
Abstract
The Asian tiger mosquito Aedes albopictus is one of the most invasive species of mosquito. The prevalence of its apicomplexan gregarine parasite Ascogregarina taiwanensis is high in natural populations across both temperate and tropical regions. However, the parasite’s oocysts cannot colonize the insect host during winter, when the mosquito lays diapausing eggs. It is therefore unclear if the parasite can survive outside of its insect host during the cold season in temperate regions. Oocysts stored for 1 month at a low temperature (representative of the temperatures that occur during periods of mosquito diapause) were as infectious as fresh oocysts, but those stored for the same period of time at a higher temperature (representative of the temperatures that occur during periods of mosquito activity) were uninfectious. We therefore suggest that the parasite has evolved traits that maximize its maintenance during periods of mosquito dormancy, while traits that would enable its long term survival during periods of mosquito activity have not been selected for.
Similar content being viewed by others


The phylum Apicomplexa, which comprises heterotrophic animal parasites, evolved from photosynthetic marine algae [1]. The Apicomplexa have been intensively studied because important human pathogens belong to this phylum, such as Toxoplasma and Plasmodium. Gregarine parasites (Gregarinasina) are of great interest from an evolutionary perspective since their lineage is the most ancient of the Apicomplexa. They are highly diverse and infect a large number of invertebrates across marine, freshwater and terrestrial ecosystems [2]. Their interactions with their hosts are complex, and range from mutualism to parasitism.
In mosquitoes, only one gregarine genus has been reported, namely Ascogregarina [3]. Species of the genus Ascogregarina mainly colonize Aedes spp., but also colonize Tripteroides dofleini and Armigeres subalbatus, and show high host specificity. They are often considered weakly pathogenic to their natural hosts but harmful to other species [4]. The Asian tiger mosquito Aedes albopictus is one of the most invasive species of mosquitoes in the world [5]. Originating in Asia, it has efficiently colonized every continent, except for Antarctica, over the past century [6]. Its ability to lay diapausing eggs which can remain dormant during the winter has enabled it to survive low temperatures and colonize temperate regions [7]. Its gregarine parasite Ascogregarina taiwanensis has a remarkable prevalence in both native and introduced populations [8,9,10,11], although some exceptions have been reported [12]. The parasites are released as oocysts (a means of environmental resistance) by adult mosquitoes when these emerge, die or defecate in freshwater [3]. The oocysts infect 1st instar mosquito larvae through ingestion in their freshwater habitat, and the biological cycle of the parasite, which occurs within the digestive tract of the insect (i.e. the gut lumen, intestinal epithelium and lumen of the Malpighian tubules) is regulated by the development of the mosquito [13, 14]. It is, however, unclear whether the parasite can remain infectious during the winter period. In this study, we aimed to test the infective ability of oocysts under winter and summer conditions.
We generated a line of mosquitoes from a focal population collected in September 2017 in Villeurbanne and Pierre-Bénite (France). The mosquitoes were reared under laboratory conditions at 28 °C. The presence of A. taiwanensis in the line was confirmed by diagnostic polymerase chain reaction following the protocol described by Reyes-Villanueva et al. [15], and by microscopic observation of the gut in pupae and Malpighian tubules in adults. The parasite was maintained in the line over 22 generations by (i) sprinkling the crushed bodies of infected adults over the water in the containers holding the larvae, (ii) not storing eggs, and (iii) never changing the water in the containers before the 2nd instar stage. A non-infected line was generated from the same focal population of mosquitoes and maintained for 18 generations. This was achieved by (i) treating F1 mosquito eggs with griseofulvin (1 mg ml−1), a fungicide that shows antiparasitic activity against gregarines [16]; and (ii) changing the water in which the 1st instar larvae were maintained. We verified the absence of the parasite by microscopy.
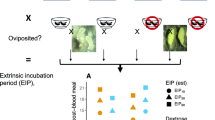
To test our hypothesis, we exposed 100 uninfected mosquito larvae to fresh oocysts or oocysts stored for 1 month at a low temperature (4 °C), or oocysts stored for 1 month at a higher temperature (26 °C). The oocysts were obtained by crushing 100 infected adult mosquitoes in 45-ml sterile water in a Dounce tissue grinder tube (mean ± SD = 2.56 ± 2.14 × 106 oocysts). The contents of each tube were then separated into three 15-ml tubes. The contents of one of these tubes was used to immediately re-infect mosquitoes, while the contents of the other tubes were stored at 4 °C or 26 °C. The experiment was replicated for three different mosquito batches per condition. A negative control consisting of uninfected mosquitoes was prepared to ensure that there was no oocyst material in the test population. We then counted the number of oocysts that had colonized the adult individuals that emerged under each of the test conditions. Survival to immature stages was slightly higher for uninfected individuals (mean ± SD = 87.67 ± 3.29%), individuals colonized with parasites stored at 26 °C (mean ± SD = 93.67 ± 0.58%) and individuals immediately reinfected with parasites (mean ± SD = 85.67 ± 5.03%) compared to those reinfected with parasites stored at 4 °C (mean ± SD = 75.33 ± 3.06%).
Individual mosquitoes were crushed with a sterile pestle in 100 µl of sterile water (Gibco, France). Oocysts were counted in 10 haemocytometer cells per mosquito at ×400 magnification under a microscope (Leica, Germany). Number of oocysts per mosquito was calculated using the following equation: \(n({\text{oocysts/mosquito}})={\text{Water volume}} \times {{\frac{\text{Average number of oocyst per cell}}{{\text{Cell volume}}}}}\). Oocyst counts were determined for a total of 10 males and 10 females per condition and per replicate (i.e. a total of 240 individuals).
Statistical analyses were performed with R software v. 4.0.2 (R Development Core Team). The data were plotted using the ggplot2 and ggpubr R packages.
Interestingly, no oocysts were retrieved from the majority of mosquitoes infected with oocysts stored at 26 °C (Fig. 1), or from the uninfected individuals. These results differed from those for mosquitoes infected with fresh oocysts or oocysts stored at 4 °C. These observations were consistent across sexes and replicate batches. Since the data were zero-inflated, the oocyst count data were transformed into presence–absence for the analysis, where presence indicated that an individual was parasitized and absence indicated that it was not. Variation in the oocyst infection rate between replicate batches, male and female mosquitoes, and experimental conditions were modelled with a generalized linear model with a binomial distribution using the brglm2 R package. The influence of each explanatory variable on the response variables was tested by type II Wald χ2 with the car R package. This showed that oocyst infection rate was significantly influenced by the experimental condition (Wald χ2, P < 2 × 10–16) and by the sex (Wald χ2, P = 0.02) of the mosquito in interaction with the replicate batch (Wald χ2, P = 0.02; Table 1). Therefore, independent analyses were repeated for each sex separately. Oocyst infection rate variation among mosquitoes of both sexes was significantly affected by the experimental conditions tested (Wald χ2, females, P < 2 × 10–16; males, P = 7.98 × 10–15), and differences were also observed between batches for males (Wald χ2, P = 3.78 × 10–4; Table 1). After averaging the values of the batch replicates, post hoc comparisons of pairwise experimental conditions were performed using Tukey honest significant difference (HSD) test with the emmeans R package. These pairwise comparisons indicated that females and males showed consistent patterns despite the batch effect observed for the males (Table 2). The pairwise comparisons showed that oocytes stored in water under a low temperature were still infectious, and that infection rates under this condition did not significantly differ from those in which fresh oocysts were used to infect mosquitoes (Tukey HSD, females, P = 0.3; males, P = 0.98). In contrast, rates of infection with oocysts stored in 26 °C water were consistently lower than those with fresh oocysts or oocysts stored at 4 °C (Tukey HSD, P < 10–2 for each comparison across both sexes), and did not significantly differ from those of uninfected control individuals (Tukey HSD, males, P = 0.99; females, P = 0.99).

Number of oocysts per adult mosquito across the experimental conditions. Females (blue) and males (yellow) were tested across three independent replicate batches. The violin plots show the log10 of total oocyst number + 1 across the experimental conditions. In the Fresh oocysts treatment, infected mosquitoes were crushed into sterile water which was immediately used to seed the water in the tray containing the unparasitized 1st instar larvae. Non-infected mosquitoes were unparasitized and used as a control to confirm that no residual parasites had colonized the test population. In the Parasite kept 1 month at 26 °C or 4 °C treatments, infected mosquitoes were crushed in sterile water and stored for 1 month at 26 °C or at 4 °C, respectively before being seeded to tray containing unparasitized 1st instar larvae. Larvae were reared to adult mosquitoes before parasitism measurement
In temperate regions, the active period of mosquitoes ranges from April/May to October/November, while during the rest of the year mosquitoes lay diapausing eggs that remain dormant until the following active period [17]. Ascogregarina taiwanensis colonizes every developmental stage of mosquitoes except for the egg stage [13]. Therefore, it cannot infect its mosquito hosts during the period of dormancy, and thus persists as a free-living oocyst until mosquito eggs hatch. However, one can hypothesize that the parasite may infect its host during the inactive period if life stages other than eggs are maintained during the winter. Adult hibernation has been reported in some mosquito species [18] but not, to the best of our knowledge, in Ae. albopictus. Mosquitoes can pursue their cycle over the winter if they find warm refuges. However, Ae. albopictus mostly colonizes open habitats [19]. The overwintering of mosquitoes in greenhouses have been reported in the Netherlands, but this remains a solitary example of this behaviour [20]. Concrete tanks were used as winter refuges for larvae and pupae in northern Vietnam [21], but the winter temperatures in this subtropical region average 14 °C, which is sufficiently high to allow the slow but complete development of the mosquito. In contrast, open bodies of water in temperate regions can reach lower temperatures in winter.
The prevalence of Ascogregarina taiwanensis in Aedes albopictus can reach 100% in temperate regions [11]. This finding is inconsistent with low survival of the parasite in mosquito refuges during the winter. However, based on our results, we can propose an explanation for this discrepancy. Since the oocysts showed no decrease in infectivity when they were stored in water at a low temperature (at least 1 month at 4 °C), we suggest that the parasite is adapted to survive outside of its host during the latter’s period of inactivity. It is interesting to note that the survival of the parasite at the higher temperature tested, which is representative of temperatures that occur during the period of mosquito activity, was poor. A possible explanation for this is that the parasite is more likely to encounter a mosquito host in warmer periods, and thus there may be less selective pressure for resistance in oocysts at temperatures prevalent during the mosquito’s active season.
In conclusion, our study provides an example of parallel phenology which may explain the success of a neglected, but widespread, invasive parasitic species in infecting its mosquito host. This profile resembles that of Cryptosporidium, an apicomplexan parasite of mammals that is transmitted in water. In that model, infectivity remained stable for at least 12 weeks when the oocysts were stored in water at 4 °C, but dropped dramatically after 6 weeks when the oocysts were stored at 25 °C [22]. This type of pattern could have epidemiological consequences, since transmission rates of viruses and filarial nematodes in Aedes mosquitoes have been shown to vary according to the degree to which the mosquitoes have been parasitized by Ascogregarina spp. [23, 24].
Availability of data and materials
The datasets supporting the conclusions of this article are included within the article and in its additional file.
References.
Woo YH, Ansari H, Otto TD, Klinger CM, Kolisko M, Michálek J, et al. Chromerid genomes reveal the evolutionary path from photosynthetic algae to obligate intracellular parasites. eLife. 2015;4:e06974.
Rueckert S, Betts EL, Tsaousis AD. The symbiotic spectrum: where do the gregarines fit? Trends Parasitol. 2019;35:687–94.
Lantova L, Volf P. Mosquito and sand fly gregarines of the genus Ascogregarina and Psychodiella (Apicomplexa: Eugregarinorida, Aseptatorina)—overview of their taxonomy, life cycle, host specificity and pathogenicity. Infect Genet Evol. 2014;28:616–27.
Aliabadi BW, Juliano SA. Escape from gregarine parasites affects the competitive interactions of an invasive mosquito. Biol Invasions. 2002;4:283–97.
Bonizzoni M, Gasperi G, Chen X, James AA. The invasive mosquito species Aedes albopictus: current knowledge and future perspectives. Trends Parasitol. 2013;29:460–8.
Goubert C, Minard G, Vieira C, Boulesteix M. Population genetics of the Asian tiger mosquito Aedes albopictus, an invasive vector of human diseases. Heredity. 2016;117:125–34.
Lacour G, Chanaud L, L’Ambert G, Hance T. Seasonal synchronization of diapause phases in Aedes albopictus (Diptera: Culicidae). PLoS ONE. 2015;10:e0145311.
Albicócco AP, Vezzani D. Further study on Ascogregarina culicis in temperate Argentina: prevalence and intensity in Aedes aegypti larvae and pupae. J Invertebr Pathol. 2009;101:210–4.
Garcia JJ, Fukuda T, Becnel JJ. Seasonality, prevalence and pathogenicity of the gregarine Ascogregarina taiwanensis (Apicomplexa: Lecudinidae) in mosquitoes from Florida. J Am Mosq Control Assoc. 1994;10:413–8.
Mayilsamy M, Arunachalam N, Rajaiah P, Paulraj P. Prevalence of Ascogregarina spp. in the container breeding Aedes albopictus from Chikungunya fever affected areas of Kerala State. India J Commun Dis. 2010;42:157–9.
Munstermann L, Wesson D. First record of Ascogregarina taiwanensis (Apicomplexa: Lecudinidae) in North American Aedes albopictus. J Am Mosq Control Assoc. 1990;6:235–43.
Alonso AC, Stein M, Micieli MV. Presence of Ascogregarina culicis and Ascogregarina sp. in natural sympatric populations of Aedes aegypti and Ae. albopictus (Diptera: Culicidae) in Argentina. An Acad Bras Cienc. 2021;93:e20200058.
Chen WJ. The life cycle of Ascogregarina taiwanensis (Apicomplexa: Lecudinidae). Parasitol Today. 1999;15:153–6.
Tseng M. Sex-specific response of a mosquito to parasites and crowding. Proc R Soc B Biol Sci. 2004;271:S186–8.
Reyes-Villanueva F, Becnel JJ, Butler JF. Morphological traits for distinguishing extracellular gamonts of Ascogregarina culicis and Ascogregarina taiwanensis in Aedes aegypti and Aedes albopictus. J Invertebr Pathol. 2001;77:227–9.
Johny S, Merisko A, Whitman DW. Efficacy of eleven antimicrobials against a gregarine parasite (Apicomplexa: Protozoa). Ann Clin Microbiol Antimicrob. 2007;6:15.
Charrel RN, de Lamballerie X, Raoult D. Seasonality of mosquitoes and chikungunya in Italy. Lancet Infect Dis. 2008;8:5–6.
Dörge DD, Cunze S, Schleifenbaum H, Zaenker S, Klimpel S. An investigation of hibernating members from the Culex pipiens complex (Diptera, Culicidae) in subterranean habitats of central Germany. Sci Rep. 2020;10:10276.
Hafeez F, Naeem-Ullah U, Akram W, Arshad M, Iftikhar A, Naeem A, et al. Habitat characterization of Aedes albopictus. Int J Trop Insect Sci. 2022;42:1555–60.
Scholte E-J, Jacobs F, Linton Y-M, Dijkstra E, Fransen J, Takken W. First record of Aedes (Stegomyia) albopictus in the Netherlands. Eur Mosq Bull. 2007;22:5–9.
Tsunoda T, Cuong TC, Dong TD, Yen NT, Le NH, Phong TV, et al. Winter refuge for Aedes aegypti and Ae. albopictus mosquitoes in Hanoi during winter. PLoS ONE. 2014;9:e95606.
King BJ, Keegan AR, Monis PT, Saint CP. Environmental temperature controls Cryptosporidium oocyst metabolic rate and associated retention of infectivity. Appl Environ Microbiol. 2005;71:3848–57.
Comiskey NM, Lowrie RC, Wesson DM. Effect of nutrient levels and Ascogregarina taiwanensis (Apicomplexa: Lecudinidae) infections on the vector competence of Aedes albopictus (Diptera: Culicidae) for Dirofilaria immitis (Filarioidea: Onchocercidae). J Med Entomol. 1999;36:55–61.
Singh D, Yadav P, Gokhale M, Barde P, Narayan N, Mishra A, et al. Role of gregarine parasite Ascogregarina culicis (Apicomplexa: Lecudinidae) in the maintenance of chikungunya virus in vector mosquito. J Eukaryot Microbiol. 2003;50:379–82.
Acknowledgements
We thank the Master’s in Microbiology Programme and the Ecology, Evolution, Microbiology and Modelling Doctoral School of the University of Lyon, to which both CDC and MG are associated.
Funding
This study was funded by the EC2CO Project (grant INTERASCO) and the ANSES Project (grant AS-CONTROL PNR EST 2019-1-095).
Author information
Authors and Affiliations
Contributions
EM, LV and GM designed the experiments. EM and LV conducted the experiments with the help of CDC, MG and GM. GM performed the statistical analysis. GM wrote the first draft of the manuscript and all the co-authors reviewed it. All the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Experiment on insects are exempted from ethics approval. To produce the mosquito eggs, the adult females were fed on anaesthetized mice (Mus musculus). This protocol was approved by an ethical committee and validated by the French Ministry for Higher Education and Research (APAFIS#31807-2021052715018315 v2).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Martin, E., Vallon, L., Da Silva Carvalho, C. et al. Gregarine parasites are adapted to mosquito winter diapause. Parasites Vectors 15, 249 (2022). https://doi.org/10.1186/s13071-022-05365-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-022-05365-w