
Abstract
In this paper, a class of virus infection models with CTLs response is considered. We incorporate an immune delay and two intracellular delays into the virus infection model. It is found that only incorporating two intracellular delays almost does not change the dynamics of the system, but incorporating an immune delay changes the dynamics of the system very greatly, namely, a Hopf bifurcation and oscillations can appear. Those results show immune delay dominates intracellular delays in some viral infection models, which indicates the human immune system has a special effect in virus infection models with CTLs response, and the human immune system itself is very complicated. In fact, people are aware of the complexity of the human immune system in medical science, which coincides with our investigating. We also investigate the global Hopf bifurcation of the system with the immune delay as a bifurcation parameter.
Similar content being viewed by others


1 Introduction
People utilize widely mathematical models to investigate viral infections currently, for example, HBV (hepatitis B virus), HCV (hepatitis C virus), HIV, and so on [1–7]. Perelson et al. proposed a standard and classic model (probably the first) for HIV dynamics in [8, 9] as follows:
Here \(x(t)\) represents the concentration of uninfected cells at time t, \(y(t)\) represents the concentration of infected cells that can produce a virus at time t, \(v(t)\) represents the concentration of viruses at time t. λ is the rate at which new healthy cells are generated. \(d_{1}\), \(d_{2}\), \(d_{3}\) are the death rates of uninfected cells, infected cells, and virus cells, respectively. \(\beta x(t)v(t)\) is the bilinear incidence between infected cells and uninfected cells. Free virus is produced from infected cells at the rate \(ky(t)\).
Many researchers also consider system (1.1) as a basic virus infection model for various other viruses, such as HBV [10, 11]; HCV [12]. It is well known that an immune response exists universally and plays an important role in many viral infections [3, 13, 14]. So a typical extension model for those infections with a cytotoxic T lymphocytes (CTLs) response is considered in [14],
Here \(z(t)\) represents the concentration of the cells of the immune response. \(d_{4}\) is the death rate of cells of the immune response. CTLs-driven elimination of infected cells is assumed to be of the form \(\mu y(t)z(t)\), where μ is the rate of CTLs elimination. The CTLs response to the infection is modeled by \(\gamma y(t)z(t)\).
In fact, we should note that there are delays in the infection process, including intracellular delay and CTLs immune response delay. We refer readers to [4, 5] and references therein. In [15], Zhu and Zou studied a HIV infection model with intracellular delay as follows:
In this paper, we incorporate two intracellular delays and immune response delay in system (1.2). Namely, we incorporate a time delay \(\tau_{1}\) to describe the period between healthy cells’ contacting with viruses and complete production of viral RNA and protein. We incorporate the time delay \(\tau_{2}\) to describe the period between complete production of viral RNA and protein and actually releasing new mature viruses. \(\tau_{3}\) represents the CTLs immune response delay. So we have the following system:
A question is how the intracellular delays \(\tau_{1}\), \(\tau_{2}\) and the immune delay \(\tau_{3}\) affect the dynamics of the system (1.4). That is the main goal of this paper. We find that only incorporating two intracellular delays \(\tau_{1}\) and \(\tau_{2}\) almost does not change the dynamics of the system, but incorporating the immune delay \(\tau_{3}\) changes the dynamics of the system very greatly, namely, a Hopf bifurcation and oscillations can occur. Those results show that the immune delay dominates the intracellular delays in this class of viral infection models, which indicates the human immune system has a special effect in virus infection models with CTLs response, and the human immune system itself is very complicated. People are aware of the complexity of the human immune system in medical science, which coincides with our investigation.
The paper is organized as follows. In the next section, the attractive region and equilibria for system (1.4) are discussed and the two threshold parameters \(R_{0}\) and \(R_{1}\) are introduced. In Section 3, by combining the linear stability theory and the LaSalle-Lyapunov theorem, the global stability of \(P_{0}\) and \(P_{1}\) when \(R_{0}<1\) and \(R_{1}<1<R_{0}\) is discussed, respectively. By analyzing the distribution of the eigenvalues, the dynamics of the system when \(R_{1}>1\) is investigated. In Section 4, we study the global Hopf branch of the system. Numerical simulations are presented in Section 5 to illustrate the analysis results. The paper ends with a brief conclusion.
2 Attractive region and equilibria
Let \(\tau=\max\{\tau_{1}, \tau_{2}, \tau_{3}\}\); we denote by \(\mathcal {C}=\mathcal{C}([-\tau,0],\mathbb{R}^{4})\) the Banach space of continuous real-valued functions on the interval \([-\tau,0]\), with norm
The nonnegative cone of \(\mathcal{C}\) is defined as
The initial conditions for system (1.4) are chosen at \(t=0\) as
Proposition 2.1
Under the initial condition (2.1), all solutions of system (1.4) are positive and ultimately bounded in \(\mathcal{C}\). Furthermore, all solutions eventually enter and remain in the following bounded region:
where
and ε is an arbitrarily small positive number.
Proof
First, we prove that \(x(t)\) is positive for \(t\geq0\). Assuming the contrary and letting \(t_{1}>0\) be the first time such that \(x(t_{1})=0\), by the first equation of system (1.4), we have \(x'(t_{1})=\lambda>0\), and hence \(x(t)<0\) for \(t\in(t_{1}-\eta ,t_{1})\) and sufficiently small \(\eta>0\). This contradicts \(x(t)>0\) for \(t\in[0,t_{1})\). It follows that \(x(t)>0\) for \(t>0\). From the fourth equation of (1.4), we use the method of the steps to prove \(z(t)>0\) for \(t>0\). From the third equation of (1.4), we can prove \(v(t)>0\) for \(t>0\). From the second equation of (1.4), we can obtain \(y(t)>0\) for \(t>0\).
Next we show that positive solutions of (1.4) are ultimately uniformly bounded for \(t\geq0\). From the first equation of system (1.4), we obtain
and thus
Adding the first two equations of (1.4) leads to
where \(\tilde{d}=\min\{d_{1},d_{2}\}\). Thus
Adding the first three equations of (1.4), we have
where \(\hat{d}=\min\{{d_{1},\frac{d_{2}}{2},d_{3}}\}\). Thus
Adding all four equations of (1.4), we have
where \(d=\min\{{d_{1},\frac{d_{2}}{2},d_{3},d_{4}}\}\). Thus
Therefore, \(x(t)\), \(y(t)\), \(v(t)\), and \(z(t)\) are ultimately uniformly bounded in \(\mathcal{C}\). □
As a consequence of Proposition 2.1, we know that the dynamics of system (1.4) can be analyzed in the following bounded region:
Furthermore, the region Γ is attractive with respect to system (1.4).
The system has two threshold parameters,
They are called the basic reproduction numbers for viral infection and for CTL response [3, 16]. We note that \(R_{1}< R_{0}\) always holds.
System (1.4) always has an infection-free equilibrium \(P_{0}=(x_{0},0,0,0)\), \(x_{0}=\frac{\lambda}{d_{1}}\). In addition to \(P_{0}\), the system can have two other equilibria \(P_{1}=(\overline {x},\overline{y},\overline{v},0)\) and \(P_{2}=(x^{*},y^{*},v^{*},z^{*})\), where x̅, y̅, v̅, \(x^{*}\), \(y^{*}\), \(v^{*}\), and \(z^{*}\) are all positive. The equilibrium \(P_{1}=(\overline{x},\overline{y},\overline{v},0)\) exists if and only if \(R_{0}>1\) and
The equilibrium \(P_{2}\) exists if and only if \(R_{1} >1\). The equilibrium \(P_{2}=(x^{*},y^{*},v^{*},z^{*})\) is given by
3 Global stability and Hopf bifurcation
We investigate stability of the equilibria and the Hopf bifurcation in this section. First, \(P_{0}\) is considered in the following.
3.1 Global stability of \(P_{0}\)
In this subsection, we rigorously show that when \(R_{0}<1\), the infection-free equilibrium \(P_{0}\) is globally asymptotically stable in Γ.
Theorem 3.1
If \(R_{0}<1\), the infection-free equilibrium \(P_{0}\) of system (1.4) is globally asymptotically stable in Γ. If \(R_{0}>1\), \(P_{0}\) is unstable.
Proof
First we prove \(P_{0}\) is locally asymptotically stable. The characteristic equation associated with the linearization of system (1.4) at \(P_{0}\) is given by
Obviously we have
and we consider the equation
Notice 0 is not a root of (3.2) because of \(R_{0}<1\). Following the method in [17], if \(\xi=i\omega\) (\(\omega>0\)) is a purely imaginary root of (3.2), we have
Squaring and adding both equations of (3.3), it follows that
Let \(u=\omega^{2}\). Then (3.4) becomes
From \(R_{0}<1\), we easily see that (3.5) has no positive root. Therefore, all roots of (3.1) have negative real parts. So \(P_{0}\) is locally asymptotically stable when \(R_{0}<1\).
Next, we prove \(P_{0}\) is globally attractive in Γ if \(R_{0}<1\). To prove this, we consider a Lyapunov functional \(L:\mathcal {C}\rightarrow\mathbb{R}\) given by
Here \(x_{t}(s)=x(t+s)\), for \(s\in[-\tau,0]\), and thus \(x(t)=x_{t}(0)\) in this notation.
Calculating the time derivative of L along solution of system (1.4), it follows that
\(R_{0}<1\) ensures that \(L'|_{\text{(1.4)}}\leq0\), and \(L'=0\) if and only if
it can be verified that the maximal invariant set in \(\{L'|_{\text{(1.4)}}=0 \}\) is the set
By the LaSalle-Lyapunov theorem, we conclude that M is globally attractive in Γ if \(R_{0}<1\). So \(P_{0}\) is globally attractive in Γ.
Therefore, \(P_{0}\) is globally asymptotically stable in Γ.
We can easily see that (3.1) has a root with a positive real part when \(R_{0}>1\). \(P_{0}\) is unstable when \(R_{0}>1\). □
Remark 3.2
Obviously \(P_{0}\) is globally asymptotically stable without any delays when \(R_{0}<1\), but after incorporating three delays (a immune delay and two intracellular delays), \(P_{0}\) is still globally asymptotically stable. Delays do not destroy the globally asymptotical stability of \(P_{0}\).
3.2 Global stability of \(P_{1}\)
Theorem 3.3
If \(R_{1}<1<R_{0}\), then the equilibrium \(P_{1}\) is globally asymptotically stable. If \(R_{1}>1\), \(P_{1}\) is unstable.
Proof
Let
Define a Lyapunov functional
in the following form:
Calculating the time derivative of V along the solution of system (1.4), we obtain
Using
we have
when \(R_{1}<1\). Furthermore,
and thus the maximal invariant set in the set \(\{V'=0\}\) is the singleton \(\{P_{1}\}\). Therefore, \(P_{1}\) is globally attractive.
The characteristic equation associated with the linearization of system (1.4) at \(P_{1}\) is given by
We first consider
When \(\tau_{3}=0\), we have
Assuming \(\tau_{3}>0\) and \(\xi=i\omega\) (\(\omega>0\)) is the purely imaginary root of this equation, then we obtain
So, \(\omega^{2}+d_{4}^{2}=(\gamma\overline{y})^{2}\), namely, \(\omega^{2}=(\gamma\overline{y})^{2}-d_{4}^{2}<0\). Obviously, this is a contradiction. Note that 0 is not the root of the equation. Therefore, all roots of this equation have a negative real part.
Next, we address the following equation:
We rewrite this equation in the following form:
When \(\tau_{1}+\tau_{2}=0\), the equation becomes
where
and \(a_{1}a_{2}-a_{3}>0\). By the Routh-Hurwitz criteria, all roots of (3.9) have negative real parts when \(\tau_{1}+\tau_{2}=0\).
Assume \(\tau_{1}+\tau_{2}>0\), and \(\xi=i\omega\) (\(\omega>0\)) is the purely imaginary root of (3.9). Substituting \(\xi=i\omega\) (\(\omega>0\)) into equation (3.9), we obtain
Separating the real and imaginary parts, we have
Squaring and adding both equations lead to
where
By the Routh-Hurwitz criteria,
has no positive root. So (3.11) has no positive root. Equation (3.9) has no pure imaginary root. Also 0 is not root of equation (3.9), therefore, all roots of equation (3.9) have negative real parts for \(\tau_{1}+\tau_{2}\geq0\). Hence \(P_{1}\) is locally asymptotically stable.
Further, \(P_{1}\) is globally asymptotically stable.
For \(R_{1}>1\), we can find the characteristic equation (3.9) has positive root. Thus \(P_{1}\) is unstable when \(R_{1}>1\). □
Remark 3.4
\(P_{1}\) is globally asymptotically stable without any delays when \(R_{1}<1<R_{0}\). Although incorporating three delays (a immune delay and two intracellular delays), \(P_{1}\) is still globally asymptotically stable. Delays do not destroy the globally asymptotical stability of \(P_{1}\).
3.3 Dynamics when \(R_{1}>1\)
When \(R_{1}>1\), there exists an interior equilibrium \(P_{2}=(x^{*},y^{*},v^{*},z^{*})\), where
The characteristic equation associated with the linearization of system (1.4) at \(P_{2}\) is given by
where
3.3.1 When \(\tau_{1}\geq0\), \(\tau_{2}\geq0\), \(\tau_{3}=0\)
Theorem 3.5
If \(R_{1} >1\), then the equilibrium \(P_{2}\) is globally attractive when
Proof
Let
Define a Lyapunov functional
in the following form:
Calculating the time derivative of V along solution of system (1.4), we obtain
Using
it follows that
This implies that
and thus the maximal invariant set in the set \(\{U'=0\}\) is the singleton \(\{P_{2}\}\). Therefore, \(P_{2}\) is globally attractive. □
Remark 3.6
It is very difficult to analyze the characteristic roots of the characteristic equation (3.12). But we conjecture that all characteristic roots of the characteristic equation (3.12) have negative real parts when \(\tau_{1}>0\), \(\tau_{2}>0\), \(\tau_{3}=0\). Namely, \(P_{2}\) is locally asymptotically stable, and \(P_{2}\) is also globally asymptotically stable when \(\tau_{1}\geq0\), \(\tau_{2}\geq0\), \(\tau_{3}=0\). We find the intracellular delays \(\tau_{1}\) and \(\tau_{2}\) do not destroy global attractability of \(P_{2}\).
3.3.2 When \(\tau_{1}= 0\), \(\tau_{2}= 0\), \(\tau_{3}>0\)
When \(\tau_{1}= 0\), \(\tau_{2}= 0\), \(\tau_{3}>0\), system (1.4) becomes
The characteristic equation of system (3.14) at \(P_{2}\) is given by
When \(\tau_{3}=0\), (3.15) becomes
where
By the Routh-Hurwitz criteria, all roots of this equation have negative real parts. Clearly, 0 is not the root of (3.15).
For \(\tau_{3}>0\), assuming \(\xi=i\omega\) (\(\omega>0\)) is a purely imaginary root of (3.15). It satisfies
Separating the real and imaginary parts, we get
Squaring and adding both above equations lead to
where
Let \(\omega^{2}=s\), we have
Then
Set
Let \(r=s+\frac{p}{4}\), then (3.19) becomes
where \(p_{1}=\frac{q}{2}-\frac{3p^{2}}{16}\), \(q_{1}=\frac{p^{3}}{32}-\frac {pq}{8}+\frac{u}{4}\).
Define
We cite the results in [18] about the existence of positive roots of the fourth-degree polynomial equation, namely, we have the following lemma.
Lemma 3.7
-
(i)
If \(v<0\), then (3.18) has at least one positive root.
-
(ii)
If \(v\geq0\) and \(\Delta\geq0\), then (3.18) has positive roots if and only if \(s_{1} > 0\) and \(F(s_{1})<0\).
-
(iii)
If \(v\geq0\) and \(\Delta< 0\), then (3.18) has positive roots if and only if there exists at least one \(s^{*}\in\{s_{1},s_{2},s_{3}\}\) such that \(s^{*} > 0\) and \(F(s^{*})<0\).
Supposing one of the above three cases in Lemma 3.7 is satisfied, (3.18) has finite positive roots \(s_{1},s_{2},\ldots, s_{k}\), \(k\leq4\). Therefore (3.17) has finite positive roots
For every fixed \(\omega_{i}\) (\(i=1,2,\ldots,k\), \(k\leq4\)), there exists a sequence
where
such that (3.16) holds. Let
Then (3.15) has a pair of purely imaginary roots \(\pm i\omega^{*}\) when \(\tau_{3}=\tau_{3}^{*}\).
After a long and tedious computation, we get the following lemma.
Lemma 3.8
Especially, supposing \(F'((\omega^{*})^{2})\neq0\), then
Remark 3.9
For the nth degree exponential polynomial
we conjecture there are similar equations to (3.21). To the best of our knowledge, it is correct for \(n=1,2,3,4\).
From Lemma 3.8, we can get the following result.
Theorem 3.10
For system (3.14), there exists
such that \(P_{2}\) is asymptotically stable when \(\tau_{3}\in[0,\tau _{3}^{*})\). Furthermore, if \(F'((\omega^{*})^{2})\neq0\) holds, and system (3.14) undergoes a Hopf bifurcation at \(P_{2}\) when \(\tau_{3}=\tau_{3}^{*}\).
Remark 3.11
We find that incorporating an immune delay can destroy the global attractability of \(P_{2}\) on proper conditions when \(R_{1}>1\), and a Hopf bifurcation occurs. That is, a periodic oscillation appears. Stability switches can appear when \(k\geq2\). Those results show immune delay dominates intracellular delays in this class of viral infection models. Those indicate the human immune system has a special effect in virus infection models with a CTLs response, and the human immune system itself is very complicated.
4 Global Hopf bifurcation analysis
Many researchers studied global Hopf bifurcations in their research, for example [19, 20]. In this section, we will investigate the global existence of periodic solutions of system (3.14) by using the global Hopf bifurcation theorem given by Wu [21] when \(R_{1}>1\). So we consider the following system:
Note that we omit the subscript ‘3’ of \(\tau_{3}\) for convenience.
Firstly, we suppose (3.18) has a unique positive root \(s^{*}\) in this section, therefore, \(\omega^{*}=\sqrt{s^{*}}\), and
where
\(\tau^{0}=\min\{\tau^{j},j=0,1,2,\ldots\}\). It is reasonable that we suppose (3.18) has a unique positive root \(s^{*}\). For example, we consider the following case:
when \((3p)^{2}-4\times6 q<0\) and \(v<0\), \(F(s)=0\) has only one positive root. Furthermore, in Section 5, we can choose proper parameters such that \(F(s)=0\) has unique positive root when we carry out numerical simulations.
From Lemma 3.8, we obtain the following lemma.
Lemma 4.1
\(\tau^{j}\), \(\omega^{*}\) are defined as above. The following holds:
Furthermore, if \(\tau\in(0,\tau^{0}]\), then all roots of (3.15) have negative real parts; if \(\tau\in(\tau^{j},\tau^{j+1}]\), \(j=0,1,2,\ldots \) , then (3.15) has exactly \(2(j+1)\) roots with positive real parts.
Lemma 4.2
System (4.1) has no nonconstant periodic solution when \(\tau=0\).
Proof
Theorem 3.5 shows \(P_{2}\) is globally attractive when \(\tau=0\). This lemma follows from the fact that \(P_{2}\) is globally attractive when \(\tau=0\). □
Lemma 4.3
All the nontrivial periodic solutions of (4.1) are positive and uniformly bounded.
Proof
The proof of this lemma can be obtained from Proposition 2.1. □
Lemma 4.4
When \(R_{1}>1\), system (4.1) has no nonconstant periodic solution of period τ. Furthermore, system (4.1) has no nonconstant periodic solution of period \(\frac{\tau}{j}\), \(j=2,3,4,\ldots \) .
Proof
We prove by contradiction. Suppose system (4.1) has a periodic solution of periodic τ, \(W(t)=(x(t),y(t),v(t),z(t))^{T}\), and \(W(t+\tau)=(x(t+\tau),y(t+\tau),v(t+\tau),z(t+\tau))^{T}=W(t)\). So \(W(t)=(x(t),y(t),v(t),z(t))^{T}\) is also τ-periodic solution of the following system:
However, this system has no periodic solutions, which follows from Theorem 3.5. Therefore, system (4.1) has no periodic solution of period τ. □
Let \(W(t)=(x(t),y(t),v(t),z(t))^{T}\), we rewrite system (4.1) as the following functional differential equation:
where \(W_{t}(\theta)=(x(t+\theta),y(t+\theta),v(t+\theta),z(t+\theta ))\in C([-\tau,0],\mathbb{R}_{+}^{4})\), and
Let
It is easy to see the assumptions (A1), (A2), and (A3) in [21] are satisfied.
Note that the periodic solutions are all bounded away from zero, which follows from Lemma 4.2, thus we need not to consider the boundary equilibria \(P_{0}\) and \(P_{1}\).
It is convenient to introduce the following notations:
Let \(C(P_{2}, \tau^{j}, \frac{2\pi}{\omega^{*}})\) denote the connected component of \((P_{2}, \tau^{j},\frac{2\pi}{\omega^{*}})\) in Σ, where \(\tau^{j}\), \(\omega^{*}\) are defined in (4.2).
Now, we are in a position to state the following global Hopf bifurcation results.
Theorem 4.5
When \(R_{1}>1\), for each \(\tau>\tau^{j}\), \(j=1,2,3,\ldots \) , system (4.1) has at least \(j+1\) positive periodic solutions, where \(\tau^{j}\) is defined in (4.2).
Proof
It is obvious that \((P_{2}, \tau^{j},\frac{2\pi}{\omega^{*}})\) are isolated centers. By Lemma 4.1, there exist \(\varepsilon>0\), \(\delta>0\), and a smooth curve \(l: (\tau^{j}-\delta, \tau^{j}+\delta)\rightarrow\mathbb{C}\), such that
for all \(\tau\in[\tau^{j}-\delta, \tau^{j}+\delta]\), where Δ is defined above, and
Let
Clearly, if \(|\tau-\tau^{j}|\leqslant\delta\) and \((u,T)\in\partial \Omega_{\varepsilon}\) such that \(\Delta_{(P_{2}, \tau, T)}(u+\frac{2\pi i}{T})=0\), then \(\tau=\tau^{j}\), \(u=0\), and \(T=\frac {2\pi}{\omega^{*}}\). So (A4) is satisfied in [21] for \(m=1\). Moreover, let
then we see, from \(\operatorname{Re}\xi'(\tau^{j})>0\), that the crossing number is
Using the local Hopf bifurcation theorem in [21], we conclude that the connected component \(C(P_{2}, \tau^{j},\frac{2\pi}{\omega^{*}})\) through \((P_{2}, \tau^{j}, \frac{2\pi}{\omega^{*}})\) in Σ is nonempty. Meanwhile, we have
By Theorem 3.3 in [21], \(C(P_{2}, \tau^{j}, \frac{2\pi}{\omega ^{*}})\) is unbounded.
Lemma 4.3 shows the projection of \(C(P_{2}, \tau^{j}, \frac{2\pi}{\omega ^{*}})\) onto W-space is bounded. Lemma 4.2 implies the projection of \(C(P_{2}, \tau^{j}, \frac{2\pi}{\omega^{*}})\) onto τ-space is bounded below.
From the definition of \(\tau^{j}\) in (4.2), we have
namely,
From Lemma 4.4, we know if
then
if
then
and so on. This shows that the projection of \(C(P_{2}, \tau^{j}, \frac{2\pi}{\omega^{*}})\) onto T-space is bounded. Therefore in order for \(C(P_{2}, \tau^{j}, \frac {2\pi}{\omega^{*}})\) to be unbounded, its projection onto the τ-space must be unbounded. The projection of \(C(P_{2}, \tau^{j}, \frac{2\pi}{\omega^{*}})\) onto the τ-space includes \([\tau^{j},+\infty)\). Note \(\frac{1}{j+1}<\frac{2\pi }{\omega^{*}\tau^{j}}<\frac{1}{j}\), \(j\geq1\), we can see that the connected components \(C(P_{2}, \tau^{j}, \frac{2\pi}{\omega^{*}})\), \(j\geq1\) are disjoint. This shows system (4.1) has at least j positive periodic solutions for each \(\tau>\tau^{j}\). The proof is completed. □
5 Numerical simulations
In this section, we shall carry out some numerical simulations for illustrating our theoretical analysis. As regards the selected parameters in this section, we refer to [15, 22].
First, we consider the following set of parameter values:
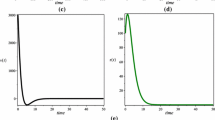
For the above parameter set, \(R_{0}=0.7407<1\), system (1.4) has a unique infection-free equilibrium \(P_{0}=(50,0,0,0)\). Figure 1 shows \(P_{0}\) is globally asymptotically stable when \(R_{0}<1\).

\(\pmb{P_{0}}\) is globally asymptotically stable. Here \(\lambda=10\), \(\beta=0.02\), \(d_{1}=0.2\), \(d_{2}=1.8\), \(d_{3}=1.5\), \(d_{4}=0.5\), \(k=2\), \(\mu =0.2\), \(\gamma=0.2\), \(\tau_{1}=1\), \(\tau_{2}=2\), \(\tau_{3}=3\), and \(R_{0}=0.7407<1\).
Next, we use the following parameters: \(\lambda=14\), \(\beta=0.02\), \(d_{1}=0.2\), \(d_{2}=1.8\), \(d_{3}=1.5\), \(d_{4}=0.5\), \(k=2\), \(\mu =0.2\), \(\gamma=0.2\), \(\tau_{1}=1\), \(\tau_{2}=2\), \(\tau_{3}=3\). For those parameters, \(R_{1}=0.7778<1<R_{0}=1.0370\), system (1.4) has a chronic-infection equilibrium \(P_{1}=(67.5,0.2778,0.3704,0)\). Figure 2 demonstrates the equilibrium \(P_{1}\) is globally asymptotically stable when \(R_{1}<1<R_{0}\).

\(\pmb{P_{1}}\) is globally asymptotically stable. Here \(\lambda=14\), \(\beta=0.02\), \(d_{1}=0.2\), \(d_{2}=1.8\), \(d_{3}=1.5\), \(d_{4}=0.5\), \(k=2\), \(\mu =0.2\), \(\gamma=0.2\), \(\tau_{1}=1\), \(\tau_{2}=2\), \(\tau_{3}=3\), and \(R_{1}=0.7778<1<R_{0}=1.0370\).
In Figures 3 and 4, we adopt the following set of parameter values:
Thus \(R_{1}=2.2222>1\), system (1.4) has the equilibrium \(P_{2}=(150,2.5,3.3333,11)\) and \(F(0)=-0.0176<0\), \(F(s)=0\) has only one positive root \(s^{*}\approx0.2927\). \(\tau_{3}^{*}\approx0.7392\), \(\tau_{3}^{1}\approx 12.3529\). Figure 3 demonstrates that \(P_{2}\) is asymptotically stable when \(R_{1}>1\) and \(\tau_{3}<\tau_{3}^{*}\), where \(\tau_{3}=0.7<\tau_{3}^{*}\). Figure 4 demonstrates that oscillations appear, where \(\tau_{3}=0.8>\tau_{3}^{*}\). Using those parameter values, global Hopf branches diagrams are shown in Figures 5 and 6.

\(\pmb{P_{2}}\) is asymptotically stable. Here \(\lambda=40\), \(\beta=0.02\), \(d_{1}=0.2\), \(d_{2}=1.8\), \(d_{3}=1.5\), \(d_{4}=0.5\), \(k=2\), \(\mu =0.2\), \(\gamma=0.2\), \(\tau_{1}=0\), \(\tau_{2}=0\), \(\tau_{3}=0.7<\tau_{3}^{*}=0.7392\), and \(R_{1}=2.2222>1\).

\(\pmb{P_{2}}\) is not asymptotically stable, and oscillations appear. Here \(\lambda=40\), \(\beta=0.02\), \(d_{1}=0.2\), \(d_{2}=1.8\), \(d_{3}=1.5\), \(d_{4}=0.5\), \(k=2\), \(\mu =0.2\), \(\gamma=0.2\), \(\tau_{1}=0\), \(\tau_{2}=0\), \(\tau_{3}=0.8>\tau_{3}^{*}=0.7392\), and \(R_{1}=2.2222>1\).

Global Hopf branches diagrams. Here \(\lambda=40\), \(\beta =0.02\), \(d_{1}=0.2\), \(d_{2}=1.8\), \(d_{3}=1.5\), \(d_{4}=0.5\), \(k=2\), \(\mu=0.2\), \(\gamma=0.2\), \(\tau _{1}=0\), \(\tau_{2}=0\), \(\tau_{3}^{*}\approx0.7392\), and \(R_{1}=2.2222>1\).

Global Hopf branches diagrams. Here \(\lambda=40\), \(\beta =0.02\), \(d_{1}=0.2\), \(d_{2}=1.8\), \(d_{3}=1.5\), \(d_{4}=0.5\), \(k=2\), \(\mu=0.2\), \(\gamma=0.2\), \(\tau _{1}=0\), \(\tau_{2}=0\), \(\tau_{3}^{1}\approx12.3529\), and \(R_{1}=2.2222>1\).
6 Conclusion
In this paper, we considered a class of virus infection models with three time lags, two intracellular delays and one immune delay. We have carried out a mathematical analysis of the dynamics of the model. We proved that \(P_{0}\) is globally asymptotically stable when \(R_{0} < 1\), and the three delays do not destroy the globally asymptotical stability of \(P_{0}\). \(P_{1}\) is globally asymptotically stable when \(R_{1} < 1 < R_{0}\), and the three delays also do not destroy the globally asymptotical stability of \(P_{1}\). When \(R_{1} > 1\), we found \(P_{2}\) has still global attractability under only incorporating two intracellular delays \(\tau _{1}\) and \(\tau_{2}\). But on only incorporating the immune delay \(\tau _{3}\), \(P_{2}\) can undergo a Hopf bifurcation on proper conditions, furthermore, oscillations and stability switches can appear. The immune delay can destroy the global attractability of \(P_{2}\). Those results show immune delay dominates intracellular delays in some viral infection models, which indicates the human immune system has a special effect in virus infection models with CTLs response, and the human immune system itself is very complicated. People are aware of the complexity of human immune system in medical science, which coincides with our investigation. Finally, we studied the global Hopf bifurcation of the system, and we obtained the global existence of periodic solutions.
References
Culshaw, RV, Ruan, S: A delay-differential equation model of HIV infection of CD4+ T-cells. Math. Biosci. 165, 27-39 (2000)
Gourley, SA, Kuang, Y, Nagy, D: Dynamics of a delay differential model of hepatitis B virus infection. J. Biol. Dyn. 2, 140-153 (2008)
Gomez-Acevedo, H, Li, MY, Jacobson, S: Multi-stability in a model for CTL response to HTLV-I infection and its consequences in HAM/TSP development and prevention. Bull. Math. Biol. 72, 681-696 (2010)
Li, MY, Shu, H: Global dynamics of a mathematical model for HTLV-I infection of CD4+ T cells with delayed CTL response. Nonlinear Anal., Real World Appl. 13, 1080-1092 (2012)
Li, MY, Shu, H: Impact of intracellular delays and target-cell dynamics on in vivo viral infections. SIAM J. Appl. Math. 70, 2434-2448 (2010)
Korobeinikov, A: Global properties of basic virus dynamics models. Bull. Math. Biol. 66, 879-883 (2004)
Sun, X, Wei, J: Global dynamics of a HTLV-I infection model with CTL response. Electron. J. Qual. Theory Differ. Equ. 2013, 40 (2013)
Perelson, A, Neumann, A, Markowitz, M, Leonard, J, Ho, D: HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time. Science 271, 1582-1586 (1996)
Perelson, A, Nelson, P: Mathematical models of HIV dynamics in vivo. SIAM Rev. 41, 3-44 (1999)
Gourley, SA, Kuang, Y, Nagy, D: Dynamics of a delay differential model of hepatitis B virus infection. J. Biol. Dyn. 2, 140-153 (2008)
Xu, R, Ma, Z: An HBV model with diffusion and time delay. J. Theor. Biol. 257, 499-509 (2009)
Wodarz, D: Hepatitis C virus dynamics and pathology: the role of CTL and antibody responses. J. Gen. Virol. 84, 1743-1750 (2003)
Burić, N, Mudrinic, M, Vasović, N: Time delay in a basic model of the immune response. Chaos Solitons Fractals 12, 483-489 (2001)
Nowak, MA, Bangham, CRM: Population dynamics of immune responses to persistent viruses. Science 272, 74-79 (1996)
Zhu, H, Zou, X: Dynamics of a HIV-1 infection model with cell-mediated immune response and intracellular delay. Discrete Contin. Dyn. Syst., Ser. B 12(2), 511-524 (2009)
van den Driessche, P, Watmough, J: Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 180, 29-48 (2002)
Ruan, S, Wei, J: On the zeros of transcendental functions with applications to stability of delay differential equations with two delays. Dyn. Contin. Discrete Impuls. Syst., Ser. A Math. Anal. 10, 863-874 (2003)
Li, X, Wei, J: On the zeros of a fourth degree exponential polynomial with applications to a neural network model with delays. Chaos Solitons Fractals 26, 519-526 (2005)
Wei, J: Bifurcation analysis in a scalar delay differential equation. Nonlinearity 20, 2483-2498 (2007)
Shu, H, Wang, L, Wu, J: Global dynamics of Nicholson’s blowflies equation revisited: onset and termination of nonlinear oscillations. J. Differ. Equ. 255, 2565-2586 (2013)
Wu, J: Symmetric functional differential equations and neural networks with memory. Trans. Am. Math. Soc. 350, 4799-4838 (1998)
Wodarz, D: Hepatitis C virus dynamics and pathology: the role of CTL and antibody responses. J. Gen. Virol. 84, 1743-1750 (2003)
Acknowledgements
The authors would like to thank the anonymous referees and the editor for pertinent suggestions and comments, which led to improvements of our paper. This research is supported by National Natural Science Foundation of China (No. 11371111), Research Fund for the Doctoral Program of Higher Education of China (No. 20122302110044) and Shandong Provincial Natural Science Foundation, China (No. ZR2013AQ023).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
The authors had equal contributions to each part of this paper. All the authors read and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Sun, X., Wei, J. Stability and bifurcation analysis in a viral infection model with delays. Adv Differ Equ 2015, 332 (2015). https://doi.org/10.1186/s13662-015-0664-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-015-0664-7