
Abstract
Background
Ticks are obligate hematophagous ectoparasites involved in transmitting viruses of public health importance. The objective of this work was to identify the **gmen tick virus in hard ticks from the Colombian Caribbean, an arbovirus of importance for public health.
Methods
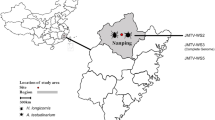
Ticks were collected in rural areas of Córdoba and Cesar, Colombia. Taxonomic identification of ticks was carried out, and pools of 13 individuals were formed. RNA extraction was performed. Library preparation was performed with the MGIEasy kit, and next-generation sequencing (NGS) with MGI equipment. Bioinformatic analyses and taxonomic assignments were performed using the Galaxy platform, and phylogenetic analyses were done using IQ-TREE2.
Results
A total of 766 ticks were collected, of which 87.33% (669/766) were Rhipicephalus microplus, 5.4% (42/766) Dermacentor nitens, 4.2% (32/766) Rhipicephalus linnaei, and 3.0% (23/766) Amblyomma dissimile. Complete and partial segments 1, 2, 3, and 4 of **gmen tick virus (JMTV) were detected in the metatranscriptome of the species R. microplus, D. nitens, and A. dissimile. The JMTVs detected are phylogenetically related to JMTVs detected in Aedes albopictus in France, JMTVs detected in R. microplus in Trinidad and Tobago, JMTVs in R. microplus and A. variegatum in the French Antilles, and JMTVs detected in R. microplus in Colombia. Interestingly, our sequences clustered closely with JMTV detected in humans from Kosovo.
Conclusions
JMTV was detected in R. microplus, D. nitens, and A. dissimile. JMTV could pose a risk to humans. Therefore, it is vital to establish epidemiological surveillance measures to better understand the possible role of JMTV in tropical diseases.
Graphical Abstract

Similar content being viewed by others

Background
Tick-borne infections constitute a significant portion of vector-borne diseases, can generate a considerable disease burden, and present an imminent threat to public health. Ticks act as biological vectors for a wide range of microorganisms, including RNA viruses that can be transmitted to humans and animals [1].
Some examples of pathogenic viruses transmitted by ticks to humans or animals are Crimean−Congo hemorrhagic fever virus (CCHF), Nairobi sheep disease virus, tick-borne encephalitis virus, Omsk hemorrhagic fever, Kyasanur forest disease virus, Powassan virus, Alkhurma hemorrhagic fever virus, severe fever with thrombocytopenia virus, Heartland virus, Colorado fever virus, Bourbon virus, and African swine fever virus (ASF) [2, 3]. In addition, of these pathogens, **gmen tick virus (JMTV) associated with disease in humans has recently been described [4].
JMTV was first detected in 2010 from China in Rhipicephalus microplus [5]. The virus belongs to the **gmen virus group (JMV), and although it remains unclassified by the International Committee on Taxonomy of Viruses (ICTV), it is related to the Flavivirus genus of the Flaviviridae family [5]. The JMV group are single-stranded RNAs in the positive sense (ssRNA+), and unlike typical flaviviruses, it has a segmented genome [6].
The JMTV genome comprises four segments: S1, S2, S3, and S4. The S1 and S3 encode nonstructural proteins (NSPs), while the S2 and S4 encode structural proteins (VPs) [7]. S1 encodes NSP1, an RNA-dependent RNA polymerase homologous to the NS5 protein of flaviviruses; S2 presents an open reading frame (ORF) that encodes a glycoprotein (VP1), S3 encodes NSP2, homologous to the flavivirus NS3-NSP2 complex; S4 presents two ORFs to encodes two proteins, the capsid protein (VP2) and the membrane protein (VP3) [5]. Although segments S1 and S3 are closely related to the NSPs genes (NS3 and NS5) of flaviviruses, the remaining two segments (S2 and S4) are unique to these viruses and lack homologs [5].
JMTV has been identified in several regions of China in ticks and humans [4, 5, 8]. Additionally, its presence has been reported in ticks from various countries including Poland, Georgia, France, French Antilles, Laos People’s Democratic Republic, Brazil, Kenya, Turkey, Japan, Romania, Trinidad and Tobago, and Colombia [9,10,11,12,13,14,15,16,17], as well as mosquitoes (Aedes albopictus) in Italy [18]. JMTV has also been detected in vertebrates, such as the red colobus monkey (Procolobus rufomitratus) in Uganda [19], bats in Cambodia [9], cattle in Brazil [10], and rodents in China [25]. De novo assembly was performed to obtain viral contigs or scaffolds using MEGAHIT v.1.2.9 [26]. Finally, the taxonomic assignment was performed with BLASTx 2.14.1 [27]. All these tools were accessed through the online platform Galaxy [28].
Phylogenetic analysis
The obtained contigs were compared with the GenBank database using BLASTp [27]. Alignments were generated with MAFFT [29] with sequences downloaded from GenBank [30]. Phylogenetic reconstructions were performed in IQ-TREE v2.2.2.6 [31] with the maximum likelihood method [32]; the best-fit amino acid substitution models were obtained using ModelFinder, and the substitution models were chosen according to the Bayesian information criterion [33] and SH-like approximate likelihood ratio test for statistical support [34]. Trees were reconstructed using 1000 bootstraps visualized in iTOL v5 [35] and edited in Inkscape v.1.1 [36].
Results
A total of 766 ticks were collected, of which 87.3% (669/766) were R. microplus, 5.4% (42/766) Dermacentot nitens, 4.2% (32/766) Rhipicephalus linnaei, and 3.0% (23/766) Amblyomma dissimile. In total, 120 pools were generated.
In the metatranscriptome of the species R. microplus and D. nitens collected in the Cesar Department, the complete genome of JMTV was detected. Additionally, complete and partial fragments of segments 1, 2, 3, and 4 that encode the proteins NSP1, VP1, NSP2, VP2, and VP3 of JMTV were detected in R. microplus, D. nitens, and A. dissimile collected in Córdoba and Cesar (Table 1).
The JMTVs detected in this study are phylogenetically related to JMTVs detected in A. albopictus from France, JMTVs detected in R. microplus from Trinidad and Tobago, JMTVs detected in R. microplus and A. variegatum from the French Antilles, and JMTVs detected in Colombia. This relationship was consistent in all constructed trees (Fig. 2). Additionally, when performing BLASTp, the amino acid sequences presented a percentage of identity between 96.23% and 99.38% with the JMTV mentioned above [7, 11, 16].

Phylogenetic reconstructions with amino acids of the four segments of JMTV. A Phylogenetic tree of the NSP1 protein built with 45 sequences (35 downloaded from GenBank and 10 own). B Phylogenetic tree of the VP1 protein built with 37 sequences (28 downloaded from GenBank and 9 own). C Phylogenetic tree of the NSP2 protein built with 35 sequences (28 downloaded from GenBank and 7 own). D VP2 phylogenetic tree built with 35 sequences (25 downloaded from GenBank and 10 own). E VP3 phylogenetic tree built with 39 sequences (33 downloaded from GenBank and 6 own). These five trees were rooted with JMTV (QFR36167) and JMTV (QFR36160). The sequences generated in this study are highlighted in red. Trees were constructed using the substitution models JTTDCMut + G4 for NSP1, FLU + F + G4 for VP1, FLU + G4 for VP2, JTT + G4 for NSP2, and VP3 protein
Additionally, there is a phylogenetic relationship between the JMTVs detected in this study and those detected in humans in Kosovo, given that in trees A, D, and E the sequences of these groups are in the same cluster (Fig. 2, tree A, D, E).
Discussion
JMTV was detected in R. microplus and D. nitens ticks collected from cattle, and A. dissimile collected from iguanas in the Colombian Caribbean (Table 1). This is the third JMTV study on R. microplus and the first virus report on D. nitens and A. dissimile from Colombia.
JMTV was first identified in R. microplus ticks in the **gmen area of Hubei Province, China, in 2010 [5], then in 2012 in Uganda, JMTV was detected by NGS in the plasma of a red colobus monkey (Procolobus rufomitratus) [19]. Between 2013 and 2015, 12 human sera samples collected in Kosovo were analyzed, and patients with CCHF coinfected with JMTV were detected [21]. Between 2014 and 2016, in France, French Antilles, Laos People’s Democratic Republic, and Cambodia, JMTV was detected in Ixodes ricinus, R. microplus, Amblyomma testudinarium, and bats, respectively [9]. In 2017, JMTV was isolated in BME/CTVM23 cells. NGS sequenced the whole genome in Amblyomma javanense collected from Chinese pangolins [4]. That same year, human patients positive for JMV were reported in China. The infection was confirmed by RT-PCR assay in blood in 86 patients with fever, headache, and a history of tick bites. Serological assays showed seroconversion in 19 patients [22]. In Kenya, between 2013 and 2019, ticks of the Amblyomma and Rhipicephalus species were collected, and JMTV was detected by PCR and NGS [11].
Recently, in 2023, in China, JMTV was detected in R. microplus ticks through NGS (MGISEQ-2000), which PCR confirmed, and the complete genome was obtained with Sanger sequencing [8]. Furthermore, they detected JMTV in I. ricinus in Poland and Georgia by NGS (nanopore) and PCR [3].
In Latin America, there are a few reports of JMTV. In Brazil, between 2015 and 2016, complete and partial JMTV genomes were obtained by high-throughput sequencing in 67% (4/6) of tick pools and 14% (5/36) of bovine sera [10]. In Colombia, in the Department of Antioquía, between 2013 and 2018, several JMTV segments were detected by NGS in ticks of the genus Rhiphicephalus [16, 17]. Many of the JMTV reports with NGS carry out a confirmation with PCR; however, in this study, complete genomes were recovered. Thus, confirmation was unnecessary.
JMTV has a worldwide distribution and has been detected in six genera of ticks: Rhipicephalus, Amblyomma, Dermacentor, Haemaphysalis, Hyalomma, and Ixodes, and 26 species [7, 37]. This work provides two new species of ticks where JMTV was detected: D. nitens and A. dissimile. However, R. linnaei was negative despite previous positive reports for this tick species [5].
Interestingly, in the present work, JMTV was detected in D. nitens parasitizing cattle, which suggests transmission to cattle by these tick species, given that there are reports of this virus in cattle from Brazil [10]. This finding indicates a probable risk of transmission to different species of ticks, animals, and humans.
Phylogenetic analyses of five JMTV proteins showed that, in four of the trees obtained (Fig. 2, tree A, C–E), most of the viruses were grouped in two clusters, one of them associated with different species of bats and ticks and the other with humans and different species of ticks.
Interestingly, our sequences of the NSP1, VP2, and VP3 trees were placed in a clade closely related to the JMTVs detected in humans from Kosovo [21]. This is a finding that should alert the medical community in the tropical regions of Colombia.
Given the close linkage between the sole JMTV sequence identified in A. albopictus in Italy and the strain characterized in the current study (Fig. 2, tree A), it is necessary to carry out epidemiological surveillance of JMTV in mosquitoes from tropical areas of Colombia.
Unfortunately, for the phylogenetic analyses, only the VP1 of JMTV previously reported in Colombia could be included given the limited availability of data; however, it presents a high phylogenetic relationship with the JMTV detected in D. nitens from Cesar, reported in the present study.
It is necessary to develop serological tests with the exclusive segments of circulating JMTV to avoid cross-reactions with other endemic flaviviruses and generate tools to contribute to diagnosing febrile syndrome without an apparent focus.
Conclusions
This is the first work in the Colombian Caribbean on ticks of importance in human and animal health. JMTV is one of the emerging viruses with a potential risk of causing epidemiological outbreaks because they are endemic globally. They are closely associated with arthropods and have caused sporadic cases of febrile illness in humans with a history of tick bites. These results are essential to establish surveillance measures to prevent possible outbreaks of these pathogens.
Availability of data and materials
Sequences are available in the Sequence Read Archive (SRA) with accession numbers SRR29433420, SRR29433419, SRR29433418, SRR29433417, SRR29433416, SRR29433415, SRR29433414, SRR29433413, SRR29433412, and SRR29433411.
References
Rochlin I, Toledo A. Emerging tick-borne pathogens of public health importance: a mini-review. J Med Microbiol. 2020;69:781–91. https://doi.org/10.1099/jmm.0.001206.
Mansfield KL, Jizhou L, Phipps LP, Johnson N. Emerging tick-borne viruses in the twenty-first century. Front Cell Infect Microbiol. 2017;7:298. https://doi.org/10.3389/fcimb.2017.00298.
Ergunay K, Bourke BP, Reinbold-Wasson DD, et al. The expanding range of emerging tick-borne viruses in Eastern Europe and the Black Sea Region. Sci Rep. 2023;13:19824. https://doi.org/10.1038/s41598-023-46879-2.
Jia N, Liu HB, Ni XB, et al. Emergence of human infection with **gmen tick virus in China: a retrospective study. EBioMedicine. 2019;43:317–24. https://doi.org/10.1016/j.ebiom.2019.04.004.
Qin XC, Shi M, Tian JH, et al. A tick-borne segmented RNA virus contains genome segments derived from unsegmented viral ancestors. Proc Natl Acad Sci U S A. 2014;111:6744–9. https://doi.org/10.1073/pnas.1324194111.
Simmonds P, Becher P, Bukh J, et al. ICTV virus taxonomy profile: flaviviridae. J Gen Virol. 2017;98:2–3. https://doi.org/10.1099/jgv.0.000672.
Wu Z, Zhang M, Zhang Y, et al. **gmen tick virus: an emerging arbovirus with a global threat. mSphere. 2023;8:e0028123. https://doi.org/10.1128/msphere.00281-23.
Li W, Li R, Tang X, et al. Genomics evolution of **gmen viruses associated with ticks and vertebrates. Genomics. 2023;115:110734. https://doi.org/10.1016/j.ygeno.2023.110734.
Temmam S, Bigot T, Chrétien D, et al. Insights into the host range, genetic diversity, and geographical distribution of **gmenviruses. mSphere. 2019;4:e00645-e719. https://doi.org/10.1128/mSphere.00645-19.
Souza WM, Fumagalli MJ, Torres Carrasco AO, et al. Viral diversity of Rhipicephalus microplus parasitizing cattle in southern Brazil. Sci Rep. 2018;8:16315. https://doi.org/10.1038/s41598-018-34630-1.
Ergunay K, Mutinda M, Bourke B, et al. Metagenomic investigation of ticks from Kenyan wildlife reveals diverse microbial pathogens and new country pathogen records. Front Microbiol. 2022;13:932224. https://doi.org/10.3389/fmicb.2022.932224.
Dinçer E, Hacıoğlu S, Kar S, et al. Survey and characterization of **gmen Tick Virus variants. Viruses. 2019;11:1071. https://doi.org/10.3390/v11111071.
Sameroff S, Tokarz R, Charles RA, et al. Viral diversity of tick species parasitizing cattle and dogs in Trinidad and Tobago. Sci Rep. 2019;9:10421. https://doi.org/10.1038/s41598-019-46914-1.
Kobayashi D, Kuwata R, Kimura T, et al. Detection of **gmenviruses in Japan with evidence of vertical transmission in ticks. Viruses. 2021;13:2547. https://doi.org/10.3390/v13122547.
Bratuleanu BE, Temmam S, Chrétien D, et al. The virome of Rhipicephalus, Dermacentor and Haemaphysalis ticks from Eastern Romania includes novel viruses with potential relevance for public health. Transbound Emerg Dis. 2022;69:1387–403. https://doi.org/10.1111/tbed.14105.
Gómez GF, Isaza JP, Segura JA, Alzate JF, Gutiérrez LA. Metatranscriptomic virome assessment of Rhipicephalus microplus from Colombia. Ticks Tick Borne Dis. 2020;11:101426. https://doi.org/10.1016/j.ttbdis.2020.101426.
Orozco Orozco M, Gómez GF, Alzate JF, Isaza JP, Gutiérrez LA. Virome analysis of three Ixodidae ticks species from Colombia: a potential strategy for discovering and surveying tick-borne viruses. Infect Genet Evol. 2021;96:105103. https://doi.org/10.1016/j.meegid.2021.105103.
Parry R, James ME, Asgari S. Uncovering the worldwide diversity and evolution of the virome of the mosquitoes Aedes aegypti and Aedes albopictus. Microorganisms. 2021;9:1653. https://doi.org/10.3390/microorganisms9081653.
Ladner JT, Wiley MR, Beitzel B, et al. A multicomponent animal virus isolated from mosquitoes. Cell Host Microbe. 2016;20:357–67. https://doi.org/10.1016/j.chom.2016.07.011.
Yu ZM, Chen JT, Qin J, et al. Identification and characterization of **gmen tick virus in rodents from **njiang, China. Infect Genet Evol. 2020;84:104411. https://doi.org/10.1016/j.meegid.2020.104411.
Emmerich P, Jakupi X, von Possel R, et al. Viral metagenomics, genetic and evolutionary characteristics of Crimean-Congo hemorrhagic fever Orthonairovirus in humans, Kosovo. Infect Genet Evol. 2018;65:6–11. https://doi.org/10.1016/j.meegid.2018.07.010.
Wang ZD, Wang B, Wei F, et al. A new segmented virus associated with human febrile illness in China. N Engl J Med. 2019;380:2116–25. https://doi.org/10.1056/NEJMoa1805068.
Dantas-Torres F, Fernandes Martins T, Muñoz-Leal S, Onofrio VC, Barros-Battesti DM. Ticks (Ixodida: Argasidae, Ixodidae) of Brazil: updated species checklist and taxonomic keys. Ticks Tick Borne Dis. 2019;10:101252. https://doi.org/10.1016/j.ttbdis.2019.06.012.
Chen S, Zhou Y, Chen Y, Gu J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 2018;34:i884–90. https://doi.org/10.1093/bioinformatics/bty560.
Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nat Methods. 2012;9:357–9. https://doi.org/10.1038/nmeth.1923.
Li D, Liu CM, Luo R, Sadakane K, Lam TW. MEGAHIT: an ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics. 2015;31:1674–6. https://doi.org/10.1093/bioinformatics/btv033.
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–10. https://doi.org/10.1016/S0022-2836(05)80360-2.
Broom BM, Ryan MC, Brown RE, et al. A galaxy implementation of next-generation clustered heatmaps for interactive exploration of molecular profiling data. Cancer Res. 2017;77:e23–6. https://doi.org/10.1158/0008-5472.CAN-17-0318.
Katoh K, Misawa K, Kuma K, Miyata T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002;30:3059–66. https://doi.org/10.1093/nar/gkf436.
Benson DA, Karsch-Mizrachi I, Lipman DJ, Ostell J, Wheeler DL. GenBank: update. Nucleic Acids Res. 2004;32:D23–6. https://doi.org/10.1093/nar/gkh045.
Nguyen LT, Schmidt HA, von Haeseler A, Minh BQ. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol Biol Evol. 2015;32:268–74. https://doi.org/10.1093/molbev/msu300.
Felsenstein J. Evolutionary trees from DNA sequences: a maximum likelihood approach. J Mol Evol. 1981;17:368–76. https://doi.org/10.1007/BF01734359.
Kalyaanamoorthy S, Minh BQ, Wong TKF, von Haeseler A, Jermiin LS. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat Methods. 2017;14:587–9. https://doi.org/10.1038/nmeth.4285.
Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol. 2010;59:307–21. https://doi.org/10.1093/sysbio/syq010.
Letunic I, Bork P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021;49:W293–6. https://doi.org/10.1093/nar/gkab301.
Yuan S, Chan HCS, Filipek S, Vogel H. PyMOL and inkscape bridge the data and the data visualization. Structure. 2016;24:2041–2. https://doi.org/10.1016/j.str.2016.11.012.
Shi M, Lin XD, Vasilakis N, et al. Divergent viruses discovered in arthropods and vertebrates revise the evolutionary history of the flaviviridae and related viruses. J Virol. 2015;90:659–69. https://doi.org/10.1128/JVI.02036-15.
Acknowledgements
We thank the University of Córdoba, where the project was carried out, and R.T. was funded by ANID becas/scholarship program/doctorado nacional/ 2020–21200182. S.M.L. was funded by Fondecyt iniciación #11220177.
Funding
This research was financed by the science, technology and innovation fund, BPIN 2020000100322. Research project: “Fortalecimiento de las capacidades de investigación con relación a las enfermedades transmitidas por vectores de las universidades de Córdoba y Cesar 2020 -2023 en Córdoba, Cesar”.
Author information
Authors and Affiliations
Contributions
Y.L. and Sa.Ma. designed the initial study. Y.L., A.C., Ye.Lo., K.G., A.G., L.R.R., G.O., L.R., P.F., and H.C. carried out the fieldwork. Y.L., Ye.Lo., A.G., D.E., C.M., B.G., R.H., A.O., E.G., M.A., J.M., V.C., V.T., G.A., L.F., and C.G. performed DNA extraction, PCRs, and sequencing. Y.L., Se.Mu., and R.T. implemented the phylogenetic analyzes. Y.L., R.T., and R.R. performed bioinformatics analyses. Y.L. and Sa.Ma. wrote the first draft of the manuscript. All authors contributed to the interpretation and review of the data.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The capture of opossums was carried out under permits of the National Authority of Environmental Licenses [ANLA], resolution No. 00914.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
López, Y., Thomas, R., Muñoz-Leal, S. et al. Hard ticks (Ixodida: Ixodidae) in the Colombian Caribbean harbor the **gmen tick virus: an emerging arbovirus of public health concern. Parasites Vectors 17, 268 (2024). https://doi.org/10.1186/s13071-024-06362-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-024-06362-x