
Abstract
Climate change and the underlying causal factors have been thoroughly described in the scientific literature. Although well understood and documented in general, we are only beginning to understand the effects of climate change on biotic systems, such as insect communities. It seems obvious that insect pest problems should be be more important in a warmer climate; temperature has a strong impact on insect physiology and demography. It must be remembered, however, that insect distribution and abundance are controlled by many factors other than temperature, e.g. host plant, natural enemies. This chapter summarizes empirical evidence for climate change-induced insect pest problems, i.e. changed distribution range and frequency of insect outbreaks. The chapter reviews cases of forest pests for which there is sufficient scientific evidence of climate change-effects on their population dynamics leading to outbreaks, strong enough to be a concern for forest ecology and management.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


22.1 Introduction
Climate change and the underlying causal factors have been thoroughly described (Field et al. 2014). Climate change, particularly increased temperature, has several consequences for the functioning of ecosystems. For instance, we know that the distribution range of some organisms has changed (Parmesan et al. 1999), tree phenology altered (Walther et al. 2002), and phenological asynchrony developed, e.g. between tree and associated insects (Visser and Both 2006). Although these effects are well understood and documented, we are only beginning to understand the effects of climate change on insect communities. This is in large part because of the complexity of their interactions with the abiotic and biotic environment.
It seems obvious that insect pest problems will be more important in a warmer climate because of the strong positive effect that temperature has on insect physiology and demography (Ayres and Lombardero 2000). However, temperature increases above optimal ranges may also be detrimental to insect fitness (Lehmann et al. 2020). In addition, it must be remembered that insect distribution and abundance are controlled by many factors other than temperature.
Klapwijk et al. (2012) reviewed climate-change associated factors affecting the outbreak potential of forest insects. They identified direct and indirect factors and provided a theoretical framework for assessing how changes in climate can be incorporated into predictive models of insect population dynamics. Similarly, Battisti and Larsson (2015) and Jactel et al. (2019) reviewed how climate change can affect the distribution range of insect pests, and provided examples of forest insect species whose ranges have been changed in a manner consistent with changes in climate.
This chapter summarizes empirical evidence for climate-change induced insect pest problems, i.e. changed distribution range and frequency of insect outbreaks. Climate change can interact with non-native insect species accidently introduced into novel areas (Brockerhoff and Liebhold 2017). The issue of invasions is discussed in Chapter 23 of this volume. In this chapter we briefly discuss, in general terms, if and how climate change can be a factor that contributes to non-native insect species being established and becoming invasive, i.e. acting as novel pests in the forest.
Throughout the chapter the focus is on how climate change affects the distribution and abundance of forest pests (directly and indirectly through biotic interactions). We acknowledge that climate change will also influence host tree vulnerability and tolerance, and thus potential future damage (Toïgo et al. 2020; Forzieri et al. 2021) (discussed in Chapter 20).
22.2 Climatic Drivers
There is general consensus among scientists that the global climate is changing at an unprecedented rate, with many regions experience warming trends, shifts in precipitation patterns, and more frequent extreme weather events (Field et al. 2014). Factors potentially affecting forest insects include temperature, precipitation, rare weather events such as wind storms and heat waves, and atmospheric carbon dioxide concentration. All these factors can act both directly and indirectly (through host plant or natural enemies) on insect pests.
Temperature is the most important driver because it has steadily increased since the beginning of the twentieth century (0.61 °C in global mean temperature from 1850–1900 to 1986–2005; Field et al. 2014), and is predicted to increase further. Forests experience different levels of climate change depending on geographic position. Upper latitudes of northern and southern hemispheres, where most of the world’s temperate and boreal forests grow, are expected to experience a higher warming. Insects, being poikilothermic organisms, respond directly to temperature as described by their specific reaction norms. Temperature also affects insects indirectly through effects on the host plant (bottom-up, see Chapter 7) and natural enemies (top-down, see Chapter 6).
Patterns in precipitation are due to a complex interaction between air circulation and temperature. Thus, an effect of temperature increase on precipitation patterns is expected. The result, however, is not as clear as the one depicted for temperature alone. Predictions on the total amount of annual precipitation vary according to the geographic area, with upper latitudes of both hemispheres experiencing more precipitation than mid latitudes, while the tropical and subtropical regions show a patchy effect (Field et al. 2014). Precipitation is also characterized by two more aspects, i.e. its distribution in the year and the intensity of the precipitation events. At upper latitudes of the northern hemisphere, precipitation increase will mainly occur in winter, while intense precipitation events will be more likely everywhere. Forests will thus experience different precipitation regimes according to geographic region. The interactions of these changes with those of temperature and solar radiation (through modified cloudiness) will likely modify the microclimatic niche experienced by forest insects. Although precipitation is known to directly affect forest insects, most of its action is indirect because water availability is crucial for tree growth, and consequently, host quality for insect herbivores.
Extreme rare weather events, such as high/low temperature and rainfall, strong wind, and their combinations, will probably occur in higher frequency, and this is considered a potentially important component of climate change (Field et al. 2014). Two factors characterize the nature of these events, timing and intensity. For example, a heat wave may suppress all the insects active in that moment because the upper thermal threshold is achieved (see Chapter 4), or a wind storm may simultaneously fell a large number of trees that may facilitate a bark beetle outbreak (see Chapter 10). The periods when such events may happen are roughly predictable, because they are associated with the yearly variation of both temperature and precipitation, although it is impossible to define exactly when and where they will occur.
Carbon dioxide, together with other greenhouse gases, is a major determinant of temperature increase (Field et al. 2014). Being a fundamental molecule for photosynthesis, the increase in carbon dioxide in the atmosphere may affect the metabolism of forest trees, including molecules of importance to tree-feeding insects (Lindroth et al. 1993), although the general effects on herbivorous insects are weak and idiosyncratic (Hillstrom et al. 2014).
The climate in the future will most likely differ in all the above-mentioned aspects. However, with respect to effects on forest insects temperature has by far been the most discussed in the literature, and thus is the factor for which there exists a reasonable amount of data. Therefore, it will be the focus of discussion in the following sections.
22.3 Insect Response to Increased Temperature
In this section we deal with temperature effects at the level of the individual insect. Temperature has a direct effect on insect development rate and survival. Development rate generally increases with increasing temperature to some maximum, above which development slows down and mortality increases (see Chapter 4). Increased development rate could lead to increased voltinism in facultative multivoltine species. Increased development rate in insect larvae could result in reduced temporal exposure to enemies or other mortality agents, with resulting higher survival.
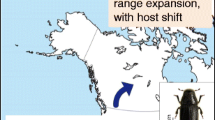
Winter mortality is likely to decrease under increasing temperatures (e.g. Ayres and Lombardero 2000), although decreased snow cover (and therefore decreased insulation of overwintering sites) can reverse that pattern (Petrucco-Toffolo and Battisti 2008). Warmer winters may permit some non–diapausing species to continue feeding and development during months that were previously too cold (Schneider et al. 2021). For example, larvae of the pine processionary moth Thaumetopoea pityocampa have a higher probability of survival if winter temperatures do not often fall below specific feeding thresholds (Battisti et al. 2005, Fig. 22.1A, B).

A The same tree can be colonized in subsequent years as shown in the photo where the remains of an old nest are visible close to two new nests with white silk. The tree is a Scots pine (Pinus sylvestris) growing at high elevation (>1,400 m) in the Southern Alps (Venosta/Vinschgau valley) where the insect has expanded its distribution in recent decades and reached the upper limit of host plant range. At even higher elevations in the Alps, the pine processionary moth is massively colonizing the dwarf mountain pine (Pinus mugo), historically not a suitable host because of being covered by snow. B The photo shows a colony that survived the winter because the limited snow cover, and higher temperatures, permitted suitable conditions for larval feeding across the winter.
Several indirect effects mediated through the host tree exist. Many insect species match their feeding activity with certain developmental stages in the host plant; for example, species associated with deciduous trees, such as the autumnal moth Epirrita autumnata, match their feeding with nutritious immature foliage during spring and early summer (Haukioja 2003).
If host trees are reasonably well matched to historically favorable climatic conditions, then it is inevitable that changing climate will lead to situations where trees are poorly matched to the new conditions, i.e. trees can be stressed. Stress-induced changes in plant tissue quality and their effects on insect survival and reproduction are well documented in experimental studies (Koricheva et al. 1998).
That plant stress can trigger insect outbreaks is a long-standing hypothesis in forest entomology. Insect outbreaks have been commonly correlated with conditions that induce stress in their host plants (e.g. Mattson and Haack 1987). This has led to speculation that there is a causal link between stress-induced changes in plant quality, and thus insect performance, and the start of outbreaks (e.g. White 1974). Experimental tests of the plant stress hypothesis, most often at the level of individual insects, have produced mixed results; species from some feeding guilds respond to experimentally stressed trees with increased performance, some are unaffected, and some respond negatively (Larsson 1989). Bark beetles constitute a globally important group of insects for which plant stress seems relevant; a long-standing paradigm is that healthy trees are resistant to most bark beetle species (and other boring insects), but that periods of stress make trees susceptible, although at high beetle density even non-stressed trees can be attacked and killed (Raffa et al. 2008).
Arthropod natural enemies can exert powerful forces on the performance of herbivorous insects (see Chapter 6), and climate change may affect their activity as much as that of their prey. In addition, the phenological synchrony between natural enemies and their hosts/prey can also be affected. However, specialist enemies should be under strong selection to track phenological changes in their prey, which might make them less likely to become temporally uncoupled from their prey (Klapwijk et al. 2010). Higher temperatures can influence parasitism and predation rates by increasing searching activity of individual parasitoids and predators. When the prey are relatively immobile (e.g. many immature insect herbivores), this should generally increase rates of detection and attack.
Insect pathogens, e.g. fungi, bacteria, and viruses, can also limit the performance of herbivorous insects (Hajek 1997). Temperature can be important for both infection rate and defense responses within the host. Different thermal optima for host and pathogen might lead to a situation where high temperatures favor the host by both optimizing defense responses and directly limiting pathogen growth (Blanford and Thomas 1999).
Insects rarely, if ever, act independently from other organisms. Therefore, it is necessary to consider the position of the forest insect in the trophic web. In other words, not only should we consider direct effects of climate change on the target insect, but we need to recognize likely interactions of climatic variables with host tree, natural enemies, and insect diseases (indirect effects). Such an approach is necessary to fully understand the potential consequences that climate change can have for pests. For climate-change driven effects to have an impact on forest ecosystems, and thus be of economic concern for forest managers, effects at the level of individual insects need to be confirmed at the level of the population. This is not a trivial step as a multitude of biotic interactions, each with its inherent uncertainty, can modify the effects when it comes to populations, as discussed in the following section.
22.4 Insect Population Response to Increased Temperature
Climate warming can influence two important aspects of insect population ecology: distribution and abundance. Many insect species have been documented to change their distribution range in response to increased temperature (Battisti and Larsson 2015). It is important to realize, however, that the dynamics of range expansion are rarely known in any detail. This is simply because populations in expansion areas are initially at very low density and thus can remain undetected for a long time. If the expanding population is a forest pest, then the expansion is likely to be discovered if the population reaches outbreak numbers.
In general, the majority of insect populations are controlled by a number of different agents and thus occur at low density. Under some conditions an insect species may escape from the controlling agents and reach outbreak densities. Outbreaks are easily observed because they are generally defined by managers as population densities so high that they are of economic concern in forestry. Outbreaks can thus be seen as a proxy for high-density events, and of course as a warning signal of potential forest health problems. Low-density populations will also vary in size, but their dynamics will most of the time be unnoticed because their densities will not result in damage to the forest.
In the following section, we present case studies to illustrate effects of climate change on insect populations. The focus is on outbreaks, which we assign to three main groups. The first group of case studies refer to Outbreaks at the core of historical range of distribution, thus evidence of climate change effects (or lack of effects) on populations in the historic range of the distribution. The second group deals with New areas of outbreaks within historic species distribution. This refers to species where no outbreaks were recorded for a portion of their historical distribution, typically in the colder areas, but where outbreaks in recent years have been observed. The third group includes Outbreaks in recently invaded geographic areas, in other words, species that have expanded their distribution range and occurred at outbreak densities that clearly can be related to warming.
22.4.1 Outbreaks at the Core of Historical Range of Distribution
Long-term surveillance data of European insect populations report a large variability in the responses of key forest pests to climate change: positive, negative, and no response to increased temperatures (Haynes et al. 2014; Lehmann et al. 2020).
The European spruce beetle Ips typographus is the most aggressive bark beetle in Europe. Analysis of 17 time-series spanning from 1980 to 2010 shows density-dependent factors to be the main drivers of population dynamics, although high temperature and summer precipitation deficit also play a role (Marini et al. 2017). In addition, temperature appears to be important for the voltinism of I. typographus, as populations may become bi- or multivoltine under favorable conditions (Wermelinger 2004). Results suggest that greater efforts should be made to integrate temperature increase, drought, and storm effects into future scenarios of outbreaks under climate change (Marini et al. 2017).
The larch bud moth Zeiraphera griseana is an example of a pest where climate warming has had negative effects on population growth. Dendrochronological analyses of host trees associated with Z. griseana outbreaks over 500 years reveal periodicities of 4, 8, and 16 years throughout the time series, except during the period 1690–1790, and since 1980. The data suggest a disruption of periodicity probably related to changes in climate; temperature decreased in the period 1690–1790 (Little Ice Age) wheras it increased since 1980 (Saulnier et al. 2017).
Responses at the species level appear idiosyncratic and no general patterns were observed in several species of defoliating insects associated with coniferous trees in southern Germany for more than 200 years (Haynes et al. 2014). A similar study in Hungary involving five species of defoliating insects associated with broadleaved tree species for a period of about 60 years also observed no clear pattern in responses (Klapwijk et al. 2013). It should be noted that for both these studies data refer to large scale events, and that changes at local scale could have gone undetected. More precise data are available for the pine processionary moth Thaumetopoea pityocampa from eight geographic zones in France (but for a shorter period, 1981–2014). Although in general, populations were controlled mainly by density-dependent agents, population growth was negatively related to precipitation in five regions and positively related to winter temperature in four regions; thus, these data suggest that the effects of weather-related factors need to be considered at a local scale using appropriate measures of population density (Toïgo et al. 2017).
22.4.2 New Areas of Outbreaks Within Historic Species Distribution
Both the autumnal moth Epirrita autumnata and the winter moth Operophtera brumata have expanded their outbreak range in recent years, presumably as a result of improved winter survival of eggs, and maintenance of synchrony (through adaptive phenological plasticity) with bud burst of their main host, the mountain birch Betula pubescens ssp. tortuosa (Jepsen et al. 2008). Winter moth populations show a pronounced north-eastern expansion of outbreaks into areas previously dominated by the autumnal moth, which in turn has expanded historically into colder areas (Tenow 1996). This has been possible because eggs of the autumnal moth are more cold tolerant than those of the winter moth. This important direct effect of increased temperature can be affected by indirect effects in the trophic interactions and in the synchronization with the bud break of the host plants. In subarctic mountain birch forests, predation rates on E. autumnata and O. brumata larvae were almost twice as high in low versus high elevation sites, indicating that release from predation pressure at high elevations can favor outbreaks in these cooler habitats (Pepi et al. 2017).
Records of spruce budworm (Choristoneura fumiferana) defoliation and tree-ring analysis indicate that the outbreak range of this insect has expanded to the north. A regional tree-ring chronology performed by Boulanger et al. (2012) represents the longest and most replicated reconstruction of outbreak dynamics in North America (1551–1995). The authors identified nine potential outbreaks and three uncertain outbreaks in a 400-year period and concluded that outbreak frequency varied with temperature, being less frequent during the 1660–1850 period (every ~ 50 years, Little Ice Age) and more frequent in warmer periods like prior to 1660 (every ~ 28 years) and during the twentieth century (every ~ 30 years). The simultaneous occurrence of a general increase in temperature in northern latitudes at the start of the last outbreak indicates a relation with climate change (Candau and Fleming 2011). An interesting indirect effect involving the host plant has been suggested. The main host of the spruce budworm is balsam fir Abies balsamea, whereas black spruce Picea mariana is a secondary host. Climate change is predicted to advance the phenology of the secondary host that is more abundant at the upper latitudinal edge, making it more susceptible to defoliation, and thus facilitating expansion of the outbreak area into higher latitudes (Pureswaran et al. 2015). This factor has been hypothesized to explain the occurrence of the new outbreak that started in 2006 about four degrees (445 km) of latitude north of the previous one (1966–1992), with a prediction for a more northern expansion in 2041–2070 (Régnière et al. 2012).
In the southern hemisphere, the defoliation of Nothofagus forests by the saturnid moths of the genus Ormiscodes have been associated with drier and warmer seasons. The outbreaks have been more frequent in southern than in northern Patagonia. Results are consistent with recent warming in southern Patagonia and suggest that outbreak frequency may continue to increase with further warming (Paritsis and Veblen 2011).
22.4.3 Outbreaks in Recently Invaded Geographic Areas
In recent decades, the pine processionary moth T. pityocampa has expanded its latitudinal and elevational distribution range (Battisti et al. 2005). Improved survival during the feeding period in winter has contributed to outbreaks in pine forests previously unoccupied in France, Italy, Spain, and Turkey. Rapid range expansion is facilitated by warm summer nights that contribute to long-distance (more than 2 km) dispersal of female moths (Battisti et al. 2006). In the newly occupied areas, however, population dynamics are driven more by density-dependent agents than by climatic drivers (Tamburini et al. 2013). Thus, once the expansion area is occupied population dynamics seem to be determined by the same factors as in the historical range, provided that specialist enemies have tracked the host in the new areas. Interestingly, T. pityocampa shows prolonged diapause facilitating persistence in the newly colonized areas even if the weather turns unfavorable for one or more years; diapause can last up to eight years with some individuals emerging every year (Salman et al. 2016).
In western Canada recent outbreaks of the mountain pine beetle Dendroctonus ponderosae have led to extensive tree mortality within at least 14 million hectares of lodgepole pine Pinus contorta forests. The start of the outbreak was facilitated by fire suppression during the last century, which created large tracts of over-mature pine stands, in combination with recent climatic patterns, viz. mild winters and warm dry summers (Raffa et al. 2008; Bentz and Jönsson 2015). However, the relative importance of large areas of susceptible pine forests and suitable climatic conditions for beetle population growth is not entirely clear (Cooke and Carroll 2017).
In general, bark beetle species associated with weakened trees are difficult to detect at low-densities, whereas damage and tree mortality become obvious during outbreaks. Therefore, the range edge generally considered is that of the epidemic range, whereas the margins of the endemic range remain largely unknown. In southeast USA, the distribution of the southern pine beetle Dendroctonus frontalis has been moved northwards due to milder winters that enhance beetle performance (Ungerer et al. 1999). Similarly, spruce bark beetle (Dendroctonus rufipennis) outbreaks may occur throughout the range of spruce in North America in the future. In its coldest locations, D. rufipennis is semivoltine, having a generation every two years and outbreaks are rare in these populations (Schebeck et al., 2017).
22.5 Invasive Species and Climate Change
The increasing problem with invasive species during the last decades may be linked to climate change although the evidence for this remain limited. Global trade and travel are the major drivers of the invasion process (Ramsfield et al. 2016; Brockerhoff and Liebhold 2017). The process of invasion is often divided into several phases (pre-transport, transport, arrival, establishment, and spread; see Chapter 23). Here we briefly discuss how changes in climate can interact with trade and travel in each of the invasion phases.
Very little can be said about the pre-establishment phases (pre-transport, transport, arrival). It is obvious that propagule pressure in the area of origin is important in order to assess the probability of transportation, but it is unclear to what extent changes in climate affect propagule pressure. The next two phases (establishment and spread) are clearly linked to climate change as they depend on the matching between the area of origin and the area of arrival. The impact of climate change on climate matching between areas of origin and destination on the establishment and spread of non-native species is difficult to assess as data about failure to establish are rarely available for forest insects. The increasing number of the incursions of ambrosia beetles from tropical and subtropical regions in temperate forests could be an example of how this category of organisms is favored by climate change (Rassati et al. 2016). The inclusion of climatic responses of pests in the risk assessment of invasive species may help to predict which ones are the most likely to get established and threat newly colonized habitats (Grousset et al. 2020).
The spread of invasive species in a newly colonized area depend on niche availability and dispersal traits of the insect, and in principle does not differ from that of native species (Pureswaran et al. 2018). The hemlock woolly adelgid Adelges tsugae was introduced into Virgina, eastern USA in the mid 1900s. Increase in mean minimum winter temperature resulted in higher survival in overwintering life stages and facilitated the expansion northwards in the eastern USA (Paradis et al. 2008). The build up of high density populations in the already colonized areas contributed greatly to hemlock (Tsuga canadensis, T. caroliniana) dieback (Fitzpatrick et al. 2012).
Once established, the response of invasive species to climatic factors may be similar to that of native species, as illustrated by the spongy moth Lymantria dispar. The population dynamics of L. dispar have been thoroughly documented in the USA, showing periods of cyclic outbreaks intermingled by periods with no cycles. The dynamics seem to be driven by trophic interactions while the role of climate appears to be negligible (Allstadt et al. 2013). It is not clear if a changing climate would cause a net increase in suitable habitat for invading insects such as spongy moth in North America, as there should be some areas that become more favorable and others that become less favorable (Tobin et al. 2014).
22.6 Conclusions
Climate change, in particular increased temperature, is certain to have qualitative and quantitative effects on insect populations, primarily because temperature ultimately sets the limit for most insect distribution ranges (Battisti and Larsson 2015). For insects on trees, however, the availability of the host tree(s) will be a critical factor because most insect species are associated with one or a few host tree species only. The expected slower range expansion by trees compared with that of the insects, because of the much longer generation time of trees, will likely slow down the successful expansion of the insects. Overall, this probably means that at a certain point in time host tree availability, rather than temperature, may set the limit for future insect range expansion. This scenario would only apply to insect species not able to switch to novel host tree species in the expansion area.
Climate-change attention is mostly on insect species expanding their range into geographic areas that have become climatically more favorable. We have to assume, however, that an equally large area may become unfavorable. In contrast to expansion, such retraction of the range at the lower edge of the distribution will not be as immediate as the expansion, mainly because plants do not react as quickly to the warming as insects do. This is why a net increase in areal distribution is expected in the short-term while in a medium-long term a general shift of the range is predicted.
A difficult task is to assess whether or not damage to forests will be more, or less, severe under climate change (Jactel et al. 2019; Lehmann et al. 2020). The degree of damage is usually positively related to the density of the insect pest population. Thus, we can reformulate the issue using outbreak as a proxy for damage and ask: are outbreaks likely to be more common under climate change?
Ideally, in order to scientifically analyze this issue we should be able to refer to the frequency of outbreaks for a scenario of no climate warming. Obviously, this is not a straightforward matter, but the literature provides important information about insect populations that can be used as a simple null model of outbreak frequency (Barbosa and Schultz 1987). Most insect populations in forest ecosystems thrive around low mean densities, far below outbreak densities (Landsberg and Ohmart 1989), meaning that they are efficiently controlled by several, mostly unknown factors. It also indicates that many insect populations often remain unnoticed for a long time (endemic) and are considered pests only when they build up epidemic populations (outbreak) (Barbosa et al. 2012).
The categorization of forest insect populations outlined above is simplistic, but still useful as a basis for the following discussion of forest damage and insect pests under future climate change. We envisage four situations:
-
1.
The extent of range expansion of non-outbreak insects is virtually unknown; this should come as no surprise because, by definition, these insect species occur at low density. It is quite likely, however, that such expansion has occurred but should be of minor importance from a management point of view, given that the population ecology of these putative species in the new area is similar to that in their core area.
-
2.
A bias exists in the literature with almost all evidence of climate-change effects coming from outbreak species, for obvious reasons (easy to observe). In the event that outbreaks occur in the expanded range, an important question will then be whether the outbreak dynamics are similar to those in the original distribution range or show new characteristics (the mountain pine beetle outbreak may be an example of this as it has invaded new host tree species, such as Pinus banksiana in Alberta, creating the potential for massive range expansion into north central and eastern north America).
-
3.
Outbreaks in the historical area can be more, or less, frequent under climate change depending on the life history of the insect and how climate affects biotic interactions (with host tree, natural enemies, insect pathogens). Forest management is changing in many parts of the world, e.g. with stands being overall more intensively managed than in the past. So far, there are no data to suggest that pest dynamics are significantly different under intensive forestry, such as nitrogen fertilization of natural stands (Kytö et al. 1996). If novel management practices, e.g. for maximizing carbon sequestration, will be introduced on a large scale, then there is certainly a risk that pest problems will follow.
-
4.
Forest health problems due to non-native insect species will most likely continue to increase in the future. Some non-natives will establish but with dynamics of the low-density type. The distribution of other non-natives will expand, perhaps as a consequence of climate change, and establish in natural forests where populations increase to outbreak level (thus becoming an invasive). An especially serious threat is the situation where non-native insects establish in plantations of non-native tree species. Here managers may be faced with a situation of intensive control practice most often not necessary in traditional forestry, such as the application of biological control with a parasitic nematode against the Sirex wood wasp in pine plantations (Slippers et al. 2015).
The science of outbreak dynamics includes data from economically important insect populations whose dynamics appear to be driven by factors that differ from those of non-outbreak species (e.g. Larsson et al. 1993). Thus, there is no overarching hypothesis based on logic (or data) that allows for specific predictions at the species (or population) level. Our approach has been to use information from the past in order to understand the future. This approach allows us to take advantage of existing scientific knowledge. Although we advocate this approach we emphasize that we also need to appreciate that the available data, and thus predictions based on these data, have a substantial degree of uncertainty. Very rarely, if ever, can outbreak data be considered replicated, due to different boundary conditions. This is a situation that is true for many ecological data sets meant to be used in policy, but is especially troublesome here because we are interested in changes over long periods of time, hundreds of years.
References
Allstadt AJ, Haynes KJ, Liebhold AM, Johnson DM (2013) Long-term shifts in the cyclicity of outbreaks of a forest-defoliating insect. Oecologia 172:141–151
Ayres MP, Lombardero MJ (2000) Assessing the consequences of global change for forest disturbance from herbivores and pathogens. Sci Total Environ 262:263–286
Barbosa P, Schultz JC (1987) Insect outbreaks. Academic Press, New York
Barbosa P, Letourneau DK, Agrawal AA (2012) Insect outbreaks revisited. Academic Press, New York
Battisti A, Stastny M, Netherer S, Robinet C, Schopf A, Roques A, Larsson S (2005) Expansion of geographic range in the pine processionary moth caused by increased winter temperatures. Ecol Appl 15:2084–2096
Battisti A, & Larsson S (2015) Climate change and insect pest distribution range. In C. Björkman & P. Niemelä, editors. Climate change and insect pests. CABI International, pp 1–15
Battisti A, Stastny M, Buffo E, Larsson S (2006) A rapid altitudinal range expansion in the pine processionary moth produced by the 2003 climatic anomaly. Glob Change Biol 12:662–671
Bentz BJ, Jönsson AM (2015) Modeling bark beetle responses to climate change. In: Vega FE, Hofstetter RW (eds) Bark Beetles. Academic Press, San Diego, pp 533–553
Blanford S, Thomas MB (1999) Host thermal biology: the key to understanding host–pathogen interactions and microbial pest control? Agric for Entomol 1:195–202
Boulanger Y, Arseneault D, Morin H, Jardon Y, Bertrand P, Dagneau C (2012) Dendrochronological reconstruction of spruce budworm (Choristoneura fumiferana) outbreaks in southern Quebec for the last 400 years. Can J For Res 42:1264–1276
Brockerhoff EG, Liebhold AM (2017) Ecology of forest insect invasions. Biol Invasions 19:3141–3159
Candau J-N, Fleming R (2011) Forecasting the response of spruce budworm defoliation to climate change in Ontario. Can J For Res 41:1948–1960
Cooke BJ, Carroll AL (2017) Predicting the risk of mountain pine beetle spread to eastern pine forests: considering uncertainty in uncertain times. For Ecol Manage 396:11–25
Field CB, Barros VR, Mach KJ et al. (2014). Climate change 2014: impacts, adaptation, and vulnerability. part A: global and sectoral aspects. Contribution of Working Group II to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, pp 35–94
Fitzpatrick MC, Preisser EL, Porter A, Elkinton J, Ellison AM (2012) Modeling range dynamics in heterogeneous landscapes: invasion of the hemlock woolly adelgid in eastern North America. Ecol Appl 22:472–486
Forzieri G, Girardello M, Ceccherini G, Spinoni J, Feyen L, Hartmann H, Beck PSA, Camps-Valls G, Chirici G, Mauri A, Cescatti A (2021) Emergent vulnerability to climate-driven disturbances in European forests. Nat Commun 12:1081
Grousset F, Grégoire J-C, Jactel H, Battisti A, Benko Beloglavec A, Hrašovec B, Hulcr J, Inward D, Orlinski A, Petter F (2020) The risk of bark and ambrosia beetles associated with imported non-coniferous wood and potential horizontal phytosanitary measures. Forests 11:342
Hajek AE (1997) Ecology of terrestrial fungal entomopathogens. Adv Microb Ecol 15:193–249
Haukioja E (2003) Putting the insect into the birch–insect interaction. Oecologia 136:161–168
Haynes KJ, Allstadt AJ, Klimetzek D (2014) Forest defoliator outbreaks under climate change: Effects on the frequency and severity of outbreaks of five pine insect pests. Glob Change Biol 20:2004–2018
Hillstrom ML, Couture JJ, Lindroth RL (2014) Elevated carbon dioxide and ozone have weak, idiosyncratic effects on herbivorous forest insect abundance, species richness, and community composition. Insect Conservation and Diversity 7:553–562
Jactel H, Koricheva J, Castagneyrol B (2019) Responses of forest insect pests to climate change: not so simple. Current opinion in Insect Science 35:103–108
Jepsen JU, Hagen SB, Ims RA, Yoccoz NG (2008) Climate change and outbreaks of the geometrids Operophtera brumata and Epirrita autumnata in subarctic birch forest: evidence of a recent outbreak range expansion. J Anim Ecol 77:257–264
Klapwijk MJ, Grobler BC, Ward K, Wheeler D, Lewis OT (2010) Influence of experimental warming and shading on host–parasitoid synchrony. Glob Change Biol 16:102–112
Klapwijk MJ, Ayres MP, Battisti A, Larsson S (2012) Assessing the impact of climate change on outbreak potential. In: Barbosa P, Letourneau DK, Agrawal AA (eds) Insect outbreaks revisited. Academic Press, New York, pp 429–450
Klapwijk MJ, Csóka G, Hirka A, Björkman C (2013) Forest insects and climate change: long-term trends in herbivore damage. Ecol Evol 3:4183–4196
Koricheva J, Larsson S, Haukioja E (1998) Insect performance on experimentally stressed woody plants: a meta-analysis. Annu Rev Entomol 43:195–216
Kytö M, Niemelä P, Larsson S (1996) Insects on trees: population and individual response to fertilization. Oikos 75:148–159
Landsberg J, Ohmart C (1989) Levels of insect defoliation in forests: patterns and concepts. Trends Ecol Evol 4:96–100
Larsson S (1989) Stressful times for the plant stress—insect performance hypothesis. Oikos 56:277–283
Larsson S, Björkman C, Kidd NAC (1993) Outbreaks in diprionid sawflies: why some species and not others? In: Wagner MR, Raffa KF (eds) Sawfly life history adaptations to woody plants. Academic Press, San Diego, pp 453–483
Lehmann P, Ammunét T, Barton M, Battisti A, Eigenbrode SD, Jepsen JU, Kalinkat G, Neuvonen S, Niemelä P, Terblanche JS, Økland B, Björkman C (2020) Complex responses of global insect pests to climate warming. Front Ecol Environ 18:141–150
Lindroth RL, Kinney KK, Platz CL (1993) Responses of deciduous trees to elevated atmospheric CO2: productivity, phytochemistry, and insect performance. Ecology 74:763–777
Marini L, Økland B, Jönsson AM, Bentz B, Carroll A, Forster B, Grégoire J-C, Hurling R, Nageleisen LM, Netherer S, Ravn HP, Weed A, Schroeder M (2017) Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography 40:1426–1435
Mattson WJ, Haack RA (1987) The role of drought stress in provoking outbreaks of phytophagous insects. In: Barbosa P, Schultz JC (eds) Insect Outbreaks. Academic Press, San Diego, pp 365–394
Paradis A, Elkinton J, Hayhoe K, Buonaccorsi J (2008) Role of winter temperature and climate change on the survival and future range expansion of the hemlock woolly adelgid (Adelges tsugae) in eastern North America. Mitig Adapt Strat Glob Change 13:541–554
Paritsis J, Veblen TT (2011) Dendroecological analysis of defoliator outbreaks on Nothofagus pumilio and their relation to climate variability in the Patagonian Andes. Glob Change Biol 17:239–253
Parmesan C, Ryrholm N, Stefanescu C, Hill JK, Thomas CD, Descimon H, Huntley B, Kaila L, Kullberg J, Tammaru T, Tennent WJ, Thomas JA, Warren M (1999) Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 399:579–583
Pepi AA, Vinstad OPL, Ek M, Jepsen JU (2017) Elevationally biased avian predation as a contributor to the spatial distribution of geometrid moth outbreaks in sub-arctic mountain birch forest. Ecological Entomology 42:430–438
Petrucco-Toffolo E, Battisti A (2008) Performances of an expanding insect under elevated CO2 and snow cover in the Alps. Iforest Biogeosciences For 1:126–131
Pureswaran DS, De Grandpré LD, Paré D, Taylor A, Barrette M, Morin H, Régnière J, Kneeshaw DD (2015) Climate-induced changes in host tree-insect phenology may drive ecological state-shift in boreal forests. Ecology 96:1480–1491
Pureswaran DS, Roques A, Battisti A (2018) Forest insects and climate change. Curr For Rep 4:35–50
Raffa KF, Aukema BH, Bentz BJ, Carroll AL, Hicke JA, Turner MG, Romme WH (2008) Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. Bioscience 58:501–517
Ramsfield TD, Bentz BJ, Faccoli M, Jactel H, Brockerhoff EG (2016) Forest health in a changing world: effects of globalization and climate change on forest insect and pathogen impacts. Forestry 89:245–252
Rassati D, Faccoli M, Haack RA, Battisti A, Marini L (2016) Bark and ambrosia beetles show different invasion patterns in the USA. PLoS One 11(7):e0158519
Régnière J, St-Amant R, Duval P (2012) Predicting insect distributions under climate change from physiological responses: spruce budworm as an example. Biol Invasions 14:1571–1586
Salman HR, Hellrigl K, Minerbi S, Battisti A (2016) Prolonged pupal diapause drives population dynamics of the pine processionary moth (Thaumetopoea pityocampa) in an outbreak expansion area. For Ecol Manage 361:375–381
Saulnier M, Roques A, Guibal F, Rozenberg P, Saracco G, Corona C, Edouard J-L (2017) Spatiotemporal heterogeneity of larch budmoth outbreaks in the French Alps over the last 500 years. Can J For Res 47:667–680
Schebeck M, Hansen E, Schopf A, Gregory R, Stauffer C, Bentz B (2017) Diapause and overwintering of two spruce bark beetle species. Physiol Entomol 42:200–210
Schneider L, Comte V, Rebetez M (2021) Increasingly favourable winter temperature conditions for major crop and forest insect pest species in Switzerland. Agric for Meteorol 298–299:108315
Slippers B, Hurley BP, Wingfield MJ (2015) Sirex woodwasp: A model for evolving management paradigms of invasive forest pests. Annu Rev Entomol 60:601–619
Tamburini G, Marini L, Hellrigl K, Salvadori C, Battisti A (2013) Effects of climate and density-dependent factors on population dynamics of the pine processionary moth in the Southern Alps. Clim Change 121:701–712
Tenow O (1996) Hazards to a mountain birch forest—Abisko in perspective. Ecol Bull 45:104–114
Tobin PC, Gray DR, Liebhold AM (2014) Supraoptimal temperatures influence the range dynamics of a non-native insect. Divers Distrib 20:813–823
Toïgo M, Barraquand F, Barnagaud J-Y, Piou D, Jactel H (2017) Geographical variation in climatic drivers of the pine processionary moth population dynamics. For Ecol Manage 404:141–155
Toïgo M, Nicolas M, Jonard M, Croisé L, Nageleisen LM, Jactel H (2020) Temporal trends in tree defoliation and response to multiple biotic and abiotic stresses. For Ecol Manage 477:118476
Ungerer MJ, Ayres MP, Lombardero MJ (1999) Climate and the northern distribution limits of Dendroctonus frontalis Zimmermann (Coleoptera: Scolytidae). J Biogeogr 26:1133–1145
Visser ME, Both C (2006) Shifts in phenology due to global climate change: the need for a yardstick. Proceedings of the royal society of London series B—Biol Sci 272:2561–2569
Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin JM, Hoegh-Guldberg O, Bairlein F (2002) Ecological responses to recent climate change. Nature 416:389–395
Wermelinger B (2004) Ecology and management of the spruce bark beetle, Ips typographus—a review of recent research. For Ecol Manage 202:67–82
White TCR (1974) Hypothesis to explain outbreaks of looper caterpillars, with special reference to population of Selidosema suavis in a plantation of Pinus radiata in New-Zealand. Oecologia 16:279–301
Acknowledgements
Many thanks to the Editors Jeremy D. Allison and Tim Paine, and to Barbara Bentz, Sandy Liebhold, and Deepa Pureswaran who kindly commented an earlier version of the chapter. The work benefited of a grant of Swedish University of Agricultural Sciences (SLU) and the European Union’s Horizon 2020 Program for Research and Innovation ‘HOMED’ (grant no. 771271).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2023 The Author(s)
About this chapter

Cite this chapter
Battisti, A., Larsson, S. (2023). Climate Change and Forest Insect Pests. In: D. Allison, J., Paine, T.D., Slippers, B., Wingfield, M.J. (eds) Forest Entomology and Pathology. Springer, Cham. https://doi.org/10.1007/978-3-031-11553-0_22
Download citation
DOI: https://doi.org/10.1007/978-3-031-11553-0_22
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-11552-3
Online ISBN: 978-3-031-11553-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)