalidomide resistance in multiple myeloma patients. Haematologica. 2018;103(8):e368–71.
Gooding S, Ansari-Pour N, Towfic F, et al. Multiple cereblon genetic changes are associated with acquired resistance to lenalidomide or pomalidomide in multiple myeloma. Blood. 2021;137(2):232–7.
Kortum KM, Mai EK, Hanafiah NH, et al. Targeted sequencing of refractory myeloma reveals a high incidence of mutations in CRBN and Ras pathway genes. Blood. 2016;128(9):1226–33.
Lopez-Girona A, Mendy D, Ito T, et al. Cereblon is a direct protein target for immunomodulatory and antiproliferative activities of lenalidomide and pomalidomide. Leukemia. 2012;26(11):2326–35.
Thakurta A, Gandhi AK, Waldman MF, et al. Absence of mutations in cereblon (CRBN) and DNA damage-binding protein 1 (DDB1) genes and significance for IMiD therapy. Leukemia. 2014;28(5):1129–31.
Kronke J, Kuchenbauer F, Kull M, et al. IKZF1 expression is a prognostic marker in newly diagnosed standard-risk multiple myeloma treated with lenalidomide and intensive chemotherapy: a study of the german myeloma study group (DSMM). Leukemia. 2017;31(6):1363–7.
Pourabdollah M, Bahmanyar M, Atenafu EG, Reece D, Hou J, Chang H. High IKZF1/3 protein expression is a favorable prognostic factor for survival of relapsed/refractory multiple myeloma patients treated with lenalidomide. J Hematol Oncol. 2016;9(1):123.
Sperling AS, Burgess M, Keshishian H, et al. Patterns of substrate affinity, competition, and degradation kinetics underlie biological activity of thalidomide analogs. Blood. 2019;134(2):160–70.
Yamamoto J, Suwa T, Murase Y, et al. ARID2 is a pomalidomide-dependent CRL4(CRBN) substrate in multiple myeloma cells. Nat Chem Biol. 2020;16(11):1208–17.
Relating clinical outcomes in multiple myeloma to personal assessment of genetic Profile-Full Text View-ClinicalTrials.gov. [Internet]. Available from: https://clinicaltrials.gov/ct2/show/NCT01454297?term=NCT01454297&draw=2&rank=1.
Rajkumar SV. Updated diagnostic criteria and staging system for multiple myeloma. Am Soc Clin Oncol Educ Book. 2016;35:e418–23.
Borsi E, Terragna C, Brioli A, Tacchetti P, Martello M, Cavo M. Therapeutic targeting of hypoxia and hypoxia-inducible factor 1 alpha in multiple myeloma. Transl Res. 2015;165(6):641–50.
Martello M, Remondini D, Borsi E, et al. Opposite activation of the Hedgehog pathway in CD138+ plasma cells and CD138-CD19+ B cells identifies two subgroups of patients with multiple myeloma and different prognosis. Leukemia. 2016;30(9):1869–76.
Borsi E, Perrone G, Terragna C, et al. Hypoxia inducible factor-1 alpha as a therapeutic target in multiple myeloma. Oncotarget. 2014;5(7):1779–92.
Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15(12):550.
Wu T, Hu E, Xu S, et al. clusterProfiler 4.0: a universal enrichment tool for interpreting omics data. Innovation (Camb). 2021;2(3):100141.
Gandhi AK, Mendy D, Waldman M, et al. Measuring cereblon as a biomarker of response or resistance to lenalidomide and pomalidomide requires use of standardized reagents and understanding of gene complexity. Br J Haematol. 2014;164(2):233–44.
Lode L, Amiot M, Maiga S, et al. Cereblon expression in multiple myeloma: not ready for prime time. Br J Haematol. 2013;163(2):282–4.
Egan JB, Kortuem KM, Kurdoglu A, et al. Extramedullary myeloma whole genome sequencing reveals novel mutations in cereblon, proteasome subunit G2 and the glucocorticoid receptor in multi drug resistant disease. Br J Haematol. 2013;161(5):748–51.
Heintel D, Rocci A, Ludwig H, et al. High expression of cereblon (CRBN) is associated with improved clinical response in patients with multiple myeloma treated with lenalidomide and dexamethasone. Br J Haematol. 2013;161(5):695–700.
Paul G, Richardson EO, Raje NS, et al. CC-92480, a potent, novel cereblon E3 ligase modulator (CELMoD) agent, in combination with dexamethasone (DEX) and bortezomib (BORT) in patients (pts) with relapsed/refractory multiple myeloma (RRMM): preliminary results from the phase 1/2 study CC-92480-MM-002. Blood. 2021;138:2731.
Lonial S, Popat R, Hulin C, et al. Iberdomide plus dexamethasone in heavily pretreated late-line relapsed or refractory multiple myeloma (CC-220-MM-001): a multicentre, multicohort, open-label, phase 1/2 trial. Lancet Haematol. 2022;9(11):e822–32.
Bjorklund CC, Kang J, Amatangelo M, et al. Iberdomide (CC-220) is a potent cereblon E3 ligase modulator with antitumor and immunostimulatory activities in lenalidomide- and pomalidomide-resistant multiple myeloma cells with dysregulated CRBN. Leukemia. 2020;34(4):1197–201.
Edmond R, Watson PJN, Matyskiela ME, Chamberlain PP, et al. Molecular glue CELMoD compounds are allosteric regulators of cereblon conformation. Science. 2022;378(6619):549–53.
Hansen JD, Correa M, Nagy MA, et al. Discovery of CRBN E3 ligase modulator CC-92480 for the treatment of relapsed and refractory multiple myeloma. J Med Chem. 2020;63(13):6648–76.
Acknowledgements
The authors would like to thank all the Study Coordinators and the Research Groups involved in the Seràgnoli Institute, Bologna, Italy.
Funding
This study was supported by AIRC—Associazione Italiana Ricerca sul Cancro (IG2014-15839) and (IG2018-22059), Ministero della Salute (RF-2016-02362532) and (RC-2023-2778976) Associazione Italiana Leucemia, Linfomi e Mieloma—AIL ODV, Bologna.
Author information
Authors and Affiliations
Contributions
EB and CT conceived and designed the study; EZ, PT, LP, KM, SR, IR, MM, IV, SA, BT, AFD, and IP provided study material or patients; EB provided genomic analysis; GM, AP, and VS analyzed bioinformatic and statistical data; EB, CT, and MC discussed and interpreted data; EB and CT wrote the manuscript, and all authors reviewed and approved the paper.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest.
Ethical approval and consent to participate
All patients included in the study provided written informed consents for biological studies and have been treated following local recommendations for clinical trials or routine clinical practice at the Seràgnoli Institute, IRCCS Azienda Ospedaliero-Universitaria of Bologna, Italy.
Consent for publication
All authors have approved the manuscript for submission.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Borsi, E., Mazzocchetti, G., Dico, A.F. et al. High levels of CRBN isoform lacking IMiDs binding domain predicts for a worse response to IMiDs-based upfront therapy in newly diagnosed myeloma patients. Clin Exp Med 23, 5227–5239 (2023). https://doi.org/10.1007/s10238-023-01205-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10238-023-01205-y