
Abstract
Estuaries are crucial feeding, nursery and resting sites for fish but can also be subject to the impacts of severe flooding. The environmental features of estuaries can mediate how they respond to these impacts. For example, the size, configuration, and context of estuarine habitats across seascapes affects the value of patches for fish, and so fish assemblages at sites with a greater habitat extent or closer to the mouth of an estuary may rebound more quickly from flooding. We investigated how a once in 100-year flood event affected fish assemblages at approximately 600 sites across 13 estuaries and six estuarine habitats (bare sediments, log snags, mangrove forests, rocky structures, saltmarsh and seagrass meadows) in southeast Queensland, Australia, and determined whether flood impacts were mediated by the position of sites within the broader estuarine seascape. Sites were surveyed annually in 2020/2021 (pre-flood) and 2022 (6 months post-flood) using underwater videography. Flooding modified the structure of the fish community and reduced the abundance of fish targeted by local fisheries in all six habitats. Crucially, flood effects on fish were greater at sites near more expansive urbanisation in some ecosystems, but lower at sites nearer to the estuary mouth. Maximising the extent of natural habitats across estuaries can mediate the effects of floods and should be priorities for restoration and management plans seeking to maintain biodiversity and fisheries productivity in the face of increasing climate-related disturbances.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Estuaries are highly dynamic systems that are periodically impacted by above average rainfall, often over very short periods of time (Gillanders et al. 2011; Robins et al. 2016; Harrison et al. 2022). However, on rare occasions, these systems will experience continued severe rainfall across surrounding catchments that results in significant flooding. Floods are increasing in size and intensity in many areas globally due to climate change (Wetz and Yoskowitz 2013), and the effects of flooding on estuaries can be worsened by the composition or extent of urban land and/or natural habitats (Harrison et al. 2022). Floods cause significant change across estuaries, including increased water flow, sedimentation and nutrient concentrations, and alterations to the physical boundary of the estuary (e.g. erosion along the estuary margin) and the structure of the habitats within (Cooper 2002; Rogers and Woodroffe 2016). These impacts are intensified by urbanisation, because natural habitats are often replaced with impervious surfaces which themselves accelerate the speed and velocity of water currents, and the concentration of nutrients and pollutants that runoff into estuarine waters. The physical effects of a flood in an estuary typically only last for a short period of time (e.g. days to weeks), but several ongoing and persistent negative consequences of flooding (e.g. habitat destruction, water quality degradation, increased water velocity) can impact estuaries for periods of up to several years (Wetz and Yoskowitz 2013; Reithmaier et al. 2021). However, the effects of substantially increased sediment deposition and water quality degradation in some estuarine habitats (e.g. mangrove forests, saltmarsh ecosystems and seagrass meadows) or physical habitat degradation due to fast flowing waters or erosion may have ongoing consequences for the continued structure and functioning of ecosystems (Olley et al. 2006; Wetz and Yoskowitz 2013).
Estuarine seascapes are comprised of a diversity of coastal habitats that are each crucial feeding, nursery and resting sites for adult and juvenile fish (Barbier et al. 2011; Sheaves et al. 2015; Whitfield 2017). Fish use a variety of these estuarine habitats throughout their lifecycle due to changes in resource availability and requirements associated with tidal, seasonal and ontogenetic movements (Irlandi and Crawford 1997; Krumme 2009; Becker et al. 2016), and so their distribution and abundance responds significantly to variation in the context (e.g. the extent of different habitats and the position of a site relative to the estuary mouth) of the seascape (Sheaves and Johnston 2008; Lacerda et al. 2014; Gilby et al. 2018). For example, fish species abundance and diversity in estuaries is generally greater in habitat patches that have a higher extent and diversity of natural habitats (e.g. seagrass and saltmarsh) that provide unique foraging and refuge opportunities. Similarly, the proximity of sites relative to the mouth of the estuary allows for movement into coastal waters which benefits some species, while others may be more abundant at sites further away (Henderson et al. 2019; Jones et al. 2020). Increasing and expanding anthropogenic impacts in the coastal zone does, however, have significant implications for the structure of estuarine seascapes and result in reductions in the habitat value for fish across all coastal habitats (Waltham and Connolly 2007; Sheaves et al. 2012; Goodridge Gaines et al. 2022). Increasing the extent of natural habitats across coastal seascapes can also work to mitigate or mediate the effects of anthropogenic impacts (Olds et al. 2012b). For example, locations with a greater extent of natural habitats nearby might be more resilient to the impacts from poor weather or floods, fishing or broader habitat loss (Olds et al. 2014). More thoroughly understanding these patterns helps to optimise coastal planning and management to improve the resilience of coastal ecosystems to change.
Habitat context, the extent of habitat and position in the estuary (position relative to other habitats and variation in water chemistry variables such as salinity and turbidity), is a strong predictor of variation in fish diversity and abundance across estuarine seascapes (Pittman et al. 2007; Gillson et al. 2012) and it is therefore likely a strong predictor of how fish respond to disturbances like floods (Carlson et al. 2016). However, such patterns remain poorly understood in most systems, meaning that it is unclear how we can incorporate the associated impacts of severe disturbances to management actions (Morrisey et al. 2003; Becker et al. 2013). In inshore reefs and seagrass meadows, the effects of habitat context are associated with positive outcomes for fish after the implementation of marine reserves and the resilience of ecosystems to natural disturbances (Olds et al. 2012a; Henderson et al. 2017). Greater extent of urbanisation across coastal seascapes might, however, reduce the extent of natural habitats, and exacerbate the effects of flooding on fishes in coastal habitats (Romme et al. 1998; Turner and Gardner 2015). This is because urban estuaries can contain a range of instream structures (e.g. rock walls, jetties, pontoons) that have replaced natural estuarine habitats (i.e. such as mangroves, seagrass and saltmarsh) (Bishop et al. 2017). Alternatively, estuaries that are surrounded by natural habitats may be able to buffer the impacts from floods or cyclones as natural habitats can filter the effects of flooding by trap** sediments and nutrients and therefore can provide resilience to those that are degraded or lost (Thrush et al. 2008; Fairchild et al. 2021). It is also well established that there are significant differences in fish communities and their responses to impacts between different estuarine habitats (Dolbeth et al. 2016). Further, the effects of habitat context vary between estuarine habitats, resulting in potentially inconsistent patterns in response to impacts such as flooding (Goodridge Gaines et al. 2022). It is, therefore, possible that different coastal ecosystems, and the fish communities that reside within them, have distinct responses to flood impacts. It remains unknown, however, whether the positive effects of habitat context (e.g. greater extent of natural habitats) in estuaries can mediate the effects of flooding on fish, whether they be positive or negative on all or certain aspects of the community, and whether these effects differ between estuarine habitats.
Distinct fish species in estuaries are likely to be impacted differently by both the short- and long-term impacts of severe flooding according to their habitat and physiological requirements over multiple temporal scales (Gillanders et al. 2011). Furthermore, in many cases, the effects of flooding on fish can be linked to changes in salinity, which alters the distribution and abundance of fishes throughout estuaries (Kimmerer 2002; Gillson et al. 2012). Different species will cope differently with the variations in salinity due to varying physiological tolerances (Gillson et al. 2012). Therefore, flooding is likely to impact different components of estuarine fish communities in different ways. For example, specialist (e.g. species with a narrow trophic niche) or sensitive species are more likely to be impacted by flood impacts (Villéger et al. 2010; Payne et al. 2015). Conversely, generalist species (e.g. species with a broad trophic niche) or those that are highly mobile would be expected to increase in abundance more quickly, change their feeding behaviours in response to altered resource availability or maintain their abundances after flooding due their ability to more effectively cope with disturbances (Henderson et al. 2020). Whether and how flooding impacts different components of fish assemblages remains relatively unknown, thereby severely limiting our understanding of the functional effects of flooding.
Floods and other severe weather events have significant impacts on the structure of coastal ecosystems by causing short term changes in salinity, light penetration and water movement, but also long term modifications such as habitat destruction (Cooper 2002; Gillson et al. 2012). Additionally, humans have significantly modified estuarine ecosystems around the world, which has resulted in significantly negative effects for the structure of estuaries, the habitats that frequent estuaries (e.g. mangroves, seagrass meadows, saltmarshes), and the species (e.g. fish and invertebrates) that reside within estuaries (Gibson et al. 2002). Flooding events do, however, present opportunities to determine how these impacts modify the fish communities that reside within estuaries as we currently lack the understanding of how to manage coastal habitats to ensure they remain resilient to these types of disturbances. In February 2022, southeast Queensland, Australia, experienced a severe, once in 100-year flooding event. In this study, we determine the effects that this flooding event had on the structure of fish assemblages across 13 estuaries in the region. Further, we determine whether fish communities from six estuarine habitats (mangroves, seagrass, saltmarsh, unvegetated sediments, rocky outcrops and log snags) respond differently to flooding according to their position across estuarine seascapes. We hypothesised that sites in estuaries with greater extents of urbanisation would have negative effects on fish diversity and abundance following floods. Furthermore, we expect that the effects of the flood on fish communities will be reduced at the mouth of the estuary (e.g. closer to the ocean), compared with further upstream, as improved flushing of this part of the estuary causes salinity and turbidity levels to rebound more quickly due to the ingress of tides.
Methods
Study system and flood impacts
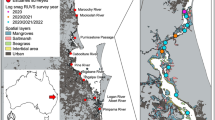
Fish surveys were undertaken in the austral winter (June to August) in each of 2020, 2021 and 2022 in 13 estuaries in southeast Queensland, Australia (Fig. 1a). The estuaries sampled contain a gradient of relatively natural systems, to those with extensive modification of the estuary shorelines with widespread urbanisation and human disturbance throughout the catchment (Goodridge Gaines et al. 2022). Within each estuary, we sampled up to six different habitats: unvegetated sediments, log snags, mangrove forests, rocky structures, saltmarsh, and seagrass (Fig. 1a–c). We sampled up to 10 sites of each habitat in each estuary (resulting in up to 60 sites per estuary), with sites positioned as evenly as possible throughout the stretch in which the habitat was present in each estuary. In some cases, we were not able to sample all habitats in each estuary as they either were not present (seagrass doesn’t appear in each estuary) or were not a large enough extent to survey ten sites. Surveyed sites were positioned from the mouth of the estuary to the maximum extent of tidal influence, which varied from 7 km in Tallebudgera Creek, to 77 km in the Brisbane River (Fig. 1b, c). This distance also coincides with the most upstream estuarine monitoring site of the local waterway monitoring program in each system (Table 1) (EHMP 2023). Sites were positioned at least 50 m apart to maximise spatial independence. This rule determined the number of sites of some ecosystems in some estuaries. For example, in a hypothetical estuary where only a single, small (< 50 m long) patch of seagrass is present, then only one site could be surveyed in this estuary.

a Location of the study estuaries in southeast Queensland, Australia. Insets illustrate the sampling of different habitats along b the longest estuary in the region, the Brisbane River, and c the shortest estuary, Tallebudgera Creek. d Rainfall during the February 2022 flooding event resulted in an average of 744 mm falling in each estuary during the month of the flood. This had a significant impact on e salinity and f turbidity. The grey section d–f indicates our period of sampling in each year
In February 2022, the southeast Queensland region experienced extensive rainfall which resulted in severe flooding occurring throughout the sampled estuaries (Australian Bureau of Meteorology 2023). Average rainfall in each sampled estuary in February 2022 was 744 mm, while the historic February average is 158 mm (Fig. 1d). Average yearly rainfall for the region is approximately 1150 mm (Australian Bureau of Meteorology 2023). Rainfall resulted in significant flooding across all surveyed catchments, including the significant inundation of residential and industrial areas across the region. The extensive rainfall in the region had significant impacts on both the salinity and turbidity of all estuaries, with salinity in the months immediately following the flood being on average lower than any other time from 2020–2022, and turbidity being higher than all months except for December 2021, a time when above average (but not flooding) rainfall was also experienced in the region (EHMP 2023) (Fig. 1e, f).
Fish surveys
We surveyed fish communities using 30 min deployments of remote underwater video system (RUVS) at each site (Colton and Swearer 2010; Gilby et al. 2018). This approach was preferred to baited cameras given our interest in the habitat-specific effects of flooding on fish and the effects of bait attracting fish from outside of the focal habitat to the field of view. RUVS consisted of a GoPro camera recording in high definition (1080p) fixed to a 3 kg weight and buoyed at the surface for retrieval. In this study, we optimised our sampling approach by focusing on appropriate times where water clarity was maximised (e.g. at high tide, avoiding times with recent rainfall and shallower environments). Therefore, all surveys were completed two hours either side of the daytime high tide and we limited sampling to times where recent rainfall had not occurred to maximise visibility (Henderson et al. 2019; Goodridge Gaines et al. 2022). When RUVS were placed in structured habitats (i.e. rocky structures, log snags, mangrove forests and seagrass meadows), the video field of view was positioned along the edge of the habitat, thereby recording fish moving in and out of the habitat and preventing the habitat itself (e.g. mangrove pneumatophores, seagrass blades) from obscuring the camera. RUVS were deployed sequentially upstream from the estuary mouth (following the ingress of the tide) and up to twelve RUVs were deployed at the same time, meaning that adjacent sites were typically surveyed within the same 30-min window, thereby further enhancing our spatial independence. Surveys were repeated at the same sites in the austral winter of 2020, 2021 and 2022. This period was selected to coincide with periods of maximum water clarity within the region. This design resulted in over 1800 sites being surveyed across all years of the study (Table S1).
Fish assemblages were quantified from videos using the standard MaxN metric; the maximum number of each species observed in a single frame in any 30 min video. MaxN therefore ensures individuals are not counted more than once at each site and is considered a conservative index of relative abundance (Ellis and DeMartini 1995; Stobart et al. 2007). From the resulting fish assemblage matrix (i.e. species by site), we also calculated the compound metrics of fish species richness (being the number of unique species identified at each site), total fish abundance (being the sum of the MaxN values for all species identified at each site) and harvested fish abundance (being the sum of MaxN values of species commercially and recreationally harvested within this region at each site).
Environmental variables
To determine whether the flood that occurred at the start of 2022 had effects on the composition of fish assemblages in estuaries, we categorised surveys completed in 2020 and 2021 as “Pre-flood” and those completed in 2022 as “Post-flood”. Previous studies in the region have found that the distribution of fish in estuaries are significantly impacted by the proximity of the site to the ocean and the extent of mangrove forests and urban land immediately around each site (Gilby et al. 2018; Henderson et al. 2019; Goodridge Gaines et al. 2022). Consequently, it was these variables that we hypothesised would have the greatest effects on flood impacts. Each of these seascape context measures were calculated in QGIS (Quantum GIS 2023), by measuring the distance from each site to the ocean and calculating the area of mangroves and urban land in a 500 m buffer. This buffer size was chosen as it represents the maximum likely movement of most estuarine fish species in a single tidal cycle and reflects the area over which most previous effects of estuarine habitat extent have been found in this region (Olds et al. 2012a; Gilby et al. 2018; Goodridge Gaines et al. 2022). All spatial layers (e.g. those used to calculate mangrove forest and urban area) were obtained from existing habitat and land use maps (Queensland Government 2019) and were ground-truthed in the field or by overlap** with aerial imagery of the region collected by NearMap (NearMap 2009).
Statistical analysis
We tested for collinearity between all of our continuous variables (e.g. mangrove area, urban area and distance to ocean) prior to analyses, with a Pearson’s correlation coefficient cut off set at 0.7 (no variables were removed due to this cut off). We used ManyGLMs in the mvabund package (Wang et al. 2022) of R (R Core Team 2023) to test how fish communities in each estuarine habitat were impacted by the effects of the flood and whether these effects were mediated by seascape context. ManyGLM is a multivariate analysis that identifies variables that best explain patterns in the fish community (i.e. a multivariate matrix of fish species by replicate site across all years, n = 1812, Table S1), as well as the fish species from that community that best correlate with the overall patterns of best-fit variables. The ManyGLM works by fitting a generalised linear model between each species in the analysis and the predictor variables and then uses resampling to test for significant community level effects based on the predictors (Wang et al. 2012). Given the likely difference in effects between habitats, we calculated separate ManyGLMs for each habitat (Goodridge Gaines et al. 2022). Additionally, to limit the effect of rare species, we only analysed ManyGLMs on the species that made up 95% of the total abundance in each habitat. Model structure included the interaction of flood effect (fixed factor, two levels; pre-flood and post-flood) with all seascape variables (i.e. mangrove area and urban area within a 500 m buffer and distance to the ocean) and the fixed effect of estuary (as we are unable to include this as a random variable in a ManyGLM). All ManyGLM models were fit on a negative binomial error distribution and all residual vs. fitted plots are provided (see Fig S1). Best-fit models for each ecosystem were identified using reverse stepwise simplification on Akaike’s information criterion (AIC), with the best-fit ManyGLM for each habitat visualised using a non-metric multidimensional scaling (nMDS) ordination plot. The “p.uni” function within "anova.manyglm” in the mvabund package was used to identify indicator species that were correlated with variables from the best-fit manyGLM model (Wang et al. 2012). The “p.uni” adjusted method returns a test statistic and p-value for each species that has been adjusted for multiple testing and is calculated using a step-down resampling algorithm, which uses the resampling to ensure inferences take into account the correlation between species (Westfall and Young 1993; Wang et al. 2012). The trajectory of the best-fit variables and indicator species were visualised over the nMDS ordination using Pearson’s vectors. When a species and significant variable vectors correlate with one another in the same direction, the species is becoming more abundant when that variables value is increasing. When these variables are perpendicular to one another, this would suggest that that species is more abundant at intermediate values of that variable.
We then used generalised additive mixed models (GAMMs) in the mgcv package (Wood and Wood 2015) to assess the effect of flood impacts and seascape context variables on species richness, fish abundance and harvested fish abundance. GAMMs were fit with a negative binomial distribution and all variables were limited to three knots to avoid model overfitting. Models were structured with three three-way interactions between flood impact, habitat and each seascape context variable, with estuary modelled as a random factor. Model selection was undertaken using the ‘dredge’ function in the MuMIn package of R, which runs all possible combinations of four variables or fewer to minimise model overfitting (Barton and Barton 2015). Best-fit models were those with the lowest AICc values.
Results
Estuarine fish assemblages
We identified 84 fish species across the 3 years across approximately 600 sites in six habitats and 13 estuaries. Six species accounted for greater than 85% of the fish sampled across the 3 years, with the most common species being estuary glassfish (Ambassis marianus, 42.7%), yellowfin bream (Acanthopagrus australis, 15.3%), southern herring (Herklotsichthys castelnaui, 8.7%), common hardyhead (Atherinomorus vaigiensis, 8.2%), common silver biddy (Gerres subfasciatus, 6%) and sea mullet (Mugil cephalus) (5.5%).
Flood effects on estuarine fish communities
Bare sediments
The structure of fish assemblages on bare sediments was best explained by urban area (χ2 = 30.46, p = 0.004) (Fig. 2a) and the non-significant effects of flooding (χ2 = 22.72, p = 0.11) and distance to ocean (χ2 = 23.42, p = 0.073). There was no significant interaction between flooding and any seascape features (Table 1, Fig. 2a). We identified two fish species that best explained the variation across sites, common silver biddy (Gerres subfasciatus) and yellowfin bream (Acanthopagrus australis) (Table 1, Fig. 2a). The abundance of yellowfin bream increased after the flood, while the abundance of common silver biddy was correlated with increased urban area (Fig. 2a).

Non-metric multi-dimensional scaling ordination (nMDS) for each habitat a unvegetated sediments, b log snag, c mangroves, d rocky, e saltmarsh and f seagrass. Pearson vector overlays show significant variables (black) and indicator species (blue) from the best fit ManyGLM. Black points were used when no significant effect of flooding was found
Log snags
The structure of fish assemblages on log snags were best explained by the flood impact (χ2 = 44.5, p = 0.008), distance to the ocean (χ2 = 114.7, p = 0.001), estuary (χ2 = 665.6, p = 0.003) and the non-significant effect of mangrove area (χ2 = 14.6, p = 0.592) (Fig. 2b). There was no significant interaction between flooding and an seascape features (Table 1, Fig. 2b). We identified eight fish species that best explained the variation across sites, estuary glassfish, yellowfin bream, common silver biddy, sea mullet, diamondfish (Monodactylus argenteus), Moses perch (Lutjanus russeli), luderick (Girella tricuspidate) and gobiidae spp. (Table 1, Fig. 2b). The abundance of all indicator species was not impacted by the effect of flooding. (Fig. 2b). The abundances of estuary glassfish, yellowfin bream, common silver biddy, luderick and gobiidae spp. were significantly modified by distance to ocean (Fig. 2b). The abundance of estuary glassfish, yellowfin bream, common silver biddy, sea mullet, diamondfish and Moses perch were modified by the effect of estuary (Fig. 2b).
Mangrove forests
The structure of fish assemblages in mangrove forests were best explained by the interaction of flood impact and mangrove area (χ2 = 20.22, p = 0.037) and estuary (χ2 = 282.74, p = 0.001) (Fig. 2c). We identified three fish species that best explained the variation across sites; estuary glassfish, yellowfin bream and common silver biddy (Table 1, Fig. 2c). The abundance of yellowfin bream decreased post-flood (Table 1, Fig. 2c).
Rocky structures
The structure of fish assemblages on rocky structures were best explained by the area of urban land (χ2 = 16.81, p = 0.254), distance to ocean (χ2 = 116.58, p = 0.001) and flood impact (χ2 = 29.1, p = 0.009) (Fig. 2d). There were no significant interactions between flooding impact and any seascape features. We found five species that best explained the variation across sites; estuary glassfish, yellowfin bream, common silver biddy, luderick and fineline surgeonfish (Table 1, Fig. 2d). There were no indicator species that significantly correlated with the impact of flooding.
Saltmarsh
The structure of fish assemblages using saltmarsh ecosystems was best explained by flood impact (χ2 = 44.5, p = 0.003), the area of mangroves (χ2 = 22.9, p = 0.005), the effect of estuary (χ2 = 225.35, p = 0.006), and the distance to ocean (χ2 = 41.15, p = 0.02) (Fig. 2e). There was no significant interaction between flooding impact and any seascape features. We found seven species that best explained the variation across sites; estuary glassfish, yellowfin bream, sea mullet, common silver biddy, Australian sardine (Sardinops sagax), Australian smelt (Retropinna semoni) and sand whiting (Sillago ciliata) (Table 1, Fig. 2e). There were no indicator species that significantly correlated with the impact of flooding.
Seagrass
The structure of fish assemblages in seagrass was best explained by the interaction between flood impact and the area of urban land around the site (χ2 = 26.7, p = 0.011), and the effect of estuary (χ2 = 107.1, p = 0.017) (Fig. 2f). We found two species that best explained the variation across sites; estuary glassfish and sea mullet (Table 1, Fig. 2f). There were no indicator species that significantly correlated with the impact of flooding.
Flood impacts on fish diversity and abundance
The abundance of fish in estuaries was not modified by the significant interaction between urban area and flooding impact. The abundance of fish in estuaries was modified significantly by the interaction of distance to ocean and flood impact (χ2 = 88.73, p < 0.001), the interaction of habitat and impact (χ2 = 152.5, p < 0.001) and the random effect of estuary (χ2 = 236.6, p < 0.001) (dev = 23.5%, Fig. 3a). There were no other models within 2 AICc values of the best fit model. While the effects of habitat and distance to ocean was significant, there was no clear impact of the effect of the flood on fish abundance, except for bare sediment habitats (Fig. 3a). Similarly, there was no consistent trends between pre- and post-flood associated with habitats and distance to ocean interaction, with pre- and post-flood samples following the same trajectory (Fig. 3a). In all habitats, except bare sediments post-flood, the total fish abundance was highest closest to the ocean (Fig. 3a).

Generalised additive mixed models (GAMMs) on best-fit models for a total fish abundance, b harvested fish abundance and c species richness. Different colours represent the pre- (yellow) and post-flood (blue). Shaded areas around the best-fit model indicate the 95% confidence interval
The abundance of harvested fish in estuaries was not modified by the significant interaction between urban area and flooding impact. The abundance of harvested fish in estuaries was significantly modified by the interaction of distance to ocean, habitat and flood impact (χ2 = 12.18, p = 0.032) and the random effect of estuary (χ2 = 332.4, p < 0.001) (dev = 28.8%, Fig. 3b). There were no other models within 2 AICc values of the best fit model. There was a significant effect of the flood on the abundance of harvested fish in log snags, mangrove forests, rocky structures and saltmarsh ecosystems, with each of these habitats having a greater number of harvested fish prior to the flood compared with after (Fig. 3b). Similarly, the effect of distance to ocean was significantly different pre- and post the flood for mangroves, rocky structures and saltmarsh ecosystems, with pre-flood samples all having a greater abundance but followed the same trajectory (Fig. 3b).
The species richness of fish in estuaries was not modified by the significant interaction between urban area and flooding impact. Species richness in estuaries was significantly modified by the main effects of habitat (χ2 = 129.39, p < 0.001), flood impact (χ2 = 90.15, p = 0.001) and the random effect of estuary (χ2 = 405.2, p < 0.001) (dev = 28.3%, Fig. 3c). There were no other models within 2 AICc values of the best fit model. All habitats had a significantly greater species richness prior to the flood (Fig. 3c). This ranged from a decrease in species richness from approximately 17% in seagrass meadows to 45% in saltmarshes (Fig. 3c). No connectivity or seascape composition variables were correlated with species richness across the sampled estuarine habitats.
Discussion
The effects of flooding and other severe natural weather events can have a significant financial and social cost for humans, leading to a decline in a variety of ecosystem services, but also contribute to the degradation or removal of natural habitats, with significant impacts for a range of animals (Preen et al. 1995; Carlson et al. 2016; Harrison et al. 2022). The effects of flooding on some species might, however, be mitigated by maintaining and managing for a greater extent of natural habitats. Natural habitats can buffer nutrients and maintain high resource availability, reducing the extent of urbanisation that exacerbate the impact of flooding along estuary verges to ensure they are resilient to the severe disturbances associated with flooding (Sutton-Grier et al. 2015; Temmerman et al. 2022), but this hypothesis has rarely been tested. Here, we found that the effects of a significant flooding event in southeast Queensland, Australia modified the composition on fish assemblages in several estuarine habitats (i.e. mangrove forests, log snags, rocky structures and saltmarsh) 6 months post flooding, and that these effects were primarily mediated by the proximity to the ocean, with additional effects for some habitats of mangrove. However, we only identified negative synergistic effects of flooding and the area of urban land on fish communities in seagrass meadows, with this effect not being found anywhere else. Overall, the habitats that experienced the most significant effects of the flooding on their inhabiting fish assemblages were mangrove forests, log snags, rocky structures, saltmarshes and seagrass habitats, with all experiencing significant changes in their community structure and reductions in harvested fish abundance post-flood. While species richness was greatest in all estuarine habitats prior to the flood, these effects were not modified by seascape positioning. While there are often short-term effects in estuaries post-flooding such as increased water flow and decreased light penetrations due to sediment, these effects are likely to be remedied relatively quickly (Wetz and Yoskowitz 2013; Reithmaier et al. 2021). Maximising the extent of natural habitats and minimising urbanisation around estuary verges, particularly towards the mouth of estuaries, can help mitigate the effects of flood impacts in estuaries on fish assemblages, and thereby maintain the populations of important species to commercial and recreational fishing.
Species richness and the abundance of all fish and harvested species responded negatively to the impact of the flood in estuarine habitats. While the effects of flooding on harvested fish abundance were mediated by variation in the seascape context, there was no such relationship for species richness. Maximising biodiversity is important in enhancing the ecological functioning of coastal seascapes, as different species use different resources that are available within, or between, habitats (Duffy et al. 2007; Henderson et al. 2020; Mosman et al. 2023). The species that reside within estuaries can be sensitive to a range of disturbances such as urbanisation and habitat fragmentation, and therefore, negative effects for biodiversity due to flooding were expected (Chapman and Blockley 2009; Bishop et al. 2017). While we document negative effects of flooding 6 months post impact here, it is unclear as to the timeframe that would be required for fish diversity and abundance to return to pre-flood levels. The short-term effects of flooding (e.g. increased turbidity and water flow) would be expected to reside for a period of days to several weeks (Wetz and Yoskowitz 2013; Reithmaier et al. 2021), and indeed this is reflected in water quality monitoring data within the region (EHMP 2023). However, we sampled fish communities 4–6 months after the flood occurred, suggesting that the impacts associated with the flood may have modified food and habitat availability in some habitats, or even removed entire populations from some estuaries (as evidenced by the overall reduction in species richness post flood) (Gillson et al. 2012; Williams et al. 2017). Many of the species that we found to be indicators of change in estuarine habitats were estuarine specialists that feed on detritus, vegetation, invertebrates or small fish, all of which are likely to be depleted by flooding due to changes in water flow and turbidity (Dittmann et al. 2015; Henderson et al. 2020). While many of these fish species could migrate back to the impacted estuary relatively quickly post-flood, it is likely that their preferred food sources have reduced and will take an extended time to recover (e.g. up to or longer than 12 months) (Gillson et al. 2012). For example, Dittmann et al. (2015) found that the abundance of some invertebrates species increased quickly after a flood event, but it could take multiple years for larger bodied invertebrates and overall biomass to return to benthic sediments. Given that extreme weather events such as floods of this magnitude are expected to become more frequent with climate change (Taherkhani et al. 2020), a more detailed understanding of these subtle ways in which flooding modifies estuaries, their biodiversity, and functioning is increasingly important.
The influence of seascape context is a well-established driver of fish community structure in estuaries and other coastal ecosystems (Meynecke et al. 2008; Nagelkerken 2009). Here, we found that the effects of flooding on total fish abundance and the abundance of harvested species were mediated by the proximity of a site to the ocean. The mouths of estuaries are frequent transition zones for many different fish species that move between estuarine and offshore environments (e.g. surf zones, headlands and offshore reefs) (Teichert et al. 2018; Jones et al. 2020), but would also be the region of the estuary that is most likely to recover most quickly. This is because the mouths of estuaries experience faster currents due to tidal exchange and can be impacted by waves, all of which is likely to make them more resilient to the effects of flooding (Whitfield et al. 2008). Furthermore, it would be expected that the habitats closer to the ocean would receive the first influx of species moving back into the estuary. Regions with a greater extent of natural habitats often support a greater abundance and diversity of fish species, even in the face of disturbances, because of their increased availability of resources and feeding opportunities (Teichert et al. 2018; Henderson et al. 2019). Managing and restoring estuarine habitats in these regions of the estuary, particularly given their importance to species that move into coastal waters, should therefore remain a key target for management seeking to optimise the resilience of landscapes to broad scale impacts. Similarly, for species that are more estuarine dependent, management should focus on maintaining or improving the extent of natural habitats throughout the entire stretch of the estuary, as structured habitats always maintained a higher abundance and diversity than unstructured habitats.
We found that the area of urban land, mangroves and the proximity to the mouth of the estuary were key drivers of changes in fish community structure and that proximity to the ocean interacted with the effect of flooding for harvested fish abundance (Gilby et al. 2018; Henderson et al. 2019). When floods occur, they are likely to have a significant influence in all estuaries, however, the duration of that impact may be modified by the extent of urban and natural land surrounding specific habitats (here mangrove forests and seagrass meadows) in an impacted estuary (Borsje et al. 2011; Temmerman et al. 2022). This is because estuaries that contain extensive urban area surrounding the waterway are typically dominated by a greater extent of impermeable surfaces that increase the transfer of water and pollutants from the land to the estuary (Waltham and Connolly 2007). Furthermore, urban estuaries are characterised by the depleted extent of natural habitats which act as a filtration system for run off (Temmerman et al. 2022). This often results in increased nutrient and sediment runoff into estuaries, which can have deleterious effects for estuarine habitats and fish (Dauer et al. 2000; Wetz and Yoskowitz 2013). Maintaining or restoring natural habitats such as mangrove forests closer to the estuary mouth can, however, help to mitigate some of these effects (Sutton-Grier et al. 2015; Temmerman et al. 2022). These habitats buffer waves and absorb nutrients when an estuary receives an influx during a flood (Adame and Lovelock 2011; Temmerman et al. 2022). Nature-based approaches are increasingly desirable attributes of coastal hazard mitigation plans (Borsje et al. 2011; Morris et al. 2018). By increasing the extent of natural habitats in estuaries, we will ideally lessen the long-term changes associated with severe floods and the fish that rely upon them.
Estuaries are highly dynamic systems; however, they also experience natural and anthropogenic disturbances that can modify the composition of fish that reside in this ecosystem. We show how the effects of a once in 100-year flood in southeast Queensland, Australia modified the fish community, abundance of key harvested species and reduced biodiversity across all estuarine habitats. It is important to note that we were not able to include unimpacted estuaries in our study as the severity of the flooding event impacted all estuaries in the region, removing the possibility of comparing our results to similar estuarine seascapes that did not experience flooding. The impacts of the flood, however, were mediated by the position of the habitat and the spatial extent of natural and anthropogenic habitats in some ecosystems. Sites with extensive mangroves are prime locations where water quality and resource availability are maximised during disturbances, and therefore, likely support flood resilience. Our results support the notion that maintaining, managing and/or restoring natural habitats in the downstream sections of estuaries is crucial in maximising the future resilience of estuaries and their animal assemblages to flood impacts. We do, however, note that not all species in estuaries are reliant on the connectivity to the ocean and that focusing restoration and management on natural habitats throughout the entirety of the system would be beneficial for many estuarine dependent species. With an increased frequency of severe floods expected to occur with sea level rise and climate change, effective storm and flood mitigation that is successful for the preservation of estuarine habitats, fish communities and broader ecosystem services is required.
Data availability
Data and code used in this study will be made available upon reasonable request to the corresponding author.
References
Adame MF, Lovelock CE (2011) Carbon and nutrient exchange of mangrove forests with the coastal ocean. Hydrobiologia 663:23–50
Australian Bureau of Meteorology (2023) Average annual, seasonal and monthly rainfall. http://www.bom.gov.au/climate/maps/rainfall
Barbier EB, Hacker SD, Kennedy C, Koch EW, Stier AC, Silliman BR (2011) The value of estuarine and coastal ecosystem services. Ecol Monogr 81:169–193
Barton K, Barton MK (2015) Package ‘mumin’. Version, 1(18), 439
Becker A, Whitfield AK, Cowley PD, Järnegren J, Næsje TF (2013) Does boat traffic cause displacement of fish in estuaries? Mar Pollut Bull 75:168–173. https://doi.org/10.1016/j.marpolbul.2013.07.043
Becker A, Holland M, Smith JA, Suthers IM (2016) Fish movement through an estuary mouth is related to tidal flow. Estuaries Coasts 39:1199–1207
Bishop MJ, Mayer-Pinto M, Airoldi L, Firth LB, Morris RL, Loke LHL, Hawkins SJ, Naylor LA, Coleman RA, Chee SY, Dafforn KA (2017) Effects of ocean sprawl on ecological connectivity: impacts and solutions. J Exp Mar Biol Ecol 492:7–30. https://doi.org/10.1016/j.jembe.2017.01.021
Borsje BW, van Wesenbeeck BK, Dekker F, Paalvast P, Bouma TJ, van Katwijk MM, de Vries MB (2011) How ecological engineering can serve in coastal protection. Ecol Eng 37:113–122
Carlson AK, Fincel MJ, Longhenry CM, Graeb BD (2016) Effects of historic flooding on fishes and aquatic habitats in a Missouri River delta. J Freshw Ecol 31:271–288
Chapman M, Blockley D (2009) Engineering novel habitats on urban infrastructure to increase intertidal biodiversity. Oecologia 161:625–635
Colton MA, Swearer SE (2010) A comparison of two survey methods: differences between underwater visual census and baited remote underwater video. Mar Ecol Prog Ser 400:19–36
Cooper J (2002) The role of extreme floods in estuary-coastal behaviour: contrasts between river-and tide-dominated microtidal estuaries. Sed Geol 150:123–137
Dauer DM, Ranasinghe JA, Weisberg SB (2000) Relationships between benthic community condition, water quality, sediment quality, nutrient loads, and land use patterns in Chesapeake Bay. Estuaries 23:80–96
Dittmann S, Baring R, Baggalley S, Cantin A, Earl J, Gannon R, Keuning J, Mayo A, Navong N, Nelson M (2015) Drought and flood effects on macrobenthic communities in the estuary of Australia’s largest river system. Estuar Coast Shelf Sci 165:36–51
Dolbeth M, Vendel AL, Pessanha A, Patrício J (2016) Functional diversity of fish communities in two tropical estuaries subjected to anthropogenic disturbance. Mar Pollut Bull 112:244–254. https://doi.org/10.1016/j.marpolbul.2016.08.011
Duffy JE, Cardinale BJ, France KE, McIntyre PB, Thébault E, Loreau M (2007) The functional role of biodiversity in ecosystems: incorporating trophic complexity. Ecol Lett 10:522–538
EHMP (2023) Healthy waterways and catchments: ecosystem health monitoring program. http://healthywaterways.org. Accessed 1 Feb 2023
Ellis D, DeMartini E (1995) Evaluation of a video camera technique for indexing abundances of juvenile pink snapper, Pristipomoides filamentosus, and other Hawaiian insular shelf fishes. Oceanogr Lit Rev 9:786
Fairchild TP, Bennett WG, Smith G, Day B, Skov MW, Möller I, Beaumont N, Karunarathna H, Griffin JN (2021) Coastal wetlands mitigate storm flooding and associated costs in estuaries. Environ Res Lett 16:074034
Gibson R, Barnes M, Atkinson R (2002) Impact of changes in flow of freshwater on estuarine and open coastal habitats and the associated organisms. Oceanogr Mar Biol Annu Rev 40:233
Gilby BL, Olds AD, Connolly RM, Maxwell PS, Henderson CJ, Schlacher TA (2018) Seagrass meadows shape fish assemblages across estuarine seascapes. Mar Ecol Prog Ser 588:179–189
Gillanders BM, Elsdon TS, Halliday IA, Jenkins GP, Robins JB, Valesini FJ (2011) Potential effects of climate change on Australian estuaries and fish utilising estuaries: a review. Mar Freshw Res 62:1115–1131
Gillson J, Suthers I, Scandol J (2012) Effects of flood and drought events on multi-species, multi-method estuarine and coastal fisheries in eastern Australia. Fish Manag Ecol 19:54–68
Goodridge Gaines LA, Henderson CJ, Mosman JD, Olds AD, Borland HP, Gilby BL (2022) Seascape context matters more than habitat condition for fish assemblages in coastal ecosystems. Oikos 2022:e09337. https://doi.org/10.1111/oik.09337
Harrison LM, Coulthard TJ, Robins PE, Lewis MJ (2022) Sensitivity of estuaries to compound flooding. Estuaries Coasts 45:1250–1269
Henderson CJ, Olds AD, Lee SY, Gilby BL, Maxwell PS, Connolly RM, Stevens T (2017) Marine reserves and seascape context shape fish assemblages in seagrass ecosystems. Mar Ecol Prog Ser 566:135–144
Henderson CJ, Gilby BL, Schlacher TA, Connolly RM, Sheaves M, Flint N, Borland HP, Olds AD (2019) Contrasting effects of mangroves and armoured shorelines on fish assemblages in tropical estuarine seascapes. ICES J Mar Sci. https://doi.org/10.1093/icesjms/fsz007
Henderson CJ, Gilby BL, Schlacher TA, Connolly RM, Sheaves M, Maxwell PS, Flint N, Borland HP, Martin TSH, Gorissen B, Olds AD (2020) Landscape transformation alters functional diversity in coastal seascapes. Ecography 43:138–148. https://doi.org/10.1111/ecog.04504
Irlandi EA, Crawford MK (1997) Habitat linkages: the effect of intertidal saltmarshes and adjacent subtidal habitats on abundance, movement, and growth of an estuarine fish. Oecologia 110:222–230. https://doi.org/10.1007/s004420050154
Jones TR, Henderson CJ, Olds AD, Connolly RM, Schlacher TA, Hourigan BJ, Goodridge Gaines LA, Gilby BL (2020) The mouths of estuaries are key transition zones that concentrate the ecological effects of predators. Estuaries Coasts. https://doi.org/10.1007/s12237-020-00862-6
Kimmerer W (2002) Effects of freshwater flow on abundance of estuarine organisms: physical effects or trophic linkages? Mar Ecol Prog Ser 243:39–55
Krumme U (2009) Diel and tidal movements by fish and decapods linking tropical coastal ecosystems. In: Nagelkerken I (ed) Ecological connectivity among tropical coastal ecosystems. Springer, pp 271–324
Lacerda CHF, Barletta M, Dantas DV (2014) Temporal patterns in the intertidal faunal community at the mouth of a tropical estuary. J Fish Biol 85:1571–1602. https://doi.org/10.1111/jfb.12518
Meynecke JO, Lee SY, Duke NC (2008) Linking spatial metrics and fish catch reveals the importance of coastal wetland connectivity to inshore fisheries in Queensland, Australia. Biol Conserv 141:981–996. https://doi.org/10.1016/j.biocon.2008.01.018
Morris RL, Konlechner TM, Ghisalberti M, Swearer SE (2018) From grey to green: efficacy of eco-engineering solutions for nature-based coastal defence. Glob Change Biol 24:1827–1842
Morrisey DJ, Turner SJ, Mills GN, Williamson RB, Wise BE (2003) Factors affecting the distribution of benthic macrofauna in estuaries contaminated by urban runoff. Mar Environ Res 55:113–136
Mosman JD, Gilby BL, Olds AD, Goodridge Gaines LA, Borland HP, Henderson CJ (2023) Multiple fish species supplement predation in estuaries despite the dominance of a single consumer. Estuaries Coasts 46:891–905. https://doi.org/10.1007/s12237-023-01184-z
Nagelkerken I (2009) Ecological connectivity among tropical coastal ecosystems. Springer, Berlin
Nearmap Nearmap-Photomap Aerial Imagery (2009) http://www.nearmap.com. Accessed 1 Feb 2023
Olds AD, Connolly RM, Pitt KA, Maxwell PS (2012a) Habitat connectivity improves reserve performance. Conserv Lett 5:56–63. https://doi.org/10.1111/j.1755-263X.2011.00204.x
Olds AD, Pitt KA, Maxwell PS, Connolly RM (2012b) Synergistic effects of reserves and connectivity on ecological resilience. J Appl Ecol 49:1195–1203. https://doi.org/10.1111/jpe.12002
Olds AD, Pitt KA, Maxwell PS, Babcock RC, Rissik D, Connolly RM (2014) Marine reserves help coastal systems cope with extreme weather. Glob Change Biol 20:3050–3058
Olley J, Wilkinson S, Caitcheon G, Read A (2006) Protecting Moreton Bay: how can we reduce sediment and nutrient loads by 50%. In: Procedings of the 9th international river symposium, Brisbane, Australia, vol 47, p 19
Payne NL, van der Meulen DE, Suthers IM, Gray CA, Walsh CT, Taylor MD (2015) Rain-driven changes in fish dynamics: a switch from spatial to temporal segregation. Mar Ecol Prog Ser 528:267–275
Pittman SJ, Caldow C, Hile SD, Monaco ME (2007) Using seascape types to explain the spatial patterns of fish in the mangroves of SW Puerto Rico. Mar Ecol Prog Ser 348:273–284
Preen AR, Lee Long WJ, Coles RG (1995) Flood and cyclone related loss, and partial recovery, of more than 1000 km2 of seagrass in Hervey Bay, Queensland, Australia. Aquat Bot 52:3–17. https://doi.org/10.1016/0304-3770(95)00491-h
Quantum GIS (2023) Quantum GIS geographic information system. Open Source Geospatial Foundation Project
Queensland Government (2019) Regional ecosystem map**. In: Queensland Government (ed) Brisbane, Australia
R Core Team (2023) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Reithmaier GMS, Chen X, Santos IR, Drexl MJ, Holloway C, Call M, Álvarez PG, Euler S, Maher DT (2021) Rainfall drives rapid shifts in carbon and nutrient source-sink dynamics of an urbanised, mangrove-fringed estuary. Estuar Coast Shelf Sci 249:107064. https://doi.org/10.1016/j.ecss.2020.107064
Robins PE, Skov MW, Lewis MJ, Giménez L, Davies AG, Malham SK, Neill SP, McDonald JE, Whitton TA, Jackson SE (2016) Impact of climate change on UK estuaries: a review of past trends and potential projections. Estuar Coast Shelf Sci 169:119–135
Rogers K, Woodroffe CD (2016) Geomorphology as an indicator of the biophysical vulnerability of estuaries to coastal and flood hazards in a changing climate. J Coast Conserv 20:127–144
Romme WH, Everham EH, Frelich LE, Moritz MA, Sparks RE (1998) Are large, infrequent disturbances qualitatively different from small, frequent disturbances? Ecosystems 1:524–534. https://doi.org/10.1007/s100219900048
Sheaves M, Johnston R (2008) Influence of marine and freshwater connectivity on the dynamics of subtropical estuarine wetland fish metapopulations. Mar Ecol Prog Ser 357:225–243
Sheaves M, Johnston R, Connolly RM (2012) Fish assemblages as indicators of estuary ecosystem health. Wetlands Ecol Manag 20:477–490
Sheaves M, Baker R, Nagelkerken I, Connolly RM (2015) True value of estuarine and coastal nurseries for fish: incorporating complexity and dynamics. Estuaries Coasts 38:401–414
Stobart B, García-Charton JA, Espejo C, Rochel E, Goñi R, Reñones O, Herrero A, Crec’hriou R, Polti S, Marcos C, Planes S, Pérez-Ruzafa A (2007) A baited underwater video technique to assess shallow-water Mediterranean fish assemblages: methodological evaluation. J Exp Mar Biol Ecol 345:158–174. https://doi.org/10.1016/j.jembe.2007.02.009
Sutton-Grier AE, Wowk K, Bamford H (2015) Future of our coasts: the potential for natural and hybrid infrastructure to enhance the resilience of our coastal communities, economies and ecosystems. Environ Sci Policy 51:137–148
Taherkhani M, Vitousek S, Barnard PL, Frazer N, Anderson TR, Fletcher CH (2020) Sea-level rise exponentially increases coastal flood frequency. Sci Rep 10:1–17
Teichert N, Carassou L, Sahraoui Y, Lobry J, Lepage M (2018) Influence of intertidal seascape on the functional structure of fish assemblages: implications for habitat conservation in estuarine ecosystems. Aquat Conserv Mar Freshw Ecosyst 28:798–809
Temmerman S, Horstman EM, Krauss KW, Mullarney JC, Pelckmans I, Schoutens K (2022) Marshes and mangroves as nature-based coastal storm buffers. Annu Rev Mar Sci 15:95–118
Thrush SF, Halliday J, Hewitt JE, Lohrer AM (2008) The effects of habitat loss, fragmentation and community homogenization on resilience in estuaries. Ecol Appl 18:12–21. https://doi.org/10.1890/07-0436.1
Turner MG, Gardner RH (2015) Landscape disturbance dynamics. In: Turner MG, Gardner RH (eds) Landscape ecology in theory and practice. Springer, pp 175–228
Villéger S, Miranda JR, Hernández DF, Mouillot D (2010) Contrasting changes in taxonomic vs. functional diversity of tropical fish communities after habitat degradation. Ecol Appl 20:1512–1522. https://doi.org/10.1890/09-1310.1
Waltham N, Connolly R (2007) Artificial waterway design affects fish assemblages in urban estuaries. J Fish Biol 71:1613–1629
Wang Y, Naumann U, Wright ST, Warton DI (2012) mvabund—An R package for model-based analysis of multivariate abundance data. Methods Ecol Evol 3:471–474
Wang Y, Naumann U, Wright S, Warton D, Wang MY, Rcpp I (2022) Package ‘mvabund’
Westfall PH, Young SS (1993) Resampling-based multiple testing: examples and methods for P value adjustment. Wiley series in probability and statistics, vol 279. Wiley
Wetz MS, Yoskowitz DW (2013) An ‘extreme’ future for estuaries? Effects of extreme climatic events on estuarine water quality and ecology. Mar Pollut Bull 69:7–18
Whitfield AK (2017) The role of seagrass meadows, mangrove forests, salt marshes and reed beds as nursery areas and food sources for fishes in estuaries. Rev Fish Biol Fish 27:75–110. https://doi.org/10.1007/s11160-016-9454-x
Whitfield A, Adams J, Bate G, Bezuidenhout K, Bornman T, Cowley P, Froneman P, Gama P, James N, Mackenzie B (2008) A multidisciplinary study of a small, temporarily open/closed South African estuary, with particular emphasis on the influence of mouth state on the ecology of the system. Afr J Mar Sci 30:453–473
Williams J, Hindell JS, Jenkins GP, Tracey S, Hartmann K, Swearer SE (2017) The influence of freshwater flows on two estuarine resident fish species show differential sensitivity to the impacts of drought, flood and climate change. Environ Biol Fish 100:1121–1137
Wood S, Wood MS (2015) Package ‘mgcv’. R package version, 1(29), 729
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This project was funded by Healthy Land and Water and University of the Sunshine Coast. The authors thank Taylor Cooper, Tyson Jones, Cody James, Edward Hay and Nicholas Ortodossi for their assistance with fieldwork and analysing videos.
Author information
Authors and Affiliations
Contributions
CH, LGG, JM, AO and BG conceptualised the project. All authors contributed to fieldwork. CH led analysis and the writing of the manuscript. All authors contributed to the manuscript drafting.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest are present for this study.
Ethic approval
An animal ethics exemption was granted by the University of the Sunshine Coast Animal Ethics Committee as the project used unbaited camera deployments.
Additional information
Responsible Editor: W. Figueira.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Henderson, C.J., Olds, A.D., Goodridge Gaines, L.A. et al. Flood effects on estuarine fish are mediated by seascape composition and context. Mar Biol 171, 138 (2024). https://doi.org/10.1007/s00227-024-04459-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-024-04459-6