
Abstract
The plant microbiota fulfils various crucial functions related to host health, fitness, and productivity. Over the past years, the number of plant microbiome studies continued to steadily increase. Technological advancements not only allow us to produce constantly increasing datasets, but also to extract more information from them in order to advance our understanding of plant-microbe interactions. The growing knowledge base has an enormous potential to improve microbiome-based, sustainable agricultural practices, which are currently poorly understood and have yet to be further developed. Cereal plants are staple foods for a large proportion of the world’s population and are therefore often implemented in microbiome studies. In the present review, we conducted extensive literature research to reflect the current state of knowledge in terms of the microbiome of the four most commonly cultivated cereal plants. We found that currently the majority of available studies are targeting the wheat microbiome, which is closely followed by studies on maize and rice. There is a substantial gap, in terms of published studies, addressing the barley microbiome. Overall, the focus of most microbiome studies on cereal plants is on the below-ground microbial communities, and there is more research on bacteria than on fungi and archaea. A meta-analysis conducted in the frame of this review highlights microbiome similarities across different cereal plants. Our review also provides an outlook on how the plant microbiota could be harnessed to improve sustainability of cereal crop production.
Similar content being viewed by others

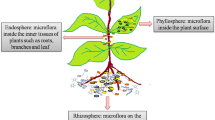
Introduction
It is assumed that the plant microbiome harbours a similar potential to improve agriculture as it happened during the Green Revolution of the 60s [1]. Microbiomes not only consist of bacteria and fungi, but also include archaea, protists as well as viruses. The whole assemblage of living organisms, this excludes viruses, is termed as the “microbiota” while their genetic elements, molecular building blocks, signalling molecules, and other constituents are known as the “microbiome” as per the latest definition [2]. The emergence of high-throughput sequencing and other meta-omics technologies facilitate the assessment of microbial community compositions and functions as well as the identification of the influencing factors. Over the past years, it was shown that the plant microbiome has certain specificities that makes it clearly distinguishable from microbial assemblages that are connected to other life forms. For example, the plant rhizosphere (a term that describes the surroundings of roots that are influenced by plant exudates) provides the main environment for microbial colonization [3]. The rhizosphere is known to be highly dynamic, enriched by microbes from local soil, and subject to specific shifts associated with various plant growth stages [4]. Moreover, its colonization is highly influenced and controlled by the composition of host-specific exudates which modify the physio-chemical properties of the soil and thus the microbiome [5]. Although the role of the rhizosphere in plant health, fitness, resilience, and productivity has been known for decades, it has increasingly been recognized in recent years that the comparatively less abundant microorganisms colonizing aboveground plant tissues also have certain implications for host resilience and pathogen defence [6, 7]. Especially endophytes, which can occur inside all plant tissues, are known to be involved in a multitude of interactions with their hosts [8, 9]. There are a number of mechanisms that were identified by which microorganisms can influence plant health, stress tolerance, and productivity [10]. Bacteria can metabolize nutrients to make them available to plants (e.g. nitrogen-fixation, phosphate solubilization and siderophore production to facilitate iron uptake), induce tolerance to abiotic and biotic stress (e.g. ACC deaminase activity) or produce phytohormones (e.g. auxins) with influence on plant development [11, 12].
In the 2020 crop year, 3 trillion tons cereals were produced worldwide, whereas it is estimated that wheat, maize, and rice cultivation account for almost 90% of it [13]. In 2011, cereals accounted directly for more than 50% of the worldwide daily caloric intake, while in addition a large proportion of the grain production has an indirect influence on human nutrition through its use as livestock feed [14]. The growing world population, which is projected to be around 9.7 billion by 2050 [15], has to face the challenge of increased agricultural productivity and yield, without wasting more land and under increasingly difficult and changing climatic conditions [16]. One way to achieve these goals could be to harness intrinsic functions of the plant microbiome and its potential to promote plant growth and positively influence crop tolerance to abiotic and biotic stressors [17]. However, the integration of beneficial microbes on a large scale in modern agriculture requires a deep understanding of the underlying plant-microbe-environment interactions [18, 19].
In this review, we focus on the microbiome of the four most commonly cultivated cereal plants: wheat, maize, rice and barley [13]. Many other cereals such as sorghum, rye, triticale, oats and millets as well as the so-called pseudocereals including amaranth, quinoa, buckwheat and various others are less commonly grown; they were not considered in this review. It is also noteworthy to mention that most of these plant species have several varieties and a scarcely assessable number of cultivars [20, 21]. In order to summarize current knowledge related to the microbiome of cereal plants, we performed an extensive literature search with the main databases PubMed and Google Scholar in the period of 02 February 2021 until 13 July 2021. The keywords “wheat”, “Triticum”, “maize”, “corn”, “Zea”, “rice”, “Oryza”, “barley”, “Hordeum”, and “cereals” were combined with either “microbiome” or “microbiota”. Several additional articles were found by searching for reviews and references that were cited within the articles that were found during the initial search. Studies not based on high-throughput sequencing and reviews were excluded to achieve greater consistency. A total of 302 articles were selected, all of which were published between 2013 and 2021. A table (Supplementary Table 1) with the microorganisms (bacteria, fungi and/or archaea) discussed in the articles, the plant compartment (soil, rhizosphere, roots, aboveground plant compartments, and specifically seeds) as well as the general topic of all included research articles was compiled during the preparation of the review. One point was given for every subject matter of the table that was addressed in the articles; for example, if a research article contained data on the microbiome of both wheat and maize, each subject matter received one point. Many studies contained more than one study matter, thus dividing through the total amount of publications would result in percentage numbers higher than 100%. We therefore decided to divide through the total number of entries in the compiled table. The same strategy was also applied for plant microhabitats and microorganism within the domains bacteria, fungi, and archaea. However, for simplicity, we always refer to “articles” and not the number of entries throughout the review. A general overview is provided in Table 1, while detailed information can be found in Supplementary Table 1. Overall, this review is meant to provide a comprehensive overview of the current knowledge base and to highlight key findings of the last years that were inferred from microbiome studies of cereal plants.
Recent findings related to the wheat microbiome
Wheat (plants from the genus Triticum L.) globally accounts for the largest cultivation area of all cereal plants. In the last 20 years, the area under cultivation for wheat has remained constant, while the yield steadily increased. Wheat is mainly cultivated in the European Union, China, and India, accounting for over 50% of the global wheat production [13].
From the 302 assessed articles, the largest fraction (32.4%; Fig. 1) included data related to the wheat microbiome. The majority of these articles focused on the below-ground microbiome, including soil, rhizosphere, and roots, accounting for 23.7%, 39.6% and 19.5%, respectively (Fig. 2). The aboveground microbiome was addressed in 17.2% of the studies assessed, with 10.7% addressing shoots, leaves and/or stems; 6.5% specifically addressed the plant’s seeds. Bacteria were more often in the focus of the research than fungi and archaea, with 65.7%, 30.1% and 4.2%, respectively (Fig. 3).

Cultivation areas of globally prevailing cereal crop plants and the number of corresponding microbiome studies. The cultivation areas are shown in the outer ring as percentages of the total area (622 million ha) used to cultivate the four crops [9]. The inner ring indicates the percentage of microbiome studies obtained by extensive literature search within publicly accessible databases

Proportions of studies addressing the microbiome of specific plant compartments. The belowground parts are most commonly addressed in the currently available studies for all major cereal plants. This is followed by the aboveground parts and studies that are specifically focused on seeds; except for barley for which more studies focus on the seeds than on other aboveground compartments

Proportions of microbiome studies targeting certain organism groups in microbiomes. Bacterial communities are most commonly addressed in the currently available studies for all major cereal plants. This is followed by fungi and archaea, which account for a substantially lower number of studies
The main research objectives related to the differences in the microbiome compared to other plant species, genotypes, plant compartments, and developmental stages (Table 1). It was shown that all these factors have a certain influence on the microbiome, although to varying degrees [34] showed that tillage and management type do not affect the richness, but the composition and structure of microbial communities in soil and plant roots. It was furthermore shown that agricultural intensification negatively influences the abundance of keystone fungal taxa and reduces the root fungal network connectivity [35]. On the other hand, Lupwayi et al.[36] found that neither tillage nor crop rotation had an effect on the alpha diversity and the relative abundance of the rhizobacterial community. Studies that focused on the plant genotype indicated that the microbiome of domesticated wheat plants is different in comparison to their ancestors and wild varieties [37, 38]. The authors provided evidence that they follow distinct community assembly strategies and that wild plants harbour more structured and more defined communities, yet they are less diverse compared to domesticated cultivars [39, 40]. In terms of agrochemical treatments, a high degree of specificity in terms of the applied substances was observed. Schlatter et al.[41, 42] showed that glyphosate had only limited effects on bacterial and fungal communities, while Qu et al.[43] observed that the herbicide S-metolachlor substantially influenced the microbial richness in the rhizosphere. Interestingly, they found that the proportion of distinct, beneficial bacteria can increase upon treatments, proposing a plant-regulated mechanism to cope with herbicide stress. In addition to the above-mentioned studies, there are also several studies that focused on the influence of pathogens, biocontrol agents, as well as different biostimuli on the microbiome. Seybold et al.[44] identified a correlation between changes in the wheat leaf microbiome and the suppression of immune-related metabolites by the pathogen Zymoseptoria tritici. Chen et al.[45] showed that the wheat-associated bacterium Pseudomonas piscium secrets a compound which suppresses growth and virulence of the fungal pathogen Fusarium graminearum by targeting a histone acetyltransferase and thereby dysregulating histone acetylation. Overall, several trends in wheat microbiome studies have become evident in recent years. Much emphasis was placed on the general assessment of the microbiome and to identify factors influencing microbial community compositions.
Microbiome-related discoveries in maize
Maize (plants from the genus Zea L.) globally accounts for the second largest cultivation area, with 0.9 hectares (ha) maize crop area per 1 ha wheat crop area. The crop area has increased constantly by 1.4 times over the last 20 years, while the harvest volume has doubled [13]. The by far leading corn-producing country is the US. It produced 31% of the world’s harvest in the harvest year 2019 and up to 91% of the grown plants were genetically engineered [13, 46]. About a third of the corn production in the US is used for feeding livestock, another third serves as main source for production of fuel ethanol. The rest is exported and used for human food and beverages as well as industrial applications, like packing material or insulin [47]. In comparison to wheat, rice and barley, maize is a C4-plant; these plants avoid photorespiration and have a higher water use efficiency. These host characteristics may be involved in sha** their microbiota.
The second largest proportion of the studied articles (31.8%; Fig. 1) included data on the maize microbiome; this is only 1.85% less than for wheat. The majority, 88.8%, of the research articles focused on the belowground microbiome, including soil, rhizosphere and roots, accounting for 26.9%, 43.8% and 18.1%, respectively (Fig. 2). The above-ground microbiome was addressed in 11.3% of the research articles, while 7.5% and 3.8% focused on the shoots and kernels, respectively (Fig. 2). Bacteria were again the focus of research (64.9%; Fig. 3), at the expense of research on fungi, which were covered in only 27.7% of the articles.
The main research objectives also related to differences in the microbiome compared to other plant species, genotypes, compartments or developmental stages. In accordance to wheat, it was shown that all these factors can have impacts on the microbiome[51] showed that BXs influence herbivore defence of the next generation of plants by changing the root and soil microbiome. There are two studies that highlight Trichoderma (T. harzianum and T. asperellum) as potential biocontrol agent (BCA) to control stalk rot caused by Fusarium graminearum. Both studies reveal differences in the microbiome community and disease reduction upon application of the BCA [52, 53]. In several other studies, it was shown that agricultural practices highly influence the maize microbiome [54, 55]. Schmidt et al.[56] found that recruitment of microorganisms differs between agricultural management types and that bacteria and fungi respond differently to the management type. They showed that microbial communities in the soil and rhizosphere respond differently to management strategies and propose that roots should be considered as a crucial factor influencing management outcomes. Ares et al.[55] compared two different maize genotypes (SinPre and Pigarro) under conventional and organic management and found that higher microbial diversity was associated with organic farming, as was the presence of AMF. Furthermore, it was shown that crop rotation had a bigger influence on the fungal soil community, than on the bacterial [54, 57]. Overall, recent studies found that the maize and wheat microbiome had many characteristics in common.
Summary of ongoing rice microbiome research
Rice (plants from the genus Oryza L.) globally accounts for the third largest cultivation area, with 0.75 ha rice crop per 1 ha wheat crop area. China and India are the main rice producers, accounting for over 50% of the global rice production in the harvest year 2019 [13].
From the 302 studied research articles, the third largest fraction (28.6%; Fig. 1) included data related to the rice microbiome. The majority, with 79.6%, of the assessed studies, focused on the belowground microbiome, with 24.1%, 31.4%, and 24.1% on the soil, rhizosphere, and roots, respectively. The aboveground microbiome was the focus of research in 22.4% of the articles, with 8% addressing seeds and 12.4% shoots (Fig. 2). When the collected literature was assessed, we noticed that the rhizosphere microbiome was less frequently (31.4%) explored compared to wheat and maize (39.6% and 43.8%, respectively) and that more emphasis was put on the root microbiome (24.1%). With 73.2% of the articles, bacteria were again studied more frequently than fungi with 18.7% and archaea with 8.1% (Fig. 3).
The main research objective addressed was targeting differences in the microbiome, as also observed for wheat and maize. Other frequent objectives included comparisons to other plant species, genotypes, developmental stages, or plant compartments (Table 1). As with wheat, the influence of agricultural practices on the microbiome was the second most addressed topic. A major focus was on the effects of different irrigation methods on the below-ground microbiome [58,59,60,61,62]. Compared to research on wheat and maize, there is less research on the impact of fertilizer use on the rice microbiome and more on the influence of soil pollution with pesticides and heavy metals, like methylmercury, arsenic, or antimony. Liu et al. [63] showed that the soil microbial community was able to quickly adapt to exposure to the three commonly used pesticides butachlor, clothianidin and tricyclazole. They found only minor changes in community composition and diversity; the differences even decreased over the growth period. Moreover, Chen et al. [64] and Qian et al. [65] showed that root exudation in rice plants increased upon exposure to the herbicide diclofop-methyl, leading to increased bacterial biomass, richness, and diversity. They hypothesized that it was possibly due to a protection mechanism of the rice plant as a response to the herbicide treatment. Moreover, it was shown several times that the rhizosphere and soil microbiome respond to heavy metal contamination. Recently, Das et al. [66] proposed fertilization with silicate as a possible mechanism to improve bacterial stress tolerance in arsenic-polluted soils. Microorganisms can substantially influence the solubility of pollutants and bio-transform (e.g. methylation, oxidation and reduction) them. Interestingly, this process has been shown to be strongly influenced by water management in rice cultivation, probably due to the different redox potentials of aerobic and anaerobic soils [59, 67, 68]. Water management in general has been shown to significantly affect the rice microbiome, and distinct bacterial communities were found in flooded and non-flooded rice fields [60]. A distinct shift in microbial communities towards more consistent compositions was also observed in extensive rice monocultures. Interestingly, it was demonstrated that this is only partly due to agricultural management, but that there is also a strong influence of the plants themselves, leading to an enrichment of distinct taxa in rice fields, e.g. methanogenic archaea [69]. In terms of rice seeds, most of the studies focused on the description of microbial communities in different host genotypes. Recently, however, a transgenerational mechanism via the seed microbiota to confer resistance against Burkholderia plantarii was discovered. The seed-endophytic bacterium Sphingomonas melonis was identified as a key player in the tripartite interaction. It confers disease resistance to a broad range of rice genotypes by producing anthranilic acid which impairs the virulence factor biosynthesis of B. plantarii [8].
Recent discoveries in the barley microbiome
Barley (plants form the genus Hordeum L.) globally accounts for the fourth largest cultivation area, with 0.23 ha barley crop per 1 ha wheat crop area. The European Union is the leading barley-growing region, accounting for over 40% of the global barley production [13]. From the 302 assessed articles, the by far smallest fraction (7.2%; Fig. 1) included data related to the barley microbiome. As with the other cereal plants, the majority of the articles focused on the below-ground microbiome (78.3%; Fig. 2). The aboveground microbiome was addressed in 21.6% of the studies, with a specific focus on the plant’s seed microbiome (16.2%, Fig. 2). Most of the studies focused on bacteria (74.1%, Fig. 3), and only to a lesser extent on fungi (25.9%; Fig. 3), while no studies were found that addressed archaea.
The vast majority of the assessed publications presented data related to differences in the microbiome compared to other plant species, genotypes, plant compartments and developmental stages (Table 1). Bulgarelli et al. [70] showed that the barley genotype has a rather small but significant impact on the root microbiome and that the community composition is influenced by the combined effect of microbe-microbe and plant-microbe interactions. Yang et al. [71] found that plants grown in autoclaved soil, which simulated a disturbed microbiome, were severely affected by additional drought stress. The reduction in biomass was connected with major changes in the microbial community composition, which indicated a shift in plant colonization from soil-derived bacteria to seed-originated endophytes. Moreover, plants grown in field soil showed a significantly higher potential to be resistant against Blumeria graminis and lower infection rates compared to plants grown in potting soil. The authors hypothesized that this was due to the fact that field soil harbours higher microbial diversity which allows enrichment of beneficial bacteria [72]. Rahman et al. [73] found that the barley seed microbiome harbours several beneficial strains and inoculation with them promoted, not only plant growth, but also resistance against Blumeria graminis. The barley seed microbiome is, compared to the other cereals, more in the focus of research. Abdullaeva et al. showed that cultivated barley (as well as wheat) has a more diverse seed microbiome, including bacterial taxa linked to the human microbiome, than its wild ancestor Hordeum spontaneum, yet it is less connected. Furthermore, they could detect indications for co-evolution between the plant and their microbiome during the domestication process [40].
Common features of cereal crop microbiomes
The proportion of microbiome studies on the four cereal crops corresponds relatively closely to the proportion of their cultivated area, with only rice being slightly overrepresented and barley being slightly underrepresented (Fig. 1). Although the microbiome of each of the cereal crops discussed here has its own characteristics, certain commonalities among the hosts have been identified. It is particularly noteworthy that in all four crops, the belowground microbiome and bacterial communities have been the focus of research in recent years. All four cereal plants had in common that they harboured Proteobacteria, Actinobacteria, Firmicutes, Bacteriodetes, Acidobacteria, and Chloroflexi in their microbiome as prevalent bacterial phyla. Reviews by Kavamura et al. [74] and Mehta et al. [75] already presented the most commonly detected bacterial genera in the wheat and maize microbiome, respectively. Pantoea, Pseudomonas, Rhizobium, Sphingomonas, and Stenotrophomonas constitute some of the bacterial genera that are commonly found in both wheat and maize. When the literature related to rice and barley was assessed, we found that these bacterial genera also commonly occurred in their microbiome.
Many of the conducted studies led to the identification of beneficial bacteria and biostimulants which refers to microorganisms with plant growth-promoting, disease-supressing, and/or other fitness-enhancing traits [76]. Interestingly, the majority of the identified strains was assigned to the bacterial phyla Proteobacteria and Firmicutes and the fungal phyla Ascomycota, more specifically the genus Trichoderma. Members of the genera Pseudomonas and Bacillus/Paenibacillus were often identified as plant-beneficial bacteria. For example, Pseudomonas stutzeri inoculation increased plant development and had a positive impact on bacterial community composition, particularly among diazotrophs and ammonia-oxidizers [77]. Similar results were obtained by Li et al. [78], showing that inoculation with Paenibacillus triticisoli led to increased dry maize biomass and a shift in the microbial community, especially in a low nitrogen environment. They both demonstrated that biostimulants not only promote plant growth by the strain’s specific characteristics, like nitrogen fixation, but can also modulate the microbiome.
Recent advances in microbiome-related techniques allow us to analyse microbial communities in detail, estimate their complexity, and study their interplay with host plants; however, most of the studies conducted to date have dealt exclusively with bacterial communities [79, 80]. In the future, increasing the proportion of research on fungi, archaea, and protists in cereal crops will be crucial to fully understand the relationship between plants, microbes, and the environment. So far, studies targeting fungal communities have shown that cereals are dominated by Ascomycota and to a lesser extent by Basidiomycota [45, 71, 73,74,75, 81,82,83,84,85,86,87,88,89,90,91]. Archaea were found to be the focus of the lowest proportion of the reviewed studies. Furthermore, archaea are only considered in metagenomic /-transcriptomic studies (38%) or included in 16S rRNA gene analyses conducted with bacteria-specific primers (38%). Only a few studies are based on archaea-specific primers (23%), which may lead to an incomplete picture of the archaeal community composition. Rice cultivation is the cause of 1.3% of the global greenhouse gas emission, because its cultivation in flooded paddies results in anaerobic soil conditions, which provide optimal growing conditions for methanogenic archaea [92]. For this reason, a major fraction of research on archaea is done in rice paddies and to a lesser extend in maize and wheat. In wheat the most commonly detected archaeal phylum was Nitrososphaerota and to a lesser extend Euryarchaeota [27, 93, 94]. However, in rice, the phyla Euryarchaeota and Crenarchaeota, including the methanogenic archaea, were detected most commonly [95, 96]. There is a substantial variation in archaeal community compositions between the studies, which may have different reasons [62, 85, 95, 96]. For one, only a low number of studies is available and in addition they focus on different compartments, treatments, and other factors. Furthermore, the use of different primers to assess the community may lead to substantial variations.
A meta-analysis conducted in the frame of this review indicates that there is no significant difference in microbial diversity between the four plant types in any compartment (Fig. 4). The utilized data were extracted from 160 manuscripts depicting alpha diversity and the methods are further described in the Supplementary Information. Certain tendencies were observed in the meta-analysis, yet there is substantial variability across the studies. This observed variability could be due to several reasons, e.g. different primer sets used to amplify marker genes, differences in sequencing platforms, non-standardized protocols to extract total community DNA, etc. Furthermore, there are substantial differences in relation to the sampling process, especially concerning (bulk) soil. We noticed that the definition of bulk soil is very broad. It ranges from soil collected from neighbouring fields to soil only loosely adhering to the plant. Soil samples not sampled in the field at least during cultivation, or from pots not planted were excluded in our meta-analysis for better consistency. A standardized protocol as proposed by Barillot et al. [97] could help overcoming these variations. In order to be able to clearly differentiate between bulk-, rhizosphere-, and rhizoplan soil fractions they suggest a 3-step protocol, starting with vigorously shaking the roots to remove loosely bound soil considered as bulk soil. Subsequently, the rhizosphere is sampled by shaking roots in a NaCl solution to catch the soil directly adhering to the roots. Finally, roots are washed and again shaken in a solution containing NaCl and Tween to collect the rhizoplane fraction. However, it will require host-specific adaptations because this protocol was established for small, herbaceous plants and may not be directly applied to crop plants grown under different environmental and soil conditions.

Assessment of bacterial (A-E) and fungal (F-J) diversity in the different compartments of cereals. The diversity numbers were extracted as Chao1 index or ASV richness from the manuscript text or plots. The diversity in each compartment is not significantly different between the cereals
There were no observable differences in microbial diversity of rhizosphere and soil samples, but the soil compartments were substantially higher compared to endosphere samples. The endorhiza was generally more diverse than endophytes found in the aboveground parts of the host plants (Fig. 4). It is important to highlight that not all samples from the endosphere were surface-sterilized. Meta-analyses of fungal communities showed similar results. The diversity in the soil compartments was much higher than in roots, leaves and seeds (Fig. 4). Interestingly, the intensely bred cereal plants maintained the evolutionary old trait to form arbuscular mycorrhizal symbioses, which offers a great potential for enhancing yield without using synthetic fertilizers [98]. However, due to the fact that less data is available for fungi than for bacteria, the analysis might be biased to a certain degree.
The overall microbiome-related objectives addressed in the assessed studies were similar for all four plants. The most commonly addressed topic was the general and comparative assessment of their microbiome, followed by agronomic practices (e.g. tillage, organic vs. conventional management, crop rotation). Differences were observed in terms of the number of studies focusing on fertilization management, which was more often in the focus for maize studies compared to the other cereals. Moreover, effects of soil contaminations were mostly addressed in rice microbiome studies. Overall, it is clear that there are many factors that influence the composition of the cereal plant microbiome; the major studied factors are depicted in Fig. 5. Interestingly, the assessed studies had only a minor focus (4.1% on average) on the impact of abiotic stress (e.g. water stress) on the microbiome and how the microbiota could be harnessed to protect host plants under these conditions. In addition, the influence of plant pathogens on the microbiome was comparatively less discussed (4% on average) in the reviewed literature. Several studies addressed the overall change in the microbiome composition when plants are confronted with pathogens [44, 45, 72, 99]. Bacterial communities, including such that promote plant growth, showed a tendency to be more diverse when a pathogen was present [99,100,101,102]. It is generally known that moderate disruptions can cause diversity increases in various ecosystems, however, lasting disturbances by plant pathogens (and abiotic stress) are mostly linked to adverse effects [103].

The plant microbiome is influenced by various factors. These influencing factors can be intrinsic (e.g. genotype, compartment and developmental stage) or external factors, including agricultural management or abiotic and biotic stress
It should be highlighted that many of the described commonalities between the four addressed crop plants are also present in various other plant species, therefore they are not only distinct to cereal crops. Moreover, it can be expected that a growing number of available datasets combined with big data analyses will allow us to better understand the underlying factors of similarities and differences in the future.
The potential of the cereal crop microbiome for future applications
The current state of knowledge indicates that functions provided by distinct microbial communities associated with plants can be harnessed to reduce the use of agrochemicals and fertilizers [104]. This is especially important for cereal crops, because they not only globally account for the largest cultivation areas, but also provide staple food for over half of the world’s population. Agriculture is an important factor in achieving climate goals of the European Union and beyond, as it has been identified as one of the main causes of climate change. The recently presented Farm to Fork Strategy as part of the European Green Deal has the ambitious goals to halve the use of pesticides, reduce overfertilization and triple the area of sustainable agriculture for a fair and environmentally friendly food system as well as to counteract the immense biodiversity loss and depletion of natural resources [105]. It is estimated that pre-harvest grain loss due to biotic factors, like pests, pathogens and weeds, or abiotic stress, can account for up to 35% of the total harvest. Another 20% is globally lost during storage and affected by growth of mycotoxigenic fungi which can also lead to a decline in grain quality [106]. Solanki et al. [107, 108] showed that traditional methods to eliminate insects before storage, like seed fumigation, have a non-target effect on the microbiome. These applications decrease bacterial diversity over time. Furthermore, they isolated several bacterial and fungal strains from the wheat seed microbiome with the potential to reduce fungal and mycotoxin contamination. The plant microbiome will likely play an important role in providing solutions for sustainable agricultural practices; previous research has demonstrated that various members of the indigenous microbiome of cereal plants can be used to increase yields while simultaneously reducing emissions as well as the use of agrochemicals and fertilizers. During the last years, several microbial strains were identified in cereals that might be suitable to substitute pesticides to combat major plant diseases. It was shown that Pseudomonas piscium and Pantoea agglomerans can be used against Fusarium head blight in wheat, Sphingomonas melonis for rice seedling blight, and Enterobacter cloacae to fight stalk and ear rot in maize [8, 45, 72, 116]. The application of microbial inoculants is highly complex and their successful introduction depends on several factors, including the specific strain, application mode and time as well as the host microbiome itself [117]. Therefore, further research will be required to identify inoculants that are compatible with the host plant as well as its microbiome under distinct, and often adverse, conditions. Although research on wheat, maize, and rice is already tremendous compared to other plants, especially concerning the general assessment of the microbiome, there is still a lack of information on the functional aspects of the microbiome and how it can be specifically manipulated and engineered to improve plant productivity and health. Furthermore, as previously highlighted, cereal microbiomes share many common characteristics, suggesting that many findings obtained with a particular plant species may be transferable to others.
Propositions for further consideration
-
1.
A large number of studies addressing microbiomes of cereal crops were conducted in recent years; however, they are mainly based on the analysis of a specific plant compartment. More research based on holistic approaches covering the whole plant will be essential to improve our understanding of microbiome assembly and dynamics.
-
2.
The majority of available studies focuses on belowground plant compartments. Aboveground compartments, and especially seeds, are currently understudied. Recent research has indicated that seeds might harbour highly effective biostimulants, plant growth promoters, and biocontrol strains [118].
-
3.
Climate change with more frequently occurring weather extremes like drought, heavy rainfalls, and in more general, conditions allowing pathogens to proliferate, highlights the importance of research on the response of the plant microbiome to abiotic and biotic stress. This will allow to identify key players to increase plant resistance to different stress factors. In addition, more studies on the effects of combined stresses, which are characteristic for the Anthropocene, on the plant microbiome and plant performance are necessary.
-
4.
Many agrochemicals were shown to have off-target effects on naturally occurring microbial populations; this also includes antimicrobial resistance formation. Most of the current studies focus on resistance formation in target organisms, but ignore the effect on the overall resistome. Future studies should subject agrochemicals to deepening analyses which includes the evaluation of representative plant resistomes.
-
5.
Most of the ongoing microbiome research is focused on bacteria, followed by fungi. The other constituents of the microbiota, including archaea, protists, and algae, are almost completely ignored by research. More holistic microbiome assessments should be conducted in the future.
-
6.
Viruses, phages, plasmids, free (relic) DNA and various mobile genetic elements also belong to the plant microbiome; however, they are completely understudied despite indications of their importance. More studies on their implications for the assembly and functioning of the microbiome are required.
-
7.
Most of the studies are based on amplicon sequencing of 16S (or 18S) rRNA gene fragments and the ITS region. However, such approaches only allow assessments of the community composition. More research on functions and activities occurring within the plant microbiome will be crucial for its deepening exploration. This requires more poly-phasic and inter-linked experimental and methodological approaches considering spatial and temporal scales. For studies targeting the plant microbiome, we especially suggest the combination of metagenomics/metatranscriptomics and cultivation-based approaches. This will allow an assessment of modes of (inter-)action.
-
8.
Microbiome research provides a knowledge base that has an enormous potential to improve agricultural practices and develop microbiome-based fertilizers and pesticides. The plant microbiome is inter-connected with other organisms and ecosystems; this has to be considered especially for health issues. The development of solutions to restore and save microbial diversity for ecosystem functioning as well as the closely connected planetary health should be a central aim of microbiome researchers in the future [103].
Data Availability
Not applicable.
References
Qiu Z, Egidi E, Liu H, Kaur S, Singh BK. New frontiers in agriculture productivity: optimised microbial inoculants and in situ microbiome engineering. Biotechnol Adv. 2019;37:107371.
Berg G, Rybakova D, Fischer D, Cernava T, Vergès M-CC, Charles T, et al. Microbiome definition re-visited: old concepts and new challenges. Microbiome. 2020;8:1–22.
Berendsen RL, Pieterse CMJ, Bakker PA. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012;17:478–86.
Chaparro JM, Badri DV, Vivanco JM. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014;8:790–803.
Zhalnina K, Louie KB, Hao Z, Mansoori N, Da Rocha UN, Shi S, et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat Microbiol. 2018;3:470–80.
Li P-D, Zhu Z-R, Zhang Y, Xu J, Wang H, Wang Z, Li H. The phyllosphere microbiome shifts toward combating melanose pathogen. Microbiome. 2022;10:56. https://doi.org/10.1186/s40168-022-01234-x.
Chen T, Nomura K, Wang X, Sohrabi R, Xu J, Yao L, et al. A plant genetic network for preventing dysbiosis in the phyllosphere. Nature. 2020;580:653–7.
Matsumoto H, Fan X, Wang Y, Kusstatscher P, Duan J, Wu S, et al. Bacterial seed endophyte shapes disease resistance in rice. Nat Plants. 2021;7:60–72.
Hardoim PR, van Overbeek LS, van Elsas JD. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008;16:463–71.
Arif I, Batool M, Schenk PM. Plant microbiome engineering: expected benefits for improved crop growth and resilience. Trends Biotechnol. 2020;38:1385–96.
Di Fan, Smith DL. Characterization of selected plant growth-promoting rhizobacteria and their non-host growth promotion effects. Microbiol Spectr. 2021;9:e00279–21.
Del Carmen Orozco-Mosqueda M, Glick BR, Santoyo G. ACC deaminase in plant growth-promoting bacteria (PGPB): an efficient mechanism to counter salt stress in crops. Microbiol Res. 2020;235:126439.
Food and Agriculture Organization of the United Nations (FAO). FAOSTAT Database. https://www.fao.org/faostat/en/#data. Accessed 14 Oct 2021.
Awika JM. Major Cereal Grains Production and Use around the World. Advances in Cereal Science: Implications to Food Processing and Health Promotion:American Chemical Society; 2011.pp. 1–13. doi:https://doi.org/10.1021/bk-2011-1089.ch001.
United Nations. World population prospects 2019: highlights. Population Division: Department of Economic and Social Affairs; 2019.
Godfray HCJ, Beddington JR, Crute IR, Haddad L, Lawrence D, Muir JF, et al. Food security: the challenge of feeding 9 billion people. Science. 2010;327:812–8.
Ali S, Tyagi A, Mushtaq M, Al-Mahmoudi H, Bae H. Harnessing plant microbiome for mitigating arsenic toxicity in sustainable agriculture.Environmental Pollution. 2022:118940.
Zhang J, Cook J, Nearing JT, Zhang J, Raudonis R, Glick BR, et al. Harnessing the plant microbiome to promote the growth of agricultural crops. Microbiol Res. 2021;245:126690.
Albright MBN, Louca S, Winkler DE, Feeser KL, Haig S-J, Whiteson KL, et al. Solutions in microbiome engineering: prioritizing barriers to organism establishment. ISME J. 2022;16:331–8.
Wrigley CW. An overview of the family of cereal grains prominent in world agriculture. Encyclopedia of Food Grains. 2016;1:73–85.
Fletcher RJ. Pseudocereals, Overview. Reference Module in Food Science:Elsevier; 2016. doi:https://doi.org/10.1016/B978-0-08-100596-5.00039-1.
**ong C, Zhu Y-G, Wang J-T, Singh B, Han L-L, Shen J-P, et al. Host selection shapes crop microbiome assembly and network complexity. New Phytol. 2021;229:1091–104. https://doi.org/10.1111/nph.16890.
Lu T, Ke M, Peijnenburg WJGM, Zhu Y, Zhang M, Sun L, et al. Investigation of Rhizospheric Microbial Communities in Wheat, Barley, and two Rice varieties at the Seedling Stage. J Agric Food Chem. 2018;66:2645–53. https://doi.org/10.1021/acs.jafc.7b06155.
Latz MAC, Kerrn MH, Sørensen H, Collinge DB, Jensen B, Brown JKM, et al. Succession of the fungal endophytic microbiome of wheat is dependent on tissue-specific interactions between host genotype and environment. Sci Total Environ. 2021;759:143804. https://doi.org/10.1016/j.scitotenv.2020.143804.
Cordero J, de Freitas JR, Germida JJ. Bacterial microbiome associated with the rhizosphere and root interior of crops in Saskatchewan, Canada. Can J Microbiol. 2020;66:71–85. https://doi.org/10.1139/cjm-2019-0330.
Chen S, Waghmode TR, Sun R, Kuramae EE, Hu C, Liu B. Root-associated microbiomes of wheat under the combined effect of plant development and nitrogen fertilization. Microbiome. 2019;7:136. https://doi.org/10.1186/s40168-019-0750-2.
Fan K, Weisenhorn P, Gilbert JA, Chu H. Wheat rhizosphere harbors a less complex and more stable microbial co-occurrence pattern than bulk soil. Soil Biol Biochem. 2018;125:251–60.
Fan K, Weisenhorn P, Gilbert JA, Shi Y, Bai Y, Chu H. Soil pH correlates with the co-occurrence and assemblage process of diazotrophic communities in rhizosphere and bulk soils of wheat fields. Soil Biol Biochem. 2018;121:185–92.
Kowalchuk GA, Buma DS, de Boer W, Klinkhamer PG, van Veen JA. Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms. Antonie Van Leeuwenhoek. 2002;81:509. https://doi.org/10.1023/A:1020565523615.
Berg G, Smalla K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol Ecol. 2009;68:1–13. https://doi.org/10.1111/j.1574-6941.2009.00654.x.
Wang J, Song Y, Ma T, Raza W, Li J, Howland JG, et al. Impacts of inorganic and organic fertilization treatments on bacterial and fungal communities in a paddy soil. Appl Soil Ecol. 2017;112:42–50. https://doi.org/10.1016/j.apsoil.2017.01.005.
Kavamura VN, Hayat R, Clark IM, Rossmann M, Mendes R, Hirsch PR, Mauchline TH. Inorganic Nitrogen Application affects both taxonomical and predicted functional structure of wheat Rhizosphere Bacterial Communities. Front Microbiol. 2018;9:1074. https://doi.org/10.3389/fmicb.2018.01074.
Ai C, Liang G, Sun J, Wang X, He P, Zhou W, He X. Reduced dependence of rhizosphere microbiome on plant-derived carbon in 32-year long-term inorganic and organic fertilized soils. Soil Biol Biochem. 2015;80:70–8. https://doi.org/10.1016/j.soilbio.2014.09.028.
Hartman K, van der Heijden MGA, Wittwer RA, Banerjee S, Walser J-C, Schlaeppi K. Crop** practices manipulate abundance patterns of root and soil microbiome members paving the way to smart farming. Microbiome. 2018;6:1–14.
Banerjee S, Walder F, Büchi L, Meyer M, Held AY, Gattinger A, et al. Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J. 2019;13:1722–36.
Lupwayi NZ, Fernandez MR, Kanashiro DA, Petri RM. Profiles of wheat rhizobacterial communities in response to repeated glyphosate applications, crop rotation, and tillage.Canadian Journal of Soil Science. 2020:1–11.
Spor A, Roucou A, Mounier A, Bru D, Breuil M-C, Fort F, et al. Domestication-driven changes in plant traits associated with changes in the assembly of the rhizosphere microbiota in tetraploid wheat. Sci Rep. 2020;10:1–12.
Kinnunen-Grubb M, Sapkota R, Vignola M, Nunes IM, Nicolaisen M. Breeding selection imposed a differential selective pressure on the wheat root-associated microbiome. FEMS Microbiol Ecol. 2020;96:fiaa196.
Hassani MA, Özkurt E, Franzenburg S, Stukenbrock EH. Ecological assembly processes of the bacterial and fungal microbiota of wild and domesticated wheat species. Phytobiomes J. 2020;4:217–24.
Abdullaeva Y, Manirajan BA, Honermeier B, Schnell S, Cardinale M. Domestication affects the composition, diversity, and co-occurrence of the cereal seed microbiota.Journal of Advanced Research. 2020.
Schlatter DC, Yin C, Burke I, Hulbert S, Paulitz T. Location, root proximity, and glyphosate-use history modulate the effects of glyphosate on fungal community networks of wheat. Microb Ecol. 2018;76:240–57.
Schlatter DC, Yin C, Hulbert S, Burke I, Paulitz T. Impacts of repeated glyphosate use on wheat-associated bacteria are small and depend on glyphosate use history.Applied and environmental microbiology. 2017;83.
Qu Q, Li Y, Zhang Z, Cui H, Zhao Q, Liu W, et al. Effects of S-metolachlor on wheat (Triticum aestivum L.) seedling root exudates and the rhizosphere microbiome. J Hazard Mater. 2021;411:125137.
Seybold H, Demetrowitsch TJ, Hassani MA, Szymczak S, Reim E, Haueisen J, et al. A fungal pathogen induces systemic susceptibility and systemic shifts in wheat metabolome and microbiome composition. Nat Commun. 2020;11:1–12.
Chen Y, Wang J, Yang N, Wen Z, Sun X, Chai Y, Ma Z. Wheat microbiome bacteria can reduce virulence of a plant pathogenic fungus by altering histone acetylation. Nat Commun. 2018;9:1–14.
USDA NASS, Acreage, Report. June 30, 2022. https://usda.library.cornell.edu/concern/publications/j098zb09z?locale=en. Accessed 18 Jul 2022.
McConnell, Outlook F. FDS-21 h. August 16, 2021. https://www.ers.usda.gov/webdocs/outlooks/101863/fds-21 h.pdf?v=4923.1. Accessed 14 Oct 2021.
Walters WA, ** Z, Youngblut N, Wallace JG, Sutter J, Zhang W, et al. Large-scale replicated field study of maize rhizosphere identifies heritable microbes. Proc Natl Acad Sci U S A. 2018;115:7368–73. https://doi.org/10.1073/pnas.1800918115.
Kudjordjie EN, Sapkota R, Steffensen SK, Fomsgaard IS, Nicolaisen M. Maize synthesized benzoxazinoids affect the host associated microbiome. Microbiome. 2019;7:59. https://doi.org/10.1186/s40168-019-0677-7.
Frey M, Schullehner K, Dick R, Fiesselmann A, Gierl A. Benzoxazinoid biosynthesis, a model for evolution of secondary metabolic pathways in plants. Phytochemistry. 2009;70:1645–51.
Hu L, Robert CAM, Cadot S, Zhang X, Ye M, Li B, et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by sha** the rhizosphere microbiota. Nat Commun. 2018;9:2738. https://doi.org/10.1038/s41467-018-05122-7.
Saravanakumar K, Li Y, Yu C, Wang Q, Wang M, Sun J, et al. Effect of Trichoderma harzianum on maize rhizosphere microbiome and biocontrol of Fusarium Stalk rot. Sci Rep. 2017;7:1–13.
He A, Sun J, Wang X, Zou L, Fu B, Chen J. Reprogrammed endophytic microbial community in maize stalk induced by Trichoderma asperellum biocontrol agent against Fusarium diseases and mycotoxin accumulation. Fungal biology. 2019;123:448–55.
Benitez M-S, Osborne SL, Lehman RM. Previous crop and rotation history effects on maize seedling health and associated rhizosphere microbiome. Sci Rep. 2017;7:1–13.
Ares A, Costa J, Joaquim C, Pintado D, Santos D, Messmer MM, Mendes-Moreira PM. Effect of Low-Input Organic and Conventional Farming Systems on Maize Rhizosphere in Two Portuguese Open-Pollinated Varieties (OPV),“Pigarro”(Improved Landrace) and “SinPre”(a Composite Cross Population).Front Microbiol. 2021;12.
Schmidt JE, Kent AD, Brisson VL, Gaudin ACM. Agricultural management and plant selection interactively affect rhizosphere microbial community structure and nitrogen cycling. Microbiome. 2019;7:1–18.
Mwakilili AD, Mwaikono KS, Herrera SL, Midega CAO, Magingo F, Alsanius B, et al. Long-term maize-Desmodium intercrop** shifts structure and composition of soil microbiome with stronger impact on fungal communities. Plant Soil. 2021;467:437–50.
Zhang Q, Chen H, Huang D, Xu C, Zhu H, Zhu Q. Water managements limit heavy metal accumulation in rice: dual effects of iron-plaque formation and microbial communities. Sci Total Environ. 2019;687:790–9.
Zecchin S, Corsini A, Martin M, Cavalca L. Influence of water management on the active root-associated microbiota involved in arsenic, iron, and sulfur cycles in rice paddies. Appl Microbiol Biotechnol. 2017;101:6725–38.
Cui H-L, Duan G-L, Zhang H, Cheng W, Zhu Y-G. Microbiota in non-flooded and flooded rice culms. FEMS Microbiol Ecol. 2019;95:fiz036.
Chialva M, Ghignone S, Cozzi P, Lazzari B, Bonfante P, Abbruscato P, Lumini E. Water management and phenology influence the root-associated rice field microbiota. FEMS Microbiol Ecol. 2020;96:fiaa146.
Breidenbach B, Brenzinger K, Brandt FB, Blaser MB, Conrad R. The effect of crop rotation between wetland rice and upland maize on the microbial communities associated with roots. Plant Soil. 2017;419:435–45.
Liu J, Song Y, Tang M, Lu Q, Zhong G. Enhanced dissipation of xenobiotic agrochemicals harnessing soil microbiome in the tillage-reduced rice-dominated agroecosystem. J Hazard Mater. 2020;398:122954.
Chen S, Li X, Lavoie M, ** Y, Xu J, Fu Z, Qian H. Diclofop-methyl affects microbial rhizosphere community and induces systemic acquired resistance in rice. J Environ Sci. 2017;51:352–60.
Qian H, Zhu Y, Chen S, ** Y, Lavoie M, Ke M, Fu Z. Interacting effect of diclofop-methyl on the rice rhizosphere microbiome and denitrification. Pestic Biochem Physiol. 2018;146:90–6.
Das S, Kim GW, Lee JG, Bhuiyan MSI, Kim PJ. Silicate fertilization improves microbial functional potentials for stress tolerance in arsenic-enriched rice crop** systems. J Hazard Mater. 2021;417:125953.
Kong T, Lin H, **ao E, **ao T, Gao P, Li B, et al. Investigation of the antimony fractions and indigenous microbiota in aerobic and anaerobic rice paddies. Sci Total Environ. 2021;771:145408.
Das S, Chou M-L, Jean J-S, Liu C-C, Yang H-J. Water management impacts on arsenic behavior and rhizosphere bacterial communities and activities in a rice agro-ecosystem. Sci Total Environ. 2016;542:642–52.
Edwards J, Santos-Medellín C, Nguyen B, Kilmer J, Liechty Z, Veliz E, et al. Soil domestication by rice cultivation results in plant-soil feedback through shifts in soil microbiota. Genome Biol. 2019;20:1–14.
Bulgarelli D, Garrido-Oter R, Münch PC, Weiman A, Dröge J, Pan Y, et al. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe. 2015;17:392–403.
Yang L, Schröder P, Vestergaard G, Schloter M, Radl V. Response of barley plants to drought might be associated with the recruiting of soil-borne endophytes. Microorganisms. 2020;8:1414.
Bziuk N, Maccario L, Douchkov D, Lueck S, Babin D, Sørensen SJ, et al. Tillage shapes the soil and rhizosphere microbiome of barley—but not its susceptibility towards Blumeria graminis f. sp. hordei. FEMS Microbiol Ecol. 2021;97:fiab018.
Rahman MM, Flory E, Koyro H-W, Abideen Z, Schikora A, Suarez C, et al. Consistent associations with beneficial bacteria in the seed endosphere of barley (Hordeum vulgare L). Syst Appl Microbiol. 2018;41:386–98.
Kavamura VN, Mendes R, Bargaz A, Mauchline TH. Defining the wheat microbiome: towards microbiome-facilitated crop production. Comput Struct Biotechnol J. 2021;19:1200.
Mehta S, Singh B, Patra A, Tripathi A, Easwaran M, Choudhary JR et al. Maize microbiome: current insights for the sustainable agriculture. In: Microbiomes and Plant Health: Elsevier; 2021. p. 267–297.
Du Jardin P. Plant biostimulants: definition, concept, main categories and regulation. Sci Hort. 2015;196:3–14.
Ke X, Feng S, Wang J, Lu W, Zhang W, Chen M, Lin M. Effect of inoculation with nitrogen-fixing bacterium Pseudomonas stutzeri A1501 on maize plant growth and the microbiome indigenous to the rhizosphere. Syst Appl Microbiol. 2019;42:248–60.
Li Y, Li Q, Chen S. Diazotroph Paenibacillus triticisoli BJ-18 drives the variation in bacterial, diazotrophic and fungal communities in the rhizosphere and root/shoot endosphere of maize. Int J Mol Sci. 2021;22:1460.
Afridi MS, Javed MA, Ali S, de Medeiros FHV, Ali B, Salam A et al. New opportunities in plant microbiome engineering for increasing agricultural sustainability under stressful conditions.Frontiers in Plant Science. 2022;13.
Glick BR, Gamalero E. Recent developments in the study of plant microbiomes. Microorganisms. 2021;9:1533.
Zhang J, Zhang C, Yang J, Zhang R, Gao J, Zhao X, et al. Insights into endophytic bacterial community structures of seeds among various Oryza sativa L. rice genotypes. J Plant Growth Regul. 2019;38:93–102.
Wang M, Eyre AW, Thon MR, Oh Y, Dean RA. Dynamic changes in the Microbiome of Rice during shoot and Root Growth Derived from Seeds. Front Microbiol. 2020;11:2183.
Wang W, Zhai Y, Cao L, Tan H, Zhang R. Endophytic bacterial and fungal microbiota in sprouts, roots and stems of rice (Oryza sativa L). Microbiol Res. 2016;188:1–8.
Wagner MR, Busby PE, Balint-Kurti P. Analysis of leaf microbiome composition of near‐isogenic maize lines differing in broad‐spectrum disease resistance. New Phytol. 2020;225:2152–65.
Peng X, **e J, Li W, **e H, Cai Y, Ding X. Comparison of wild rice (Oryza longistaminata) tissues identifies rhizome-specific bacterial and archaeal endophytic microbiomes communities and network structures. PLoS ONE. 2021;16:e0246687.
Østlie HM, Porcellato D, Kvam G, Wicklund T. Investigation of the microbiota associated with ungerminated and germinated norwegian barley cultivars with focus on lactic acid bacteria. Int J Food Microbiol. 2021;341:109059.
Milazzo CB, Zulak KG, Muria-Gonzalez MJ, Jones D, Power M, Bransgrove K et al. High-throughput metabarcoding characterises fungal endophyte diversity in the phyllosphere of a barley crop.Phytobiomes Journal. 2021.
Kumar M, Kumar A, Sahu KP, Patel A, Reddy B, Sheoran N, et al. Deciphering core-microbiome of rice leaf endosphere: Revelation by metagenomic and microbiological analysis of aromatic and non-aromatic genotypes grown in three geographical zones. Microbiol Res. 2021;246:126704.
Kim H, Lee Y-H. The rice microbiome: a model platform for crop holobiome. Phytobiomes J. 2020;4:5–18.
Huang W, Gilbert S, Poulev A, Acosta K, Lebeis S, Long C, Lam E. Host-specific and tissue-dependent orchestration of microbiome community structure in traditional rice paddy ecosystems. Plant Soil. 2020;452:379–95.
Chen W, Turkington TK, Lévesque CA, Bamforth JM, Patrick SK, Lewis CT et al. Geography and agronomical practices drive diversification of the epiphytic mycoflora associated with barley and its malt end product in western Canada. Agriculture, Ecosystems & Environment. 2016;226:43–55.
Ritchie and Roser. CO2 and Greenhouse Gas Emissions: Published online at OurWorldInData.org. 2020. https://ourworldindata.org/co2-and-other-greenhouse-gas-emissions. Accessed 31 Mar 2022.
Simonin M, Dasilva C, Terzi V, Ngonkeu ELM, Diouf D, Kane A, et al. Influence of plant genotype and soil on the wheat rhizosphere microbiome: evidences for a core microbiome across eight african and european soils. FEMS Microbiol Ecol. 2020;96:fiaa067.
Prudence SMM, Newitt JT, Worsley SF, Macey MC, Murrell JC, Lehtovirta-Morley LE, Hutchings MI. Soil, senescence and exudate utilisation: characterisation of the Paragon var. Spring bread wheat root microbiome. Environ microbiome. 2021;16:1–21.
Yuan C-L, Zhang L-M, Wang J-T, Hu H-W, Shen J-P, Cao P, He J-Z. Distributions and environmental drivers of archaea and bacteria in paddy soils. J Soils Sediments. 2019;19:23–37.
Long X-E, Yao H. Phosphorus input alters the assembly of rice (Oryza sativa L.) root-associated communities. Microb Ecol. 2020;79:357–66.
Barillot CDC, Sarde C-O, Bert V, Tarnaud E, Cochet N. A standardized method for the sampling of rhizosphere and rhizoplan soil bacteria associated to a herbaceous root system. Ann Microbiol. 2013;63:471–6.
Sawers RJH, Gutjahr C, Paszkowski U. Cereal mycorrhiza: an ancient symbiosis in modern agriculture. Trends Plant Sci. 2008;13:93–7.
Masson A-S, Ho Bich H, Simonin M, Nguyen Thi H, Czernic P, Moulin L, Bellafiore S. Deep modifications of the microbiome of rice roots infected by the parasitic nematode Meloidogyne graminicola in highly infested fields in Vietnam. FEMS Microbiol Ecol. 2020;96:fiaa099.
Yang F, Zhang J, Zhang H, Ji G, Zeng L, Li Y, et al. Bacterial blight induced shifts in endophytic microbiome of rice leaves and the enrichment of specific bacterial strains with pathogen antagonism. Front Plant Sci. 2020;11:963.
Yin C, Casa Vargas JM, Schlatter DC, Hagerty CH, Hulbert SH, Paulitz TC. Rhizosphere community selection reveals bacteria associated with reduced root disease. Microbiome. 2021;9:1–18.
Tian D, Chen Z, Lin Y, Liang T, Chen Z, Guo X, et al. The interaction between rice genotype and Magnaporthe oryzae regulates the assembly of rice root-associated microbiota. Rice. 2021;14:1–15.
Berg G, Cernava T. The plant microbiota signature of the Anthropocene as a challenge for microbiome research. Microbiome. 2022;10:1–12.
Suman J, Rakshit A, Ogireddy SD, Singh S, Gupta C, Chandrakala J. Microbiome as a key player in sustainable agriculture and human health.Frontiers in Soil Science. 2022:12.
European Commission. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions: A Farm to Fork Strategy for a fair, healthy and environmentally-friendly food system. 2020. https://eur-lex.europa.eu/resource.html?uri=cellar:ea0f9f73-9ab2-11ea-9d2d-01aa75ed71a1.0001.02/DOC_1&format=PDF.
Mesterházy Á, Oláh J, Popp J. Losses in the grain supply chain: causes and solutions. Sustainability. 2020;12:2342.
Solanki MK, Abdelfattah A, Sadhasivam S, Zakin V, Wisniewski M, Droby S, Sionov E. Analysis of Stored Wheat Grain-Associated Microbiota reveals Biocontrol Activity among Microorganisms against Mycotoxigenic Fungi. J Fungi. 2021;7:781.
Solanki MK, Abdelfattah A, Britzi M, Zakin V, Wisniewski M, Droby S, Sionov E. Shifts in the composition of the microbiota of stored wheat grains in response to fumigation. Front Microbiol. 2019;10:1098.
Xu S, Liu Y-X, Cernava T, Wang H, Zhou Y, **a T, et al. Fusarium fruiting body microbiome member Pantoea agglomerans inhibits fungal pathogenesis by targeting lipid rafts. Nat Microbiol. 2022;7:831–43.
Niu B, Paulson JN, Zheng X, Kolter R. Simplified and representative bacterial community of maize roots. Proc Natl Acad Sci U S A. 2017;114:E2450–9.
Jiao X, Takishita Y, Zhou G, Smith DL. Plant associated rhizobacteria for biocontrol and plant growth enhancement. Front Plant Sci. 2021;12:634796.
Kanjanasopa D, Aiedhet W, Thitithanakul S, Paungfoo-Lonhienne C. Plant growth promoting rhizobacteria as biological control agent in rice. Agricultural Sci. 2021;12:1.
Ji C, Chen Z, Kong X, **n Z, Sun F, **ng J et al. Biocontrol and plant growth promotion by combined Bacillus spp. inoculation affecting pathogen and AMF communities in the wheat rhizosphere at low salt stress conditions.Frontiers in Plant Science. 2022;13.
Zhang J, Liu Y-X, Zhang N, Hu B, ** T, Xu H, et al. NRT1. 1B is associated with root microbiota composition and nitrogen use in field-grown rice. Nat Biotechnol. 2019;37:676–84.
Hone H, Mann R, Yang G, Kaur J, Tannenbaum I, Li T, et al. Profiling, isolation and characterisation of beneficial microbes from the seed microbiomes of drought tolerant wheat. Sci Rep. 2021;11:1–12.
Beirinckx S, Viaene T, Haegeman A, Debode J, Amery F, Vandenabeele S, et al. Tap** into the maize root microbiome to identify bacteria that promote growth under chilling conditions. Microbiome. 2020;8:1–13.
Berg G, Kusstatscher P, Abdelfattah A, Cernava T, Smalla K. Microbiome Modulation—Toward a better understanding of Plant Microbiome response to Microbial Inoculants. Front Microbiol. 2021;12:803.
Alibrandi P, Cardinale M, Rahman MM, Strati F, Ciná P, de Viana ML, et al. The seed endosphere of Anadenanthera colubrina is inhabited by a complex microbiota, including Methylobacterium spp. and Staphylococcus spp. with potential plant-growth promoting activities. Plant Soil. 2018;422:81–99.
Acknowledgements
The authors appreciate the provision of the “Wheat” icon by Alex Muravev, “Corn” icon by Annette Spithoven, “barley” icon by Basti Steinhauer, and the “rice” icon by Bend Didier for implementation in figures; the icons are all freely available on the Noun Project (https://thenounproject.com/).
Funding
The authors received funding from the Austrian Science Fund (FWF; grant no. I 5085) in the project NAPERDIV that is coordinated by BioDiversa (BioDivClim) under the Horizon 2020 ERA-NET COFUND scheme.
Open access funding provided by Austrian Science Fund (FWF).
Author information
Authors and Affiliations
Contributions
TC conceived the idea for the review. KM collected all relevant literature. TC and KM drafted the manuscript. GB provided specific information related to the plant microbiome. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Conflict of Interest
The authors declare no conflict of interest.
Consent for publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Michl, K., Berg, G. & Cernava, T. The microbiome of cereal plants: The current state of knowledge and the potential for future applications. Environmental Microbiome 18, 28 (2023). https://doi.org/10.1186/s40793-023-00484-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40793-023-00484-y