

Abstract
The accumulation of senescent cells is a major cause of age-related inflammation and predisposes to a variety of age-related diseases1. However, little is known about the molecular basis underlying this accumulation and its potential as a target to ameliorate the ageing process. Here we show that senescent cells heterogeneously express the immune checkpoint protein programmed death-ligand 1 (PD-L1) and that PD-L1+ senescent cells accumulate with age in vivo. PD-L1− cells are sensitive to T cell surveillance, whereas PD-L1+ cells are resistant, even in the presence of senescence-associated secretory phenotypes (SASP). Single-cell analysis of p16+ cells in vivo revealed that PD-L1 expression correlated with higher levels of SASP. Consistent with this, administration of programmed cell death protein 1 (PD-1) antibody to naturally ageing mice or a mouse model with normal livers or induced nonalcoholic steatohepatitis reduces the total number of p16+ cells in vivo as well as the PD-L1+ population in an activated CD8+ T cell-dependent manner, ameliorating various ageing-related phenotypes. These results suggest that the heterogeneous expression of PD-L1 has an important role in the accumulation of senescent cells and inflammation associated with ageing, and the elimination of PD-L1+ senescent cells by immune checkpoint blockade may be a promising strategy for anti-ageing therapy.




Similar content being viewed by others
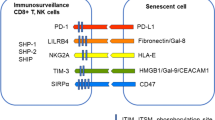
Data availability
The scRNA-seq data described in this article are available at Gene Expression Omnibus (GEO) under accessions GSE132042 and GSE155182. The bulk RNA-seq data are available under accessions GSE198396 and GSE198397. All other data needed to evaluate the conclusions in the paper are presented in the paper and/or the supplementary information.
Code availability
The code and scripts used for the RNA-seq analyses are available at Github (https://github.com/adslsars/PD-L1-senescence and https://github.com/Genomon-Project). Other analysis scripts are available from contacts under GEO accessions GSE198396 and GSE198397.
References
Childs, B. G. et al. Senescent cells: an emerging target for diseases of ageing. Nat. Rev. Drug Discov. 16, 718–735 (2017).
Kang, T. W. et al. Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature 479, 547–551 (2011).
Krizhanovsky, V. et al. Senescence of activated stellate cells limits liver fibrosis. Cell 134, 657–667 (2008).
Pardoll, D. M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 12, 252–264 (2012).
Johmura, Y. et al. Necessary and sufficient role for a mitosis skip in senescence induction. Mol. Cell 55, 73–84 (2014).
Pieren, D. K. J., Smits, N. A. M., van de Garde, M. D. B. & Guichelaar, T. Response kinetics reveal novel features of ageing in murine T cells. Sci. Rep. 9, 5587 (2019).
Lages, C. S., Lewkowich, I., Sproles, A., Wills-Karp, M. & Chougnet, C. Partial restoration of T-cell function in aged mice by in vitro blockade of the PD-1/PD-L1 pathway. Aging Cell 9, 785–798 (2010).
Channappanavar, R., Twardy, B. S., Krishna, P. & Suvas, S. Advancing age leads to predominance of inhibitory receptor expressing CD4 T cells. Mech. Ageing Dev. 130, 709–712 (2009).
Decman, V. et al. Defective CD8 T cell responses in aged mice are due to quantitative and qualitative changes in virus-specific precursors. J. Immunol. 188, 1933–1941 (2012).
Omori, S. et al. Generation of a p16 reporter mouse and its use to characterize and target p16high cells in vivo. Cell Metab. 32, 814–828.e6 (2020).
Sadasivam, S. & DeCaprio, J. A. The DREAM complex: master coordinator of cell cycle-dependent gene expression. Nat. Rev. Cancer 13, 585–595 (2013).
Helmbold, H., Kömm, N., Deppert, W. & Bohn, W. Rb2/p130 is the dominating pocket protein in the p53–p21 DNA damage response pathway leading to senescence. Oncogene 28, 3456–3467 (2009).
Fan, J. & Bertino, J. R. Functional roles of E2F in cell cycle regulation. Oncogene 14, 1191–1200 (1997).
Dyson, N., Guida, P., Münger, K. & Harlow, E. Homologous sequences in adenovirus E1A and human papillomavirus E7 proteins mediate interaction with the same set of cellular proteins. J. Virol. 66, 6893–6902 (1992).
Janky, R. et al. iRegulon: from a gene list to a gene regulatory network using large motif and track collections. PLoS Comput. Biol. 10, e1003731 (2014).
Verfaillie, A., Imrichova, H., Janky, R. & Aerts, S. iRegulon and i-cisTarget: reconstructing regulatory networks using motif and track enrichment. Curr. Protoc. Bioinformatics 52, 2 (2015).
Zhang, J. et al. Cyclin D–CDK4 kinase destabilizes PD-L1 via cullin 3-SPOP to control cancer immune surveillance. Nature 553, 91–95 (2018).
Lim, S. O. et al. Deubiquitination and stabilization of PD-L1 by CSN5. Cancer Cell 30, 925–939 (2016).
Rechsteiner, M. & Hill, C. P. Mobilizing the proteolytic machine: cell biological roles of proteasome activators and inhibitors. Trends Cell Biol. 15, 27–33 (2005).
Radhakrishnan, S. K. et al. Transcription factor Nrf1 mediates the proteasome recovery pathway after proteasome inhibition in mammalian cells. Mol. Cell 38, 17–28 (2010).
Zhang, Y. & Hayes, J. D. The membrane-topogenic vectorial behaviour of Nrf1 controls its post-translational modification and transactivation activity. Sci. Rep. 3, 2006 (2013).
Rodier, F. et al. Persistent DNA damage signalling triggers senescence-associated inflammatory cytokine secretion. Nat. Cell Biol. 11, 973–979 (2009).
Muñoz-Espín, D. & Serrano, M. Cellular senescence: from physiology to pathology. Nat. Rev. Mol. Cell Biol. 15, 482–496 (2014).
Griffin, G. K. et al. Epigenetic silencing by SETDB1 suppresses tumour intrinsic immunogenicity. Nature 595, 309–314 (2021).
Zhang, S. M. et al. KDM5B promotes immune evasion by recruiting SETDB1 to silence retroelements. Nature 598, 682–687 (2021).
Kalbasi, A. et al. Uncoupling interferon signaling and antigen presentation to overcome immunotherapy resistance due to JAK1 loss in melanoma. Sci. Transl. Med. 12, eabb0152 (2020).
Grosse, L. et al. Defined p16High senescent cell types are indispensable for mouse healthspan. Cell Metab. 32, 87–99.e6 (2020).
Maeso-Díaz, R. et al. Effects of aging on liver microcirculatory function and sinusoidal phenotype. Aging Cell 17, e12829 (2018).
Vaddepally, R. K., Kharel, P., Pandey, R., Garje, R. & Chandra, A. B. Review of indications of FDA-approved immune checkpoint inhibitors per NCCN guidelines with the level of evidence. Cancers 12, 738 (2020).
Liu, J. Y. et al. Cells exhibiting strong p16INK4a promoter activation in vivo display features of senescence. Proc. Natl Acad. Sci. USA 116, 2603–2611 (2019).
Schulte, H., Mühlfeld, C. & Brandenberger, C. Age-related structural and functional changes in the mouse lung. Front. Physiol. 10, 1466 (2019).
Gregg, S. Q. et al. A mouse model of accelerated liver aging caused by a defect in DNA repair. Hepatology 55, 609–621 (2012).
Sheth, K. A. et al. Muscle strength and size are associated with motor unit connectivity in aged mice. Neurobiol. Aging 67, 128–136 (2018).
Tung, V. W. K., Burton, T. J., Quail, S. L., Mathews, M. A. & Camp, A. J. Motor performance is impaired following vestibular stimulation in ageing mice. Front. Aging Neurosci. 8, 12 (2016).
Ogrodnik, M. et al. Cellular senescence drives age-dependent hepatic steatosis. Nat. Commun. 8, 15691 (2017).
Matsumoto, M. et al. An improved mouse model that rapidly develops fibrosis in non-alcoholic steatohepatitis. Int. J. Exp. Pathol. 94, 93–103 (2013).
Chang, J. et al. Clearance of senescent cells by ABT263 rejuvenates aged hematopoietic stem cells in mice. Nat. Med. 22, 78–83 (2016).
Kazankov, K. et al. The role of macrophages in nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Nat. Rev. Gastroenterol. Hepatol. 16, 145–159 (2019).
Gate, D. et al. Clonally expanded CD8 T cells patrol the cerebrospinal fluid in Alzheimer’s disease. Nature 577, 399–404 (2020).
Hashimoto, K. et al. Single-cell transcriptomics reveals expansion of cytotoxic CD4 T cells in supercentenarians. Proc. Natl Acad. Sci. USA 116, 24242–24251 (2019).
McDonnell, W. J. et al. High CD8 T-cell receptor clonality and altered CDR3 properties are associated with elevated isolevuglandins in adipose tissue during diet-induced obesity. Diabetes 67, 2361–2376 (2018).
Koda, Y. et al. CD8+ tissue-resident memory T cells promote liver fibrosis resolution by inducing apoptosis of hepatic stellate cells. Nat. Commun. 12, 4474 (2021).
Baruch, K. et al. PD-1 immune checkpoint blockade reduces pathology and improves memory in mouse models of Alzheimer’s disease. Nat. Med. 22, 135–137 (2016).
Rosenzweig, N. et al. PD-1/PD-L1 checkpoint blockade harnesses monocyte-derived macrophages to combat cognitive impairment in a tauopathy mouse model. Nat. Commun. 10, 465 (2019).
Pfister, D. et al. NASH limits anti-tumour surveillance in immunotherapy-treated HCC. Nature 592, 450–456 (2021).
Martins, F. et al. Adverse effects of immune-checkpoint inhibitors: epidemiology, management and surveillance. Nat. Rev. Clin. Oncol. 16, 563–580 (2019).
Kondo, T. et al. Notch-mediated conversion of activated T cells into stem cell memory-like T cells for adoptive immunotherapy. Nat. Commun. 8, 15338 (2017).
Hogquist, K. A. et al. T cell receptor antagonist peptides induce positive selection. Cell 76, 17–27 (1994).
Simma, O. et al. Identification of an indispensable role for tyrosine kinase 2 in CTL-mediated tumor surveillance. Cancer Res. 69, 203–211 (2009).
Merry, T. L. et al. The CSF1 receptor inhibitor pexidartinib (PLX3397) reduces tissue macrophage levels without affecting glucose homeostasis in mice. Int. J. Obesity 44, 245–253 (2020).
Nakanishi, M. et al. Exit from G0 and entry into the cell cycle of cells expressing p21Sdi1 antisense RNA. Proc. Natl Acad. Sci. USA 92, 4352–4356 (1995).
Parrinello, S. et al. Oxygen sensitivity severely limits the replicative lifespan of murine fibroblasts. Nat. Cell Biol. 5, 741–747 (2003).
Johmura, Y. et al. SCFFbxo22-KDM4A targets methylated p53 for degradation and regulates senescence. Nat. Commun. 7, 10574 (2016).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Anders, S., Pyl, P. T. & Huber, W. HTSeq-A Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Liao, Y., Smyth, G. K. & Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 47, e47 (2019).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2009).
Ritchie, M. E. et al. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43, e47 (2015).
Yu, G., Wang, L. G., Han, Y. & He, Q. Y. ClusterProfiler: an R package for comparing biological themes among gene clusters. OMICS 16, 284–287 (2012).
Storer, J., Hubley, R., Rosen, J., Wheeler, T. J. & Smit, A. F. The Dfam community resource of transposable element families, sequence models, and genome annotations. Mob. DNA 12, 2 (2021).
Wolf, F. A., Angerer, P. & Theis, F. J. SCANPY: Large-scale single-cell gene expression data analysis. Genome Biol. 19, 15 (2018).
Lewis, M. D. et al. A reproducible method for the expansion of mouse CD8+ T lymphocytes. J. Immunol. Methods 417, 134–138 (2015).
Kitamura, T., Doughty-Shenton, D., Pollard, J. W. & Carragher, N. O. Real time detection of in vitro tumor cell apoptosis induced by CD8+ T cells to study immune suppressive functions of tumor-infiltrating myeloid cells. J. Vis. Exp. https://doi.org/10.3791/58841 (2019).
Acknowledgements
We thank C. Konishi, Y. Chiba, T. Ando, T. Kanai and Y. Zhang for technical assistance. The super-computing resource was provided by the Human Genome Center (University of Tokyo). This study was supported by Imaging Core Laboratory and FACS Core Laboratory, Institute of Medical Science, University of Tokyo. This study was supported by AMED under Grant Numbers JP22zf0127003h (M.N. and A.Y.), JP20gm5010001s (M.N.), JP20ck010655h (M.N.), JP21gm6410014h (Y.J.), JP22gm1110009 (A.Y.) and MEXT/JSPS KAKENHI under grant numbers JP20H00514 (M.N.), JP20K21497 (M.N.), JP19H05740 (M.N.), JP19H03431 (Y.J.), JP20H04940 (Y.J.), JP21H05044 (A.Y.), JP22K1944 (A.Y.) and Princess Takamatsu Cancer Research Fund (M.N.).
Author information
Authors and Affiliations
Contributions
M.N. and Y.J. conceived the project. T.-W.W., Y.J. and M.N. planned the experiments. T.-W.W., Y.J., N.S. and S.O. performed the experiments. T.-W.W., Y.J., T.M., K.Y., S.H., S.Y., E.S., S.I., Y.F., A.Y. and M.N. analysed the results, and M.N. and T.-W.W. wrote the manuscript with editing by all the other authors.
Corresponding authors
Ethics declarations
Competing interests
M.N. is a scientific advisor and a shareholder of reverSASP Therapeutics. S.Y. is a co-founder of Celaid Therapeutics.
Peer review
Peer review information
Nature thanks Jesus Gil and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Heterogeneous PD-L1 expression levels in senescent cells.
a, The population of cells expressing the indicating immune checkpoint proteins in quiescent (Quie) or nutlin3a-induced senescent (n-Sen) HCA2 cells. Gating strategy was shown in the left lower panels. The representative scatterplots were shown in the lower right panels. b, Expression of SASP factors in Tom- and Tom+CD45- cells sorted from 6 months old male p16-Tom mice was analyzed by qPCR (n = 3 for each group). c, Gating strategy for the experiments shown in Fig. 1c. d, The ratio of PD-L1+ cells in Tom- cells determined by FACS in young (2-month-old, n=7) and old (24- to 26-month-old, n=4) p16-Tom mice. Data are presented as means ± SEM of independent experiments. Unpaired t-test (b and d) was performed. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001. The precise p-values were described in Methods.
Extended Data Fig. 2 Regulation of PD-L1 expression by p130-E2F axis.
a, Immunoblotting of lysates from quiescent (Quie), replicative senescent (r-Sen), nutlin3a senescent (n-Sen), d-Sen, and proliferative (Prolifer) HCA2 cells using the indicated antibodies. b, qPCR analysis of PD-L1 and CDC20 expression in sorted G1, S, and G2/M HCA2 cells. The results are shown as relative fold changes. c, Immunoblotting of lysates from HCA2 cells expressing mock, p130, and Flag-E2F97 using the indicated antibodies. d, qPCR analysis of PD-L1 and CDC20 expression in HCA2 cells as in c. e, Immunoblotting of lysates from proliferative and d-Sen HCA2 cells expressing doxycycline (Doxy)-inducible 3xFlag-E7 in the presence or absence of Doxy (1 μg/ml) using the indicated antibodies. The duration of senescence induction by doxorubicin were shown (Day 3 or Day 12). f, A volcano plot of differentially expressed genes (DEGs) between PD-L1+ and PD-L1− d-Sen HCA2 cells. Blue dots indicate the DEGs identified by FDR < 0.05. A red arrow indicates Cd274 (PD-L1). g, TF candidates enriched on promoter regions of PD-L1 positively correlated genes (R2 > 0.95) in RNA-seq results as in f. The threshold of normalized enrichment score was greater than 3. The transcriptional regulatory network was showed in the right panel. (Pink circle: targeted correlated gene; Blue circle: untargeted correlated gene; Green circle: TF candidate; Red box: E2F1 and CD274) h, Immunoblotting of lysates from sorted PD-L1− and PD-L1+ d-Sen HCA2 cells using the indicated antibodies. i, Gating strategy for the experiments shown in Fig. 1e. Relative quantified values of PD-L1 was labeled. Data are presented as means ± SEM of three independent experiments. All immunoblotting results were repeated independently for three times. One-way ANOVA with post-hoc Tukey HSD test (b) and Dunnett’s test (d). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. The precise p-values were described in Methods.
Extended Data Fig. 3 Heterogeneous expression of PD-L1 and inflammatory transcriptome in senescent cells.
a, SA-β-Gal staining of Quie, n-Sen, and d-Sen MPFs. The representative images were shown in the right panels. Scale bar: 500 μm b, Immunoblotting of lysates from MPFs as in a using the indicated antibodies. c, The representative histograms of the results shown in Fig. 2b (right panels). Gating strategy of in vitro T cell killing assay was shown in the left panels. d, The category-gene-net plot of DEGs between n-Sen and Quie MPFs involving in GO results as in Fig. 2d. e, GSEA of RNA-seq result between n-Sen and Quie MPFs. All GSEA terms were identified by p.adj < 0.05, which was adjusted by B-H method. f, Immunoblotting of lysates from the doxy-inducible 3xFlag-IκBα(S32/36A) expressing n-Sen MPFs using the indicated antibodies with or without 1 μg/ml doxy treatments for 2 days. g, qPCR analysis of SASP factors expression in n-Sen MPFs with or without doxy-inducible IκBα(S32/36A). h, The representative histogram of the results shown in Fig. 2e. Data are presented as means ± SEM of three independent experiments. All immunoblotting results were repeated independently for three times. One-way ANOVA with Tukey HSD test was performed. ****P < 0.0001. The precise p-values were described in Methods.
Extended Data Fig. 4 Heterogeneous expression of PD-L1 in senescent cells reduces cytotoxicity of T cells.
a, qPCR analysis of ERVs expression in Tom- and Tom+ CD45- cells sorted from 6 months old male p16-Tom mice. b, The representative histogram of the results shown in Fig. 2g. c, Cytotoxicity of the sorted PD-L1+ and PD-L1− n-Sen MPFs (Target cells) was evaluated by incubating with mouse activated CD8+ T cells (Effector cells) from OT-1 mice for 18 hrs. During the co-culture, ovalbumin (OVA) was added with 1 μg/ml to trigger the antigen specific response. The representative histograms were shown in the right panels. d, Immunoblotting of lysates from the mock, PD-L1 overexpression (OE) MPFs using the indicated antibodies. e, The representative histogram of the results shown in Fig. 2h. f, Cytotoxicity of the n-Sen MPFs expressing mock or PD-L1 with 10 μg/ml isotype IgG or αPD-1 treatments during the incubation with CD8+ T cells from OT-1 mice. The representative histograms were shown in the right panels. Data are presented as means ± SEM of three independent experiments. All immunoblotting results were repeated independently for three times. Unpaired t-test (a) and Two-way ANOVA with Sidak’s test were performed (c and f). **P < 0.01, ***P < 0.001, ****P < 0.0001. The precise p-values were described in Methods.
Extended Data Fig. 5 The inflammation-related phenotypes in PD-L1+ p16high cells in vivo.
a, UMAP visualization of tdTomato positive (Tom+) population (upper panels) and Cd274 (PD-L1) expression levels (lower panels) in the indicating scRNA-seq datasets and cell types. The NASH samples were prepared from a 7-month-old p16-Tom mouse with feeding CDA-HFD for 6 months. The kidney sample was from a 5.5-month-old p16-Tom mouse. b, Violin plots for the expression levels of Cd274 (PD-L1) in Tom+ and Tom- cells of the indicating cell types. The red dotted lines represent the cut-off threshold for determining PD-L1+ and PD-L1− cells. c-e, Volcano plots of DEGs between PD-L1+ and PD-L1− Tom+ cells in the indicated samples. The DEGs were identified by an FDR < 0.05 and a Log2FC > 0.15. Red dots represent significant DEGs. f-h, The enriched GO terms in upregulated DEGs of Tom+PD-L1+ cells compared with Tom+PD-L1− cells in the indicating samples.
Extended Data Fig. 6 Immune checkpoint blockade treatment promotes the CD8+ T cell-dependent immune clearance of senescent cells in vivo.
a, FACS analysis of splenic CD8+ T cells of αPD-1 and/or αCD8 treated mice as in Fig. 4a. Gating strategy and representative scatterplots were shown in the right panels. (IgG+IgG: n = 6, αPD-1+IgG: n = 6, IgG+αCD8: n = 7, αPD-1+αCD8: n = 5) b, The representative images of immunofluorescent staining shown in Fig. 4a with the indicating treatments in lung, liver, and kidney. (Blue: DAPI; Red: PD-L1; Yellow: tdTomato; Green: CD3) Scale bar: 500 μm c, The proportion of PD-L1+Tom+ cells (upper panels) and PD-L1−Tom+ cells (lower panels) from immunofluorescent staining sections of lung, liver, kidney with the indicating treatments. Fold changes (FC) between IgG and αPD-1 groups without αCD8 treatment were shown on figures. (IgG+IgG: n = 7, αPD-1+IgG: n = 7, IgG+αCD8: n = 6, αPD-1+αCD8: n = 5) Data are presented as means ± SEM of three or more independent experiments. Two-way ANOVA with Sidak’s test was performed (a and c). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. The precise p-values were described in Methods.
Extended Data Fig. 7 Immune checkpoint blockade treatment alters senescent population and ageing phenotypes in vivo.
a, The representative images of immunofluorescent staining in liver with IgG and αPD-1 treatments as shown in Fig. 4a. (Blue: DAPI; Red: PD-L1; Yellow: tdTomato; Green: F4/80) Scale bar: 500 μm b, The quantified proportion of F4/80- in Tom+ cells from immunofluorescent staining sections of liver from mice with the indicating treatments. (IgG: n = 5, αPD-1: n = 6) c, The proportion of Tom+ cells in F4/80- (left panel) and F4/80+ cells (right panel) from immunofluorescent staining sections as in b. (IgG: n = 5, αPD-1: n = 6) d, The proportion of PD-L1+ cells in Tom+/F4/80- from immunofluorescent staining sections as in b. (IgG: n = 5, αPD-1: n = 6) e, The motor performance test of young (3-month-old) and old (17.5-month-old) wild type mice. The staying time was recorded from starting rotation of rotarod model to the mouse falling on the switch. (Young IgG: n = 6, Young αPD-1: n = 6, Old IgG: n = 7, Old αPD-1: n = 8) f, The grip strength test of mice as in e. (Young IgG: n = 6, Young αPD-1: n = 6, Old IgG: n = 7, Old αPD-1: n = 8) Data are presented as means ±SEM of three or more independent experiments. Unpaired two-sided t-test was performed (b, c, and d). Two-way ANOVA with Sidak’s test was performed (e and f). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. The precise p-values were described in Methods.
Extended Data Fig. 8 αPD-1 or ABT263 treatments improve NASH pathologies.
a, The activation and infiltration of hepatic CD8+ T cells analyzed by FACS from wild type mice fed with normal diet (Normal) or L-amino acid diet with 60 kcal % fat, 0.1 % methionine, and no added choline (CDA-HFD) for 10 weeks combined with αPD-1 or ABT263 treatments as in Fig. 4e. Gating strategy was shown in the right panel. (Normal placebo: n = 6, Normal αPD-1: n = 6, Normal ABT263: n = 5, CDA-HFD placebo: n = 6, CDA-HFD αPD-1: n = 6, CDA-HFD ABT263: n = 6) b, The serum levels of AST, ALT, and LDH from the mice as in a. (Normal placebo: n = 6, Normal αPD-1: n = 6, Normal ABT263: n = 5, CDA-HFD placebo: n = 6, CDA-HFD αPD-1: n = 6, CDA-HFD ABT263: n = 6) c, The Sirius red staining of livers from the mice as in a. The representative images were shown in upper panels. The percentage of red area (collagen area) in each filed was quantified and described as the hepatic fibrogenesis. Scale bar: 100 μm (n = 6 for each group) d, Body weights and serum total cholesterol (T-CHO) from the mice as in a. (n = 6 for each group) Data are presented as means ± SEM of three or more independent experiments. Two-way ANOVA with Sidak’s test was performed. **P < 0.01, ****P < 0.0001. The precise p-values were described in Methods.
Extended Data Fig. 9 Treatments of αPD-1 and CSF1R inhibitor improve NASH pathologies.
a, Experimental schedule of the PLX3397 treatment on young (3-month-old, n = 5) and old (17.5-month-old, n = 5) wild type mice. The treatment of 40 mg/kgBW PLX3397 was administrated by oral-gavage twice per week in a total 3 weeks period. (All the male mice in each group were adapted in two independent cages.) b, The motor performance test of the indicated mice as in a. The staying time was recorded from starting rotation of rotarod model to the mouse falling on the switch. c, The grip strength test of mice as in a. d, The quantification of alveolar size of the indicated mice as in a. Representative images (H&E staining) were shown in the right panels. e, Experimental schedule of the αPD1 and PLX3397 treatment on wild type mice fed with normal diet or CDA-HFD. (n = 5 for each group) f, The serum levels of AST, ALT, and LDH from the mice as in e. g, Representative images (lower panels: H&E staining) and quantification of lipid drop area (upper panel) of livers from the indicated mice as in e. h, The Sirius red staining of livers from the mice as in e. The representative images were shown in lower panels. Scale bars, 100 μm. Data are presented as means ±SEM of independent experiments. Two-way ANOVA with Sidak’s test was performed. ****P < 0.0001. The precise p-values were described in Methods.
Supplementary information
Supplementary Figure 1
Uncropped gel source data for Figure 1 and Extended Data Figures 2-4.
Supplementary Data
This zipped file contains source data for the main Fgures and Extended Data Figures.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Wang, TW., Johmura, Y., Suzuki, N. et al. Blocking PD-L1–PD-1 improves senescence surveillance and ageing phenotypes. Nature 611, 358–364 (2022). https://doi.org/10.1038/s41586-022-05388-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-022-05388-4
- Springer Nature Limited
This article is cited by
-
Regulatory mechanisms of PD-1/PD-L1 in cancers
Molecular Cancer (2024)
-
CDK4/6 inhibition enhances T-cell immunotherapy on hepatocellular carcinoma cells by rejuvenating immunogenicity
Cancer Cell International (2024)
-
Microfluidics-enabled fluorinated assembly of EGCG-ligands-siTOX nanoparticles for synergetic tumor cells and exhausted t cells regulation in cancer immunotherapy
Journal of Nanobiotechnology (2024)
-
Role of cellular senescence in inflammation and regeneration
Inflammation and Regeneration (2024)
-
The interaction between ageing and Alzheimer's disease: insights from the hallmarks of ageing
Translational Neurodegeneration (2024)