
Abstract
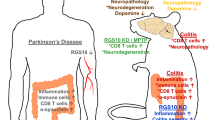
The link between the gut and the brain in Parkinson’s disease (PD) pathogenesis is currently a subject of intense research. Indeed, gastrointestinal dysfunction is known as an early symptom in PD and inflammatory bowel disease (IBD) has recently been recognised as a risk factor for PD. The leucine-rich repeat kinase 2 (LRRK2) is a PD- and IBD-related protein with highest expression in immune cells. In this study, we provide evidence for a central role of LRRK2 in gut inflammation and PD. The presence of the gain-of-function G2019S mutation significantly increases the disease phenotype and inflammatory response in a mouse model of experimental colitis based on chronic dextran sulphate sodium (DSS) administration. Bone marrow transplantation of wild-type cells into G2019S knock-in mice fully rescued this exacerbated response, proving the key role of mutant LRRK2 in immune cells in this experimental colitis model. Furthermore, partial pharmacological inhibition of LRRK2 kinase activity also reduced the colitis phenotype and inflammation. Moreover, chronic experimental colitis also induced neuroinflammation and infiltration of peripheral immune cells into the brain of G2019S knock-in mice. Finally, combination of experimental colitis with overexpression of α-synuclein in the substantia nigra aggravated motor deficits and dopaminergic neurodegeneration in G2019S knock-in mice. Taken together, our results link LRRK2 with the immune response in colitis and provide evidence that gut inflammation can impact brain homeostasis and contribute to neurodegeneration in PD.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Parkinson’s disease (PD) is the second most common neurodegenerative disorder, affecting over 10 million people worldwide. The hallmarks of the disease are the presence of cytoplasmic protein inclusions containing mainly misfolded α-synuclein (termed Lewy Bodies and neurites), the degeneration of the dopaminergic neurons in the substantia nigra pars compacta (SNpc) and neuroinflammation. Dopaminergic dysfunction ultimately leads to the characteristic motor symptoms of PD (tremor, bradykinesia, rigidity and postural instability), but other non-motor symptoms are commonly present in PD patients. More specifically, the importance of the gut-to-brain axis in PD pathogenesis already postulated by Braak has received increasing attention recently. Indeed, constipation is present in approximately 60% of the PD patients and, as it precedes motor symptoms by decades, the research criteria of the Movement Disorder Society describe it as a predictive symptom for prodromal PD [31, 64]. In addition, inflammatory bowel disease (IBD) is a chronic condition that includes Crohn’s disease (CD) and ulcerative colitis (UC) and has been identified as a risk factor for PD [48, 60,61,62, 66]. Furthermore, anti-TNF-α treatment of IBD patients eliminated this increased risk for PD, supporting the relevance of gut inflammation in PD pathogenesis [50].
The leucine-rich repeat kinase 2 (LRRK2) was genetically linked to PD in 2004 with the most common mutation G2019S accounting for 4% of familial and 1% of PD sporadic cases [27]. Mutations in LRRK2 increase the kinase activity of the protein and therefore the clinical use of LRRK2 kinase inhibitors is currently being evaluated. Despite intense research in the field, the function of LRRK2 and its involvement in the pathogenesis of PD is not fully understood. A great body of evidence supports the role of LRRK2 in immune cells and inflammatory diseases (reviewed in Cabezudo et al. 2020)). Indeed, the highest expression of this protein is found in monocytes, neutrophils, dendritic cells and to a lower extent, T- and B cells [13, 14, 20, 24, 58]. Additionally, G2019S patients and asymptomatic carriers were reported to have higher levels of inflammatory cytokines [17].
Despite the high prevalence of LRRK2 mutations in familial and sporadic PD patients, the penetrance is relatively low (~ 25–40% for the G2019S mutation) [21, 36, 41]. A combination with other environmental triggers, such as inflammation, might be needed for the mutant LRRK2 to elicit its pathogenic effect. IBD, as a chronic inflammatory condition linked to PD, can potentially be such a trigger. In this context, genome wide association studies (GWAS) have associated the LRRK2 locus and IBD, and some functional variants of LRRK2 increase the risk for both CD and PD, such as the G2019S and N2081D [28, 34, 39]. However, the association between N2081D and PD is still under debate, as it is in linkage disequilibrium with the PD-linked variant rs76904798 [8, 33]. LRRK2 has been detected in immune cells from the gastrointestinal tract, and there is evidence that this expression is upregulated in antigen-presenting cells from CD patients [20]. The involvement of LRRK2 in gastrointestinal inflammation has also been suggested by some studies in rodents. An experimental colitis model in mice commonly used to study IBD and colonic inflammation is based on administration of dextran sulphate sodium (DSS). DSS was shown to increase IFN-γ and LRRK2 phosphorylation in mice, suggesting an interplay between both proteins in the gut under inflammatory conditions [53]. In addition, both LRRK2 knock-out and BAC transgenic overexpression mice demonstrated enhanced vulnerability to DSS-induced colitis [40, 56]. And recently, chronic DSS administration in a G2019S LRRK2 overexpressing transgenic mouse, induced higher gut pathology and aggravated the ongoing dopaminergic degeneration [37]. In conclusion, further research in more relevant preclinical models is needed to better understand the link between LRRK2 and colitis (Reviewed in detail in Tsafaras and Baekelandt, 2022).
In this study, we aimed to characterise in detail the role of LRRK2 in gut inflammation by modelling IBD in G2019S mutant knock-in mice. This mouse model is currently accepted to better represent the human condition than previously developed transgenic models that rely on overexpression and exogenous promoters, since the LRRK2 levels and pattern of expression remain unaltered. We found that the presence of the LRRK2 mutation significantly enhanced inflammation not only in the colon but also in the brain. We found that this response was regulated by LRRK2 specifically in the immune cells. Additionally, colonic inflammation was reduced by pharmacological inhibition of LRRK2 kinase activity. Finally, we demonstrate that the higher inflammation in the presence of the G2019S mutation impacts neuronal survival in an α-synuclein-based PD model.
Materials and methods
Animals and ethical approval
All animal experiments were carried out in accordance with the European Communities Council Directive of November 24, 1986 (86/609/EEC) and approved by the Bioethical Committee of the KU Leuven (Belgium) (ECD projects 051/2020 and 156/2022). For this study 8–10-week-old male C57BL/6 J (NTG, JAX 000664, RRID: IMSR_JAX:000664) and B6.Cg-Lrrk2tm1.1Hlme/J (GS LRRK2, JAX:030961, RRID:IMSR_JAX:030961) were used. Animals were housed in individually ventilated cages with free access to food and water, under a controlled light–dark cycle (12-h light–12-h dark) and temperature (21 ± 1 °C).
Chronic DSS Scheme
DSS (molecular weight, 40 kDa, TdB Labs) was added to the drinking water at a concentration range of 0.5–2% (w/v) for 28 days. The DSS concentration was gradually increased by 0.5% w/v every 5 days and replaced with regular animal facility water for 2 days in between).
Bone marrow transplantation
C57BL/6-Ly5.1 (B6.SJL-PtprcaPepcb/BoyCrCrl, Charles river) and B6.Cg-Lrrk2tm1.1Hlme/J were used as donors. Donor mice were euthanised by cervical dislocation without anaesthesia and the humerus, femur and tibia were used for the bone marrow (BM) isolation. Upon aseptic conditions, BM cells were isolated, filtered through a 100 μm cell strainer, washed in ice-cold PBS, and resuspended in serum free medium (RPMI, Fisher scientific). Ten-week-old recipient C57BL/6 J and B6.Cg-Lrrk2tm1.1Hlme/J mice, lethally irradiated at 9.5 Gy for 10 min, were transplanted with approximately 10 × 106 cells/mouse by retro-orbital injection. Recipient mice were treated with 1 mM sulfamethoxazole and 0.18 mM trimethoprim in the drinking water for 2 weeks. Twelve weeks post transplantation the DSS administration protocol was initiated.
MLi-2 treatment
Mice were fed with food pellets containing either MLi-2 (MedChemExpress) at a dose of 25 mg/kg of body weight per day (Research Diets, Inc.) or placebo starting 2 days prior to DSS administration and maintained for the duration of the experiment. Animals were randomly assigned to the compound or placebo groups.
Stereotactic injections
All surgical procedures were performed using aseptic techniques and mice were treated with ketamine (60 mg/kg, i.p., Ketalar, Pfizer, Puurs) and medetomidine (0.4 mg/kg, Dormitor, Pfizer) anaesthesia. Following anaesthesia, the rodents were placed in a stereotactic head frame (Stoelting, Wood Dale, IL). Injections were performed with a 30-gauge needle and a 10-μl Hamilton syringe (Hamilton, Bonaduz, GR, Switzerland). LRRK2 GS or WT mice were injected with 2 μl rAAV2/7 αSYN vector (CMVenh-synapsin-intron-Hs αSyn WT) at a dose of 3 × 109 genome copies/ml. The right SNpc was targeted using the following stereotactic coordinates: anteroposterior − 3.1 mm, mediolateral − 1.2 mm, and dorsoventral 4.3 mm (counting from the skull) using bregma as reference. The injection rate was 0.25 μl/min, with a 5-min interval following initial needle placement. The needle was left in place for an additional 5 min period after the injection procedure before being slowly retracted.
Behavioural analysis
The motor phenotype of the rAAV2/7 αSYN vector-injected mice was assessed one month post-injection. One week prior to behavioural analysis, mice were handled on a daily basis to minimise stress and anxiety. Before each behavioural test, mice were allowed to acclimate to the testing room for a minimum of 30 min. Two behavioural tests were implemented to evaluate locomotion and forelimb asymmetry. For the rotarod, mice were placed on 5 rotating rods and were left walking face forward under steady rotation speed (4 rpm), for at least one minute (revolutions per minute). Past training time, the speed of the rotation was progressively increased from 4 to 40 rpm over 5 min. Latency to fall or passive rotation was recorded. Three trials were conducted with 1 h resting intervals in between. The mean latency for all three trials was used for analysis. For the cylinder test, mice were placed in a transparent glass cylinder and the front paw contacts to the cylinder wall were recorded as left, right, or both paws, with a total of 25 contacts per animal. The number of contralateral forelimb contacts was expressed as a percentage of total forelimb contacts.
Flow cytometry
The animals were killed with an overdose of sodium pentobarbital (Dolethal, 200 mg/kg, i.p.), followed by intracardial perfusion with ice-cold saline. Half brains were dissected and enzymatically digested with Collagenase D (Merck Life Science BV) and DNAse I (Roche) at 37 °C, followed by passage through a 70 μm cell strainer. Cells were washed with Hank’s Balanced Salt Solution (HBSS) and centrifuged at 1000 g for 10 min. The pellet was resuspended with 38% Percoll (GE Healthcare Bio-Sciences) and centrifuged for 25 min at 800 g with slow acceleration and no brake. Cells were collected from the bottom of the tube and washed once with Dulbecco’s Modified Eagle Medium (DMEM) containing 10% foetal bovine serum (FBS) and once in FBS stain buffer (BD Biosciences).
Equal parts of distant colon samples were dissected and cleaned. After incubation in HBSS with 10% FBS and 5 mM EDTA at 37 °C, the colon pieces were washed with DMEM and minced finely. Then, each sample was enzymatically digested with Collagenase D and DNAse I at 37 °C followed by passage through a 100 μm cell strainer. Cells were washed in DMEM with 10% FBS and centrifuged at 1000 g for 10 min. The pellet was resuspended in a 40% Percoll in DMEM underlaid by a layer of 80% Percoll in DMEM and centrifuged for 25 min at 800 g with slow acceleration and no brake. The interphase was collected and washed in FBS stain buffer.
For both protocols, the cells were then blocked with mouse anti-rat FcγII receptor (CD32) (D34-485, BD Biosciences) in FBS stain buffer and stained for the following surface markers: CD45 30-F11, PE-Cy7 (Biolegend); CD11b M1/70, PerCPCy5.5- (Biolegend); Ly-6G 1A8, PE (BioLegend); CD4 GK1.5, AlexaFluor 488 (BioLegend); CD8a 53–6.7, Pacific Blue (BioLegend); CD25 PC61, PE-Cy5 (BioLegend); MHC II M5/114.15.2, APC (BioLegend); Ly-6C HK1.4, Brilliant Violet 605 (BioLegend); CD19 6D5, Brilliant Violet 570 (BioLegend); CD11c, eFluor™ 506 (ThermoFisher eBioscience™); CD161 PK136, Brilliant Violet 650 (BioLegend); CD68 FA-11, Brilliant Violet 711 (BioLegend); CD3 17A2, Brilliant Violet 750 (BioLegend); CD45.2 104, APC (BioLegend); CD45.1 A20, FITC (BioLegend). A fixable viability dye eFluor 780 (eBioscience) was used in each condition. Cells were fixed in 4% PFA, and data were acquired on the following day. Flow cytometry/FACS was performed at the VIB-KU Leuven FACS Core Facility using a BD FACSymphony A5 cytometer with the FACSDiva software. Data were analysed using FlowJo v10.0.7 software. Fifty thousand to one hundred thousand events were collected for each brain sample and one hundred fifty thousand events for the colon samples.
Immunohistochemistry
Mice were killed with an overdose of sodium pentobarbital (200 mg/kg, i.p., Dolethal, Vetoquinol) followed by intracardial perfusion with saline and/or 4% PFA in PBS. After perfusion and fixation, colons were embedded in paraffin blocks, cut into 6 μm sections and mounted on glass slides. Slides were stained using standard immunohistochemistry as described below. Slides were de-paraffinised with 2 sequential 5-min washes in xylenes, followed by 5-min washes in a descending series of ethanol concentrations: 100%, 100%, 95%, 70% and finally incubated in deionised water for 5 min prior to antigen retrieval. Slides were stained for haematoxylin and eosin (H & E) staining. The histological score for the colitis was assessed based on different parameters as follows; crypts architecture: from normal morphology to loss of entire crypts morphology (0–3); presence of oedema: oedema (0–3); goblet cell depletion (0–1). The infiltration of immune cells into the lamina submucosa was scored from 0 to 3. The total histological score was calculated as the sum of the individual scores of these parameters.
For immunohistochemical analysis slides were treated with 3% hydrogen peroxide for 10 min and incubated overnight with rat anti-mouse MHC Class II (I-A/I-E) (eBioscience™) primary antibody in 10% rabbit serum (DakoCytomation) and as secondary antibody we used biotinylated anti-rabbit IgG (DakoCytomation), followed by incubation with streptavidin–horseradish peroxidase complex (DakoCytomation). MHC II immunoreactivity was visualised using 3' diaminobenzidine tetrahydrochloride (DAB; Merck Life Science BV). For immunofluorescent staining the slides were incubated overnight in PBS-0.1% triton X-100 with 10% donkey serum (Jackson ImmunoResearch Laboratories) and the following antibodies: goat Myeloperoxidase (MPO, eBioscience™) and rat F4/80 (Biolegend). After three rinses in PBS-0.1% triton X-100, the sections were incubated in the dark for 2 h in fluorochrome-conjugated secondary antibodies: donkey anti-goat Alexa 488 (Abcam) (MPO) and donkey anti-rat Alexa 555 (Abcam) (F4/80).
For the immunohistochemical stainings of the brain, after the post-fixation overnight in 4% PFA, 25 µm thick coronal brain sections were made with a vibrating microtome (HM 650 V, Microm). Immunohistochemistry was performed on free-floating sections using antibodies against tyrosine hydroxylase (TH, rabbit polyclonal, Merck Millipore). Sections were pretreated with 3% hydrogen peroxide for 10 min and incubated overnight with primary antibody in 10% rabbit serum (DakoCytomation) and as secondary antibody we used biotinylated anti-rabbit IgG (DakoCytomation), followed by incubation with streptavidin–horseradish peroxidase complex (DakoCytomation). TH immunoreactivity was visualised using Vector SG (SK-4700, Vector Laboratories).
Stereological quantification of dopaminergic neurons
For the assessment of the integrity of the SNpc, the number of TH + cells was determined by stereological measurements using the optical fractionator method in a computerised system as described before [3] (Stereo Investigator; MicroBrightField, MBF). Every 8th section throughout the entire SNpc and a total of 5 sections for each animal, was analysed for TH immunoreactivity. The coefficients of error, calculated according to the procedure of Schmitz and Hof as estimates of precision, varied between 0.05 and 0.10. Percentages represent the loss of dopaminergic neurons compared to the contralateral, non-injected side. All analyses were performed by an investigator blinded to the different groups.
Protein extraction
Colon pieces were freshly isolated, weighted and snap-frozen. Samples were homogenised in 20 volumes, respectively, of RIPA buffer (50 mM Tris–HCl, 150 mM NaCl, 0.1% (w/v) SDS, 1% (v/v) Triton-X100, 0.5% (w/v) Sodium Deoxycholate, 1.0 mM EDTA pH 7.4) containing a protease inhibitor cocktail (Roche cOmplete) and phospho-STOP EASYPACK (Roche) using a tissue homogeniser (TH, Omni Tissue Homogenizer). After 30-min incubation at 4 °C, the samples were spun for 10 min at 3000 g at 4 °C, the supernatant and was further spun for 30 min at 20.000 g at 4 °C, and the resulting supernatants represented the protein fraction. A second colon piece was homogenised in 10 volumes of urea buffer (8 M urea, 100 mM Tris–HCl, 10% glycerol, 1% SDS, 5 mM DTT, 1 mM EDTA, 1 mM EGTA, pH 6.8) containing protease and phosphatase inhibitor using a tissue homogeniser. After 30-min incubation at 4 °C, the samples were spun for 10 min at 6000 g at 4 °C, and the resulting supernatants represented the protein fraction. Protein sample concentration was determined by BCA protein assay (Thermo Scientific, MA, USA) according to the manufacturer’s directions.
Western blot
To assess LRRK2 and LRRK2 P-S1292 by Western blot analysis, 40 μg protein of colon samples extracted in urea buffer were loaded on NuPAGE 3–8% Tris–acetate gradient gels (Invitrogen, Waltham). Separated proteins were transferred to a polyvinylidene fluoride (PVDF) membrane (Bio-Rad). Nonspecific binding sites were blocked for 15 min in PBS with 0.1% Triton X-100 (PBS-T) and 5% nonfat milk. After overnight incubation at 4 °C with primary antibodies (anti-LRRK2 N241A/34, NeuroMab; anti-LRRK2 (P-S1292), Abcam; anti-vinculin V9131, Sigma), blots were washed 3 times with PBS-T and incubated with horseradish peroxidase-conjugated secondary antibody (Dako) for 1 h and washed again 3 times. Bands were visualised using Clarity Western ECL (Bio-Rad) and developed with a GE ImageQuant 800 (GE Healthcare). Densitometric analysis was performed using ImageQuant software (GE Healthcare).
Meso scale V-plex assay
Pro- and anti-inflammatory proteins were analysed in the protein fraction of the colon homogenates using a 96-well V-PLEX Mouse cytokine 19-plex (Proinflammatory Panel-1 and Cytokine Panel-1) (#K15255D; Meso Scale). Each mouse colon extract (homogenised in RIPA buffer) was diluted two fold with diluents and incubated for 2 h at RT. After washing with 0.05% Tween‐20 in 0.01 M PBS, the plates were incubated with detection antibodies for 2 h at RT. Data were generated from the V-PLEX Mouse cytokine 19-plex kits by washing away detection antibody solution and adding reading buffer to generate an electrochemiluminescence signal measured in a Meso Quickplex SQ120 (Meso Scale) and quantified with the Discovery Workbench 4.0 software.
Microscopy and image analysis
H&E slides were scanned using an Aperio ScanScope CS whole slide scanner at 40X magnification. Fluorescent staining images were obtained with Leica DM6 B upright microscope using Leica Application Suite 3.6 Download. For the counting of F4/80 and MPO positive cells, tiled images of three distant colon sections/animal were counted using QuPath 0.3.2 software. For the imaging analysis, Fiji v2.0.0 software was utilised.
Statistical analysis
Graph creation and statistical analysis were performed using GraphPad Prism for Windows (GraphPad Software, Inc.), version 9.0. Results are presented as means ± standard error of the mean. Normality of data was tested using the Shapiro–Wilk test. Data not showing normal distribution were transformed before applying the parametric ANOVA. Statistical significance was assessed using t-test or when multiple groups were analysed simultaneously, one-way ANOVA followed by Tukey’s multiple comparison post hoc test, two-way ANOVA followed by Sidak’s post hoc test, or three-way ANOVA followed by Sidak’s post hoc test. Significance was represented as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001.
Results
The G2019S LRRK2 mutation increases the susceptibility to DSS-induced colitis in mice
Administration of DSS in mice induces disruption of the distal colonic intestinal epithelial layer, resulting in histological damage with morphological and symptomatic resemblance to human UC. The severity and acuteness of the inflammation can be modulated by the dose and administration pattern of DSS. In this study, we first established a chronic and progressive colitis model by administration of increasing concentrations of DSS during a period of 28 days and compared the response between knock-in mice carrying the G2019S LRRK2 mutation (GS LRRK2) and control wild-type mice (WT LRRK2) (Fig. 1a). In agreement with the literature, our preliminary analysis suggested that female mice are partially protected against DSS [2, 22]. Since analysing both sexes separately was deemed necessary for a thorough investigation, we decided to exclusively utilise male mice in all subsequent experiments to ensure the appropriate statistical power.

The G2019S LRRK2 mutation increases the sensitivity to experimental colitis in mice. a DSS administration protocol. Mice received DSS in drinking water at increasing concentrations each time for 5 days with a pause of 2 days during one month. b Disease index score evaluation of stool consistency and haematochezia measured three times per week revealed a more severe phenotype in GS LRRK2 mice. Graph represents mean ± SD. Statistical differences were assessed using three-way ANOVA and Tukey’s post hoc test, n = 10 animals/group. ***p < 0.001. c Representative image of Haematoxylin and Eosin (H&E) staining of the distal colon showing the damage to the epithelium and inflammation after DSS. Scale bar = 100 µm. d Histological scoring of the H&E images based on crypt architecture, infiltration of immune cells, presence of oedema and depletion of the goblet cells. e–g Immunoblot analysis of total and S-1292 phosphorylated LRRK2 in distal colon protein extracts of DSS-treated and control GS and WT mice. Graphs represent mean ± SD. Statistical differences were assessed using two-way ANOVA and Tukey’s post hoc test, n = 10 animals/group. *p < 0.05, **p < 0.01, ***p < 0.001
Our DSS protocol moderately reduced the body weight but did not cause lethality, with no differences between GS LRRK2 and WT LRRK2 mice (Sup Fig. 1a). The severity of the colitis was scored three times per week by assessing the stool consistency and presence of blood in the stool (Sup Fig. 1b, c). Both parameters, as well as the combined score (disease index score), show that our chronic administration scheme induced mild experimental colitis in WT mice (Fig. 1b). Nevertheless, a significantly increased sensitivity was observed in the GS LRRK2 mice, which developed severe colitis symptoms. Additionally, the GS LRRK2 mice presented severe shrinkage of the colon, which is linked to the severity of the inflammation (Sup Fig. 1d). The histological score based on crypt architecture, infiltration of immune cells, presence of oedema and goblet cell depletion also demonstrated more damage to the colonic epithelium after DSS in the GS LRRK2 mice compared to the controls (Fig. 1c, d).
The G2019S LRRK2 mutation has been reported to increase kinase activity by approximately 4 times [65]. No difference in the basal LRRK2 protein levels was found in the colon of untreated GS and WT LRRK2 mice (Fig. 1e, f). In response to DSS, however, a significant increase in LRRK2 was found in the colon of the GS LRRK2 mice. In order to assess the kinase activity of LRRK2, we measured the autophosphorylation on Serine 1292 (Fig. 1g). Phosphorylation on S-1292 was significantly increased in GS LRRK2 mice, but to the same extent in control and DSS conditions, indicating that the kinase activity of LRRK2 remains unaltered during colitis.
To characterise the inflammation in the colon we first used flow cytometry immunophenoty** (Fig. 2a). Different populations of immune cells were quantified according to the gating strategy shown in the Supplementary Fig. 2. An overall increase in leukocytes (CD45+ cells) was found in the GS animals after DSS treatment. When looking at different subpopulations, we observed a significantly higher response in the GS LRRK2 animals treated with DSS for the non-classical (Ly6C−) monocytes, dendritic cells (CD11c+) and the lymphoid population (CD45hi, CD11b−), more specifically the CD8+ and CD4+ T cells. A similar trend that did not reach statistical significance was found for the myeloid cells (CD45hi, CD11b+), the classical (Ly6C+) monocytes and the granulocytes (defined as Ly6G+ , Ly6C+ , CD11b+ cells).

Increased inflammatory response in the colon of GS LRRK2 mice after DSS. a Flow cytometry immunophenoty** of the colon of WT and GS LRRK2 mice treated with DSS. Number of total leukocytes (CD45+), myeloid cells (CD45+ CD11b+), Ly6C+ and Ly6C− monocytes, dendritic cells (CD11c+), neutrophils (Ly6G+), lymphocytes (CD45+ CD11b−) and T cells (CD8+ and CD4+) are shown. Graphs represent mean ± SD. Statistical differences were assessed using Student’s t-test between WT and GS DSS groups, n = 5 (DSS), 2 (CTR) animals/group. *p < 0.05, **p < 0.01. b Immunostaining of paraffin-embedded distal colon sections. Representative images of macrophage (F4/80), neutrophil (MPO) and MHC II immunostaining in the distal colon of WT and GS LRRK2 mice. Quantification of macrophages c, neutrophils d, and MHC II expressing cells. e Graphs represent mean ± SD. Statistical differences were assessed using two-way ANOVA and Tukey’s post hoc test, n = 10 animals/group. *p < 0.05, ***p < 0.001
Next, we performed immunostaining in the distal colon for macrophages (F4/80), neutrophils (MPO) and for the major histocompatibility complex class II (MHC II) (Fig. 2b). In agreement with the higher level of inflammation detected by flow cytometry, the three markers were significantly increased in the GS LRRK2 mice treated with DSS (Fig. 2c, d, e). Finally, a panel of cytokines was measured in colon homogenates from these animals (Fig. 3). An overall increase in cytokine levels was present in GS LRRK2 mice after experimental colitis. More specifically, significant differences were found between WT and GS LRRK2 mice for IL-33, CXCL10, CCL2, CCL3, CXCL2, IL-10, IL-12p70, IL-2 and TNF-α, as well as a consistent trend for IFN-γ, IL-1β, IL-5, IL-6 and CXCL1.

DSS induces higher levels of pro-inflammatory cytokines in the inflamed colon of GS LRKK2 mice. Cytokines were measured in colon protein extracts from WT and GS LRRK2 mice after DSS administration as well as WT and LRRK2 control mice (CTR). Increased levels of IL-33, IP-10, MCP-1, MIP-1α, MIP-2, IL-10, IL12p27, IL-2 and TNF-α, as well as trends towards increased levels of IFN-γ, IL-1β, IL-5, IL-6 and CXCL1 were found in GS mice. Graphs represent mean ± SD. Statistical differences were assessed using Student’s t-test between WT and GS DSS groups, n = 8 (DSS), 2 (CTR) animals/group. *p < 0.05, **p < 0.01
Bone marrow transplantation of wild type into G2019S LRRK2 mice reverts the susceptibility to DSS-induced colitis
We hypothesised that the presence of mutant LRRK2 specifically in the immune cells might be responsible for the exacerbated inflammatory response of the GS mice towards DSS. Therefore, we transplanted the bone marrow of WT mice into lethally irradiated WT (WTWT) and GS LRRK2 mice (GSWT), as well as GS bone marrow into recipient GS LRRK2 (GSGS) mice. To control for proper engraftment, we transplanted in parallel bone marrow from C57BL/6-Ly5.1 mice carrying the CD45.1 alloantigen of CD45 to our GS and WT LRRK2 mice which express CD45.2. 10 weeks post transplantation, engraftment was evaluated through flow cytometry (Fig. Sup. 3a). An average of 98.7% of engraftment was measured in every control animal. Next, we induced experimental colitis in the WTWT, GSGS and GSWT mice 10 weeks after bone marrow transplantation. The disease index score of the GSWT group was improved compared to the GSGS group and reaching similar levels as the WTWT mice (Fig. 4a). The colon length was normalised in the GSWT mice treated with DSS (Fig. Sup. 3b). Also, the histological score confirmed a reduction in the colonic epithelial damage in GSWT mice (Fig. 4b; Fig. Sup. 3c).

Bone marrow transplantation of WT into GS LRRK2 mice reverts the susceptibility to DSS-induced colitis. a Disease index score of GS mice is normalised to WT levels after bone marrow transplantation from WT mice. Graph represents mean ± SD. Statistical differences were assessed using two-way ANOVA and Tukey’s post hoc test, n = 8–9 animals/group. ***p < 0.001. b Histological score of GS mice is reduced to WT levels after bone marrow transplantation. Graph represents mean ± SD. Statistical differences were assessed using one-way ANOVA and Tukey’s post hoc test, n = 8–9 animals/group. *p < 0.05. c Colonic immunophenoty** by flow cytometry of the GSWT as well as the WTWT and GSGS mice treated with DSS. Quantification of total myeloid cells (CD45+ CD11b+), Ly6C+ and Ly6C− monocytes, neutrophils (Ly6G+), dendritic cells (CD11c+), MHC II+ antigen-presenting cells, natural killer cells (CD161+), lymphocytes (CD45+ CD11b−), T cells (CD3+), CD4 T cells, CD25+ CD4 T cells, CD8 T cells and B cells (CD19+). Graphs represent mean ± SD. Statistical differences were assessed using one-way ANOVA and Tukey’s post hoc test, n = 8–9 animals/group. *p < 0.05, **p < 0.01
Flow cytometry analysis of the colon revealed overall comparable immune cell numbers in the WTWT and GSWT animals in contrast to the increase seen in the GSGS mice (Fig. 4c). More specifically, a rescue was found by the WT bone marrow transplantation in the GSWT mice for total myeloid cells (CD45+ CD11b+), Ly6C+ and Ly6C− monocytes, granulocytes (Ly6G+), dendritic cells (CD11c+), MHC II + leukocytes, NK cells (CD161+), CD4+ T cells, CD25+ expressing CD4 T cells and B cells (CD19+). These results demonstrate that the increased inflammation present in the GS LRRK2 mice after DSS is completely rescued by the bone marrow transplantation, suggesting that the DSS response is modulated by the activity of LRRK2 in the immune cells.
Inhibition of LRRK2 kinase activity partially protects G2019S LRRK2 mice from DSS-colitis
To determine whether or not the increased LRRK2 kinase activity is responsible for the enhanced DSS response in the GS mice, we treated the animals with the LRRK2 inhibitor MLi-2 [19]. MLi-2 was administered using an in-diet treatment protocol as previously described for 30 days starting 2 days before the DSS treatment [49]. The dose of ingested compound was estimated at 25 mg/kg/day (mean per animal). The treatment was well tolerated and no differences in animal weight between the treatment and placebo groups were observed (Fig. Sup 4a). LRRK2 inhibition was confirmed by quantifying LRRK2 dephosphorylation at S935 in colon through western blot (Fig. Sup 4b). Our protocol results in a reduction of 80% in the phosphorylation of S935, which suggests partial inhibition of LRRK2.
Pharmacological inhibition of LRRK2 significantly ameliorated the colitis phenotype based on the disease index score, especially in the last two weeks of the treatment (Fig. 5a). Nevertheless, protection with the treatment was partial as the MLi-2-treated animals still responded more severely than the WT LRRK2 mice. The length of the colon was not significantly altered by the inhibition of LRRK2, suggesting that LRRK2 kinase inhibition at this dose and administration route could not completely counteract the GS mutation effect (Fig. Sup. 4c).

The LRRK2 kinase inhibitor MLi-2 partially rescues the enhanced susceptibility of GS LRRK2 mice to DSS. a Disease index score evaluation in WT and GS mice treated with placebo or MLi-2, as well as controls (CTR). Graph represents mean ± SD. Statistical differences were assessed using three-way ANOVA and Tukey’s post hoc test, n = 9–15 animals/group. ***p < 0.001. b Colonic immunophenoty** by flow cytometry of GS mice treated with MLi-2 or placebo, as well as WT mice, after DSS administration. Quantification of total number of myeloid cells (CD45+ CD11b+), Ly6C+ and Ly6C− monocytes, neutrophils (Ly6G+), dendritic cells (CD11c+), MHC II+ antigen-presenting cells, natural killer cells (CD161+), lymphocytes (CD45+ CD11b−), T cells (CD3+), CD4 T cells, CD25+ CD4 T cells, CD8 T cells and B cells (CD19+). Graphs represent mean ± SD. Statistical differences were assessed using one-way ANOVA and Tukey’s post hoc test, n = 9–15 animals/group. *p < 0.05, **p < 0.01
Quantification of the immune cells in the colon by flow cytometry revealed a reduction in the MLi-2 treated mice for the different immune populations tested, with the exception of Ly6C− monocytes, which did not reach statistical significance (Fig. 5b). Overall, the results suggest a reduction of inflammation in the MLi-2-treated GS mice.
Colitis aggravates dopaminergic neurodegeneration in G2019S LRRK2 mice
Inflammatory bowel disease has been reported to increase the risk for develo** PD by 20–90% [48, 54, 60]. Furthermore, anti-inflammatory treatment of IBD patients with anti-TNF-α significantly reduces this risk [50]. Despite being protected by the blood–brain barrier, communication between the brain and the peripheral organs has been described. Inflammation in the gastrointestinal tract can have an effect on the central nervous system (CNS) homeostasis and induce neuroinflammation [Full size image
Motor behaviour was examined using different tests. Rotarod test was used to evaluate the muscular coordination and grip strength of the animals 5 weeks after rAAV injection (Fig. 6c). Remarkably, a significant decrease in motor performance was found in the GS injected animals compared to the WT controls in the absence of DSS. DSS increased the motor phenotype in the WT mice, but no additional effect was observed in the GS mice. In parallel, the cylinder test was used to measure the forelimb asymmetry at 3 and 5 weeks post-injection (Fig. 6d). We observed a progressive reduction in the use of the contralateral left paw which only reached significance at the last time point between the WT and GS mice injected with α-synuclein rAAV combined with DSS, suggesting a synergistic effect between the α-synuclein overexpression and the colon inflammation.
Stereological quantification of the dopaminergic neurons in the substantia nigra corroborated the behavioural data (Fig. 6e, f). A trend towards increased neurodegeneration was observed in the GS LRRK2 mice after α-synuclein overexpression. Furthermore, DSS significantly aggravated this TH+ cell loss in the GS LRRK2 mice but not in the WT control mice. These results confirm that in GS LRRK2 mice, DSS-experimental colitis can aggravate α-synuclein-mediated dopaminergic neurodegeneration.
