
Abstract
Eutrophication is a main threat to continental aquatic ecosystems. Prevention and amelioration actions have been taken under the assumption of a stable climate, which needs reconsideration. Here, we show that reduced precipitation can bring a lake ecosystem to a more productive regime even with a decline in nutrient external load. By analyzing time series of several decades in the largest lake of the Iberian Peninsula, we found autocorrelated changes in the variance of state variables (i.e., chlorophyll and oxygen) indicative of a transient situation towards a new ecosystem regime. Indeed, exceptional planktonic diatom blooms have occurred during the last few years, and the sediment record shows a shift in phytoplankton composition and an increase in nutrient retention. Reduced precipitation almost doubled the water residence time in the lake, enhancing the relevance of internal processes. This study demonstrates that ecological quality targets for aquatic ecosystems must be tailored to the changing climatic conditions for appropriate stewardship.
Similar content being viewed by others

Introduction
Eutrophication has been the main worldwide concern for inland aquatic ecosystems during the last decades since the Great Acceleration1,2. Amelioration, restoration, and prevention actions have proliferated across countries as the problem was recognized: initially, with initiatives to reduce nutrient loadings3 and, progressively, develo** sophisticated regulations to achieve sustainable water and ecosystem quality targets4. Climate change challenges these actions, formerly conceived under the assumption of a stable climate at human social scales5. The knowledge of climate and nutrient dynamics interactions is still limited, particularly concerning their combined influence as drivers of aquatic communities6,7. This understanding is becoming more urgent as climate change progresses and aquatic systems are affected across planet8. Anticipating ecosystem regime shifts is a socially urgent and scientifically challenging issue under currently shifting environmental conditions9. The climatically driven transition and eventual new situation could be socially perceived as a failure in water quality management without necessarily being the case. Traditional monitoring and new observation systems10,11,12 of lake key state variables may provide sufficient mid-term data to characterize and sometimes anticipate significant ecosystem transitions driven by climate shifts.
Any lake ecosystem shows a dynamic state (regime) with characteristic stochastic fluctuations and cycles resulting from internal feedback at the ecosystem scale in response to a hierarchical coupling with the external drivers at some characteristic time scales13. Lake communities reflect the ecosystem regime14. The two primary external drivers are climate forcing and nutrient loading from the catchment, each with their own dynamic regimes. Changes in external nutrient loading regimes have shown rapid lake responses, evidencing eutrophication and recovery within decadal scales15,16,17. On the other hand, paleolimnological records show examples of lake regime shifts at centennial to decadal scales coupled with major climatic transitions14. Nevertheless, whether relatively mild climatic shifts occurring over a few decades can drive a lake regime shift without increasing external nutrient loading, only modifying the internal lake dynamics, is unclear and difficult to evaluate exclusively based on sedimentary records.
Air temperature is increasing throughout the planet, and the focus on its impact on aquatic ecosystems has been mainly on warming effects18,19. Precipitation is usually considered in a more extended temporal framework than air temperature20. Undoubtedly, a large reduction in water inflow to a lake, affecting the water mass balance, will cause an extraordinary ecosystem shift21. However, in areas of relatively high precipitation, external nutrient loading may still be considered the main driver of the lake ecosystem state. Nevertheless, declining precipitation in these areas, although not altering lake level, may substantially modify lake water renewal, affecting nutrient retention, water column recycling, and internal loading from sediments, thus changing lake productivity and metabolism22. Consequently, it can be hypothesized that reduced precipitation could bring a lake ecosystem to a new, more productive regime, even with a decline in external nutrient loading. If the precipitation decline is incremental over decades, it may happen that the lake ecosystem experimented a gradual transition or an abrupt, rapid regime shift. The second case is of particular interest in the context of water quality management.
A regime shift is the process by which an ecosystem rapidly changes from one fundamental state to another. There is debate about the classification of these ecosystem rapid transitions concerning the mechanisms driving them9,23 and whether the regime shift can always be anticipated23,24,25. However, there is agreement that transitions should be indicated by ecosystem state variables showing a shift in their probability space, reflected in the autocorrelation patterns of their variance26. The critical aspect is the selection of those state variables and the appropriate temporal scale of observation and sampling frequency27. Increasing variance and autocorrelation in time series are early warnings of ecosystem shift13. In particular, conditional heteroscedasticity, which means that variance changes in an autocorrelated way, is a powerful indicator of regime shifts28. Variance in a time window step is not independent of the variance in the previous step. That is, periods of high variance are likely to follow periods of high variance, and conversely, those of low variance are more likely to be preceded by periods of low variance. Therefore, the potential non-linear effects of declining precipitation on lake ecosystems should be indicated by conditional heteroscedasticity in the time series of central ecosystem state variables such as chlorophyll, phytoplankton biomass, deep oxygen levels, or any other variable representative of the entire ecosystem metabolism. In the management context, when indications of significant conditional heteroscedasticity are found in monitoring time series of such key variables, the potential drivers have to be investigated to evaluate the mechanism that can be acting behind the abrupt change. Long-term monitoring of lakes has been extremely valuable for understanding eutrophication and the effectiveness of restoration actions by external nutrient loading reduction29,30. Nowadays, it can also provide the basis for a solid understanding of the effects of climate change31. Here, we describe a case study showing how a progressive decline in precipitation can produce an increase in lake productivity by enhancing the internal nutrient loading and shifting the ecosystem to a new regime. In the discussion, we consider the necessary general conditions required to increase lake productivity when the external nutrient load declines by enhancing the use efficiency and recycling of the nutrients.
Over thirty years of monthly monitoring in Lake Sanabria (northwestern Spain, 42°7′12.2″N, 6°42′27.9″W, 1004 m a.s.l, Fig. 1) have indicated a stable state in this oligotrophic lake until recently. The stability was not unexpected because the human population in its catchment is low; water treatment plants were built in the main villages during the 1990s; land use, primarily extensive livestock farming, progressively declined since the mid-twentieth century; and the atmospheric nitrogen deposition in the area has been historically low32. Surprisingly, sudden planktonic diatom blooms occurred in isolated years (2013 and 2017), embedded within apparently "normal" annual cycles. The blooms were of a single diatom species in each case (i.e., Tabellaria flocculosa and Asterionella ralfsii var americana, respectively). The puzzling situation raised social and scientific interest immediately after the first diatom proliferation, notably because Lake Sanabria is the largest natural lake in Spain. Since there was no evident proximate driver of the blooms, the question became whether the lake ecosystem was progressively changing to another state and for which reason.

The long-term time series of chlorophyll and oxygen measurements in Lake Sanabria—as primary indicators of lake metabolism—showed significant conditional heteroscedasticity earlier than the diatom blooms happened. The study of the sediment record showed a shift in the phytoplankton community, confirmed by comparing two years of intensive study with previous early phytoplankton records. However, the eventual drivers of the ecosystem regime shift were not evident. Therefore, we contrasted two main hypotheses: (1) increased diffuse external nutrient loading, perhaps related to the increasing afforestation of the catchment or the recreational use, or (2) increased internal nutrient loading and recycling within the lake basin, eventually related to changing climate conditions in the region33. The external nutrient loading trends were evaluated using (1) 30-year monthly nutrient data from the main inflow and (2) modeling external nutrient sources informed by a 2-year intensive sampling of the primary sources in the catchment (i.e., atmospheric deposition, livestock, recreation, runoff, erosion, subsurface flow, wastewater). Saharan dust intrusion events and occasional forest fires were also considered potential change drivers34. No evidence of external nutrient increase was found. Therefore, changes in internal nutrient use and recycling eventually linked to climate shift should be responsible for the lake regime shift. Weather decadal trends indicated a marked decline in precipitation, with no significant trend in average air temperature. The 30-year time series of lake monitoring showed an increase in the water retention time in the lake, which favored a more efficient use of nutrients in the water column, higher storage in the sediments, and conditions for nutrient return from the sediments. Overall, the Lake Sanabria case study provides evidence that progressive decline in precipitation driving incremental changes in water residence time may result in abrupt changes in lake productivity, resulting in a lake ecosystem regime shift.
Results
Lake metabolism regime shift
The indicators of lake metabolism, such as chlorophyll and oxygen levels, have shown changing patterns of interannual variation during the last decade compared to the previous three decades of lake monitoring (Fig. 2). The water column-integrated chlorophyll achieved values several times higher than the usual long-term range—of 5 and threefold in 2013 and 2017, respectively. Chlorophyll maxima, which usually occurred relatively deep in the water column (12–20 m), also increased the concentration with similar ratios. The phytoplankton blooms were primarily due to a single but different diatom species in each period. For 2013, ad hoc surveys indicated that the bloom was due to Tabellaria flocculosa, and monthly assessments during 2017 showed that the proliferation was, in essence, of diatoms (Fig. S3), mostly Asterionella ralfsii var americana even though Tabellaria was also present (Fig. S4). The chlorophyll variation was within a narrow range for over two decades before 2013. From 1995 to 2005, there was a period of slightly lower chlorophyll values and higher water transparency (e.g., > 9 m Secchi disk depths were typical), but the chlorophyll oscillations were similar before and after this period. Although the chlorophyll mean and coefficient of variation did not start to change until 2013 (Fig. 3a), conditional heteroscedasticity showed a significant change earlier, towards 2007 (Fig. 3b), anticipating a potential regime shift.

Time series (1986–2018) in Lake Sanabria of water column integrated chlorophyll (a), and chlorophyll (b), and oxygen (c) variation across depth, based on monthly measurements at every 2.5 m depth in the D02 station in the deepest part of the eastern basin.

Changes in the mid-term statistics of chlorophyll at 15 m depth, where chlorophyll maxima usually occur, and oxygen at ca. 50 m depth, near the bottom of the deepest point. A 36-month rolling window was considered to estimate the statistical descriptors (a,c), and the conditional heteroscedasticity tests28 (b,d): values above the red line indicate significant conditional heteroscedasticity (P < 0.05).
The two exceptional algal blooms did not produce enhanced oxygen saturation within the photic zone, and they were also not apparent in oxygen depletion deeper in the water column. The oxygen time series were flatter than the chlorophyll series (Fig. 2). The only apparent feature in the series was an increase in the frequency of low oxygen years in the deepest layers during summer. Indeed, the time series of oxygen measurements near the bottom showed large fluctuations for the whole period (Fig. 2c). However, only recently (ca. 2016), the series started to show significant conditional heteroscedasticity (Fig. 3b), supporting the anticipation of a regime shift indicated by chlorophyll.
External nutrient loads
The Tera River is the main inflow to the lake (91% of total modeled water inputs). Nutrient concentrations just before the river joins the lake suggest no increase in nutrient loading in recent years (Fig. S5), especially considering the inflow decline during the last decade (see below). In 1998, total phosphorus concentration declined when wastewater treatment plants were implemented in the main villages of the catchment, although dissolved phosphorus maintained similar values. Nitrate concentrations have been relatively stable; they did not decline after the early treatment plant implementation but did so following an improvement in plant management during the last decade. Occasional peaks of nitrate concentration, notably in 2006, were related to forest fires. However, these peaks showed no discernible response in the lake metabolism indicators. In summary, trends of nutrient loads by the main inflow showed an opposite direction to the increased metabolism experienced by the lake in recent times.
There are no indications that small stream inflows and diffuse nutrient loads could be more relevant than direct Tera River inflows. Nevertheless, we performed an overall estimation of the nutrient loads to the lake considering land use, population, and recreational activities in the catchment, atmospheric deposition, and hydrological modeling of runoff and subsurface flows. The current main contribution is soil runoff for phosphorus and nitrogen (Fig. 4). Livestock was the main phosphorus contributor in the 1960s, and its influence has declined progressively since then. The reduction of this activity was associated with a landscape change. Oak forests increased, mainly around the lake, and following the main river course, meadows declined, and crops almost disappeared (Fig. 4a). The landscape change has resulted in a lower nutrient contribution from soils, with a more substantial decline in nitrogen. Although recreational use has recently increased during the summer period (pers. comm., Sanabria Natural Park, based on a vehicle gauging system), the overall human impact in the immediate zones around the lake has declined progressively because of decades of livestock and crop reduction. The current nutrient contribution by atmospheric deposition directly into the lake and catchment runoff is less than 10%. In conclusion, no evidence exists of external nutrient loading that could have fostered higher lake productivity and metabolism during the last decades.

Contrasting land cover situations between 1956 and the present (a) in the Lake Sanabria watershed allows the comparison of phosphorus (b) and nitrogen (c) external load change according to the modeled contribution of primary sources.
Climate and lake hydrodynamics shift
The most apparent climate shift in the Lake Sanabria watershed has been a decline in annual precipitation during the last decades (Fig. 5). For the period 1960–1980, annual values above 2000 L/m2 were common (~ 50%), and years below 1000 L/m2 were infrequent (< 10%). In contrast, during the last decade, annual precipitation rarely has achieved 1500 L/m2, and years with values barely above 500 L/m2 are common (30%). The monthly mean of the average daily temperature does not show any significant trend during the last fifty years (Fig. 5), although maximum (increasing) and minimum (slightly declining) temperatures show opposing significant trends.

Weather trends in the Lake Sanabria area during the last decades. (a) Precipitation at Puente Porto (M02 station), and (b–d) temperature at Puebla de Sanabria (AEMET 2770B station). The latter is located outside the lake watershed but at a short distance. According to the Mann–Kendall trend test, linear tendencies (black line) are significant for precipitation (P = 0.001, n = 666), maximum (P = 0.0006, n = 666), and minimum temperature (P = 0.017, n = 666). The red line indicates a one-year moving average.
These recorded climatic shifts should have had consequences in lake hydrodynamics. At first glance, the heat balance and water column thermal structure did not show remarkable changes (Fig. 6a,b). However, a more detailed consideration showed significant long-term trends in the holomixis patterns. The first date of isothermy was, on average, delayed, and the temperature of complete mixing declined about one degree (Fig. 6c). These changes were related to a stronger stratification indicated by a trend of increasing maximum buoyancy frequency (Fig. 6d) and a slight deepening of this maximum from about 8 to 10 m depth. The onset of the stratification has also been delayed. Consequently, the duration of stratification changed less than the shift in the starting and ending dates.

Lake Sanabria physical trends during the last decades. (a) Water column heat and (b) temperature patterns. (c) First date and temperature of lake isothermy (Mann–Kendall trend test, n = 31, P = 0.005, P = 0.0008, respectively). (d) Highest monthly buoyancy frequency during the stratification period (Mann–Kendall test, n = 32, P = 0.02). (e) Number of times that water renews annually for some selected depth layers across the water column. The black line indicates the average for 1986–2018 and bars above (blue) and below (orange) deviations.
The water column physical changes and the inflow decline increased the lake's average water residence time (Fig. 6e). On average, layers around 10 m depth renew once per year, upper layers about twice yearly, and deep layers renewal time usually takes more than two years (Fig. 6e). However, there was a substantial difference around these average values before and after ca. 2003. Since then, renewal time has been significantly longer, particularly for layers below the seasonal thermocline (~ 10 m depth).
Lake nutrient retention and recycling
During the last decades, external nutrient loading to the lake has declined mainly because of the recession in watershed livestock and the lower inflow. Conversely, the longer water residence time could have increased nutrient retention efficiency, bioavailability, and recycling—that is, internal loading. No direct experimental evidence of these processes was available, but from observations, there were several indicators of enhanced recycling: (a) the lower the Tera River flow, the higher the reduction between inlet and outlet total phosphorus concentration (Fig. S6a). The difference was significant for both stratification and mixing periods, although the reduction was more substantial during the former. (b) Nitrate concentration was systematically higher in the water column than at the lake inflow (Table 1), indicating organic nitrogen recycling and posterior nitrification. The nitrate formation was higher during stratification, and there was a noisy, albeit significant, correlation with phosphorus retention (Fig. S6b). (c) Organic matter was enriched in carbon the deeper the sediment traps were located (Fig. S7), and the difference between the epilimnetic trap (lower C/N) and the rest (higher C/N) was higher during stratification when the C/N ratio was lower, suggestive of higher algal contribution. Furthermore, layers below the photic zone showed two significant complementary trends (Fig. S8) that explain the progressive higher oxygen depletion observed (Fig. 2). On the one hand, oxygen consumption rates increased in deep layers, a few meters above sediments. On the other, layers 30 to 40 m depth did not show this tendency but prolonged their periods of oxygen decline during stratification. Overall, these trends indicate a context of enhanced nutrient recycling and diffusion from the sediments.
Sediment records
The Vega de Tera dam failure about 10 km upstream of the lake on 9 January 1959 caused a flood that deposited a thick sandy-silt layer over the lake basin (Fig. S9). This event sealed the upper biogeochemically active sediment, causing a transient oligotrophication of the lake evidenced by the lower sediment carbon, nitrogen, and, particularly, phosphorus content during the following two decades (Fig. 7a). Similar conditions in phosphorus accumulation to those before the catastrophic event were achieved by the early 1980s. Since 2003, phosphorus accumulation maintained high values, with maxima during the last years. The sediments increased carbon and nitrogen content by the late 1980s, with a progressively more depleted signature in δ13C and δ15N, suggesting higher algal contribution. The P contrasting patterns with C and N during 1960–1990 indicated a progressive building of a P internal loading capacity.

Sediment indicators of main functional and community changes in Lake Sanabria during the last decades (a) Organic carbon, total nitrogen, and total phosphorus sediment accumulation in the deepest part of the lake. Carbon and nitrogen stable isotopic compositions are also indicated (red line). Note the marked 1959 oscillation corresponding to the Vega de Tera dam collapse that reset the lake's biogeochemical dynamics. See Fig. S7 for the depth-age model. (b) Main chrysophyte cyst changes. (c) Diatom remains (Asterionella ralfsii var americana, Tabellaria flocculosa, Aulacoseira subborealis/pseudodistans, and Cyclotella stelligera), and representative algal pigments (pheophytin-a, bulk phytoplankton indicator; lutein, chlorophytes; echinenone, cyanobacteria; alloxanthin, cryptophytes).
Some key indicators summarize the main changes in the phytoplankton community since 1959 (Fig. 7b,c). Pheophytin-a indicates overall algal biomass; the values were fairly low until ~ 1990 and have risen since then. Lutein and echinenone, pigments respectively exclusive of chlorophytes and cyanobacteria, followed the same trend. Alloxanthin, a cryptophytes' indicator, also increased recently, confirmed by α-carotene (not shown), another pigment related to cryptophytes. Alloxanthin also showed high values, indicative of cryptophyte blooms, during the decade following the dam failure. The general patterns in the sediment record agreed with two periods of direct phytoplankton observations in the water column. Comparison between 1987 and 198935 and our study 2015–2017 monthly observations (Fig. S10) indicated a decline in flagellated groups (chrysophytes, cryptophytes, and dinoflagellates) and an increase in chlorophytes, xanthophytes, and cyanobacteria.
Diatom valves and chrysophyte cysts in the sediments indicate highly fluctuating populations (Fig. 7b,c). Nonetheless, some species have declined (e.g., Cyclotella stelligera, cysts S046, S101, S050/52) during the period in which the pheophytin-a was increasing. Conversely, a few species markedly peaked during the last decade. T. flocculosa was the most conspicuous, and, more recently, A. ralfsii, both observed in the water column survey. Chrysophyte cyst S189 has increased since the 1990s and recently peaked. During the 2015–2017 study, we found this cyst within colonies of Stichogloea doederleinii, a coccal phaeothammniaceae species, which commonly peaks towards the end of the stratification period. The temporal stratification displacement may favor the Stichogloea-associated chrysophyte that has not yet been identified.
In summary, the sediment record indicates an increasing nutrient export to the sediments and a shifting phytoplankton community towards assemblages indicative of slightly higher productivity or, at least, different productivity patterns.
Discussion
Eutrophication symptoms in lake ecosystems are usually associated with increasing external nutrient loads36. In Lake Sanabria, this has not been the case. On the contrary, nutrient loads were lower during the last decades because of livestock decline, forest expansion, improved wastewater treatment, and reduced river flow. Atmospheric deposition represents a minimal contribution of nutrients, and aerosol analyses indicated a low influence of anthropogenic activities in the area34. Conversely, there were indications that the increased lake productivity was related to a higher internal nutrient load, exemplifying the complex interplay between physical and biological mechanisms37.
We found that lower river flows resulted in higher nutrient retention efficiency and cycling processes, such as nitrification in the water column. Furthermore, progressively declining flushing rates during the last decades increased carbon, nitrogen, and phosphorus accumulation rates in the sediments that, associated with higher oxygen consumption rates during stratification, favor the nutrient diffusion from sediments, modifying the feedbacks of the biogeochemical dynamics of the lake38. Considering the small C/N sediment range during the last decades, indicative of low variability in terrestrial vegetation inputs, the recent δ13C decrease in Sanabria was likely attributed to higher lake primary productivity39,40, as also indicated by the shape of the P upper sediment profile41. The post-1980 decreasing trend in δ15N was coherent with higher productivity in a non-limited N environment facilitated by a higher internal sediment recycling42. Therefore, the lake's shifting trophic conditions were ultimately related to climate and the hierarchical links with proximate drivers43.
Climate is a multi-variate primary driver sha** the probability space in which the lake ecosystem oscillates44. If the regional climate changes, the mechanistic cascading from climate to the lake is affected, and, as a consequence, the stationary lake regime may be modified45. A regime shift is generally understood as a relatively sharp (non-linear) ecosystem change compared to more progressive (linear) trends in the drivers46. In Lake Sanabria, the climatic shift mainly consists of a persistent decline in precipitation during the last decades, which has been associated with the North Atlantic Oscillation and Scandinavian atmospheric circulation modes47. The declining flushing rates did not apparently produce lake ecosystem changes until the sudden diatom blooms. However, the analysis of variation of chlorophyll already anticipated that the transition started before.
Chlorophyll and oxygen concentrations are simple but powerful and complementary variables to characterize the lake's metabolic state. The chlorophyll conditional heteroscedasticity test anticipated the later more conspicuous changes in Lake Sanabria. The shift indicator was significant in 2007, whereas diatom blooms in the water column appeared in 2013 and 2017. Oxygen in deep layers showed a delayed response, which makes sense in a variable that integrates more extended periods. The sediment record confirmed a reorganization of the phytoplankton community beyond the apparent peaks of the two diatom species. The shifts in the sediment indicators were apparent by 2008, again before the blooms.
Sanabria changes in the lake community were most conspicuous in a few species, but sediment pigment records indicate that other groups change in abundance. Historical data for short periods and occasional annual samples52 compared to the biannual intensive recent study are inconclusive, beyond the diatom case, because of the high species richness and seasonal fluctuations. Nevertheless, provided the significant changes in relative dominance at high taxonomic levels (Fig. S10), a relevant shift in species composition is very likely.
We may wonder if the lake is shifting to a new regime without precedents or is switching to a state that has been present before. Indeed, Sanabria's Holocene sediment records show periods of high Tabellaria flocculosa accumulation, the latter about 3200 years ago, which lasted for ~ 800 years48. There were other similar periods in the Mid and Early Holocene. Interestingly, past T. flocculosa peaks occurred when geochemical and sedimentological indicators point to lower precipitation48. Therefore, we might hypothesize that the lake is shifting to a state already visited long ago. However, the temporal scales between the Holocene sediment record and the recent changes are substantially different, warning against using past fluctuations as direct analogs of the current situation.
Even in the case of similar past situations, it would not mean that the lake should oscillate between two alternative equilibria and, thus, be in a critical transition situation at present49. Some criteria have to be fulfilled to infer that a regime shift involves a critical transition23: (1) an abrupt shift in the time series has to occur, (2) driven by positive feedback mechanisms, (3) in response to an incremental increase in control parameters, which (4) results in a bimodal distribution of state variables, and (5) displays hysteresis in the transitions50. Some of these requirements are accomplished in the L. Sanabria case if it is assumed that the control parameter is the water renewal time and the feedback relies on nutrient cycling. Bimodality is incipient because the period of high chlorophyll values is shorter than the period of low, so at present, there is only a fat cue in the high values of the probability density function of the chlorophyll record. Hysteresis would require a more extended time series for an accurate test, with some back-and-forth oscillation between low and high precipitation years. Indeed, there is much debate about whether bimodality and the alternative state concept help to explain long-term patterns in lakes, even in the shallow lakes where the theory was inspired51, the temporal scale of observation being critical. The conditional heteroscedasticity test of state variables is a valuable operational option for anticipating an abrupt shift in lake ecosystems26. Understanding the limnological, environmental, and climatic mechanisms involved will provide the cues for adequate management.
The case of Lake Sanabria demonstrates that increased internal nutrient loading resulting from longer water residence time in a lake can drive an ecosystem regime shift towards a more productive state, even in cases with declining total external nutrient loading because of reduced precipitation. To generalize, a deliberately simple model of nutrient cycling within a lake (Appendix S1) helps to understand that reduced precipitation could more likely increase productivity in lake ecosystems when (1) the relative decline in external loading is lower than the precipitation decline, that is, nutrient concentrations in the inflow are maintained or increase; (2) there is an increase in the efficiency of the nutrient loading conversion to living biomass within the water column; for instance, by increasing the time for phosphatase action53, mineralization of organic nitrogen54 or recycling of refractory materials55, and (3) by enhancing exchanges with the sediments56 because of longer water residence time, favoring higher sedimenting biomass proportions, or more extended periods of nutrient release to the water column. Not all these requirements must be fulfilled, but they must be met in adequate relative proportions (Appendix S1). In the end, what is relevant is the relative (not absolute) shift in the recycling processes with respect to the previous state. Consequently, those lakes with initial low productivity yield of the external nutrient load or relatively low internal recycling will be more prompt to significant changes by reduced precipitation in their watersheds.
In Lake Sanabria, a large part of P and N loading is not in the form of simple inorganic compounds readily assimilable. Therefore, the increased residence time of water may enhance the use of a nutrient fraction that otherwise will circulate through the system in high-flow conditions. In other lakes, inorganic forms of limiting nutrients may be more relevant in the external loading and, thus, have less capacity to compensate for the decline in external loading by increasing assimilation efficiency57.
On the other hand, longer water residence time enhances the fraction of water column biomass that can sink to sediments and be recycled. A large proportion of sedimented P is in insoluble fractions, and only some (i.e., metal oxides) will release exchangeable P under reduced conditions58. The release will increase the longer the stratification period and the higher the oxygen demand in deep waters, as in Lake Sanabria. In Lake Sanabria, the water residence time during rainy periods was already much higher than the characteristic time for biomass sedimentation (Appendix S1). Consequently, the lake acted, already in the past, as an efficient trap with a limited margin for increase, and the sediment record reflected lake productivity. The enhanced exchange between water and sediment should not be restricted to deep waters; warming and reduced oxygen may happen across the benthic system, enhancing nutrient release and algae production56. Lakes experiencing more extended changes in water column stratification than Lake Sanabria may show a more substantial contribution by sediment nutrient release38.
The documented Lake Sanabria transition and the eventual new situation could be socially perceived as a failure in water quality management, particularly if blooms reach the surface, as happened in 2013. Many European countries have adopted the concept of reference conditions to determine the ecological states of their aquatic ecosystems, lakes in particular59. The reference conditions for a good ecological state are established under the assumption that lakes without human influence maintain communities and biogeochemical processes within a stationary range of oscillations driven by climate. Operatively, the concept was implemented by defining reference sites with low human impact, which provided an acceptable range of variation for lakes within the same category. The statistics could come from time series in those sites or from a statistical population of lakes considered to be of the same functional category4. Lake Sanabria was defined as unique in its category and became its own reference site, provided the human impact in the watershed was low52. Under changing climate, lake reference conditions will require overall revision, especially regarding productivity indicators such as phytoplankton and chlorophyll. Lake Sanabria provides a paradigmatic example of the difficulties in defining shifting reference conditions because climate change and the lake transition may last for decades and move towards no analog scenarios60. It could happen that there is not a shift from a stationary state to another but a permanent nonstationary state.
Consequently, authorities and administrations should shift from managing the state to managing change. Stewardship measures should complement, occasionally replace, conservation and restoration targets61. Those measures require understanding the drivers cascading from global climate to within lake processes and develo** management safe-operating spaces to avoid undesirable watershed human actions that may synergistically interfere with shifting climate effects in unwanted ways. Non-stationarity in climate demands new theoretical and operative ways to deal with ecological dynamics62. The Lake Sanabria case demonstrates that ecological quality targets of aquatic ecosystems must be tailored to the new climatic conditions, even in localities with no remarkable water quality pressures. This study shows that precipitation decline in rainy regions can bring some lake ecosystems to the edge of regime shifts by modifying seasonal hydrodynamics, nutrient recycling, and internal loading.
Methods
Study site
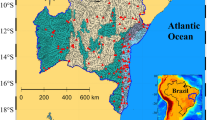
Lake Sanabria (42°7′12.2″N, 6°42′27.9″W) constitutes a unique ecosystem among the inland waters of Spain, being the largest natural lake in the Iberian Peninsula (area 3.536 km2, volume 99.114 hm3). The lake has a relatively small watershed (~ 122.16 km2) (Fig. 1, S1), mainly of granitic bedrock, with the highest altitude at 2127 m a.s.l. (Peña Trevinca Peak) and the lake at 1004 m a.s.l. The climate is temperate, with precipitation distributed throughout the year but lower during the warm summer, showing high interannual variability (Table 1). The Tera River constitutes the main inflow, which since the late 1950s has been affected by the discharge of the hydroelectric power system of Moncabril in one of its main tributaries. In addition, small streams and underwater springs are discharging in the lake. Oak forests and meadows associated with small villages constitute the dominant vegetation around the lake. Above 1500 m a.s.l., scrublands, mountain pastures, and significant areas of peat bogs are the predominant categories. Lake Sanabria morphology is the result of glacier over-excavation at the bottom of a U-shaped cross-section valley closed at its eastern end by a terminal moraine63. The lake presents two sub-basins (Fig. 1e, S10): the western basin, receiving the Tera discharge, is slightly shallower (43 m) than the eastern (50 m), with the outlet. Almost 70% of the lake volume is above 20 m depth and ~ 10% below 30 m; this last volume is exposed to high contact with fine bottom sediments. The lake thermal regime is typically holomictic, with stratification from April to December. The lake has been experiencing epilimnetic autumn warming and hypolimnetic annual cooling during the last decades33. The temperature range in the upper layers is wide (Table 1), with epilimnetic waters > 20 °C during the summer months. According to the bedrock characteristics, ionic strength and acid-neutralizing capacity are low, and pH is circumneutral (Table 1). Chlorophyll, nutrient levels, and transparency indicate an oligotrophic state (Table 1), also reflected in the oxygen levels, which do not achieve elevated over-saturation nor anoxia in the water column. At the end of the stratification period, oxygen minima (~ 20–30% saturation) occur in the lake deepest part. Dissolved organic carbon (DOC) levels are relatively high for temperate mountain lakes (2–3 mg C L−1) because of the forested catchment and some peatlands in the upper catchment. The lake holds remarkable species-rich phytoplankton, zooplankton, and littoral communities64.
Meteorological data
There are several running meteorological stations in the lake watershed (Fig. 1). The longest data series, covering the 1963–2018 period, were in Puente Porto reservoir (M02 station, 42°7′1″N, 6°49′52″W, 1645 m a.s.l), concerning precipitation, and Puebla de Sanabria (AEMET 2770B station, 42°3′15″N, 6°38′2″W, 960 m a.s.l), concerning temperature. The latter is located outside the lake watershed but not far away and shows high coherence with those in the lake's catchment. Gaps in these two meteorological series were filled using nearby stations and the Climatol R package (v 3.1.2)65. For air temperature, there is a high coherence among the regional stations. We used nine extra stations (including M02) to fill the gaps in the AEMET 2770B time series, which showed 22% of missing data. The adjusted R2 between the AEMET 2770B time series and the others was high (mean and median 0.83, min. 0.75, max. 0.9). The spatial coherence for precipitation is lower than for air temperature. However, the number of missing data was very low (7%), and the M02 station is located in the central part of the lake's watershed. We used 85 stations in a 150-km radius from the lake to fill the gaps. The adjusted R2 between the MO2 station and the other stations was uneven (adj. R2 median 0.62, min. 0.13, max. 0.80). The significance of the observed trends was assessed using the Mann–Kendall trend test.
Lake long-term dynamics
Long-term lake monitoring (1986–2018) included monthly inlet and outlet samples and profiles in the eastern sub-basin (station D02) for temperature, conductivity, dissolved oxygen, Secchi disk depth, soluble reactive phosphorus (SRP), total phosphorus (TP), nitrate, silica, and chlorophyll. There was a gap in nitrogen and phosphorus data for some months of 2012 and 2013 due to laboratory equipment problems. Water samples were collected at 2.5 m intervals from the surface to 50 m depth, and the same lab made measurements throughout the period. Additionally, there was a period (2015–2017) of more comprehensive sampling at both sub-basins, including phytoplankton samples and littoral points.
Nitrate and SRP were determined by ion chromatography (940 Professional IC Vario TWO/SeS/PP Metrohm following EPA 300.1 method); total nitrogen (TN) and TP using segmented continuous microflow AutoAnalizer AA3; silica by spectrophotometry (V650 Jasc Standard Methods 4500-SiO2 C Ed 22); and DOC by high-temperature catalytic combustion (Formacs NC Analyzer with Nondispersive infrared detector (NDIR), Skalar). Chlorophyll was determined using Whatman® GF/F fiberglass filters, acetone extraction (90%), and spectrometry, and it was calculated according to Jeffrey and Humphrey66. Phytoplankton samples were fixed with Lugol's solution and counted with a Nikon Eclipse Ti-S inverted microscope following Utermöhl's method. Counting and biovolume determination followed European standards (EN 15204:2006, EN 16695:2015). Taxa were identified under 1000 × light microscopy, and, for diatoms, scanning electron microscopy was additionally used.
The water column's physical structure was considered by calculating the buoyancy frequency (N2) from the monthly temperature and conductivity profiles,
where g is the acceleration due to gravity, ρ is the water density, and z indicates depth. We derived some variables from N2 data to obtain time series that could reflect long-term changes in the lake water column physical dynamics: (a) the isothermy date, determined as the day with the lowest water column Schmidt stability67; (b) temperature at water column isothermy; (c) highest N2 during stratification; and (d) the depth where the highest N2 occurs.
The inflow density indicated that the river mixed with lake water above the thermocline during the stratification period. Therefore, assuming that the epilimnion (and the whole water column during lake isothermy) was mixed and the outflow was exclusively through the lake surface outlet of Tera River, we estimated the number of times each 1 m layer volume was renewed annually. In-lake phosphorus retention was estimated monthly from inlet and outlet concentrations and river flow.
During the stratification, the oxygen consumption in layers below 30 m depth was characterized by two parameters: (1) the duration of the layer isolation, considering the number of days between annual maximum and minimum oxygen values at the measurement depth, and (2) the rate of consumption, estimated from the slope of the regression of the oxygen measurements made during this period against time.
Evaluation of changes in catchment nutrient sources
During the last decades, land use and human activities in the catchment have changed markedly. To evaluate whether potential nutrient loads could have increased, we modeled the nutrient loads delivered by the most relevant sources (i.e., direct precipitation, livestock, recreation areas, runoff, stream bed erosion, subsurface flow, and wastewater) for two scenarios corresponding to 1956, for which aerial images and land cover maps were available, and the period of intensive study (2015–2017), which provided information for the model adjustment. The lake's water inflow and nutrient loads were assessed by dynamic modeling on a 5 m spatial resolution digital elevation model68 validated with the intensive biennial sampling of catchment tributaries and atmospheric deposition. The hydrological model was calibrated and validated in two years of contrasting precipitation, 1620 L/m2 in 2016 and 1010 L/m2 in 2017. In each of these years, about 65% of the data was used for calibration, and the remaining 35% was used for validation. The nutrient export model was calibrated by land cover information and measurements at different points of the catchments (Fig. 1) and validated using nutrient data (a total of 21) from the river Tera at the lake inlet.
The watershed and stream network river delineation was computed using TauDEM (Terrain analysis using digital elevation models) tools69, implemented in QGIS-v2.870. The river network was subsequently adapted to the reality of the terrain by checking on orthophotos and maps (i.e., channel rooting) and field observations (i.e., minimum order of the active channel). A total of 26 discontinuity points were established considering hydro-morphological and land use discontinuities: stream confluences, diversions and canals, reservoirs, primary land use typologies, and the main (Tera River) and secondary watercourses draining into the lake. The contribution areas to these points define basins or hydrologic response units, with areas ranging from approximately 20 ha to a maximum of 2500 ha.
The hydrological and nutrient load modeling was performed using MapShed (v1.5.1)71, which is a GIS-based tool that incorporates the generalized watershed loading functions (GWLF) model and enhances the functionality using a free GIS software package (MapWindow). MapShed combines hydrology, land cover, soils, topography, weather, pollutant discharges, and other data to determine the amount of daily nutrient and sediment loads in small watersheds. It is considered a combined distributed/aggregated catchment model and has been successfully applied in rural watershed studies72. The model simulates surface runoff using the curve number equation73 based on daily precipitation and temperature data and distinguishes between direct runoff and infiltration based on rainfall, soil, and land cover data. For each contributing area, a daily water balance was made based on precipitation, snowmelt, initial state and capacity of the unsaturated zone, and evapotranspiration. Streamflow consisted of total watershed runoff from all source areas plus groundwater discharge from a shallow saturated zone, also connected with a deep saturated zone (aquifer). Both fluxes were modelled by means of recession coefficients. It was assumed that there was no deep seepage connection with a regional aquifer system consistent with the glacial character of the basin. The solid fraction contribution was estimated as the product of the monthly sediment load and the average sediment nutrient concentration. Erosion was estimated using the universal soil loss equation (USLE algorithm), which used monthly values of the soil erodibility, topographic, cover and management, and soil and water conservation coefficients for each generating area, i.e., each combination of land use/cover and soil type. The sediment delivery ratio, the critical factor for computing sediment yield, was based on the watershed size and transport capacity.
Dissolved nutrient load and sediment transport through rural areas were computed by multiplying their respective coefficients by runoff. In the GWLF, all the N and P from the urban areas were considered to be in a solid state, and the model used exponential accumulation and wash-off function to estimate urban loading. The sub-surface losses in the watershed were estimated using dissolved N and P concentrations, where the watershed was considered a single lumped-parameter contributing area. The 5-cm upper soil was sampled and chemically analyzed twice (winter and summer) in six plots in catchment areas representative of the main land uses (i.e., grassland, scrubland, oak forest, and pastures with low and medium cattle pressure). The soil water content was measured and chemically analyzed at these plots, and an extra one was located in a peatland area at 20 and 40 cm depths for one year. These measurements were used to calibrate the soil and hydro-chemical module for erosion, runoff, and subsurface flows.
Livestock farming and urban wastewater contributions were accounted for in each lake watershed basin. Livestock comprised beef cattle and sheep in a seasonal grazing mode and was considered in the "open land" areas where it was known to be present. The model estimates contributions by basin using numbers and weight for each animal type. Nitrogen and phosphorus produced by farm animal populations are transported to nearby water bodies as losses due to animal grazing. The transport includes runoff from grazing land and direct deposits to streams where access is unimpeded.
The nitrogen and phosphorus loads from point sources were estimated considering two wastewater effluents from unitary sewage treatment systems, with direct discharge to the river Tera, 100 and 600 m upstream of the lake, and another discharging to the terrain at a distance 100 m to the lake coast. They were secondary treatments with an activated sludge process. Following Evans and Corradini71, a default nutrient retention factor was applied to the total exported loads to those basins with reservoirs as outlets.
Although the GWLF model could be used without calibration74, a tailored calibration technique for some relevant parameters and specific values for the Sanabria watershed was applied (Table S1). The model performance was evaluated by comparison with measurements at the river Tera inlet using data from November 2015 to September 2017, daily for discharge and monthly for nutrient concentration (Table S2). The agreement with the observed data was satisfactory (Fig. S11). For instance, the Nash–Sutcliffe efficiency statistics (NSE) showed values of 0.61 for river discharge, 0.64 for TP concentration, and 0.61 for TN (Table S2, Fig. S11). Generally, the simulation is considered satisfactory for this model when NSE > 0.575.
The parameterized model was applied to two different periods: the year 1956 and the intensive sampling period (2015–2017), with different scenarios regarding weather, land use, population (both inhabitants and visitors), and wastewater treatments. Land use in 1956 was determined by performing a supervised classification of orthoimagery generated from the American Army Map Service flights from January 1956 to November 1957, available at the Spanish National Center for Geographic Information (CNIG) (https://www.ign.es/web/en/ign/portal/qsm-cnig).
The hydro-chemical module was fed with nutrient deposition data from measurements of the gaseous, particulate, and dissolved-in-rain concentration of the main chemical forms of N and P (ammonia, nitrogen oxides, nitric acid vapor, particulate, and dissolved ammonium nitrate, phosphate, and total phosphorus), performed for the period June 2016–December 2017, following Garcia-Gomez, Izquieta-Rojano76. Measurement plots were located at M01 (lower-altitude area, with human presence) and M02 (upper area, with occasional human presence). The results were extrapolated to the entire catchment, considering altitude, sub-basin, land use, and precipitation. Atmospheric inputs were introduced with daily resolution in the hydro-chemical module as (1) a contribution to the concentration of inorganic N forms in each sub-basin runoff and (2) direct inputs of inorganic N and P to the lake.
Sediment traps and sediment record
Cylindrical sediment traps (Hydro-Bios Saarso type) were installed at three depths (6, 14, and 40 m) in the deepest area of the western sub-basin. Several surface sediment cores were obtained at the deepest area of the eastern sub-basin using a Glew-type gravity corer and were sliced at 2 mm intervals. An age model was obtained using Pb-210 following Sanchez-Cabeza, Masque77. Subsamples from each core slice or trap material samples were analyzed to determine: TP (segmented continuous microflow Autoanalizer AA3); TN and total organic carbon (TOC) (high-temperature catalytic combustion by Skalar Primacs SNC Analyzer); ẟ15N/ẟ13C (Flash Dynamic Combustion method by Thermo Scientific FlashEA1112 Thermo-Finnigan Nitrogen and Carbon analyzer with DELTA plus (Finnigan MAT) isotope ratio mass spectrometer); and pigments (high-performance liquid chromatography with diode array detection (HPLC 1200—G1315D, Agilent Technologies), previous lyophilization of sediment subsamples and extraction with acetone:methanol:water (80: 15: 5), using trans-β-Apo-8′-carotenal as an internal standard solution, sonication, and centrifugation). Furthermore, diatom clean frustules and chrysophyte cyst suspensions were obtained by oxidizing organic matter with hot hydrogen peroxide 30% v/v, and microscopic slides were mounted using a refractive resin (Naphrax®). Taxa were identified under 1000 × light microscopy (LM) with a 100X/1.4 Zeiss Pan-Apochromat objective mounted in a Zeiss scope-A1 microscope equipped with Differential Interference Contrast (Nomarski) optics and dual oil immersion).
Ecosystem regime shift
The Lagrange multiplier test for conditional heteroscedasticity28 was used to determine whether the lake ecosystem could be in a critical transition. This test consists of (1) establishing whether there is an interannual trend in the series and, if so, detrending it; (2) fitting a time series model (Autoregressive integrated moving average, ARIMA) to the series without trend; (3) calculating the squared residuals of the series based on the optimized model; (4) regressing the squared residuals on themselves lagged one-time step; and (5) checking the slope between the two residuals: if the slope is 0 or negative, there is no conditional heteroscedasticity; if the slope is positive, the correlation coefficient of the regression is multiplied by the sample size (n); (6) finally, the value obtained in the previous step is compared with a χ2-distribution with one degree of freedom.
Data availability
The data that support the findings are available in Dryad at "Lake Sanabria ecosystem shift (1986–2019)", https://doi.org/10.5061/dryad.3j9kd51q2
References
Smith, V. H. Eutrophication of freshwater and coastal marine ecosystems a global problem. Environ. Sci. Pollut. R. 10, 126–139 (2003).
Naderian, D. et al. A water quality database for global lakes. Resour. Conserv. Recycl. 202, 107401 (2024).
Lewis, W. M. Jr., Wurtsbaugh, W. A. & Paerl, H. W. Rationale for control of anthropogenic nitrogen and phosphorus to reduce eutrophication of inland waters. Environ. Sci. Technol. 45, 10300–10305 (2011).
Birk, S. et al. Three hundred ways to assess Europe’s surface waters: An almost complete overview of biological methods to implement the Water Framework Directive. Ecol. Indic. 18, 31–41 (2012).
Kopf, R. K., Finlayson, C. M., Humphries, P., Sims, N. C. & Hladyz, S. Anthropocene baselines: Assessing change and managing biodiversity in human-dominated aquatic ecosystems. Bioscience 65, 798–811 (2015).
Adrian, R. et al. Lakes as sentinels of climate change. Limnol. Oceanogr. 54, 2283–2297 (2009).
Huang, S., Zhang, K., Lin, Q., Liu, J. & Shen, J. Abrupt ecological shifts of lakes during the Anthropocene. Earth Sci. Rev. 227, 103981 (2022).
Gilarranz, L. J., Narwani, A., Odermatt, D., Siber, R. & Dakos, V. Regime shifts, trends, and variability of lake productivity at a global scale. Proc. Natl. Acad. Sci. U. S. A. 119, e2116413119 (2022).
Capon, S. J. et al. Regime shifts, thresholds and multiple stable states in freshwater ecosystems: A critical appraisal of the evidence. Sci. Total Environ. 534, 122–130 (2015).
Mozafari, Z., Noori, R., Siadatmousavi, S. M., Afzalimehr, H. & Azizpour, J. Satellite-based monitoring of eutrophication in the Earth’s largest transboundary lake. Geohealth 7, e2022GH000770 (2023).
Dörnhöfer, K. & Oppelt, N. Remote sensing for lake research and monitoring—recent advances. Ecol. Indic. 64, 105–122 (2016).
Marcé, R. et al. Automatic high frequency monitoring for improved lake and reservoir management. Environ. Sci. Technol. 50, 10780–10794 (2016).
Scheffer, M. et al. Early-warning signals for critical transitions. Nature 461, 53–59 (2009).
Spanbauer, T. L. et al. Body size distributions signal a regime shift in a lake ecosystem. Proc. R. Soc. B Biol. Sci. 283, 20160249 (2016).
Zhao, J., Gao, Q., Liu, Q. & Fu, G. Lake eutrophication recovery trajectories: Some recent findings and challenges ahead. Ecol. Indic. 110, 105878 (2020).
Schindler, D. W. The dilemma of controlling cultural eutrophication of lakes. Proc. R. Soc. B Biol. Sci. 279, 4322–4333 (2012).
Anderson, N. J., Jeppesen, E. & Sondergaard, M. Ecological effects of reduced nutrient loading (oligotrophication) on lakes: An introduction. Freshw. Biol. 50, 1589–1593 (2005).
Oleksy, I. A., Baron, J. S., Leavitt, P. R. & Spaulding, S. A. Nutrients and warming interact to force mountain lakes into unprecedented ecological states. Proc. Biol. Sci. 287, 20200304 (2020).
Chang, C. W. et al. Long-term warming destabilizes aquatic ecosystems through weakening biodiversity-mediated causal networks. Glob. Change Biol. 26, 6413–6423 (2020).
Linderholm, H. W. et al. Arctic hydroclimate variability during the last 2000 years: Current understanding and research challenges. Clim. Past 14, 473–514 (2018).
Micklin, P. The past, present, and future Aral Sea. Lakes Reserv. Sci. Policy Manag. Sustain. Use 15, 193–213 (2010).
Schindler, D. W. Recent advances in the understanding and management of eutrophication. Limnol. Oceanogr. 51, 356–363 (2006).
O’Brien, D. A. et al. Early warning signals have limited applicability to empirical lake data. Nat Commun 14, 7942 (2023).
Burthe, S. J. Do early warning indicators consistently predict nonlinear change in long-term ecological data?. J. Appl. Ecol. 53, 666–676 (2015).
Gsell, A. S. et al. Evaluating early-warning indicators of critical transitions in natural aquatic ecosystems. Proc. Natl. Acad. Sci. U. S. A. 113, E8089–E8095 (2016).
Dakos, V. et al. Methods for detecting early warnings of critical transitions in time series illustrated using simulated ecological data. PLOS ONE 7, e41010 (2012).
Spears, B. M. et al. Ecological instability in lakes: A predictable condition? Environ. Sci. Technol. 50, 3285–3286 (2016).
Seekell, D. A., Carpenter, S. R., Cline, T. J. & Pace, M. L. Conditional heteroskedasticity forecasts regime shift in a whole-ecosystem experiment. Ecosystems 15, 741–747 (2012).
Edmondson, W. T. Phosphorus, nitrogen, and algae in Lake Washington after diversion of sewage. Science 169, 690–691 (1970).
May, L., Olszewska, J., Gunn, I. D. M., Meis, S. & Spears, B. M. Eutrophication and restoration in temperate lakes. IOP Conf. Ser. Earth Environ. Sci. 535, 012001 (2020).
Gerten, D. & Adrian, R. Climate-driven changes in spring plankton dynamics and the sensitivity of shallow polymictic lakes to the North Atlantic Oscillation. Limnol. Oceanogr. 45, 1058–1066 (2000).
Garcia-Gomez, H. et al. Nitrogen deposition in Spain: Modeled patterns and threatened habitats within the Natura 2000 network. Sci. Total Environ. 485, 450–460 (2014).
Ramos-Fuertes, A. et al. Thermal response of Sanabria Lake to global change (NW Spain). Limnetica 39, 455–468 (2020).
Pérez-Pastor, R. et al. Characterization of organic aerosols at the Natura 2000 remote environment of Sanabria Lake (Spain): Evaluating the influence of African dust and regional biomass burning smoke. Atmos. Environ. 298, 119634 (2023).
De Hoyos, C. Limnología del Lago de Sanabria. Variabilidad interanual del fitoplancton.) (Universidad de Salamanca, 1996).
Smith, V. H. & Schindler, D. W. Eutrophication science: Where do we go from here?. Trends Ecol. Evol. 24, 201–207 (2009).
Genkai-Kato, M. & Carpenter, S. R. Eutrophication due to phosphorus recycling in relation to lake morphometry, temperature, and macrophytes. Ecology 86, 210–219 (2005).
Tu, L. et al. The nexus among long-term changes in lake primary productivity, deep-water anoxia, and internal phosphorus loading, explored through analysis of a 15,000-year varved sediment record. Glob Planet Change 207, 103643 (2021).
Gu, B., Chapman, A. D. & Schelske, C. L. Factors controlling seasonal variations in stable isotope composition of particulate organic matter in a softwater eutrophic lake. Limnol. Oceanogr. 51, 2837–2848 (2006).
Teranes, J. L. & Bernasconi, S. M. Factors controlling δ13C values of sedimentary carbon in hypertrophic Baldeggersee, Switzerland, and implications for interpreting isotope excursions in lake sedimentary records. Limnol. Oceanogr. 50, 914–922 (2005).
Carey, C. C. & Rydin, E. Lake trophic status can be determined by the depth distribution of sediment phosphorus. Limnol. Oceanogr. 56, 2051–2063 (2011).
Leng, M. J. et al. Isotopes in Lake Sediments (Springer, New York, 2006).
Ryo, M., Aguilar-Trigueros, C. A., Pinek, L., Muller, L. A. H. & Rillig, M. C. Basic principles of temporal dynamics. Trends Ecol. Evol. 34, 723–733 (2019).
Woolway, R. I. et al. Phenological shifts in lake stratification under climate change. Nat. Commun. 12, 2318 (2021).
Woolway, R. I. & Merchant, C. J. Worldwide alteration of lake mixing regimes in response to climate change. Nat. Geosci. 12, 271–276 (2019).
Andersen, T., Carstensen, J., Hernández-García, E. & Duarte, C. M. Ecological thresholds and regime shifts: Approaches to identification. Trends Ecol. Evol. 24, 49–57 (2009).
Hernandez, A. et al. Sensitivity of two Iberian lakes to North Atlantic atmospheric circulation modes. Clim. Dyn. 45, 3403–3417 (2015).
Jambrina-Enriquez, M. et al. Timing of deglaciation and postglacial environmental dynamics in NW Iberia: The Sanabria Lake record. Q. Sci. Rev. 94, 136–158 (2014).
Scheffer, M., Carpenter, S., Foley, J. A., Folke, C. & Walker, B. Catastrophic shifts in ecosystems. Nature 413, 591–596 (2001).
Ratajczak, Z. et al. Abrupt change in ecological systems: Inference and diagnosis. Trends Ecol. Evol. 33, 513–526 (2018).
Davidson, T. A. et al. Bimodality and alternative equilibria do not help explain long-term patterns in shallow lake chlorophyll-a. Nat. Commun. 14, 398 (2023).
Pahissa, J., Fernandez-Enriquez, C. & de Hoyos, C. Water quality of Lake Sanabria according to phytoplankton. A comparison with historical data. Limnetica 34, 527–540 (2015).
Nedoma, J., Garcia, J., Comerma, M., Šimek, K. & Armengol, J. Extracellular phosphatases in a Mediterranean reservoir: Seasonal, spatial and kinetic heterogeneity. Freshw. Biol. 51, 1264–1276 (2006).
Kim, C., Nishimura, Y. & Nagata, T. Role of dissolved organic matter in hypolimnetic mineralization of carbon and nitrogen in a large, monomictic lake. Limnol. Oceanogr. 51, 70–78 (2006).
Catalán, N., Marcé, R., Kothawala, D. N. & Tranvik, L. J. Organic carbon decomposition rates controlled by water retention time across inland waters. Nat. Geosci. 9, 501–504 (2016).
Hampton, S. E. et al. Warming-induced changes in benthic redox as a potential driver of increasing benthic algal blooms in high-elevation lakes. Limnol. Oceanogr. Lett. 9, 1–6 (2024).
Hiatt, D. L., Back, J. A. & King, R. S. Effects of stream velocity and phosphorus concentrations on alkaline phosphatase activity and carbon:phosphorus ratios in periphyton. Hydrobiologia 826, 173–182 (2019).
Hupfer, M. & Lewandowski, J. Oxygen controls the phosphorus release from lake sediments—a long-lasting paradigm in limnology. Int. Rev. Hydrobiol. 93, 415–432 (2008).
Union, E. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 establishing a framework for community action in the field of water policy. Off. J. Eur. Commun. L327, 1–72 (2000).
Bennion, H. et al. Defining ecological and chemical reference conditions and restoration targets for nine European lakes. J. Paleolimnol. 45, 415–431 (2011).
Catalan, J., Ninot, J. M. & Aniz, M. M. The high mountain conservation in a changing world. In High Mountain Conservation in a Changing World (eds Catalan, J. et al.) (Springer, New York, 2017).
Chesson, P. AEDT: A new concept for ecological dynamics in the ever-changing world. PLOS Biol. 15, e2002634 (2017).
Vega, J. C., De Hoyos, C., Aldasoro, J., De Miguel, J. & Fraile, H. Nuevos datos morfométricos para el Lago de Sanabria. Limnetica 24, 115–122 (2005).
Vega, J. C., De Hoyos, C. & Aldasoro, J. The Sanabria Lake: The largest natural freshwater lake in Spain. Limnetica 8, 49–57 (1992).
Guijarro, J. A. Homogenization of climatic series with Climatol (2021).
Jeffrey, S. W. & Humphrey, G. F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 167, 191–194 (1975).
Boehrer, B. & Schultze, M. Density stratification and stability. In Lake Ecosystem Ecology: A Global Perspective (ed. Likens, G. E.) (Elsevier, New York, 2010).
CNIG. Digital Terrain Model—DTM05 of Spain. (2015).
Tarboton, D. G., Dash, P. & Sazib, N. TauDEM 5.3: Guide to Using the TauDEM Command Line Functions (Utah State University, Logan, 2015).
QGIS-v2.8. (2014).
Evans BM, Corradini KJ. MapShed Version 1.5 Users Guide. Prepared for Penn State Institutes of Energy and the Environment (University Park, 2016).
Pokhrel, B. K. & Paudel, K. P. Assessing the efficiency of alternative best management practices to reduce nonpoint source pollution in a rural watershed located in Louisiana, USA. Water 11, 1714 (2019).
Soil Conservation Service. Urban Hydrology for Small Watersheds (Engineering Division, Soil Conservation Service, US Department of Agriculture, 1986).
Schneiderman, E. M. Modeling phosphorus with the generalized watershed loading functions (GWLF) model. In Modeling Phosphorus in the Environment (eds Radcliffe, D. E. & Cabrera, M. L.) (CRC Press, Boca Raton, 2006).
Moriasi, D. N. et al. Model evaluation guidelines for systematic quantification of accuracy in watershed simulations. Trans. ASABE 50, 885–900 (2007).
Garcia-Gomez, H. et al. Joining empirical and modelling approaches to estimate dry deposition of nitrogen in Mediterranean forests. Environ. Pollut. 243, 427–436 (2018).
Sanchez-Cabeza, J. A., Masque, P. & Ani-Ragolta, I. Pb-210 and Po-210 analysis in sediments and soils by microwave acid digestion. J. Radioanal. Nucl. Chem. 227, 19–22 (1998).
Acknowledgements
We acknowledge Pablo Seisdedos and Noemí Hernández (Confederación Hidrográfica del Duero) for promoting and managing the limnological monitoring of Lake Sanabria (Project 2015-2019), the Spanish State Meteorological Agency (AEMET) and ENDESA company for the meteorological information, and the Natural Park of Lake Sanabria (Regional Government of Castilla y León) for authorizing the work and offering its laboratory in the natural park to facilitate the sampling campaigns. Héctor Calvete-Sogo (CIEMAT) and Santiago Robles helped with atmospheric deposition sampling, Samuel Arias (CEDEX) with fieldwork campaigns, Ana Alonso and Guillermo Cobo (CEDEX) with chemical analyses, and María E. Pérez with phytoplankton counting and biovolume estimation. Dolors Planas and Caridad De Hoyos provided helpful comments on early drafts.
Funding
Project "Control limnológico bianual intensivo del lago de Sanabria," funded by the Confederación Hidrográfica del Duero (Ministry for the Ecological Transition and the Demographic Challenge), commissioned to the Centre for Hydrographic Studies (CEDEX), CIEMAT and other research institutions (Ref. 452-A-640.01.01/2014), to study the ecological functioning of Lake Sanabria and its basin (2015–2019).
Author information
Authors and Affiliations
Contributions
M.T. conceived and led the 2015–2017 intensive study; J.C.V. led and carried out the long-term Lake Sanabria monitoring; A.P.M. led the hydrological and external nutrient loading model; B.V.G. led the sediment core study; A.I.N. led and performed the phytoplankton study; A.D. led the chemical analyses; H.G.-G. and R.A. led and performed the atmospheric deposition study; M.P. and C.N. performed sediment, and pigment analyses; J.L.V., and N.R. led the characterization of sewage discharges; H.M.L. performed the diatom study; S. P.-R. performed the cyst chrysophyte study; all CEDEX authors carried our the limnological sampling during 2015–2017; J.C., J.P., and M. B. performed data analysis; all authors contributed to results discussion; J.C. wrote a first draft of the manuscript and all authors provided feedback and contributed to the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Catalan, J., Monteoliva, A.P., Vega, J.C. et al. Reduced precipitation can induce ecosystem regime shifts in lakes by increasing internal nutrient recycling. Sci Rep 14, 12408 (2024). https://doi.org/10.1038/s41598-024-62810-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-62810-9
- Springer Nature Limited