
Abstract
Background
First-generation epidermal growth factor receptor tyrosine kinase inhibitors (EGFR-TKIs), such as erlotinib, have been shown to target tumors with L858R (exon 21) and exon 19 deletions, resulting in significant clinical benefits. However, acquired resistance often occurs due to EGFR mutations. Therefore, novel therapeutic strategies for treatment of patients with EGFR-positive tumors are needed. Berberine (BBR) is an active alkaloid extracted from pharmaceutical plants such as Coptis chinensis. Berberine has been shown to significantly inhibit EGFR activity and mediate anticancer effects in multiple preclinical studies. We investigated whether combining BBR with erlotinib could augment erlotinib-induced cell growth inhibition of EGFR-positive cells in a mouse xenograft model.
Methods
We examined the antitumor activities and potential mechanisms of erlotinib in combination with berberine in vitro and in vivo using the MTT assay, immunoblotting, flow cytometry, and tumor xenograft models.
Results
In vitro studies with A431 cells showed that synergistic cell growth inhibition by the combination of BBR and erlotinib was associated with significantly greater inhibition of pEGFR and pAKT, and inhibition of cyclin D and Bcl-2 expression compared to that observed in response to BBR or erlotinib alone. The efficacy of the combination treatment was also investigated in nude mice. Consistent with the in vitro results, BBR plus erlotinib significantly reduced tumor growth.
Conclusion
Our data supported use of BBR in combination with erlotinib as a novel strategy for treatment of patients with EGFR positive tumors.
Similar content being viewed by others


Introduction
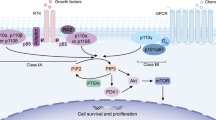
Epidermal growth factor receptor (EGFR) tyrosine kinase is a classic receptor tyrosine kinase that mediates cell proliferation in response to multiple ligands [1]. Abnormal activation of EGFR is associated with breast, lung, and colon tumors. Therefore, EGFR is the target of several cancer therapeutics [2, 3]. Epidermal growth factor receptor and related members of the ErbB family, such as ErbB2, ErbB3, and ErbB4, contain a glycosylated extracellular ligand binding domain, and single pass TM domain, and an intracellular juxtamembrane, tyrosine kinase, and autophosphorylation domain [4, 5]. Ligand binding promotes receptor dimerization and activation of intracellular protein tyrosine kinase activity. Epidermal growth factor receptor is the target of several cancer therapeutics including monoclonal antibodies targeted to the extracellular part of EGFR and small molecule inhibitors of the EGFR kinase [6]. Monoclonal antibodies include cetuximab and nimotuzumab. Nimotuzumab (h-R3) is a monoclonal antibody that targets EGFR through binding to the extracellular domain, which inhibits EGF binding [7]. The first generation of EGFR TKIs includes erlotinib, gefitinib, and lapatinib. Erlotinib is a reversible EGFR TKI used to treat NSCLC with in-frame deletions of exon 19 and an exon 21 L858R point activating mutation [8]. Nearly all patients develop resistance to erlotinib within 9–14 months of treatment [9,10,11]. Non-small cell lung cancer accounts for 85% of all lung cancers and over 60% express WT EGFR [12]. Patients with WT EGFR tumors are relatively insensitive to EGFR TKIs and patients with tumors that express WT EGFR in the absence of other targetable mutations have limited treatment options [13, 14]. The majority of EGFR positive cancers do not respond to TKIs or to mAb. Therefore, development of novel therapeutic strategies is urgently needed. Many recent studies on the efficacy of monotherapies have been severely limited by tumor cell population heterogeneity and redundant growth and survival pathways [15, 16]. Several studies showed that bevacizumab combined with erlotinib prolonged PFS in patients with NSCLC [17, 18]. Zhang et al. showed that EGCG combined with erlotinib synergistically suppressed tumor growth by targeting the common EGFR/AKT signaling pathways [19].
Natural products are widely used and their anti-cancer activities have attracted considerable attention [20, 21]. Berberine (BBR) is an active alkaloid extracted from pharmaceutical plants such as Coptis chinensis [22]. Berberine has been extensively studied in chemically induced rodent carcinogenesis models and in several types of cancers including human colon cancer, lung cancer, and breast cancer. Berberine has been shown to inhibit cancer cell growth, induce cell cycle arrest, and promote apoptosis [23,24,25]. Studies have shown that BBR (from 8.4 to 33.6 µg/mL) reduced the viability of A549 and NCI-H1299 cells in time- and concentration-dependent manners [26]. In addition, BBR was reported to inhibit tumor cell growth through suppression of phosphorylation of EGFR and other signaling mediators such as ERBB2 and VEGF [27]. A previous study showed that BBR combined with irinotecan induced apoptosis in colon cancer cells [28]. In addition, BBR combined with evodiamine synergistically suppressed MCF-7 cell proliferation [29]. These studies indicated that combination therapies may prevent drug resistance and enhance efficacy. Therefore, the goal of this study was to determine whether combination treatment with BBR and erlotinib could exert antitumor effects against A431 cells (EGFR overexpressing, WT).
The results showed that the combination of BBR with erlotinib induced synergistic antitumor effects in A431 cells in vitro and in vivo, as evidenced by inhibition of cell proliferation and induction of apoptosis.
Material and methods
Chemicals and reagents
Erlotinib was purchased from Dalian Meilun Biotechnology Co., Ltd (Dalian, People’s Republic of China). Berberine (> 98% pure) was purchased from MCE (shanghai, People’s Republic of China). Molecular biology grade dimethyl sulfoxide (DMSO) was purchased from VWR Life Science (Atlanta, GA, USA). Dulbecco’s modified Eagle’s medium (DMEM) and all chemicals used for cell culture were purchased from HyClone (Logan, UT, USA). All antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA), except total EGFR, which was purchased from Santa Cruz Animal Health (Santa Cruz, CA, USA). Secondary anti-rabbit and anti-mouse antibodies were purchased from R&D Systems (Minnesota, MN, USA). 3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide was purchased from Sigma-Aldrich (MTT; St. Louis, MO, USA).
Cell lines and cell culture
A431 cells were maintained in DMEM culture medium supplemented with 10% fetal bovine serum (FBS; Biological Industries, Ness Ziona, Israel). NCI-H1666, NCI-H441, and NCI-H1781 cells were maintained in 1640 culture medium supplemented with 10% FBS.
Cell proliferation assay
A431 and NCI-H1975 cells (1.5 × 106 cells/mL) were treated with different concentrations (0, 0.625, 1.25, 2.5, 5, and 10 µg/mL) of BBR and 0.01 µM erlotinib. NCI-H441 cells (1.5 × 106 cells/mL) were treated with different concentrations (0, 2.5, 5, 10, 20, and 40 µg/mL) of BBR and 0.01 µM erlotinib. NCI-H1781 cells (1.5 × 106 cells/mL) were treated with different concentrations (0, 5, 10, 20, 40, and 80 µg/mL) of BBR and 0.01 µM erlotinib. Cells were treated for 48 h. Cell viability was measured spectrophotometrically using the MTT assay in which DMSO was used to dissolve formazan. To evaluate the synergistic effects of BBR and erlotinib, the Q-value was calculated using King’s formula.
Western blot
A431 cells (2.5 × 106 cells/plate) in 60-mm plates were treated with BBR, erlotinib, or BBR plus erlotinib in a 60 mm plate. After treatment the cells were collected in RIPA lysis buffer containing protease inhibitors (50:1). Cell lysates were centrifuged and the supernatants were collected, mixed with sample loading buffer, and boiled for 10 min. Protein quantification was performed using BCA Protein Assay Reagent according to the manufacturer’s instructions (Beyotime, Shanghai, People’s Republic of China). Western blot analyses were performed following 8–10% SDS-PAGE and transfer to PVDF membranes. After blocking (5% skim milk) for 1 h, membranes were incubated with total EGFR, ERK, AKT, phospho-EGFR, phospho-ERK, and phospho-AKT antibodies, and anti-β-tubulin (1:1,000) at 4 ˚C overnight. The membranes were then treated with secondary anti-rabbit or anti-mouse antibodies (1:5,000) at room temperature for 1 h.
Colony formation assay
A431 cells (1 × 103 cells/plate) were treated with BBR (2.5 µg/mL), erlotinib (0.01µM), or BBR plus erlotinib in 6-well plates. After 10 days, the colonies were fixed in 4% paraformaldehyde for 15 min and stained with 0.1% crystal violet for 30 min. After washing, images were captured. Crystal violet was dissolved in 2% sodium dodecyl sulfate for 1 h and absorbance was measured at 570 nm.
Flow cytometry for evaluation of apoptotic cell death
Induction of A431 cell apoptosis by BBR and erlotinib was quantitatively determined by flow cytometry using the Annexin V-conjugated Alexafluor 488 (Alexa488) Apoptosis Detection Kit according to the manufacturer’s instructions (Becton, Dickinson, shanghai, People’s Republic of China). A431 cells were treated with BBR (2.5 µg/mL), erlotinib (0.01µM), or BBR (2.5 µg/mL) plus erlotinib (0.01µM) for 48 h. The cells were harvested, washed with PBS, and incubated with Alexa488 and propidium iodide (PI) for cellular staining at room temperature for 15 min in the dark.
In vivo tumor growth
Animal experiments were performed in accordance with ARRIVE guidelines and were approved by the Committee on Animal Handling of Yunnan Agricultural University (YNAU2019LLWYH003-1b). Male athymic nude mice were purchased from Cavens Lab Animal (Changzhou, China). They were housed in the Animal Care Facility in the Key Laboratory of Pu-er Tea Science under a 12 h light-dark cycle (lights on at 9:00 am; lights off at 9:00 pm). Mice had free access to water and food in a room controlled at 22–25 °C and 50–60% humidity. A431 cells (5 × 106) were suspended in 200 µL of normal saline and subcutaneously implanted into 6-7-week-old male athymic nude mice. Tumors began to appear 3 days after the tumor cells were injected. When the tumor volume reached approximately 50 mm3, the tumor-bearing mice received daily (five times per week) intraperitoneal injections with vehicle control, BBR (15 mg/kg), erlotinib (25 mg/kg), or the combination of BBR (15 mg/kg) and erlotinib (25 mg/kg) for 26 days. During treatment mouse body weight and tumor volume were recorded every two days. Tumor volume was calculated using the following formula: of A × B2 × 0.5 (A is solid tumor length, B is width). After 26 days the mice were euthanized by intraperitoneal injection of sodium pentobarbital. One portion of the tumor was fixed in formalin and the other portion of the tumor was frozen at − 80 °C.
Immunohistochemical staining (IHC)
Xenograft tumor tissue sections were incubated at 65 °C for 3 h, then deparaffinized and rehydrated. Deparaffinized and rehydrated sections were treated for antigen retrieval using sodium citrate buffer (pH 6.0). The slides were blocked using serum albumin for 20 min at 37 °C. The slides were then incubated with primary antibody at 4 °C overnight. Then, the slides were incubated with secondary antibody for 30 min at 37 °C. Target proteins and cell nuclei were visualized using DAB substrate and hematoxylin, respectively. Slides were visualized using a light microscope. Images were captured using 400× magnification.
Statistical analysis
All data were analyzed using SPSS17.0 software (mean ± SEM, one-way ANOVA). Student’s t-test was used to determine tumor volume statistical significance. P < 0.05 was considered significant, P < 0.01 was a significant difference, and P < 0.001 was extremely significant. The experimental data were plotted using GraphPad Prism software.
Results
Berberine combined with erlotinib decreased proliferation of EGFR positive cells
To study the sensitivity of EGFR-positive cell lines to BBR and erlotinib, we examined 4 cell lines (A431, NCI-H441, NCI-H1781 and NCI-H1975), of which A431, NCI-H441, and NCI-H1781 cells expressed wild-type EGFR and NCI-H1975 cells had EGFRL858R/T790M double mutation. Epidermal growth factor receptor positive cells were treated with 0 to 80 µg/mL BBR. As shown in Fig. 1a, b and c, and 1d, BBR inhibited growth of all 4 cell lines in a dose-dependent manner at 48 h. A previous report showed that BBR did not induce toxicity in normal human epidermal keratinocytes [28]. Next, we treated with BBR with erlotinib and evaluated cell viability using the MTT assay. As shown in Fig. 1a and d, treatment with BBR and erlotinib was significantly more effective than either agent alone in each of the 4 cell lines. The degree of response to the combination therapy varied between cell lines. Different sensitivities to various treatments were partly attributed to tumor heterogeneity. Berberine and erlotinib decreased cell viability up to 28.69% at 2.5 µg/mL and 7.68% at 0.01µM, respectively (lane 2 and lane 7, Fig. 1a). Moreover, the combination of BBR and erlotinib further decreased cell viability (63.58% inhibition) compared to either agent alone in A431 cells (lane 8, Fig. 1a). Berberine and erlotinib exerted more pronounced synergistic effects in A431 cells than in other cells based on Q-value (Fig. 1e). Berberine and erlotinib monotherapies induced dose-dependent inhibition of A431 cell expansion and 2.5 µg/mL BBR and 0.01µM erlotinib were identified as the best concentrations for assessment of interactions.

Combination therapy inhibited EGFR positive cell proliferation. (a) (b) (c) (d) Endothelial growth factor receptor positive cells were treated with BBR combined with erlotinib for 48 h, and cell viability was determined using the MTT assay. Data are expressed in terms of the percent of control cells (DMSO treated) as the mean ± SEM of 3 replicates. (e) Q-value. The Q-value was calculated using King’s formula. (f) Colony formation assay was used to determine the inhibitory effect of BBR combined with erlotinib on A431 cells. Representative images of colonies from 60-mm plates in the colony formation assay. (g) Quantification of the colony number of A431 cells. Data were analyzed using one-way ANOVA, #P < 0.05; ##P < 0.01; ###P < 0.001 vs. control. *P < 0.05; **P < 0.01; ***P < 0.001 vs. combination-therapy group, &P < 0.05; &&P < 0.01; &&&P < 0.001 vs. erlotinib
These results were further confirmed using a clonogenic assay. Anchorage independent colony formation assays further demonstrated the synergistic effect of BBR and erlotinib on cell proliferation. The results showed that co-treatment with BBR and erlotinib (22.37% survival rate) significantly inhibited NCI-H441 colony formation compared with either BBR (40.79% survival rate) or erlotinib (78.21% survival rate) alone, (Fig. 1f and g). These results indicated that the combination of BBR and erlotinib synergistically inhibited cell growth in A431 cells.

Berberine combined with erlotinib inhibited the EGFR signaling pathway in A431 cells. (a) A431 cells were treated with a range of BBR doses for 48 h. Phosphorylation of EGFR, AKT, and ERK were evaluated using western blot. The p-EGFR, p-ERK1/2 p-AKT, EGFR, ERK1/2, AKT, and β-tubulin proteins were separated using 8% SDS-PAGE. Each experiment was performed 3 times independently. (c) A431 cells were treated with BBR (2.5 µg/mL) combined with erlotinib (0.01µM) for 30 min. Phosphorylation of each protein was evaluated using western blot. Each experiment was also performed 3 times independently. (b, d) Quantification of EGFR and downstream signaling proteins. Data were analyzed using one-way ANOVA, #P < 0.05; ##P < 0.01; ###P < 0.001 vs. control. *P < 0.05; **P < 0.01; ***P < 0.001 vs. combination-therapy group
Berberine combined with erlotinib inhibited the expression of EGFR and its downstream targets
We examined the effects of BBR and erlotinib on EGFR signal transduction in A431 cells using immunoblotting. As shown in Fig. 2a-b, BBR inhibited EGFR phosphorylation in a dose-dependent matter (2.5 to 10 µg/mL) in A431cells. We then evaluated the effects of BBR combined with erlotinib on the EGFR pathway. Western blot analysis showed that 30 min treatment with BBR and erlotinib inhibited the EGFR pathway (p < 0.05), as evidenced by reduced phosphorylation of EFGR (Y1045, Y1068), extracellular regulated protein kinases (ERK), and protein kinase B (AKT) (lane 4, Fig. 2c and d). These results indicated that combined treatment with BBR with erlotinib inhibited the EGFR pathway.
Combined treatment with BBR and erlotinib induced apoptosis in A431cells
We evaluated the mechanism of combined BBR and erlotinib therapy on A431 cell proliferation and apoptosis. As shown in Fig. 3a-b, A431 cells were treated with BBR, erlotinib, or both for 48 h, then stained with annexin V-FITC/PI and analyzed using flow cytometry. Combination treatment induced more apoptosis (32.63%) than the untreated control (16.15%), BBR (29.47%), or erlotinib (21.84%). We also determined the expression levels of Bax and Bcl-2, which are important in apoptosis. Compared to the untreated control, BBR significantly reduced the Bcl-2/Bax ratio a concentration-dependent manner (Fig. 3c and d). Compared to the untreated control (p < 0.001) and the individual drugs (p < 0.05), BBR plus erlotinib significantly reduced the Bcl-2/Bax ratio (Fig. 3e and f). These results showed that erlotinib did not effectively induce apoptosis, but the combination of BBR and erlotinib significantly increased apoptotic cell death.

Expression level of apoptosis proteins in response to different drug treatments in A431 cells. (a) Berberine (2.5 µg/mL) and erlotinib (0.01 µM) synergistically enhanced apoptosis in A431 cells as determined by flow cytometry using the Annexin V-conjugated Alexafluor 488 (Alexa488) Apoptosis Detection Kit. (b) The ratio of apoptotic cells in each group. (c) Treatment of A431 cells with varying concentrations of BBR for 48 h. The expression levels of the apoptotic proteins Bcl-2/Bax were determined using western blot. Bcl-2, Bax, and cyclin D1 protein were separated using 10% SDS-PAGE. Each experiment was performed 3 times independently. (e) Treatment of A431 cells with BBR (2.5 µg/mL) combined with erlotinib (0.01 µM) for 48 h. The expression of the apoptotic proteins Bcl-2/Bax was determined using western blot. (d, f) Quantification of Bcl-2/Bax and cyclin D1 proteins. Each experiment was performed 3 times independently. Data were analyzed using one-way ANOVA, #P < 0.05; ##P < 0.01; ###P < 0.001 vs. control. *P < 0.05; **P < 0.01; ***P < 0.001 vs. combination therapy group
We then evaluated the effects of BBR plus erlotinib on cell cycle progression in A431 cells. Treatment with erlotinib alone had minimal effects on cyclin D in A431 cells (p > 0.05). Combined treatment with BBR and erlotinib led to significantly lower cyclin D1 expression than that in cells treated with BBR (p < 0.01) or erlotinib (p < 0.001, Fig. 3e and f).
Effect of BBR in combination with erlotinib on tumor growth in A431 xenograft models
We determined the effects of BBR, erlotinib, and the combination of BBR and erlotinib on tumor growth using a xenograft model generated by subcutaneous dorsal implantations of A431 cells into nude mice. To evaluate the in vivo toxicity of BBR combined with erlotinib, we monitored mouse body weight. The results showed that treatment with BBR and/or erlotinib was well-tolerated without significant weight loss (p > 0.05, Fig. 4a). The combination of BBR plus erlotinib delayed in vivo tumor growth compared with that observed in the vehicle control group, as evidenced by inhibition of tumor growth in A431 cells (tumor growth inhibition (TGI) value of 78.06%). This TGI value was higher than that observed in the BBR (67.73%) and erlotinib (64.58%) groups (Fig. 4b, c and d). We evaluated the mechanisms of action of BBR and erlotinib using IHC staining. Immunohistochemistry showed that combination therapy resulted in greater inhibition of pEGFR and pERK1/2 than that in the other groups, but did not induce degradation of EGFR (p < 0.05). Evaluation of the expression of the proliferation marker protein Ki-67 showed that combined treatment resulted in greater inhibition of proliferation in the xenograft tumors than that observed in response to treatment with BBR or erlotinib alone (p < 0.05, Fig. 4e and f). Protein extracted from the xenograft tumor tissues was examined to evaluate whether the inhibition of pEGFR by treatment with BBR and erlotinib observed in vitro was also observed in vivo. Phosphorylation of EGFR was significantly inhibited by combined treatment with BBR and erlotinib in tumor samples obtained from mice (Fig. 4g).

Berberine combined with erlotinib inhibited tumor growth in vivo. Body weights (a) and tumor volumes (b) of A431 xenograft tumors treated with vehicle control, BBR (15 mg/kg), erlotinib (25 mg/kg), and BBR plus erlotinib. (c) Representative photographs of tumor tissues. (d) Tumor weights. (e) Representative image of IHC (original magnification ×400). (f) Immunohistochemical staining analysis of pEGFR, pERK1/2, and Ki67 in A431 xenograft tumors. (g) The expression levels of EGFR and pEGFR in A431 xenograft tumors. Data were analyzed using one-way ANOVA and Student’s t-test, *P < 0.05; **P < 0.01; ***P < 0.001 vs. combination-therapy group. #P < 0.05; ##P < 0.01; ###P < 0.001 vs. control
Discussion
Lung cancer can be categorized as non-small cell lung cancer (NSCLC) or small cell lung cancer (SCLC). Abnormal activation of EGFR is observed in approximately 50% of patients with NSCLC. Therefore, targeting EGFR has been identified as an effective anticancer strategy, and EGFR has become a well-established target for treatment of NSCLC [2, 3]. The two classes of anti-EGFR agents currently used are MAbs and TKIs. Although MAbs and TKIs exert anti-tumor effects, most patients develop drug resistance within 9–14 months [9,10,11]. Only a small fraction of EGFR positive advanced colorectal cancers expressing wild type KRAS respond to anti-EGFR mAbs, and acquired resistance also commonly occurs [30]. Therefore, the majority of EGFR positive cancers do not respond to TKIs or to mAb. As such, there is an obvious need to identify and develop novel treatment strategies that can complement current EGFR targeted therapy.
Berberine is an isoquinoline quaternary alkaloid derived from Coptis chinensis that has been used as a therapeutic agent for treatment of cancer, bacterial infections, diabetes, and cardiovascular and inflammatory diseases [31]. The EGFR signaling pathways play a critical role in proliferation, invasion, and survival. The PI3K/Akt and RAS/RAF/MEK/ERK pathways are the two main downstream pathways of EGFR signaling. Previous studies have shown that BBR decreased the phosphorylation of PI3K/AKT, ERK, and GSK3β in B16F10 melanoma cells and inhibited the activation of EGFR in these tumors [32, 33]. The specific antitumor activity of BBR on EGFR signaling pathways suggested its potential as an inexpensive, relatively non-toxic therapy when combined with EGFR-TKIs based on the premise that combination treatment could exert synergistic antitumor effects compared with monotherapies. We showed that BBR and erlotinib synergistically suppressed tumor growth by acting on the common EGFR/AKT signaling pathways in EGFR positive cells.
We used A431 cells to identify novel combination therapies targeting therapy-refractory lung cancer. We showed that combined treatment with BBR and erlotinib synergistically inhibited colony formation (Fig. 1) in vitro and reduced tumor growth in vivo (Fig. 4). Treatment with BBR and erlotinib resulted in inhibition of phosphorylation of EGFR and its downstream effectors (pEGFR-Y1045, pEGFR-Y1068, pERK, and pAKT) (Fig. 2). Berberine exhibited synergistic antitumor activity when combined with erlotinib and did not induce significant side effects in our animal experiment. Neither BBR nor erlotinib alone exerted significant therapeutic effects in the A431 model. In contrast, combination therapy led to tumor regression, significant reductions in pEGFR and pERK1/2, and significantly decreased Ki67 staining (Fig. 4). The present study suggested a novel mechanism by which the combination of erlotinib and BBR resulted in decreased EGFR pathway activity.
Members of the Bcl-2 family such as Bax and Bcl-2 play important roles in regulating the mitochondria-dependent apoptotic pathway. The Bax/Bcl-2 expression ratio is critical for induction of apoptosis [34]. Treatment of A431 cells with BBR has been shown to increase the expression of Bax and decrease the expression of Bcl-2, resulting in an increase in the Bax/Bcl-2 ratio [35, 36]. Our findings showed that treatment of A431 cells with BBR and erlotinib led to a decrease in the Bcl-2/Bax ratio (Fig. 3e and f). Epidermal growth factor receptor and its downstream effectors ERK/MAPK, AKT, STAT, and cyclin D are involved in cell proliferation and differentiation. Cyclin D may be an essential element in the pathway that connects EGFR-mediated mitogenic signals to the cell cycle at the G1/S boundary [37, 38]. Studies have shown that BBR suppressed cancer cells by inducing G1-phase cell cycle arrest and reducing cyclin D expression in lung and hepatoma cells [36, 39]. Consistent with these findings, our results showed that combined treatment inhibited the expression of phosphorylated ERK and AKT, and the expression of cyclin D in A431 cells. (Fig. 3e and f).
Conclusion
In conclusion, our results showed that combined treatment with BBR and erlotinib synergistically inhibited A431 tumor growth in vitro and in vivo. Mechanistic studies showed that synergistic cell growth inhibition by the combination of BBR and erlotinib was associated with significantly greater inhibition of pEGFR. Combined treatment resulted in significantly greater inhibition of tumor growth through increased apoptosis and decreased cell proliferation compared to those observed in response to BBR or erlotinib alone. Our study suggested that combination therapy may provide a novel potential therapeutic strategy for treatment of EGFR positive cells.
Data Availability
All data generated or analyzed during this study are included in this published article.
Abbreviations
- Bax:
-
Bcl-2-associated X protein
- Bcl-2:
-
B cell lymphoma 2
- DMEM:
-
Dulbecco’s Modified Eagle’s Medium
- DMSO:
-
dimethyl sulfoxide
- MTT:
-
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
- EGFR-TKIs:
-
epidermal growth factor receptor tyrosine kinase inhibitors
- BBR:
-
Berberine
References
Macdonald-Obermann JL, Pike LJ. Different epidermal growth factor (EGF) receptor ligands show distinct kinetics and biased or partial agonism for homodimer and heterodimer formation. J Biol Chem. 2014;289(38):26178–88.
Scharadin T, Saldana M, Schlein M, Hoang-Phou S, Trans D, Chang D, et al. Abstract 3321: using NLPs to study EGFR structure, activation, and inhibition. Cancer Res. 2014;74:3321.
Al-Akhrass H, Naves T, Vincent F, Magnaudeix A, Durand K, Bertin F, et al. Sortilin limits EGFR signaling by promoting its internalization in lung cancer. Nat Commun. 2017;8:1182.
Hynes NE, Lane HA. ERBB receptors and cancer: the complexity of targeted inhibitors. Nat Rev Cancer. 2005;5:341–54.
Prenzel N, Fischer OM, Streit S, Hart S, Ullrich A. The epidermal growth factor receptor family as a central element for cellular signal transduction and diversification. Endocr-Relat Cancer. 2001;8:11–31.
José B, Carlos LA. Critical update and emerging trends in epidermal growth factor receptor targeting in cancer. J Clin Oncol. 2005;23:2445–59.
Takeda M, Okamoto I, Nishimura Y, Nakagawa K. Nimotuzumab, a novel monoclonal antibody to the epidermal growth factor receptor, in the treatment of non-small cell lung cancer. Lung Cancer (Auckl). 2011;2:59–67.
Zhou C, Wu YL, Chen G, Feng J, Liu XQ, Wang C, et al. Erlotinib versus chemotherapy as first-line treatment for patients with advanced EGFR mutation-positive non-small-cell lung cancer (OPTIMAL, CTONG-0802): a multicentre, open-label, randomised, phase 3 study. Lancet Oncol. 2011;12:735–42.
Viloria-Petit AM, Kerbel RS. Acquired resistance to EGFR inhibitors: mechanisms and prevention strategies. Int J Radiat Oncol Biol Phys. 2004;58:914–26.
Zhang Z, Lee JC, Lin L, Olivas V, Au V, LaFramboise T, et al. Activation of the AXL kinase causes resistance to EGFR-targeted therapy in lung cancer. Nat Genet. 2012;44:852–60.
Chen MC, Chen CH, Wang JC, Tsai AC, Liou JP, Pan SL, et al. The HDAC inhibitor, MPT0E028, enhances erlotinib-induced cell death in EGFR-TKI-resistant NSCLC cells. Cell Death Dis. 2013;4:e810.
Itchins M, Clarke S, Pavlakis N. Do EGFR tyrosine kinase inhibitors (TKIs) still have a role in EGFR wild-type pre-treated advanced non-small cell lung cancer (NSCLC)?-the shifting paradigm of therapeutics. Transl Lung Cancer Res. 2018;7(Suppl 1):S39–S45.
Laurie SA, Goss GD. Role of epidermal growth factor receptor inhibitors in epidermal growth factor receptor wild-type non-small-cell lung cancer. J Clin Oncol. 2013;31:1061–9.
Lee JK, Hahn S, Kim DW, Suh KJ, Keam B, Kim TM, et al. Epidermal growth factor receptor tyrosine kinase inhibitors vs conventional chemotherapy in non-small cell lung cancer harboring wild-type epidermal growth factor receptor: a meta-analysis. JAMA. 2014;311:1430–7.
Herbst RS. Erlotinib (Tarceva): an update on the clinical trial program. Semin Oncol. 2003;30:34–46.
Zhang X, Chen ZG, Choe MS, Lin Y, Sun SY, Wieand HS, et al. Tumor growth inhibition by simultaneously blocking epidermal growth factor receptor and cyclooxygenase-2 in a xenograft model. Clin Cancer Res. 2005;11:6261–9.
Wang Y, Wang H, Jiang Y, Zhang Y, Wang X. A randomized phase III study of combining erlotinib with bevacizumab and panitumumab versus erlotinib alone as second-line therapy for chinese patients with non-small-cell lung cancer. Biomed Pharmacother. 2017;89:875–9.
Zhao B, Zhang W, Yu D, Wei Y. Erlotinib in combination with bevacizumab has potential benefit in non-small cell lung cancer: a systematic review and meta-analysis of randomized clinical trials. Lung Cancer. 2018;122:10–21.
Zhang X, Zhang H, Tighiouart M, Lee J, Shin H, Khuri F, et al. Synergistic inhibition of head and neck tumor growth by green tea (-)-epigallocatechin-3-gallate and EGFR tyrosine kinase inhibitor. Int J Cancer. 2010;123(5):1005–14.
Jung S, Lee M, Lim D, Jeong C, Kim E, Lim T, et al. Butein, a novel dual inhibitor of MET and EGFR, overcomes gefitinib-resistant lung cancer growth. Mol Carcinogen. 2015;54:322–31.
Li J, Guo Q, Lei X, Zhang L, Su C, Liu Y, et al. Pristimerin induces apoptosis and inhibits proliferation, migration in H1299 lung cancer cells. J Cancer. 2020;11:6348–55.
Chen C, Yu Z, Li Y, Fichna J, Storr M. Effects of berberine in the gastrointestinal tract - a review of actions and therapeutic implications. Am J Chinese Med. 2014;42:1053–70.
Li J, Liu F, Jiang S, Liu J, Chen X, Zhang S, et al. Berberine hydrochloride inhibits cell proliferation and promotes apoptosis of non-small cell lung cancer via the suppression of the MMP2 and Bcl-2/Bax signaling pathways. Oncol Lett. 2018;15:7409–14.
Park JJ, Seo SM, Kim EJ, Yoon-** L, Ko YG, Ha J, et al. Berberine inhibits human colon cancer cell migration via AMP-activated protein kinase-mediated downregulation of integrin b1 signaling. Biochem Biophys Res Commun. 2012;426:461–7.
Tak J, Sabarwal A, Shyanti R, Singh R. Berberine enhances posttranslational protein stability of p21/cip1 in breast cancer cells via down-regulation of akt. Mol Cell Biochem. 2019;458(1–2):49–59.
Katiyar SK, Meeran SM, Katiyar N, Akhtar S. p53 cooperates berberine-induced growth inhibition and apoptosis of non-small cell human lung cancer cells in vitro and tumor xenograft growth in vivo. Mol Carcinogen. 2009;48:24–37.
Kaboli PJ, Rahmat A, Ismail P, Ling KH. Targets and mechanisms of berberine, a natural drug with potential to treat cancer with special focus on breast cancer. Eur J Pharmacol. 2014;740:584–95.
Ruan H, Zhan YY, Hou J, Xu B, Chen B, Tian Y, et al. Berberine binds RXRα to suppress β-catenin signaling in colon cancer cells. Oncogene. 2017;36:6906–18.
Du J, Sun Y, Lu YY, Lau E, Zhao M, Zhou QM, et al. Berberine and evodiamine act synergistically against human breast cancer MCF-7 cells by inducing cell cycle arrest and apoptosis. Anticancer Res. 2017;37:6141–51.
Thomas R, Zhang. Rethink of EGFR in cancer with its kinase independent function on board. Front Oncol. 2019;9:800.
Liu Y, Liu X, Zhang N, Yin M, Dong J, Zeng Q, et al. Berberine diminishes cancer cell PD-L1 expression and facilitates antitumor immunity via inhibiting the deubiquitination activity of CSN5. Acta Pharm Sin B. 2020;10(12):2299–312.
Song YC, Lee Y, Kim HM, Hyun MY, Lim YY, Song KY, et al. Berberine regulates melanin synthesis by activating PI3K/AKT, ERK and GSK3β in B16F10 melanoma cells. Int J Mol Med. 2015;35:1011–6.
Zou K, Li Z, Zhang Y, Zhang HY, Li B, Zhu WL, et al. Advances in the study of berberine and its derivatives: a focus on anti-inflammatory and anti-tumor effects in the digestive system. Acta Pharmacol Sin. 2017;38:157–67.
Garcia-Saez AJ. The secrets of the Bcl-2 family. Cell Death Differ. 2012;19:1733–40.
Ma X, Zhou J, Zhang CX, Li XY, Li N, Ju RJ, et al. Modulation of drug-resistant membrane and apoptosis proteins of breast cancer stem cells by targeting berberine liposomes. Biomaterials. 2013;34:4452–65.
Mantena SK, Sharma SD, Katiyar SK. Berberine inhibits growth, induces G1 arrest and apoptosis in human epidermoid carcinoma A431 cells by regulating cdki-cdk-cyclin cascade, disruption of mitochondrial membrane potential and cleavage of caspase 3 and PARP. Carcinogenesis. 2006;27:2018–27.
Du H, Xu B, Wu C, Li M, Ran F, Cai S, et al. Effects of CS-1 on A431 cell proliferation, cell cycle, and epidermal growth factor receptor signal transduction. Acta Biochim Biophys Sin (Shanghai). 2012;44(2):136–46.
Busse D, Doughty RS, Ramsey TT, Russel WE, Price JO, Flanagan W, et al. Reversible G1 arrest induced by inhibition of the epidermal growth factor receptor tyrosine kinase requires up-regulation of p27kip1 independent of MAPK activity. J Biol Chem. 2000;275:6987–95.
Wang N, Wang X, Tan HY, Li S, Tsang CM, Tsao SW, et al. Berberine suppresses cyclin D1 expression through proteasomal degradation in human hepatoma cells. Int J Mol Sci. 2016;17:1899.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the National Science Foundation of China (81960653), the Yunnan Fundamental Research Project (202101AU070085), the Research Start-up Fund of Yunnan Agricultural University (KY2019-17), the Yunnan Provincial Key Programs of Yunnan Eco-Friendly Food International Cooperation Research Center Project (2019ZG00904 and 2019ZG00909), the Major Scientific and Technological Special Project of Yunnan Province (2017ZF003, 2018ZG010, and 2018ZG013), and the Science and Technology Plan Project of Yunnan Province (2018IA060).
Author information
Contributions
Jun Sheng, Xuanjun Wang, and Yan** Huang conceived and designed the experiments. ** Huang wrote the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All experiments were performed in accordance with relevant guidelines and regulations. Specifically, all animal procedures were performed in accordance with the ARRIVE guidelines and were approved by the Committee on Animal Handling of Yunnan Agricultural University (YNAU2019LLWYH003-1b).
Consent for publication
Not applicable.
Conflict of interest
The authors declare no conflicts of interest in this work.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Supplementary Information
Not applicable.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Cuan, X., Yang, X., Zhu, W. et al. Antitumor effects of erlotinib in combination with berberine in A431 cells. BMC Pharmacol Toxicol 24, 29 (2023). https://doi.org/10.1186/s40360-023-00661-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40360-023-00661-2