
Abstract
Serotonin (5-HT) receptors are increasingly recognized as major targets for cognitive enhancement in schizophrenia. Several lines of evidence suggest a pathophysiological role for glutamate NMDA receptors in the prefrontal cortex in schizophrenia and associated disorders in attention and executive functioning. We investigated how the interactions between 5-HT1A and 5-HT2A and glutamate NMDA receptor mechanisms in the medial prefrontal cortex (mPFC) contribute to the control of different aspects of attentional performance. Rats were trained on a five-choice serial reaction time (5-CSRT) task, which provides indices of attentional functioning (percentage of correct responses), executive control (measured by anticipatory and perseverative responses), and speed. The competitive NMDA receptor antagonist CPP (50 ng/side) was infused directly into the mPFC 5 min after infusion of either 8-OH-DPAT (30 and 100 ng/side) or M100907 (100 and 300 ng/side) into the same brain area. Impairments in attentional functioning induced by CPP were completely abolished by both doses of 8-OH-DPAT or M100907. In addition, M100907 abolished the CPP-induced anticipatory responding but had no effects on perseverative over-responding, while 8-OH-DPAT reduced the perseverative over-responding but had no effects on anticipatory responding induced by CPP. The selective 5-HT1A receptor antagonist WAY100635 (30 ng/side) antagonized the effects of 8-OH-DPAT (100 ng/side). 8-OH-DPAT at 30 ng/side reduced the latency of correct responses in controls and CPP-injected rats and lowered the percentage of omissions in CPP-injected rats. The data show that 5-HT1A and 5-HT2A receptors in the mPFC exert opposing actions on attentional functioning and demonstrate a dissociable contribution of 5-HT1A and 5-HT2A receptors in the mPFC to different aspects of executive control such as impulsivity and compulsive perseveration.
Similar content being viewed by others

INTRODUCTION
Cognitive deficits, including attention disorders and deficits in working memory and executive functions, are a central feature of schizophrenia (Braff, 1993; Frith, 1987; Kay and Sevy, 1990). Dysregulation of prefrontal glutamate NMDA receptor function may contribute to the pathophysiology of schizophrenia and to cognitive dysfunctions (Goff and Coyle, 2001; Konradi and Heckers, 2003). In normal humans, NMDA receptor antagonists cause cognitive deficits analogous to those in schizophrenic patients (Javitt and Zukin, 1991; Krystal et al, 1994). In experimental animals, administration of NMDA receptor antagonists systemically or locally into the medial prefrontal cortex (mPFC) dysregulates the firing and bursting activity of pyramidal neurons (Jackson et al, 2004) and cortical glutamate release (Ceglia et al, 2004; Moghaddam and Adams, 1998; Verma and Moghaddam, 1996), and produce a range of behavioral impairments reminiscent of frontal lobe dysfunction (Carli et al, 2004; Egerton et al, 2005; Higgins et al, 2003a; Jentsch and Roth, 1999; Le Pen et al, 2003; Murphy et al, 2005; Rodefer et al, 2005).
Serotonin (5-HT) receptors are increasingly recognized as major targets for cognitive enhancement in schizophrenia (Meltzer et al, 2003). Indeed, the superior efficacy of current atypical antipsychotics on aspects of attention such as vigilance and to some extent executive functioning in patients with schizophrenia may be due partly, to their direct or indirect effects on serotonin 5-HT2A and 5-HT1A receptors (Harvey et al, 2003a, 2003b; Harvey and Keefe, 2001; Meltzer et al, 2003; Meltzer and McGurk, 1999).
The mPFC receives extensive 5-HT innervation from the dorsal (DR) and median raphè (MR) nuclei and contains several 5-HT receptors, with a particular abundance of 5-HT1A and 5-HT2A subtypes (Azmitia and Segal, 1978; Barnes and Sharp, 1999; Blue et al, 1988; Steinbusch, 1984). These receptors are highly colocalized (80%) in nearly half of the glutamatergic pyramidal neurons of the PFC and they are also present in about 25% of GABA interneurons (Santana et al, 2004). The activation of 5-HT1A receptors in the PFC inhibits the neuronal output of pyramidal neurons by activation of a hyperpolarizing potassium current; 5-HT2A facilitates output through a reduction of potassium conductance, reduction of the after-hyperpolarization, and increase in excitatory postsynaptic currents and discharge rate (Aghajanian and Marek, 1997; Andrade and Nicoll, 1987; Araneda and Andrade, 1991; Tanaka and North, 1993). Inhibitory effects of 5-HT2A receptors on pyramidal cell activity have also been reported (Ashby et al, 1994; Puig et al, 2003; Zhou and Hablitz, 1999).
The 5-HT1A and 5-HT2A receptors have an important role in cognitive processes related to frontal lobe functions such as attention, executive function, and working memory (Carli and Samanin, 1992, 2000; Koskinen et al, 2000; Williams et al, 2002; Winstanley et al, 2003). While activation of 5-HT1A and 5-HT2A receptors in the mPFC has opposite effects on attentional functioning (Winstanley et al, 2003), the 5-HT2A but not 5-HT1A receptors appear particularly involved in processes controlling response inhibition (Koskinen et al, 2000; Winstanley et al, 2003). In addition, a selective antagonist at 5-HT2A receptors, M100907, reverses the effects of systemic or intra-mPFC NMDA antagonists on attentional functioning and some aspects of executive function (Carli et al, 2004; Higgins et al, 2003b).
The five-choice serial reaction time (5-CSRT) task used to assess attentional and executive functions in rodents is functionally analogous to the continuous performance test in humans (Carli et al, 2004; Higgins et al, 2003b; Robbins, 2002) and schizophrenic subjects reliably show impairments in the task, compared to controls (Kurtz et al, 2001; Orzack and Kornetsky, 1966). The 5-CSRT task taxes attentional capacity, as indicated by accuracy of correctly reporting the location of a brief visual stimulus, in addition to inhibitory response control related to executive attentional processes that permit accurate response selection in the face of distraction and interference (Robbins, 1998; Shallice, 1982).
Two different aspects of inhibitory response control may be indexed (Robbins, 2002). First, anticipatory responding represents a failure of response inhibition during the orienting behavior in anticipation of the target stimulus, which occur in anticipation of visual stimulus. This behavior has been operationally defined as ‘impulsive’ after Soubrié (1986), although it is recognized that impulsivity, as it occurs clinically, may also take other forms. Second, perseverative responding represents a failure to stop an apparently aimless ‘compulsive’ repetition by which rats continue to respond in stimulus holes after a correct response has been made. This perseverative behavior is akin to that reported in the frontal-lobe and schizophrenic patients (Owen et al, 1993 #76; Lyon and Gerlach, 1988 #167; Goldberg and Weinberger, 1994 #166), and animals with PFC lesions (Chudasama et al, 2003; Mishkin, 1964). Deficits of perseveration and impulsivity and inattention may all contribute to the dys-executive syndromes such as in schizophrenia as well as in obsessive-compulsive disorder, ADHD, and Parkinson's disease (Aron et al, 2003; Baxter, 1990; Cools et al, 2002; Frith, 1987).
The present study investigated the contribution of mPFC 5-HT1A and 5-HT2A receptor activity to various aspects of performance in the 5-CSRT task in conditions of disrupted glutamate NMDA neurotransmission in the PFC. Various doses of a selective 5-HT1A receptor agonist 8-hydroxy-2-(di-n-propylamino) tetraline (8-OH-DPAT) (Hoyer et al, 1994; Peroutka, 1986) or 5-HT2A receptor antagonist [R-(+)-(2,3-dimethoxyphenyl)-l-[2-(4-fluorophenylethyl)]-4-piperidine-methanol] (M100907) (Kehne et al, 1996) were microinjected into the mPFC and their effects examined on attentional performance deficits induced by the competitive NMDA receptor antagonist 3-(R)-2-carboxypiperazin-4-propyl-1-phosphonic acid (CPP) (Lehmann et al, 1987) injected into the same cortical area. The selective 5-HT1A antagonist N-[2-[4-(2-methoxyphenyl)-1-piperazinyl)ethyl]-N-2-pyridinylcyclohexanecarboxamide, WAY100635 (Forster et al, 1995) was used to antagonize any effect of 8-OH-DPAT on rats' performance.
Stimulation of 5-HT1A or blockade of 5-HT2A receptors in the mPFC prevented the CPP-induced deficit in attentional functioning. The CPP-induced loss of executive control such as the increase in impulsivity was prevented by M100907, while 8-OH-DPAT decreased compulsive perseveration.
MATERIALS AND METHODS
Animals
Male hooded Lister rats (Charles River, Italy) weighing between 300 and 350 g before surgery were used. They were housed in pairs until surgery and then singly in a temperature-controlled room (21°C) with a day/night cycle (0700–1900 h). Water was available ad libitum. Limited access to food (about 15 g of Altromin pellets for rats) at the end of each day's testing kept the animals at 85–90% of their initial free-feeding weight. Procedures involving animals and their care were conducted in conformity with the institutional guidelines that are in compliance with the national (D.L. no. 116, G.U., suppl., 40, 18 Febbraio, 1992, Circolare no. 8, G.U., 14 luglio 1994) and international laws and policies (EEC Council Directive 86/609, OJ L 358,1, December 12, 1987; Guide for the Care and Use of Laboratory Animals, US National Research Council, 1996).
Apparatus
The test apparatus has been described in detail (Carli et al, 1983). It consisted of two 25 × 25 cm aluminum chambers built in the Department of Experimental Psychology, University of Cambridge. The rear wall of each box was concavely curved, and had set into its full arc nine square holes, 4 cm deep and 2 cm above floor level. Each hole had an infrared beam crossing the entrance vertically and illuminating a photoelectric cell. A standard 3 W bulb at the rear of each hole provided illumination. Food pellets (Sandown Scientific, UK) were delivered to a tray at the front of the box. A hinged panel blocked the entrance to the tray. A 3 W house-light was installed centrally in the box roof. Each apparatus was controlled on-line and data were collected by a Control Universal Cube microcomputer system (Cambridge, UK), with software written in ONLIBASIC.
Behavioral Procedures
Animals were trained to a stable performance in the 5-CSRT task as previously described (Carli et al, 1983). The start of the session was signalled by illumination of the house-light and delivery of a single food pellet. Opening the panel to collect the pellet began the first trial. After a fixed delay (the inter-trial interval, ITI), the light at the rear of one of the holes came on for a short period. The light stimulus was presented the same number of times in each hole during the course of a complete session, with the order of presentation randomized by the computer. While the light was on, and for a short period afterwards (the limited hold), responses in the hole that was illuminated (correct responses) resulted in the delivery of a food pellet. Responses in the holes that had not been illuminated (incorrect responses) or failure to respond within the limited hold (omissions) caused the house-light to be turned off for a short period (time out). Responses made in the holes while the house-light was off restarted the time out.
After the delivery of food, or at the end of time out, the rat started the next trial by opening the panel at the front of the chamber. Responses made in the holes after a correct response (perseverative responses), or after the end of time out before opening the panel, resulted in a period of time out. Responses made in the holes during the ITI (anticipatory responses) also resulted in a period of time out. After anticipatory responses, however, opening the panel restarted the current trial. Each daily session consisted of 100 trials or 30 min of testing, whichever was completed sooner, after which all lights were turned off and further responses had no effect.
In the first session of the test schedule, the stimulus and limited hold each lasted 1 min and, depending on individual performances, they were progressively reduced to 0.5 and 5 s, respectively. The ITI and time out both lasted 2 s during the first session and the ITI was raised to 5 s in subsequent sessions; time out was not changed. When the rats reached a stable performance with a mean of 80% correct responses and no more than 15% omissions, they were allocated to different treatment schedules. Each rat had only one session per day on the 5-CSRT task throughout the experiments.
Surgery
Rats previously trained to a stable level of performance were anesthetized by an intraperitoneal (I.P.) injection (2 ml/kg) containing 40 mg/ml ketamine and 5 mg/ml xylazine. All animals received I.P. injections of 0.1 mg/kg atropine sulphate. The animals were secured in a stereotaxic frame (Kopf Ins., USA) with the incisor bar set at −3.3 mm relative to the inter-aural line. Bilateral 23-gauge, stainless-steel guide cannulae (Cooper's Needles, UK) were implanted in the medial region of the prefrontal cortex (mPFC) using standard stereotaxic techniques and secured to the skull using three bone screws and dental cement. The coordinates used were: anteroposterior +3.1 mm from bregma, lateral ±0.7 mm from midline, and dorsoventral −2.8 from dura (Paxinos and Watson, 1982). Thirty-gauge stainless-steel stylets were inserted flush with the end of the guide cannulae. After surgery, rats were housed singly and had 1 week of recovery without training on the task. After recovery, all rats were retrained on the task to re-establish the presurgery level of baseline performance.
Microinfusion Procedure
On testing days, the rat was held and the stylets were removed and two injection units terminating 2 mm below the tip of the guides were inserted. A volume of 1 μl per hemisphere of various doses of drugs or vehicle were delivered at a rate of 0.5 μl/min by a 10-μl Hamilton syringe, mounted in a CMA/100 infusion pump (CMA Microdialysis, Sweden), connected by PP10 tubing to the injection units. Injection units were left in place for 1 min to allow for diffusion.
Histology
After completion of the behavioral testing, rats were deeply anesthetized with chloral hydrate (400 mg/kg, I.P.) and killed by decapitation. Brains were removed and postfixed in 4% formalin solution. Subsequently, the brains were transferred to 20% sucrose in 0.2 M phosphate buffer saline. The next day the brains were frozen in n-pentane and stored at −20°C. Coronal sections were cut at 30 μm in a Cryo-Cut and stained with cresyl violet. Inspection of the stained slides under the light microscope and the trajectory of gliosis produced by the cannula allowed its location and tip to be estimated and mapped on the atlas (Paxinos and Watson, 1982). Only data from rats in which the cannulae were in the desired area were included in the results. Three rats were excluded because of infection at the injection site.
Drugs and Experimental Design
CPP (Tocris, UK), and 8-OH-DPAT (Tocris, USA) and WAY100635 (Pharmacia, Nerviano, Italy) were dissolved in the phosphate buffer saline (PBS composition in mM: NaCl 137, KCl 2.7, Na2HPO4 8.0, KH2PO4 1.8, pH 7.4). M100907 (Aventis, USA) was dissolved in vehicle (PBS containing a few drops of HCl 1 M). The pH of the solution was adjusted to 7 with NaOH 1 M.
A group of rats (n=11) received 1 μl vehicle (PBS) or 30 and 100 ng/μl 8-OH-DPAT into the mPFC 5 min before an injection of 1 μl PBS or 50 ng/μl CPP into the same cortical area. At 10 min after the last cortical injection, rats were put into the box and the test session started. A different group of rats (n=14) received a volume of 1 μl vehicle (PBS) or 100 and 300 ng/μl M100907 into the mPFC 5 min before 1 μl PBS or 50 ng/μl CPP into the same cortical region. At 10 min later rats were put into the box and the test session started.
Eight rats were used to assess the effects of various combinations of vehicle, WAY100635 and 8-OH-DPAT on CPP-induced performance deficits. WAY100635 (30 ng/0.5 μl) or vehicle (0.5 μl) were mixed with the solution of 8-OH-DPAT (100 ng/0.5 μl) or vehicle (0.5 μl) and injected in a volume of 1 μl into the mPFC 5 min before CPP (50 ng/μl). Using thin-layer chromatography, it was shown that WAY100635 and 8-OH-DPAT do not form stable complexes when mixed in a solution (Carli et al. 1998). On each test day, drugs solutions were administered according to a Latin-square design. At least 2 days were left between test days. Rats were always tested on these intervening days to re-establish the baseline and check for long-lasting effects of drugs.
Statistical Analysis
The main dependent variables selected for analysis were: (a) the percentage of correct responses (total correct responses/total correct + total incorrect responses); (b) percentage of omissions (total omissions/total number of trials); (c) the number of anticipatory responses in the holes during the ITI, (d) the number of perseverative responses in the holes after a correct response, (e) mean correct response latency (to the nearest 0.01 s), and (f) mean latency to collect the reinforcement (to the nearest 0.01 s).
Correct responses and omissions, as percentages, were transformed according to the formula 2arcsin(SQRT (%X/100)). The mean latencies to respond correctly and to collect the reinforcement were transformed by log 10. These transformations were performed in order to normalize the distributions in accordance with the ANOVA model (Winer, 1971). Data from 11 rats were used to examine the effects of 8-OH-DPAT combined with saline or CPP and data from 14 rats were used to test the effects of M100907 combined with saline or CPP. Data were analyzed by a within-subject 2 × 2 ANOVA with factors 8-OH-DPAT or M100907 and CPP. The data of eight rats testing the effects of WAY100635 plus 8-OH-DPAT were analyzed by a 2 × 2 repeated measure ANOVA. The mean values of individual treatments were compared using Tukey's HDS test. Statistical software (SAS Institute Inc., USA.) was run on a Micro VAX 3500 computer (Digital, USA).
RESULTS
Histology
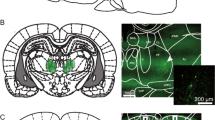
Examination of stained coronal sections of the brains showed that multiple injections into the mPFC produced limited tissue damage. Additionally, in the between-injections days rats returned to perform at very high level of accuracy (80–90%), making less than 20% omissions and only a small number of anticipatory and perseverative responses. Thus, the various treatments and multiple injections did not have a permanent effect on performance. Figure 1 shows a photograph of the representative histological section of the rat brain.

Some representative photographs of the histological sections showing the cannula tracks and the infusion sites in two (a and b) different rats.
Effects of the 5-HT1A Receptor Agonist on CPP-Induced Deficits
Figure 2a shows that 8-OH-DPAT prevented the CPP-induced decrease in the percentage of correct responses (8-OH-DPAT × CPP, F2,50=3.6, P=0.03; 8-OH-DPAT, F2,50=11.0, P=0.0001; CPP, F1,50=47.8, P<0.0001). Multiple comparison of the various treatment group means by Tukey's HDS test indicated that 30 and 100 ng/μl 8-OH-DPAT (both P<0.05) prevented the decrease in correct responses induced by CPP (P<0.05). By itself 8-OH-DPAT had no such effect (P>0.05).

The effects of 8-OH-DPAT and CPP alone and in combination on the percentage of correct responses (a), the number of anticipatory (b) and the number of perseverative (c) responses. Vehicle 1 μl (V) or 8-OH-DPAT at doses of 30 ng/μl (D 30) and 100 ng/μl (D 100) were injected into the mPFC 5 min before bilateral injections of 1 μl vehicle (VEHICLE) or 50 ng/μl CPP (CPP) into the same area. After 10 min, the rats started the test sessions. The various treatment combinations were administered at least 48 h apart, according to a Latin-square design. The histograms represent the mean±SEM of 11 rats. *P<0.05 vs V (+VEHICLE); #P<0.05 vs V (+CPP) (Tukey's test).
The CPP-induced increase in the number of anticipatory responses (Figure 2b) was not affected by 8-OH-DPAT (8-OH-DPAT × CPP, F2,50=1.5, P=0.2; 8-OH-DPAT, F2,50=0.3, P=0.7; CPP, F1,50=5.6, P=0.02). By itself 8-OH-DPAT tended to increase anticipatory responses, but not significantly (P>0.05). The CPP-induced perseverative over-responding (Figure 2c) was significantly reduced by 8-OH-DPAT (8-OH-DPAT × CPP, F2,50=3.9, P=0.02; 8-OH-DPAT, F2,50=1.0, P=0.4; CPP, F1,50=16.5, P=0.0002). Doses of 30 and 100 ng/μl 8-OH-DPAT by themselves had no effect on perseverative responses but dose-dependently lowered the effects of CPP (P<0.05).
Table 1 shows the effects of 8-OH-DPAT on the CPP-induced increase in the proportion of omissions (8-OH-DPAT × CPP, F2,50=2.6, P=0.08; 8-OH-DPAT, F2,50=3.5, P=0.03; CPP, F1,50=30.5, P=0.0001) and the mean latency to make a correct response (8-OH-DPAT × CPP, F2,50=1.9, P=0.15; 8-OH-DPAT, F2,50=5.3, P=0.008; CPP, F1,50=22.3, P=0.0001). Further analysis by comparing individual treatment means indicated that by itself 8-OH-DPAT had no effect on omissions but reduced the effects of CPP on omissions, although only at 30 ng/μl (P<0.05). In the control condition, 8-OH-DPAT speeded up correct responding, although only at 30 ng/μl (P<0.05). Similarly, the CPP-induced increases in correct response latency were reduced by 30 ng/μl (P<0.05) but not 100 ng/μl 8-OH-DPAT.
Effects of WAY100635 Plus 8-OH-DPAT on CPP-Induced Deficits
We examined the effects of 30 and 100 ng/μl WAY100635 on performance of rats injected with vehicle (1 μl) or CPP (50 ng/μl) into the mPFC. WAY100635 had no effects on any measure of performance in rats receiving vehicle (1 μl) into the mPFC (data not shown). However, 100 ng/μl WAY100635 interfered with the performance of rats given CPP (50 ng/μl); they stopped performing and made a large proportion of omissions (data not shown). Thus, 30 ng/μl WAY100635 was selected to examine the selectivity of 8-OH-DPAT's effects on CPP-induced performance deficit.
All animals received bilateral injections of 50 ng/μl CPP into the mPFC. This experiment showed that 8-OH-DPAT increased accuracy (F1,21=27.7, P<0.0001) and that this effect was blocked by WAY100635 (WAY100635 × 8-OH-DPAT, F1,21=4.6, P=0.04; WAY100635, F1,21=12.6, P=0.002). Multiple comparison of the treatment group means by Tukey's HDS test is illustrated in Table 2; after 100 ng/μl 8-OH-DPAT in the mPFC rats made a higher proportion of correct responses than after vehicle (P<0.05) thus replicating in a new group of rats the results of the experiment presented in Figure 2a. WAY100635 30 ng/μl abolished the effect of 100 ng/μl 8-OH-DPAT on the percentage of correct responses since rats receiving both drugs made fewer correct responses than animals receiving vehicle with 8-OH-DPAT (P<0.05). Similarly, WAY100635 blocked the effects of 8-OH-DPAT on the number of perseverative responses (WAY100635 × 8-OH-DPAT, F1,21=7.6, P=0.05). Table 2 shows that after 8-OH-DPAT rats made fewer perseverative responses than after vehicle (P<0.05) whereas when WAY100635 was added to 8-OH-DPAT, rats made more perseverative responses than those receiving only 8-OH-DPAT (P<0.05). Other measures of rats' performance such as anticipatory responses, proportion of omissions, and correct response latencies were not affected by 8-OH-DPAT, WAY100635, or the combination (data not shown) and are not commented further.
Effects of the 5-HT2A Receptor Antagonist on CPP-Induced Deficits
Figure 3a shows that doses of 100 and 300 ng/μl M100907 by themselves had no effect on accuracy (% correct responses) but prevented the CPP-induced impairment in accuracy (M100907 × CPP, F2,65=5.8, P=0.004; M100907, F2,65=5.4, P=0.006; CPP, F1,65=31.7, P<0.0001). Both doses of M100907 were equally potent in preventing the accuracy impairment induced by CPP (both P<0.05).

The effects of M100907 and CPP alone and in combination on the percentage of correct responses (a), the number of anticipatory (b) and the number of perseverative (c) responses. Vehicle 1 μl (V) or M100907 at doses of 100 ng/μl (M 100) and 300 ng/μl (M 300) were injected into the mPFC 5 min before bilateral injections of 1 μl vehicle (VEHICLE) or 50 ng/μl CPP (CPP) into the same area. After 10 min, the rats started the test sessions. The various treatment combinations were administered at least 48 h apart, according to a Latin-square design. The histograms represent the mean±SEM of 14 rats. *P<0.05 vs V (+VEHICLE); #P<0.05 vs V (+CPP) (Tukey's test).
The CPP-induced increase in anticipatory responses (Figure 3b) was dose-dependently reduced by 100 and 300 ng/μl M100907 (M100907 × CPP, F2,65=7.9, P=0.0009; M100907, F2,65=13.2, P<0.0001; CPP, F1,65=52.2, P<0.0001). In the control condition, 300 ng/μl M100907 tended to reduce anticipatory responses, but not significantly, possibly because the number of anticipatory responses was already low. Figure 3c shows that M100907 had no effect by itself on perseverative responses nor did it affect CPP-induced perseverative over-responding (M100907 × CPP, F2,65=1.2, P=0.3; M100907, F2,65=0.9, P=0.4; CPP, F1,65=116.1, P<0.0001).
As shown in Table 3, in the control condition M100907 did not affect omissions or the speed of correct responding. The CPP-induced increases in omission (M100907 × CPP, F2,65=0.6, P=0.6; M100907, F2,65=1.4, P=0.2; CPP, F1,65=32.9, P<0.0001) and mean latency to a correct response were unaffected by M100907 (M100907 × CPP, F2,65=1.6, P=0.2; M100907, F2,65=2.5, P=0.08; CPP, F1,65=33.6, P<0.0001).
DISCUSSION
This is the first study to demonstrate a dissociable contribution of serotonin 5-HT1A and 5-HT2A receptors in the mPFC to aspects of executive control such as impulsivity and compulsive perseveration. It also shows that these receptors exert opposite action on attentional functioning. The selective and competitive glutamate NMDA receptor antagonist CPP (Lehmann et al, 1987), injected into the mPFC, impaired accuracy and enhanced anticipatory and perseverative responses. These effects were accompanied by increases in omissions and latencies for correct detection, confirming previous reports (Mirjana et al, 2004; Murphy et al, 2005). As shown by Carli et al (Mirjana et al, 2004) the deficit may be recovered to a certain degree, when, by increasing stimulus duration, the attentional load on performance is reduced, suggesting that it is attentional in nature.
Like systemic M100907 (Mirjana et al, 2004), intra-mPFC injections of 100 and 300 ng/μl M100907 abolished the deficit in attentional accuracy and anticipatory but not perseverative responding induced by 50 ng/μl CPP. Various behavioral deficits induced by NMDA antagonists have been associated with enhanced glutamate release in the mPFC (Moghaddam et al, 1997; Moghaddam and Adams, 1998) and findings in our laboratory indicate that CPP in the mPFC increases glutamate efflux locally and that this was prevented by systemic or intra-mPFC M100907 (Ceglia et al, 2004). Thus, 5-HT2A receptors in the mPFC play a major role in controlling CPP-induced glutamate release and some aspects of attentional performance in a 5-CSRT task.
Impairments in attentional accuracy induced by CPP were completely abolished by 30 and 100 ng/μl 8-OH-DPAT injected into the mPFC. The 5-HT1A receptors are found in a large proportion (50–60%) in glutamate neurons of the mPFC (Santana et al, 2004) thus raising the possibility that 5-HT1A receptors may exert some of their functions by acting on glutamatergic signalling. Additionally, the 5-HT1A receptor subtype can be considered functionally antagonistic to the 5-HT2A receptors. They are highly colocalized (80%) in pyramidal neurons of the PFC (Santana et al, 2004) and electrophysiological studies have shown that M100907 potentiate 8-OH-DPAT suppression on firing rate (Ashby et al, 1994). In addition, 5-HT2A receptor antagonists ICI 180,809 and ritanserin potentiate the 5-HT syndrome produced by 8-OH-DPAT (Backus et al, 1990; Sharp et al, 1990) whereas 8-OH-DPAT inhibits head twitching behavior induced by systemic DOI (Berendsen and Broekkamp, 1990; Darmani et al, 1990; Dursun and Handley, 1993) or microinjection into the mPFC of (−) DOB, a congener of DOI and a 5-HT2A/2C receptors agonist (Granhoff et al, 1992). Clearly, the opposition between the two 5-HT receptor subtypes suggests that the improvement produced by M100907 and 8-OH-DPAT on CPP-induced accuracy deficit may result from a functionally antagonistic activity of these receptors on a common intracellular mechanism.
8-OH-DPAT but not M100907 had some additional effects on rats' attentional functioning but only at the low dose of 30 ng/μl; it speeded up correct response latencies and reduced CPP-induced omissions. Substantial evidence implicates the dopaminergic (DA) system in decision processes in this task (Robbins, 2002). The speeding up of correct responses and a decrease in omissions in a 5-CSRT task had been observed after systemic amphetamine and the dopamine D1 receptor agonist SKF 38393 in the mPFC (Granon et al, 2000; Robbins, 2002). Thus, the fact that intra-mPFC 8-OH-DPAT increases DA efflux in this cortical region (Sakaue et al, 2000) may have contributed to its effect on speed and omissions.
In contrast to the effects of M100907, injections of 8-OH-DPAT into the mPFC did not have any effect on CPP-induced anticipatory responding. However, CPP-induced perseverative over-responding was significantly reduced by pretreatment with 8-OH-DPAT. These results clearly demonstrate the selectivity of executive control processes and indicate that impulsivity and perseveration may be dissociated by 5-HT1A and 5-HT2A receptor mechanisms in the PFC. Evidently response inhibition operates independently for preparing responses and for monitoring performance, thus providing behavioral flexibility. This conclusion is generally consistent with emerging evidence of distinct neural systems in the control of ‘impulsive’ behavior, as, for example, after lesions of the infralimbic (IL) prefrontocortical region and in ‘compulsive’ behaviors associated with lesions of the prelimbic and orbitofrontal regions of the rat PFC (Chudasama and Muir, 2001; Chudasama et al, 2003; Passetti et al, 2002).
A selective 5-HT1A receptor antagonist, WAY100635, blocked the effects of 8-OH-DPAT on CPP-induced accuracy deficits and perseverative over-responding suggesting that the effects of 8-OH-DPAT were due to selective activation of 5-HT1A receptors in the mPFC.
Clinical and experimental evidence shows that the tendency to act without foresight, that is, ‘impulsivity’ may manifest itself in several ways (Evenden, 1999a, 1999b). In reaction time tasks (such as the 5-CSRT task), it might be detected as enhanced errors or inappropriate anticipatory responding (Evenden, 1999b; Robbins, 2002). The 5-HT system has been implicated in the regulation of different forms of impulsive behavior (Evenden, 1999a, 1999b; Mobini et al, 2000; Soubrié, 1986). In fact, impulsivity in a 5-CSRT task as measured by anticipatory responses was associated with high 5-HT turnover (Puumala and Sirvio, 1998) and release in the mPFC (Dalley et al, 2002). Stimulation of 5-HT2A receptors by a variety of 5-HT2A agonists increased whereas blockade of 5-HT2A receptors (by antagonists) reduced anticipatory responses (Higgins et al, 2003b; Koskinen et al, 2000; Mirjana et al, 2004; Passetti et al, 2003b; Winstanley et al, 2003).
Interestingly, opposite behavioral effects of 8-OH-DPAT were often reported depending on whether the drug was administered directly into the DR or into its projecting areas (Carli et al, 1995, 1998; Warburton et al, 1997). Systemic 8-OH-DPAT increased impulsivity and impaired accuracy in a 5-CSRT task through stimulation of presynaptic 5-HT1A autoreceptors in the DR nucleus (Carli and Samanin, 2000) whereas stimulation of postsynaptic 5-HT1A receptors in the mPFC had no effect on impulsivity but improved accuracy (Winstanley et al, 2003; present results). However, facilitation of accuracy in various cognitive tasks has been reported after low brain concentrations of 8-OH-DPAT (Carli et al, 2000; Cole et al, 1994; Winstanley et al, 2003) and the selective 5-HT lesions of the DR nucleus (Harrison et al, 1997b).
The NMDA receptor antagonists either infused into the mPFC or injected systemically increase the release of 5-HT in the mPFC (Ceglia et al, 2004; Martin et al, 1998). Therefore, overactivation of 5-HT2A but not 5-HT1A receptors in the mPFC as a consequence of elevated 5-HT release may be an important mechanism that increases active responding in anticipation of reward. This view is challenged by findings that global 5-HT depletion consistently enhanced anticipatory responding in a 5-CSRT task in the rat (Harrison et al, 1997a, 1997b).
The enhanced tendency to perseverate is presumably an expression of behavioral inflexibility after blockade of NMDA receptors in the mPFC, preventing the rats from suppressing irrelevant responses and shifting their attention to the next relevant response in a well-learned sequence. The ‘compulsive’ perseveration appears to be a distinctive trait of frontal-lesion animals (Mishkin, 1964; Muir et al, 1996), and frontal-lobe and schizophrenic patients when required to suppress previously reinforced responses (Owen et al, 1993 #76; Lyon and Gerlach, 1988 #167; Goldberg and Weinberger, 1994 #166).
That enhanced glutamate and 5-HT release may not be involved in CPP's effects on perseverative responses is indicated by studies showing that lowering CPP-induced glutamate release by M100907 does not abolish perseverative responding (Carli et al, 2004; Ceglia et al, 2004; the present results). In addition, reducing but not increasing 5-HT function in the PFC leads to response perseveration in tasks such as reversal learning (Clarke et al, 2004, 2005) and in some instances in a 5-CSRT (Winstanley et al, 2004).
CPP like other NMDA receptor antagonists increased DA release in the mPFC (Del Arco and Mora, 1999; Feenstra et al, 2002; Moghaddam et al, 1997). Accordingly, increasing DA transmission by systemic D-amphetamine increased perseverative responses in a 5-CSRT task (Baunez and Robbins, 1999). The exact mechanism by which 8-OH-DPAT might reduce CPP-induced perseverative responding is not clear. Both systemic and intra-mPFC 8-OH-DPAT increased mPFC DA efflux (Arborelius et al, 1993; Sakaue et al, 2000). However, 8-OH-DPAT actually reduced the rise in DA release in the mPFC induced by amphetamine, stress, and isolation rearing (Ago et al, 2002; Kuroki et al, 1996; Rasmusson et al, 1994) and attenuated the locomotor effects of amphetamine (Przegalinski and Filip, 1997). Therefore, the decrease in perseverative responding after intra-mPFC 8-OH-DPAT might be due to its effects on the increase in cortical DA release induced by CPP.
It can be speculated that the segregation of 5-HT2A receptors to apical dendrites of glutamatergic pyramidal neurons (Jakab and Goldman-Rakic, 1998) and to GABAergic interneurons specialized in the perisomatic inhibition of pyramidal cells (Jakab and Goldman-Rakic, 2000) affects excitatory glutamate input (Aghajanian and Marek, 1997) whereas 5-HT1A receptors in the axon hillock (Czyrak et al, 2003; DeFelipe et al, 2001) can suppress the generation of action potentials along the axon and influence the activity in their subcortical projection areas. Thus 5-HT, acting on 5-HT1A and 5-HT2A receptors, may finely tune the complex activity of glutamate pyramidal neurons, differently influencing various aspects of cognitive functions. It is of particular interest that mPFC neurons expressing 5-HT1A and 5-HT2A receptors simultaneously project to the 5-HT cells of DR nucleus and DA cells of the ventral tegmental areas (VTA) and influence their activity (Carr and Sesack, 2000; Celada et al, 2001; Hajos et al, 2003; Sesack and Bunney, 1989; Thierry et al, 1983). Cognitive functions of the prefrontal cortex are influenced by the 5-HT system (Robbins, 2000) and by an optimal level of mesocortical dopamine (DA) function (Arnsten, 1997; Granon et al, 2000; Roberts et al, 1994; Zahrt et al, 1997). The ability of some 5-HT2A receptors antagonists to enhance the effects of DA D2 antagonists such as haloperidol on DA function in the mPFC has been attributed to their direct or indirect agonist activity on 5-HT1A receptors (Ichikawa et al, 2001; Liegeois et al, 2002; Meltzer et al, 2003) and may be relevant to how atypical antipsychotics improve cognitive functions in schizophrenic patients (Harvey and Keefe, 2001; Meltzer and McGurk, 1999).
In conclusion, blockade of NMDA receptors in the mPFC, by inducing inhibitory deficits of impulsivity and perseveration as well as inattention, might offer a model of dys-executive syndrome of schizophrenia (Frith, 1987). The data indicate that 5-HT1A and 5-HT2A receptors in the mPFC exert opposing actions on the attentional impairment induced by blockade of NMDA receptors. Furthermore, this is the first demonstration that multiple executive mechanisms that cooperate to preserve accurate response selection can be dissociated at the levels of 5-HT2A and 5-HT1A receptor mechanisms. A complex interaction of glutamate NMDA receptor mechanisms with serotonergic and dopaminergic systems in the prefrontal cortex may be essential to preserve attentional selectivity and executive functioning. Such mechanisms may also help explain how atypical antipsychotics improve cognitive ability in schizophrenia.
References
Aghajanian GK, Marek GJ (1997). Serotonin induces excitatory postsynaptic potentials in apical dendrites of neocortical pyramidal cells. Neuropharmacology 36: 589–599.
Ago Y, Sakaue M, Baba A, Matsuda T (2002). Selective reduction by isolation rearing of 5-HT1A receptor-mediated dopamine release in vivo in the frontal cortex of mice. J Neurochem 83: 353–359.
Andrade R, Nicoll RA (1987). Pharmacologically distinct actions of serotonin on single pyramidal neurones of the rat hippocampus recorded in vitro. J Physiol 394: 99–124.
Araneda R, Andrade R (1991). 5-Hydroxytryptamine 2 and 5-hydroxytryptamine 1A receptors mediate opposing responses on membrane excitability in rat association cortex. Neuroscience 40: 399–412.
Arborelius L, Nomikos GG, Hacksell U, Svensson TH (1993). (R)-8-OH-DPAT preferentially increases dopamine release in rat medial prefrontal cortex. Acta Physiol Scand 148: 465–466.
Arnsten AF (1997). Catecholamine regulation of the prefrontal cortex. J Psychopharmacol 11: 151–162.
Aron AR, Fletcher PC, Bullmore ET, Sahakian BJ, Robbins TW (2003). Stop-signal inhibition disrupted by damage to right inferior frontal gyrus in humans. Nat Neurosci 6: 115–116.
Ashby Jr CR, Edwards E, Wang RY (1994). Electrophysiological evidence for a functional interaction between 5-HT1A and 5-HT2A receptors in the rat medial prefrontal cortex: an iontophoretic study. Synapse 17: 173–181.
Azmitia EC, Segal M (1978). An autoradiographic analysis of the differential ascending projections of the dorsal and median raphe nuclei in the rat. J Comp Neurol 179: 641–667.
Backus LI, Sharp T, Grahame-Smith DG (1990). Behavioural evidence for a functional interaction between central 5-HT2 and 5-HT1A receptors. Br J Pharmacol 100: 793–799.
Barnes NM, Sharp T (1999). A review of central 5-HT receptors and their function. Neuropharmacology 38: 1083–1152.
Baunez C, Robbins TW (1999). Effects of dopamine depletion of the dorsal striatum and further interaction with subthalamic nucleus lesions in an attentional task in the rat. Neuroscience 92: 1343–1356.
Baxter LR (1990). Brain imaging as a tool in establishing a theory of brain pathology in obsessive compulsive disorder. J Clin Psychiatry 51(Suppl): 22–25; discussion 26.
Berendsen HH, Broekkamp CL (1990). Behavioural evidence for functional interactions between 5-HT-receptor subtypes in rats and mice. Br J Pharmacol 101: 667–673.
Blue ME, Yagaloff KA, Mamounas LA, Hartig PR, Molliver ME (1988). Correspondence between 5-HT2 receptors and serotonergic axons in rat neocortex. Brain Res 453: 315–328.
Braff DL (1993). Information processing and attention dysfunctions in schizophrenia. Schizophr Bull 19: 233–259.
Carli M, Samanin R (1992). Serotonin2 receptor agonists and serotonergic anorectic drugs affect rats' performance differently in a five-choice serial reaction time task. Psychopharmacology 106: 228–234.
Carli M, Samanin R (2000). The 5-HT(1A) receptor agonist 8-OH-DPAT reduces rats' accuracy of attentional performance and enhances impulsive responding in a five-choice serial reaction time task: role of presynaptic 5-HT(1A) receptors. Psychopharmacology (Berlin) 149: 259–268.
Carli M, Balducci C, Samanin R (2000). Low doses of 8-OH-DPAT prevent the impairment of spatial learning caused by intrahippocampal scopolamine through 5-HT(1A) receptors in the dorsal raphe. Br J Pharmacol 131: 375–381.
Carli M, Baviera M, Invernizzi RW, Balducci C (2004). The serotonin 5-HT2A receptors antagonist M100907 prevents impairment in attentional performance by NMDA receptor blockade in the rat prefrontal cortex. Neuropsychopharmacology 29: 1637–1647.
Carli M, Bonalumi P, Samanin R (1998). Stimulation of 5-HT1A receptors in the dorsal raphe reverses the impairment of spatial learning caused by intrahippocampal scopolamine in rats. Eur J Neurosci 10: 221–230.
Carli M, Luschi R, Garofalo P, Samanin R (1995). 8-OH-DPAT impairs spatial but not visual learning in a water maze by stimulating 5-HT1A receptors in the hippocampus. Behav Brain Res 67: 67–74.
Carli M, Robbins TW, Evenden JL, Everitt BJ (1983). Effects of lesions to ascending noradrenergic neurones on performance of a 5-choice serial reaction task in rats; implications for theories of dorsal noradrenergic bundle function based on selective attention and arousal. Behav Brain Res 9: 361–380.
Carr DB, Sesack SR (2000). Projections from the rat prefrontal cortex to the ventral tegmental area: target specificity in the synaptic associations with mesoaccumbens and mesocortical neurons. J Neurosci 20: 3864–3873.
Ceglia I, Carli M, Baviera M, Renoldi G, Calcagno E, Invernizzi RW (2004). The 5-HT receptor antagonist M100,907 prevents extracellular glutamate rising in response to NMDA receptor blockade in the mPFC. J Neurochem 91: 189–199.
Celada P, Puig MV, Casanovas JM, Guillazo G, Artigas F (2001). Control of dorsal raphe serotonergic neurons by the medial prefrontal cortex: Involvement of serotonin-1A, GABA(A), and glutamate receptors. J Neurosci 21: 9917–9929.
Chudasama Y, Muir JL (2001). Visual attention in the rat: a role for the prelimbic cortex and thalamic nuclei? Behav Neurosci 115: 417–428.
Chudasama Y, Passetti F, Rhodes SE, Lopian D, Desai A, Robbins TW (2003). Dissociable aspects of performance on the 5-choice serial reaction time task following lesions of the dorsal anterior cingulate, infralimbic and orbitofrontal cortex in the rat: differential effects on selectivity, impulsivity and compulsivity. Behav Brain Res 146: 105–119.
Clarke HF, Dalley JW, Crofts HS, Robbins TW, Roberts AC (2004). Cognitive inflexibility after prefrontal serotonin depletion. Science 304: 878–880.
Clarke HF, Walker SC, Crofts HS, Dalley JW, Robbins TW, Roberts AC (2005). Prefrontal serotonin depletion affects reversal learning but not attentional set shifting. J Neurosci 25: 532–538.
Cole BJ, Jones GH, Turner JD (1994). 5-HT1A receptor agonists improve the performance of normal and scopolamine-impaired rats in an operant delayed matching to position task. Psychopharmacology (Berlin) 116: 135–142.
Cools R, Stefanova E, Barker RA, Robbins TW, Owen AM (2002). Dopaminergic modulation of high-level cognition in Parkinson's disease: the role of the prefrontal cortex revealed by PET. Brain 125: 584–594.
Czyrak A, Czepiel K, Mackowiak M, Chocyk A, Wedzony K (2003). Serotonin 5-HT1A receptors might control the output of cortical glutamatergic neurons in rat cingulate cortex. Brain Res 989: 42–51.
Dalley JW, Theobald DE, Eagle DM, Passetti F, Robbins TW (2002). Deficits in impulse control associated with tonically-elevated serotonergic function in rat prefrontal cortex. Neuropsychopharmacology 26: 716–728.
Darmani NA, Martin BR, Pandey U, Glennon RA (1990). Do functional relationships exist between 5-HT1A and 5-HT2 receptors? Pharmacol Biochem Behav 36: 901–906.
DeFelipe J, Arellano JI, Gomez A, Azmitia EC, Munoz A (2001). Pyramidal cell axons show a local specialization for GABA and 5-HT inputs in monkey and human cerebral cortex. J Comp Neurol 433: 148–155.
Del Arco A, Mora F (1999). Effects of endogenous glutamate on extracellular concentrations of GABA, dopamine, and dopamine metabolites in the prefrontal cortex of the freely moving rat: involvement of NMDA and AMPA/KA receptors. Neurochem Res 24: 1027–1035.
Dursun SM, Handley SL (1993). The effects of alpha 2-adrenoceptor antagonists on the inhibition of 1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane (DOI)-induced head shakes by 5-HT1A receptor agonists in the mouse. Br J Pharmacol 109: 1046–1052.
Egerton A, Reid L, McKerchar CE, Morris BJ, Pratt JA (2005). Impairment in perceptual attentional set-shifting following PCP administration: a rodent model of set-shifting deficits in schizophrenia. Psychopharmacology (Berlin) 179: 77–84.
Evenden JL (1999a). The pharmacology of impulsive behaviour in rats VII: the effects of serotonergic agonists and antagonists on responding under a discrimination task using unreliable visual stimuli. Psychopharmacology (Berlin) 146: 422–431.
Evenden JL (1999b). Varieties of impulsivity. Psychopharmacology (Berlin) 146: 348–361.
Feenstra MG, Botterblom MH, van Uum JF (2002). Behavioral arousal and increased dopamine efflux after blockade of NMDA-receptors in the prefrontal cortex are dependent on activation of glutamatergic neurotransmission. Neuropharmacology 42: 752–763.
Forster EA, Cliffe IA, Bill DJ, Dover GM, Jones D, Reilly Y et al (1995). A pharmacological profile of the selective silent 5-HT1A receptor antagonist, WAY-100635. Eur J Pharmacol 281: 81–88.
Frith CD (1987). The positive and negative symptoms of schizophrenia reflect impairments in the perception and initiation of action. Psychol Med 17: 631–648.
Goff DC, Coyle JT (2001). The emerging role of glutamate in the pathophysiology and treatment of schizophrenia. Am J Psychiatry 158: 1367–1377.
Goldberg TE, Weinberger DR (1994). Schizophrenia, training paradigms, and the Wisconsin Card Sorting Test redux. Schizophr Res 11: 291–296 (Review).
Granhoff MI, Lee C, Jackson A, Patel K, Martinez Y, Ashby Jr CR et al (1992). The interaction of 5-HT1A and 5-HT2 receptors in the rat medial prefrontal cortex: behavioural studies. Neuroscience Abstract Vol 19, ed. Neuroscience Sf., pp 1980.
Granon S, Passetti F, Thomas KL, Dalley JW, Everitt BJ, Robbins TW (2000). Enhanced and impaired attentional performance after infusion of D1 dopaminergic receptor agents into rat prefrontal cortex. J Neurosci 20: 1208–1215.
Hajos M, Gartside SE, Varga V, Sharp T (2003). In vivo inhibition of neuronal activity in the rat ventromedial prefrontal cortex by midbrain-raphe nuclei: role of 5-HT1A receptors. Neuropharmacology 45: 72–81.
Harrison AA, Everitt BJ, Robbins TW (1997a). Central 5-HT depletion enhances impulsive responding without affecting the accuracy of attentional performance: interactions with dopaminergic mechanisms. Psychopharmacology (Berlin) 133: 329–342.
Harrison AA, Everitt BJ, Robbins TW (1997b). Doubly dissociable effects of median- and dorsal-raphe lesions on the performance of the five-choice serial reaction time test of attention in rats. Behav Brain Res 89: 135–149.
Harvey PD, Keefe RS (2001). Studies of cognitive change in patients with schizophrenia following novel antipsychotic treatment. Am J Psychiatry 158: 176–184.
Harvey PD, Geyer MA, Robbins TW, Krystal JH (2003a). Cognition in schizophrenia: from basic science to clinical treatment. Psychopharmacology (Berlin) 169: 213–214.
Harvey PD, Green MF, McGurk SR, Meltzer HY (2003b). Changes in cognitive functioning with risperidone and olanzapine treatment: a large-scale, double-blind, randomized study. Psychopharmacology (Berlin) 169: 404–411.
Higgins GA, Ballard TM, Huwyler J, Kemp JA, Gill R (2003a). Evaluation of the NR2B-selective NMDA receptor antagonist Ro 63-1908 on rodent behaviour: evidence for an involvement of NR2B NMDA receptors in response inhibition. Neuropharmacology 44: 324–341.
Higgins GA, Enderlin M, Haman M, Fletcher PJ (2003b). The 5-HT2A receptor antagonist M100,907 attenuates motor and ‘impulsive-type’ behaviours produced by NMDA receptor antagonism. Psychopharmacology (Berlin) 170: 309–319.
Hoyer D, Clarke DE, Fozard JR, Hartig PR, Martin GR, Mylecharane EJ et al (1994). International Union of Pharmacology classification of receptors for 5-hydroxytryptamine (Serotonin). Pharmacol Rev 46: 157–203.
Ichikawa J, Ishii H, Bonaccorso S, Fowler WL, O'Laughlin IA, Meltzer HY (2001). 5-HT(2A) and D(2) receptor blockade increases cortical DA release via 5-HT(1A) receptor activation: a possible mechanism of atypical antipsychotic-induced cortical dopamine release. J Neurochem 76: 1521–1531.
Jackson ME, Homayoun H, Moghaddam B (2004). NMDA receptor hypofunction produces concomitant firing rate potentiation and burst activity reduction in the prefrontal cortex. Proc Natl Acad Sci USA 101: 8467–8472.
Jakab RL, Goldman-Rakic PS (1998). 5-Hydroxytryptamine2A serotonin receptors in the primate cerebral cortex: possible site of action of hallucinogenic and antipsychotic drugs in pyramidal cell apical dendrites. Proc Natl Acad Sci USA 95: 735–740.
Jakab RL, Goldman-Rakic PS (2000). Segregation of serotonin 5-HT2A and 5-HT3 receptors in inhibitory circuits of the primate cerebral cortex. J Comp Neurol 417: 337–348.
Javitt DC, Zukin SR (1991). Recent advances in the phencyclidine model of schizophrenia. Am J Psychiatry 148: 1301–1308.
Jentsch JD, Roth RH (1999). The neuropsychopharmacology of phencyclidine: from NMDA receptor hypofunction to the dopamine hypothesis of schizophrenia. Neuropsychopharmacology 20: 201–225.
Kay SR, Sevy S (1990). Pyramidical model of schizophrenia. Schizophr Bull 16: 537–545.
Kehne JH, Baron BM, Carr AA, Chaney SF, Elands J, Feldman DJ et al (1996). Preclinical characterization of the potential of the putative atypical antipsychotic MDL 100,907 as a potent 5-HT2A antagonist with a favorable CNS safety profile. J Pharmacol Exp Ther 277: 968–981.
Konradi C, Heckers S (2003). Molecular aspects of glutamate dysregulation: implications for schizophrenia and its treatment. Pharmacol Ther 97: 153–179.
Koskinen T, Ruotsalainen S, Puumala T, Lappalainen R, Koivisto E, Mannisto PT et al (2000). Activation of 5-HT2A receptors impairs response control of rats in a five-choice serial reaction time task. Neuropharmacology 39: 471–481.
Krystal JH, Karper LP, Seibyl JP, Freeman GK, Delaney R, Bremner JD et al (1994). Subanesthetic effects of the noncompetitive NMDA antagonist, ketamine, in humans. Psychotomimetic, perceptual, cognitive, and neuroendocrine responses. Arch Gen Psychiatry 51: 199–214.
Kuroki T, Ichikawa J, Dai J, Meltzer HY (1996). R(+)-8-OH-DPAT, a 5-HT1A receptor agonist, inhibits amphetamine-induced serotonin and dopamine release in rat medial prefrontal cortex. Brain Res 743: 357–361.
Kurtz MM, Ragland JD, Bilker W, Gur RC, Gur RE (2001). Comparison of the continuous performance test with and without working memory demands in healthy controls and patients with schizophrenia. Schizophr Res 48: 307–316.
Le Pen G, Grottick AJ, Higgins GA, Moreau JL (2003). Phencyclidine exacerbates attentional deficits in a neurodevelopmental rat model of schizophrenia. Neuropsychopharmacology 28: 1799–1809.
Lehmann J, Schneider J, McPherson S, Murphy DE, Bernard P, Tsai C et al (1987). CPP, a selective N-methyl-D-aspartate (NMDA)-type receptor antagonist: characterization in vitro and in vivo. J Pharmacol Exp Ther 240: 737–746.
Liegeois JF, Ichikawa J, Meltzer HY (2002). 5-HT(2A) receptor antagonism potentiates haloperidol-induced dopamine release in rat medial prefrontal cortex and inhibits that in the nucleus accumbens in a dose-dependent manner. Brain Res 947: 157–165.
Lyon N, Gerlach J (1988). Perseverative structuring of responses by schizophrenic and affective disorder patients. J Psychiatr Res 22: 261–277.
Martin P, Carlsson ML, Hjorth S (1998). Systemic PCP treatment elevates brain extracellular 5-HT: a microdialysis study in awake rats. Neuroreport 9: 2985–2988.
Meltzer HY, McGurk SR (1999). The effects of clozapine, risperidone, and olanzapine on cognitive function in schizophrenia. Schizophr Bull 25: 233–255.
Meltzer HY, Li Z, Kaneda Y, Ichikawa J (2003). Serotonin receptors: their key role in drugs to treat schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry 27: 1159–1172.
Mirjana C, Baviera M, Invernizzi RW, Balducci C (2004). The serotonin 5-HT2A receptors antagonist M100907 prevents impairment in attentional performance by NMDA receptor blockade in the rat prefrontal cortex. Neuropsychopharmacology 29: 1637–1647.
Mishkin M (1964). Perseveration of central sets after frontal lesions in monkeys. In: Warren JM, Akert K (eds). The Frontal Granular Cortex and Behaviour. McGraw-Hill: New York.
Mobini S, Chiang TJ, Ho MY, Bradshaw CM, Szabadi E (2000). Effects of central 5-hydroxytryptamine depletion on sensitivity to delayed and probabilistic reinforcement. Psychopharmacology (Berlin) 152: 390–397.
Moghaddam B, Adams BW (1998). Reversal of phencyclidine effects by a group II metabotropic glutamate receptor agonist in rats. Science 281: 1349–1352.
Moghaddam B, Adams B, Verma A, Daly D (1997). Activation of glutamatergic neurotransmission by ketamine: a novel step in the pathway from NMDA receptor blockade to dopaminergic and cognitive disruptions associated with the prefrontal cortex. J Neurosci 17: 2921–2927.
Muir JL, Everitt BJ, Robbins TW (1996). The cerebral cortex of the rat and visual attentional function: dissociable effects of mediofrontal, cingulate, anterior dorsolateral, and parietal cortex lesions on a five-choice serial reaction time task. Cereb Cortex 6: 470–481.
Murphy ER, Dalley JW, Robbins TW (2005). Local glutamate receptor antagonism in the rat prefrontal cortex disrupts response inhibition in a visuospatial attentional task. Psychopharmacology (Berlin) 179: 99–107.
Orzack MH, Kornetsky C (1966). Attention dysfunction in chronic schizophrenia. Arch Gen Psychiatry 14: 323–326.
Owen AM, Roberts AC, Hodges JR, Summers BA, Polkey CE, Robbins TW (1993). Contrasting mechanisms of impaired attentional set-shifting in patients with frontal lobe damage or Parkinson's disease. Brain 116(Part 5): 1159–1175.
Passetti F, Chudasama Y, Robbins TW (2002). The frontal cortex of the rat and visual attentional performance: dissociable functions of distinct medial prefrontal subregions. Cereb Cortex 12: 1254–1268.
Passetti F, Levita L, Robbins TW (2003b). Sulpiride alleviates the attentional impairments of rats with medial prefrontal cortex lesions. Behav Brain Res 138: 59–69.
Paxinos G, Watson C (1982). The Rat Brain in Stereotaxic Coordinates. Academic Press: Sydney.
Peroutka SJ (1986). Pharmacological differentiation and characterization of 5-HT1A, 5-HT1B, and 5-HT1C binding sites in rat frontal cortex. J Neurochem 47: 529–540.
Przegalinski E, Filip M (1997). Stimulation of serotonin (5-HT)1A receptors attenuates the locomotor, but not the discriminative, effects of amphetamine and cocaine in rats. Behav Pharmacol 8: 699–706.
Puig MV, Celada P, Diaz-Mataix L, Artigas F (2003). In vivo modulation of the activity of pyramidal neurons in the rat medial prefrontal cortex by 5-HT2A receptors: relationship to thalamocortical afferents. Cereb Cortex 13: 870–882.
Puumala T, Sirvio J (1998). Changes in activities of dopamine and serotonin systems in the frontal cortex underlie poor choice accuracy and impulsivity of rats in an attention task. Neuroscience 83: 489–499.
Rasmusson AM, Goldstein LE, Deutch AY, Bunney BS, Roth RH (1994). 5-HT1a agonist +/-8-OH-DPAT modulates basal and stress-induced changes in medial prefrontal cortical dopamine. Synapse 18: 218–224.
Robbins TW (1998). Dissociating executive functions of the prefrontal cortex. In: Robberts AC, Robbins TW, Weiskrantz L (eds). The Prefrontal Cortex: Executive and Cognitive Functions. Oxford University Press: Oxford, New York. pp 117–130.
Robbins TW (2000). Chemical neuromodulation of frontal-executive functions in humans and other animals. Exp Brain Res 133: 130–138.
Robbins TW (2002). The 5-choice serial reaction time task: behavioural pharmacology and functional neurochemistry. Psychopharmacology (Berlin) 163: 362–380.
Roberts AC, De Salvia MA, Wilkinson LS, Collins P, Muir JL, Everitt BJ et al (1994). 6-Hydroxydopamine lesions of the prefrontal cortex in monkeys enhance performance on an analog of the Wisconsin Card Sort Test: possible interactions with subcortical dopamine. J Neurosci 14: 2531–2544.
Rodefer JS, Murphy ER, Baxter MG (2005). PDE10A inhibition reverses subchronic PCP-induced deficits in attentional set-shifting in rats. Eur J Neurosci 21: 1070–1076.
Sakaue M, Somboonthum P, Nishihara B, Koyama Y, Hashimoto H, Baba A et al (2000). Postsynaptic 5-hydroxytryptamine(1A) receptor activation increases in vivo dopamine release in rat prefrontal cortex. Br J Pharmacol 129: 1028–1034.
Santana N, Bortolozzi A, Serrats J, Mengod G, Artigas F (2004). Expression of serotonin1A and serotonin2A receptors in pyramidal and GABAergic neurons of the rat prefrontal cortex. Cereb Cortex 14: 1100–1109.
Sesack SR, Bunney BS (1989). Pharmacological characterization of the receptor mediating electrophysiological responses to dopamine in the rat medial prefrontal cortex: a microiontophoretic study. J Pharmacol Exp Ther 248: 1323–1333.
Shallice T (1982). Specific impairments of planning. Philos Trans R Soc Lond B Biol Sci 298: 199–209.
Sharp T, Backus LI, Hjorth S, Bramwell SR, Grahame-Smith DG (1990). Further investigation of the in vivo pharmacological properties of the putative 5-HT1A antagonist, BMY 7378. Eur J Pharmacol 176: 331–340.
Soubrié P (1986). Reconciling the role of central serotonin neurons in human and animal behaviour. Behav Brain Sci 9: 319–364.
Steinbusch HWM (1984). Serotonin-immunoreactive neurons and their projections in the CNS. In: Bjorklund A, Hokfelt T, Kuhar MJ (eds). Handbook of Chemical Neuroanatomy. Elsevier Science: Amsterdam. pp 68–125.
Tanaka E, North RA (1993). Actions of 5-hydroxytryptamine on neurons of the rat cingulate cortex. J Neurophysiol 69: 1749–1757.
Thierry AM, Deniau JM, Chevalier G, Ferron A, Glowinski J (1983). An electrophysiological analysis of some afferent and efferent pathways of the rat prefrontal cortex. Prog Brain Res 58: 257–261.
Verma A, Moghaddam B (1996). NMDA receptor antagonists impair prefrontal cortex function as assessed via spatial delayed alternation performance in rats: modulation by dopamine. J Neurosci 16: 373–379.
Warburton EC, Harrison AA, Robbins TW, Everitt BJ (1997). Contrasting effects of systemic and intracerebral infusions of the 5-HT1A receptor agonist 8-OH-DPAT on spatial short-term working memory in rats. Behav Brain Res 84: 247–258.
Williams GV, Rao SG, Goldman-Rakic PS (2002). The physiological role of 5-HT2A receptors in working memory. J Neurosci 22: 2843–2854.
Winer BJ (1971). Statistical Principles in Experimental Design. McGraw-Hill: Tokyo.
Winstanley CA, Chudasama Y, Dalley JW, Theobald DE, Glennon JC, Robbins TW (2003). Intra-prefrontal 8-OH-DPAT and M100907 improve visuospatial attention and decrease impulsivity on the five-choice serial reaction time task in rats. Psychopharmacology (Berlin) 167: 304–317.
Winstanley CA, Theobald DE, Dalley JW, Glennon JC, Robbins TW (2004). 5-HT2A and 5-HT2C receptor antagonists have opposing effects on a measure of impulsivity: interactions with global 5-HT depletion. Psychopharmacology (Berlin) 176: 376–385.
Zahrt J, Taylor JR, Mathew RG, Arnsten AF (1997). Supranormal stimulation of D1 dopamine receptors in the rodent prefrontal cortex impairs spatial working memory performance. J Neurosci 17: 8528–8535.
Zhou FM, Hablitz JJ (1999). Activation of serotonin receptors modulates synaptic transmission in rat cerebral cortex. J Neurophysiol 82: 2989–2999.
Acknowledgements
We thank Professor Trevor W Robbins for helpful discussion of these studies and for his comments on the manuscript, Dr S Kongsamut (Aventis Pharmaceutical, USA) for the generous gift of M100907. Funding for these studies was provided by the Italian Ministry for University and Research (MIUR) Grant RBAU01ZS5C.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Carli, M., Baviera, M., Invernizzi, R. et al. Dissociable Contribution of 5-HT1A and 5-HT2A Receptors in the Medial Prefrontal Cortex to Different Aspects of Executive Control such as Impulsivity and Compulsive Perseveration in Rats. Neuropsychopharmacol 31, 757–767 (2006). https://doi.org/10.1038/sj.npp.1300893
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.npp.1300893
- Springer Nature Switzerland AG
Keywords
This article is cited by
-
The Therapeutic Effects of Classic Psychedelics in Alcohol Use Disorder
Current Addiction Reports (2024)
-
5-MeO-DMT: An atypical psychedelic with unique pharmacology, phenomenology & risk?
Psychopharmacology (2023)
-
Brain 5-HT2A receptor binding and its neural network related to behavioral inhibition system
Brain Imaging and Behavior (2022)
-
Impulsive prepotent actions and tics in Tourette disorder underpinned by a common neural network
Molecular Psychiatry (2021)
-
Δ-9-Tetrahydrocannabinol and Cannabidiol produce dissociable effects on prefrontal cortical executive function and regulation of affective behaviors
Neuropsychopharmacology (2019)