
Abstract
Receptor for activated C kinase 1 (RACK1) has been confirmed to take part in multiple biological events and the mechanism supporting abnormal RACK1 expression in ovarian cancer (OC) remains to be characterized. Here, we identified Smad ubiquitin regulatory factor 2 (SMURF2) as a bona fide E3 ligase of RACK1 in OC. SMURF2 effectively added the K6, K33 and K48 ubiquitin chains to the RACK1, resulting in polyubiquitination and instability of RACK1. PCAF promoted acetylation of RACK1 at K130, leading to SMURF2-mediated RACK1 ubiquitination inhibited and promote OC progression. The expression levels of SMURF2 and RACK1 were negatively correlated. SMURF2 was abnormal low expression in human ovarian cancer, resulting in decreased ubiquitination of RACK1 and increased stability, which promoted OC progression, and strongly associated with poor patients’ prognosis. In general, our results demonstrated that SMURF2 plays a pivotal role in stabilizing RACK1, which in turn facilitates tumorigenesis in OC, suggesting that SMURF2-RACK1 axis may prove to be potential targets for the treatment of OC.
Similar content being viewed by others

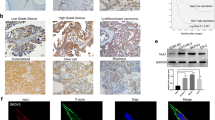

Introduction
Ovarian cancer (OC) is one of the most common cancers in the female genital tract, which is the third most common cancer behind cervical cancer and endometrial cancer, and the most deadly, being the 8th most common cause of cancer deaths among women [1]. The overall survival of OC patients has not improved significantly over the past decades due to the lack of typical and easily detectable early clinical signs, the lack of durable and effective treatment, and the elevated risk of chemoresistance and recurrence, with most cases being clinically advanced and with distant metastases at the time of diagnosis [2]. Therefore, in order to provide a theoretical basis for improving clinical outcomes in OC, there is an urgent need to better understand the potential pathogenesis of OC and to significantly improve our understanding of tumor progression and metastasis.
Post-translational modifications (PTMs) are a dynamic and reversible post-translational epigenetic modification of proteins with low metabolic cost and sensitive response [3]. It is an important mechanism for increasing the variety and functional diversity of proteins and plays a key role in regulating metabolism, signal transduction, reproductive development, tumor inflammation and other physiological and pathological conditions. Ubiquitination is an important type of PTMs, plays a crucial role in regulating the “quantity” and “quality” of substrate proteins, thus hel** to ensure homeostasis of the intracellular environment and the smooth running of life processes. Malfunctions of the ubiquitin-proteasome system are responsible for more than 80% of proteins are degraded in cells and have been demonstrated to cause pathologies, particularly malignant tumors [4]. Ubiquitination is a cascading process in which ubiquitin, a universally expressed 76 amino acid protein, first binds to ubiquitin-activating enzymes (E1s) and activates ubiquitin, then transfers the activated ubiquitin molecule to ubiquitin-binding enzymes (E2s). Ubiquitin ligases (E3s) then transfer ubiquitin molecules from the E2s to the underlying protein, ultimately leading to its degradation by the proteasome. Due to their relative specificity in recognizing substrate proteins, E3 ligases play a key role in the overall ubiquitination process.
Smad ubiquitin regulatory factor 2 (SMURF2) is a HECT-type E3 ubiquitin (Ub) ligase that regulates many key functional proteins, including SATB1 [5], RNF20 [6], YY1 [7], and Smad2 [8], which are involved in oncogenic or tumor suppressor functions. Recent studies have shown that SMURF2 interacts with SIRT1 to mediate the degradation of SIRT1, while deletion of SMURF2 expression leads to upregulation of SIRT1, inducing tumor initiation and the invasive metastasis of colorectal cancer in vivo and in vitro [9]. In addition, SMURF2 induces enhanced tumor metastasis in nude mice models of breast cancer [10]. Nevertheless, the role of SMURF2 in the progression of OC has not yet been the subject of in-depth investigation.
The receptor for activated C-kinase 1 (RACK1) is involved in a wide range of cellular signaling pathways, as a member of the tryptophan-aspartate repeat (WD repeat) family of proteins. Identified as a classic scaffolding protein for a variety of kinases and receptors, RACK1 is involved in a number of intracellular signaling pathways and plays a key role in a variety of physiological processes including cell growth, migration and differentiation [11,12,13,Protein expression and purification For the production of proteins from bacteria, Escherichia coli BL21 (DE3) cells containing the GST, GST-SMURF2 and GST-SMURF2 C716A plasmids were induced for protein expression using 0.5 mM IPTG at 37 °C for 4–6 h. The cells were lysed in buffered saline. Cell lysis was performed with lysis buffer (0.5% Triton X-100, pH 7.5, 1 mM DTT, 50 mM Tris-Cl, 200 mM NaCl, 10% glycerol and 1 mM PMSF and sonicated). Lysates were centrifuged and incubated with glutathione-Sepharose 4B (GE Healthcare) at 4 °C for 4 h or overnight. The resin was washed three times with lysis buffer plus 300 mM NaCl and then washed two more times with PBS. Immobilization on glutathione-Sepharose beads was verified by SDS-PAGE and aliquoted for storage at −80 °C. 6×His-RACK1 was purified using nickel-nitrilotriacetic acid (Ni-NTA) matrices (QIAGEN). Cells were transfected with the indicated plasmids, then treated with 20 μM MG132 for 8 h for the in vivo RACK1 ubiquitylation assay. The cells were harvested and lysed in RIPA lysis buffer plus 1% SDS, 20 µM MG132, and protease inhibitors. The lysates were incubated with anti-RACK1 or anti-Myc antibodies for 12 h and with Protein A/G magnetic beads for an additional 12 h at 4 °C. The precipitated protein was boiled in SDS-PAGE loading buffer for 10 min and then treated with IB. Bacterially expressed GST, GST-SMURF2, or GST-SMURF2 C716A was bound to glutathione-Sepharose 4B beads (GE Healthcare). The complexes were mixed with Myc-RACK1 expressed in HEK293T cells for 2 h at 4 °C. After incubation, the complexes were washed with GST binding buffer for at least 3 times, then eluted with SDS-PAGE loaded buffer by boiling and treated with IB antibody shown. GraphPad Prism (version 8.0) was used for all statistical analyses. All in vitro experiments were carried out in at least three replicates and the data presented are from one representative experiment. The data are expressed as mean ± standard deviation. Double-tailed Student’s t-test or two-factor analysis of variance was used to assess the statistical significance of differences. The correlation between SMURF2 and RACK1 expression in OC patients was calculated by Pearson correlation analysis. Overall survival was assessed by the Kaplan-Meier method and compared by the log-rank test. P < 0.05 was considered statistically significant.In vivo ubiquitination assay
GST pull-down assay
Statistics and reproducibility
Data availability
Mass spectrometry data was uploaded in the Supplementary Table 1. Other datasets are available from the corresponding author on reasonable request.
References
Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin. 2023;73:17–48.
Schoutrop E, Moyano-Galceran L, Lheureux S, Mattsson J, Lehti K, Dahlstrand H, et al. Molecular, cellular and systemic aspects of epithelial ovarian cancer and its tumor microenvironment. Semin Cancer Biol. 2022;86:207–23.
Chen L, Liu S, Tao Y. Regulating tumor suppressor genes: post-translational modifications. Signal Transduct Target Ther. 2020;5:90.
Chen YJ, Wu H, Shen XZ. The ubiquitin-proteasome system and its potential application in hepatocellular carcinoma therapy. Cancer Lett. 2016;379:245–52.
Yu L, Dong L, Wang Y, Liu L, Long H, Li H, et al. Reversible regulation of SATB1 ubiquitination by USP47 and SMURF2 mediates colon cancer cell proliferation and tumor progression. Cancer Lett. 2019;448:40–51.
Blank M, Tang Y, Yamashita M, Burkett SS, Cheng SY, Zhang YE. A tumor suppressor function of Smurf2 associated with controlling chromatin landscape and genome stability through RNF20. Nat Med. 2012;18:227–34.
Fu SH, Lai MC, Zheng YY, Sun YW, Qiu JJ, Gui F, et al. MiR-195 inhibits the ubiquitination and degradation of YY1 by Smurf2, and induces EMT and cell permeability of retinal pigment epithelial cells. Cell Death Dis. 2021;12:708.
Lin B, Huang D, Lin G, Miao Y, Wang J, Fan Z, et al. Smurf2-induced degradation of SMAD2 causes inhibition of hair follicle stem cell differentiation. Cell Death Discov. 2022;8:160.
Yu L, Dong L, Li H, Liu Z, Luo Z, Duan G, et al. Ubiquitination-mediated degradation of SIRT1 by SMURF2 suppresses CRC cell proliferation and tumorigenesis. Oncogene. 2020;39:4450–64.
Tsao SM, Hsu HY. Fucose-containing fraction of Ling-Zhi enhances lipid rafts-dependent ubiquitination of TGFbeta receptor degradation and attenuates breast cancer tumorigenesis. Sci Rep. 2016;6:36563.
Cao J, Zhao M, Liu J, Zhang X, Pei Y, Wang J, et al. RACK1 promotes self-renewal and chemoresistance of cancer stem cells in human hepatocellular carcinoma through stabilizing Nanog. Theranostics. 2019;9:811–28.
Duan F, Wu H, Jia D, Wu W, Ren S, Wang L, et al. O-GlcNAcylation of RACK1 promotes hepatocellular carcinogenesis. J Hepatol. 2018;68:1191–202.
Ling J, Tiwari M, Chen Y, Sen GL. RACK1 prevents the premature differentiation of epidermal progenitor cells by inhibiting IRF6 expression. J Invest Dermatol. 2022;142:1499–502.e4.
Wu B, Chang N, ** H, **ong J, Zhou Y, Wu Y, et al. PHB2 promotes tumorigenesis via RACK1 in non-small cell lung cancer. Theranostics. 2021;11:3150–66.
Yang H, Zhu Q, Cheng J, Wu Y, Fan M, Zhang J, et al. Opposite regulation of Wnt/beta-catenin and Shh signaling pathways by Rack1 controls mammalian cerebellar development. Proc Natl Acad Sci USA. 2019;116:4661–70.
Fan Y, Si W, Ji W, Wang Z, Gao Z, Tian R, et al. Rack1 mediates Src binding to drug transporter P-glycoprotein and modulates its activity through regulating Caveolin-1 phosphorylation in breast cancer cells. Cell Death Dis. 2019;10:394.
Sutton P, Borgia JA, Bonomi P, Plate JM. Lyn, a Src family kinase, regulates activation of epidermal growth factor receptors in lung adenocarcinoma cells. Mol Cancer. 2013;12:76.
Zhong X, Li M, Nie B, Wu F, Zhang L, Wang E, et al. Overexpressions of RACK1 and CD147 associated with poor prognosis in stage T1 pulmonary adenocarcinoma. Ann Surg Oncol. 2013;20:1044–52.
Caron C, Boyault C, Khochbin S. Regulatory cross-talk between lysine acetylation and ubiquitination: role in the control of protein stability. Bioessays. 2005;27:408–15.
Narita T, Weinert BT, Choudhary C. Functions and mechanisms of non-histone protein acetylation. Nat Rev Mol Cell Biol. 2019;20:156–74.
Shimizu K, Gi M, Suzuki S, North BJ, Watahiki A, Fukumoto S, et al. Interplay between protein acetylation and ubiquitination controls MCL1 protein stability. Cell Rep. 2021;37:109988.
Wu Y, Wang X, Xu F, Zhang L, Wang T, Fu X, et al. The regulation of acetylation and stability of HMGA2 via the HBXIP-activated Akt-PCAF pathway in promotion of esophageal squamous cell carcinoma growth. Nucleic Acids Res. 2020;48:4858–76.
Zhang N, Zhang Y, Miao W, Shi C, Chen Z, Wu B, et al. An unexpected role for BAG3 in regulating PARP1 ubiquitination in oxidative stress-related endothelial damage. Redox Biol. 2022;50:102238.
Wang C, Wan X, Yu T, Huang Z, Shen C, Qi Q, et al. Acetylation stabilizes phosphoglycerate dehydrogenase by disrupting the interaction of E3 ligase RNF5 to promote breast tumorigenesis. Cell Rep. 2020;32:108021.
Lin Y, Cui M, Teng H, Wang F, Yu W, Xu T. Silencing the receptor of activated C-kinase 1 (RACK1) suppresses tumorigenicity in epithelial ovarian cancer in vitro and in vivo. Int J Oncol. 2014;44:1252–8.
Li Y, Sun X, Gao D, Ding Y, Liu J, Chen J, et al. Dual functions of Rack1 in regulating Hedgehog pathway. Cell Death Differ. 2020;27:3082–96.
Wang J, Chen X, Hu H, Yao M, Song Y, Yang A, et al. PCAT-1 facilitates breast cancer progression via binding to RACK1 and enhancing oxygen-independent stability of HIF-1alpha. Mol Ther Nucleic Acids. 2021;24:310–24.
Zhang C, Jiang H, Yuan L, Liao Y, Liu P, Du Q, et al. CircVPRBP inhibits nodal metastasis of cervical cancer by impeding RACK1 O-GlcNAcylation and stability. Oncogene. 2023;42:793–807.
Wu H, Song S, Yan A, Guo X, Chang L, Xu L, et al. RACK1 promotes the invasive activities and lymph node metastasis of cervical cancer via galectin-1. Cancer Lett. 2020;469:287–300.
Morreale FE, Walden H. Types of ubiquitin ligases. Cell. 2016;165:248–248.e1.
Adhikari A, Chen ZJ. Diversity of polyubiquitin chains. Dev Cell. 2009;16:485–6.
Wang Q, Xu C, Cai R, An W, Yuan H, Xu M. Fbxo45-mediated NP-STEP(46) degradation via K6-linked ubiquitination sustains ERK activity in lung cancer. Mol Oncol. 2022;16:3017–33.
Acknowledgements
We thank Zhaocong Zhang kindness of providing the help on data analysis.
Funding
This work was supported by the National Natural Science Foundation of China (No. 82173238), Key Program of Natural Science Foundation of Heilongjiang Province (ZD2020H007).
Author information
Authors and Affiliations
Contributions
GL designed, supervised the study, and revision and final approval of the manuscript. YNP contributed to data research, analysis, and manuscript writing. QSF, FSS, ZQW, YZ, DJC, and YML contributed to the data collection and analysis. All the authors have read the manuscript and provided useful comments.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
The research was carried out according to the World Medical Association Declaration of Helsinki and was approved by the Ethics Committee at Harbin Medical University Cancer Hospital.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Pi, Y., Feng, Q., Sun, F. et al. Loss of SMURF2 expression enhances RACK1 stability and promotes ovarian cancer progression. Cell Death Differ 30, 2382–2392 (2023). https://doi.org/10.1038/s41418-023-01226-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41418-023-01226-w
- Springer Nature Limited