
Abstract
Cataract refers to opacities of the lens that impede the passage of light. Mutations in heat shock transcription factor 4 (HSF4) have been associated with cataract; however, the mechanisms regarding how mutations in HSF4 cause cataract are still obscure. In this study, we generated an hsf4 knockout zebrafish model using TALEN technology. The mutant zebrafish developed an early-onset cataract with multiple developmental defects in lens. The epithelial cells of the lens were overproliferated, resulting in the overabundance of lens fiber cells in hsf4null zebrafish lens. Consequently, the arrangement of the lens fiber cells became more disordered and irregular with age. More importantly, the terminal differentiation of the lens fiber cell was interrupted as the organelles cannot be cleaved in due time. In the cultured human lens epithelial cells, HSF4 could stabilize and retain p53 in the nucleus to activate its target genes such as fas cell surface death receptor (Fas) and Bcl-2-associated X apoptosis regulator (Bax). In the hsf4null fish, both p53 and activated-caspase3 were significantly decreased. Combined with the finding that the denucleation defect could be partially rescued through microinjection of p53, fas and bax mRNA into the mutant embryos, we directly proved that HSF4 promotes lens fiber cell differentiation by activating p53 and its downstream regulators. The data we presented suggest that apoptosis-related genes are involved in the lens fiber cell differentiation. Our finding that HSF4 functions in the upstream to activate these genes highlighted the new regulatory modes of HSF4 in the terminal differentiation of lens fiber cell.
Similar content being viewed by others

Main
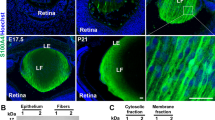
Cataract is a major cause of adult blindness and congenital cataract is a major cause of childhood blindness.1 At least 44 genetic loci and over 40 genes have been linked to congenital or early-onset cataract.2, 3, 4 Heat shock transcription factor 4 (HSF4) has been associated with isolated cataract5 and it belongs to the heat shock transcription factor family (including HSF1, HSF2, HSF3 and HSF4). These HSFs can respond to various stress stimuli and protect cells against proteotoxic damage.6, 7 More importantly, HSFs are also involved in regulating differentiation and development.8 HSF4 differs from other HSFs in that it lacks C-terminal HR-C domain, which is responsible for the negative regulation of trimerization.7, 9 Moreover, whereas HSF1 and HSF2 are expressed in most tissues, HSF4 is predominantly expressed in the lens.10, 11
To investigate the function of HSF4, three Hsf4 knockout mouse models have been constructed.12, 13, 14 All these three models developed early postpartum cataract. Histological analysis revealed that the nucleus and some cellular organelles of the secondary fiber cell persisted and the number of the epithelial cell increased. Associated with the altered biological processes, expression levels of the Hsf4 target genes were significantly changed. These genes include the ones encoding heat shock proteins (Hsp70, Hsp60 and Hsp27), diverse types of γ-crystallins, various forms of fibroblast growth factors or the cognate receptor (FGF1, 2, 4, 7 and FGFR1), the beaded filament proteins (Bfsp1/2)12, 13, 14 and several other target genes (SKAP2, Vimentin).15, 16 Besides the knockout models, the HSF4 p.Arg116His mutation and the Hsf4(exon1)-DBD-EGFP hybrid gene transgenic mouse models have also been constructed. Both models developed postnatal lamellar cataract, which is similar to the phenotype caused by the mutations in the DBD of human HSF4 gene.17, 18 These transgenic mice provide excellent models to study the lamellar cataract. Together, these studies revealed important functional aspects of the HSF4 gene. However, the exact mechanisms by which Hsf4 regulates lens development and whose mutations cause cataract still remain largely unknown.
Lens differentiation is a process through which an epithelial cell containing a full spectrum of cellular organelles is converted into a fiber cell characterized by the accumulation of high concentrations of lens-specific proteins and the loss of essential organelles.19 Recent studies demonstrated that lens differentiation is regulated by the same set of regulators responsible for the control of apoptosis.19, 20 These regulators include the tumor suppressor p53,21, 22, 23, 24, 25, 26, 27, 28 Bcl-2 family members,25, 29, 30, 31, 46 Tumor suppressor p53 has been implicated in regulating lens development. During mouse lens development, the expression of p53/Mdm2 was spatiotemporally regulated.24 Loss of p53 activity through expression of viral genes or the endogenous gene knockout induces posterior subcapsular cataracts.21, 47, 48 Besides, overexpressing human p53 in mouse lens led to microphthalmia.22 At the molecular level, p53 has been shown to regulate both major lens transcription factors c-Maf, Prox-126 and differentiation-related crystalline genes.49, 50 In addition, p53 regulates numerous apoptotic genes, some of which are implicated in regulating lens differentiation. For example, Fas and Bax mediate both extrinsic and intrinsic death pathways, which are merged to activate the downstream executional caspase3. It has been discovered that caspase3 is a key regulator of lens development.29, 30, 31, Full size image
hsf4 knockout in zebrafish causes early-onset cataract
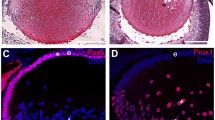
A slit lamp examination of the hsf4null zebrafish eye revealed clear presence of cataract formation at different stages including 2M, 3M and 12M (Figure 2a). By 12M, the hsf4null zebrafish lens became completely opaque (Figure 2a). In contrast, the WT lenses were transparent at all ages examined (Figure 2a). Defects in degenerating organelles are major causes of cataract. Thus, by observing denucleation status we can detect the onset of this defect. Using semiquantitative reverse transcription and polymerase chain reaction, we first determined that hsf4 was highly expressed in eyes at 48 hpf (Supplementary Figure 1). Shortly after that, denucleation began at 50 hpf, indicating that hsf4 was critically important in this process. Consistent with the expression pattern, no significant difference was observed between WT and hsf4null zebrafish before 48 hpf. After 2 dpf, great differences have been discovered. The WT zebrafish completed denucleation of the primary fiber cells before 3 dpf, and the secondary fiber cells successfully disintegrated their nuclei. In contrast, in hsf4null zebrafish lens, almost all the primary fiber cells in the lens core contained nuclei at 3 dpf. By 5 dpf, there were some nuclei still remaining in the lens core. The denucleation of the primary fiber cells eventually completed at 7 dpf, which indicated that the denucleation of the primary fiber cells was delayed but not abrogated when hsf4 was deleted (Figure 2b).

Knockout of hsf4 in zebrafish caused early-onset cataract. (a) Slit lamp examination of the WT and hsf4null zebrafish. Opacity of the eye lens could be detected from hsf4null zebrafish at all the ages we examined. However, the WT lens were transparent. (b) DAPI staining of the WT and hsf4null zebrafish lens at 2, 3, 5 and 7d. Scale bar: 20 μm. (c) Enlarged image of white box in the lens of 5-d-old hsf4null lens in b. (d) Enlarged image of white box in the lens of 7-d-old hsf4null lens in b. The white arrows in c and d indicate the spherical nucleus. Scale bar: 20 μm. (e) DAPI staining of the adult WT and hsf4null zebrafish lens at 2 and 6M. The middle column is the enlarged images of white boxes in 2M WT and hsf4null lens. The white arrows indicate the undegraded nucleus deposited in the hsf4null lens. The red arrows indicate the spherical nucleus existing in hsf4null lens. Scale bar: 500 μm
The denucleation of the secondary fiber cells seemed more severely affected within hsf4null zebrafish. By the ages of 2 and 6M, we discovered that most of the fiber cells in hsf4null lens contained intact nuclei, which were deposited together and even reached the most inner part of the lens. In the WT lens, however, the denucleation of the fiber cells was normal for we could only detect nuclei in the superficial differentiating fiber cells (Figure 2e). To our surprise, we found some spherical nuclei in the secondary fiber cells (Figures 2c–e). When the lens fiber cell differentiation initiates, nuclei start to elongate and become ovoid. Studies in bovine and chicken revealed that the volume and shape of the nuclei changed during disintegration.55, 56 Normally, the breakdown of the nucleus is a very rapid process; thus, the degenerating spherical nuclei are seldom found. The spherical nuclei we detected in the hsf4null lens maybe the fiber cells that were about to degrade. As the hsf4null lens contained an overwhelming majority of secondary fiber cells with ovoid nuclei, we reasoned that only a small proportion of the secondary fiber cells could disintegrate their nuclei in an hsf4-independent manner. Without hsf4, the denucleation of the secondary fiber cells appeared to come to a standstill.
hsf4 loss interrupts terminal differentiation of Zebrafish lens fiber cells
Previous research discovered that nuclei were preserved in the differentiating fiber cells as confirmed in hsf4null zebrafish; it is not clear whether the cellular non-nucleus organelles are also preserved in hsf4 knockouts. Thus, we performed transmission electron microscopy of 5-M-old hsf4null and WT zebrafish. As shown in Figure 3Af, the WT lens fiber cells were transparent and organelle-free, i.e., no dark stained structures interfering with the focusing of inside structure. In contrast, the fiber cells of the hsf4null zebrafish contained intact organelles, mitochondria, endosome and lysosome besides nuclei, suggesting that organelle degradation was also interrupted without hsf4 (Figure 3Aa–e). In the inner part of the lens, we could also detect intact nucleus (Figure 3Ac) and also observed that the connection between cortical lens fiber cells was loose (Figure 3Aa and b. In addition, the mutant fiber cells contained clustered vesicular structures (Figure 3Ad and e). These structures were early endosome, late endosome (LE) and LE–lysosome fusion body as observed from enlarged figures (Figure 3c). Usually, LE will be degraded when it is translocated to and fused with lysosome at the perinuclear region. In the hsf4null zebrafish lens, however, the deposited endosomes were mislocalized and left undegraded. Together, our study clearly revealed that the non-nucleus organelle degradation was also interrupted in the hsf4null lens fiber cells.

Interrupted organelle degradation in the lens fiber cells of hsf4null zebrafish. (A) Transmission electron micrographs of lens fiber cells from 5-M-old hsf4null zebrafish and WT zebrafish. Nucleus (red arrow), mitochondria (black arrow) and vesicular structures (white arrow) are obvious features in hsf4null lens fiber cells (a–e). In addition, the abbreviations including N (nucleus), M (mitochondria), EE (early endosome), LE (late endosome) and L (lysosome) were marked in the picture to emphasize the organelles' persistence. The connection between the fiber cells is loose in the cortical fiber region (a and b). (c) The fiber cells in the inner region of lens have an intact nucleus. (d and e) The mitochondria and vesicular structures including EE, matured LE and LE/lysosome fusion structure depositing in the lens fiber cells. The white arrows indicate the LE/lysosome fusion structure. (f) Showing that no cell apparatus existed in the WT lens fiber cells. (B) Enlargement pictures of the white box labeling areas showing that intact mitochondria existed in lens fiber cells. Subpanel a corresponds to white box in Aa. (b and c) Upper and lower white boxes in Ad, respectively. (C) Enlargement pictures of black box labeling areas in Ae showing the endosomes at different stages. (a) EE, (b) LE and (c) LE/lysosome fusion structure
hsf4 loss interrupts balanced cell proliferation and differentiation, which leads to pathological lens fiber architecture in Zebrafish lens
To testify whether the number of the lens epithelium cell was also increased in hsf4null zebrafish, we performed paraffin section and DAPI staining on 5-M-old zebrafish. Under microscopy observation, we confirmed that the epithelial cells were overproliferated. In adult lens, the proliferation activity at the central region of the lens epithelium is extremely low; thus, overproliferation was more obvious and significant in the germinal zone (Supplementary Figure 2).
We performed frozen section and phalloidin staining to determine whether the overproliferated cells at the germinal zone would produce excessive number of lens fiber cells. Under microscopy observation, we found that the fiber cell was labeled with phalloidin immunofluorescence, forming a hexagonal fluorescence circle. However, these circles stack together to generate a regular structure (Supplementary Figure 4). The WT lenses from every time point we checked displayed the regular arrangement pattern. In contrast, notable differences showed in hsf4null lenses. Compared with the WT lens, the fluorescence signals in hsf4null lens were denser but well organized at 2M, suggesting that more fiber cells existed in hsf4null lens (Figure 4a). At 6M, the standard arrangement pattern was severely interrupted (Figure 4b). By the age of 8M, the signals almost completely disappeared (Figure 4c). In addition, we could also find some nuclei mislocalized in the anterior region of the 6 and 8M hsf4null lenses. The above findings indicated that the arrangement of the fiber cells was disturbed (Figures 4b and c). Together, these results suggest that the overproliferation of the lens epithelial cells contributes to the excessive accumulation of fiber cells in the hsf4null lens with age, which causes chaotic arrangement of the fiber cells in the limited space, leading to cataractogenesis.

Progressively disordered arrangement of lens fiber cells in the in the hsf4null zebrafish lens. Immunostaining the WT and hsf4null zebrafish lens from different ages with antiphalloidin antibody. (a) The signals are more denser in the hsf4null zebrafish than the WT zebrafish at 2M of age. (b) The fiber cell structures becoming more disorganized in the hsf4null zebrafish at 6M of age. (c) The irregular signal disappeared in the hsf4null zebrafish at 8M of age. The white arrows indicate the mislocated fiber cell nucleus in the anterior region of the lens. Scale bar: 20 μm
HSF4-governed p53 stability and nuclear localization are essential to activate the Fas-mediated apoptotic signaling pathway
Continued with our previous research, we further investigated the roles of HSF4 and p53 in fiber cell differentiation. Firstly, we confirmed that HSF4 could stabilize p53 in human lens epithelial cell lines (HLECs). As expected, cells expressing GFP-HSF4 showed a stark enrichment of endogenous p53 in the nuclei (Figure 5a). Moreover, the endogenous p53 was predominantly localized in nuclei in a diffused pattern in cells transfected with GFP vector. In contrast, p53 was more focused and oriented in the nuclei but not in the cytoplasm of the cells expressing GFP-HSF4 fusion protein (Figure 5a). Western blot analysis of p53 levels in the separated nuclear and cytoplasmic fractions confirmed our immunofluorescence data (Figure 5b). On the other hand, p53 was obviously decreased in the nucleus when HSF4 was silenced using either of two HSF4-specific siRNAs (Figure 5c).

HSF4 stabilizes and orients p53 in the nucleus to initiate. (a) p53 is retained in the nucleus by HSF4. HLECs were transfected with GFP-tagged HSF4 (green) plasmids and negative control GFP vectors, respectively. Cells were harvested 48 h after transduction followed by immunofluorescence stains with anti-p53 antibody. Arrows indicated the overlap of the p53 signals (red) and the HSF4 signals (green). Scale bar: 10 μm. (b) The nuclear distribution of p53 was increased when HSF4 was overexpressed in HLECs. (c) Nuclear-oriented p53 was decreased when HSF4 was silenced in HLE cells. The cytoplasmic and nuclear proteins were separated and subjected to western blot detection. Tubulin and FBL were used as specific markers for the cytoplasm and nuclear components, respectively
As previous studies have revealed that apoptotic regulators are implicated in regulation of lens differentiation, we tested Fas and Bax, two target genes of p53 that encode proteins mediating extrinsic and intrinsic apoptotic pathways. First, through real-time PCR analysis, we found out that both Fas and Bax were significantly upregulated when HSF4 was overexpressed in HLECs (Figure 6a). In contrast, if HSF4 was silenced both Fas and Bax mRNA were significantly decreased (Figure 6b). Western blot results confirmed the changes of Fas and Bax at the protein level parallel with them at the mRNA level (Figures 6d–g). In addition, we noticed that the executor caspase3 was significantly activated in the cells expressing GFP-HSF4 compared with GFP control (Figure 6d). Consistent with this observation, the cleaved-caspase3 was decreased when we silenced HSF4 in HLECs (Figure 5f). Second, we determined whether HSF4-mediated upregulation of Fas and Bax was p53-dependent. To do so, the p53-null H1299 cells were transiently transfected with GFP vector or GFP-HSF4 fusion protein expression constructs. After 48 h, these cells were harvested for real-time PCR and western blot analysis. The results showed that both Fas and Bax were not altered at either mRNA or protein level (Figures 6c and h). Increased activation of caspase3 could not be detected in H1299 cells expressing GFP-HSF4 (Figure 6h). Thus, our results demonstrated that the activation of the Fas-mediated apoptotic signaling pathway by HSF4 was p53-dependent.

HSF4 can initiate the Fas-mediated apoptosis signaling pathway in HLECs. (a) Fas and Bax were upregulated at the mRNA level when HSF4 was overexpressed in HLECs. GFP-tagged HSF4 and negative control GFP plasmids were transfected into HLE cells. And then cells were harvested for RNA extraction and real-time PCR detection. Five independent experiments were performed and the relative mRNA levels were normalized to β-actin. (b) Fas and Bas were downregulated at the mRNA level when HSF4 was silenced in HLECs. The HSF4-specific siRNAs were transfected into HLECs. And then cells were harvested for RNA extraction and real-time PCR detection. Five independent experiments were performed and the relative mRNA levels were normalized to β-actin. (c) The upregulation of FAS and BAX was p53-dependent. GFP-tagged HSF4 and negative control GFP plasmids were transfected into H1299 cells. And then cells were harvested for RNA extraction and real-time PCR detection. Five independent experiments were performed and the relative mRNA levels were normalized to β-actin. (d) HLECs were transfected with GFP-HSF4 plasmids and negative control GFP vectors. Cells were harvested 48 h after transduction followed by protein extraction and western blot detection. (e) Statistical analysis of western blot detection result in d. The relative protein level was normalized by α-tubulin. Three independent experiments were performed. (f) The HSF4-specific siRNAs were transfected into HLECs. And then cells were harvested for protein extraction and western blot detection. (g) Statistical analysis of western blot detection result in f. The relative protein level was normalized by α-tubulin. Three independent experiments were performed. (h) H1299 cells were transfected with GFP-HSF4 and negative control GFP vectors. After 48 h later, cells were harvested for protein extraction and western blot detection. (i) Statistical analysis of western blot detection result in f. The relative protein level was normalized by the α-tubulin. Three independent experiments were performed. The overexpression and RNA-interfering efficiency were validated simultaneously, and the results were listed in Supplementary Figure 5 and Supplementary Figure 6
hsf4 loss can be partially rescued by overexpression of hsf4, p53 and fas
As HSF4 can regulate p53 stability and control Fas and Bax in a p53-dependent manner, we next sought to determine the functions of HSF4 regulation of p53 and its downstream target genes in governing lens development. To do so, we extracted proteins from hsf4null and WT zebrafish lenses. Western blot analysis revealed that p53 was significantly downregulated in hsf4null lens (Figures 7a and b). As p53 was downregulated, the activation of caspase3 was also significantly decreased, whereas the protein level of procaspase3 was not affected at two different time points we tested (Figures 7a and b). Together, these results demonstrated that HSF4 can induce caspase activation by stabilizing p53 in vivo.

hsf4 loss can be partially rescued by overexpression of hsf4, p53 and fas. (a) Three-M-old WT and hsf4null zebrafish lenses were isolated and subjected to protein extraction. Western blot results indicated that p53 and the activation of caspase3 was significantly decreased in the hsf4null zebrafish. (b) Western blot analysis of the proteins from 6-M-old WT and hsf4null zebrafish lenses. The results revealed that p53 and cleaved-caspase3 were also obviously decreased in the 6-M-old hsf4null zebrafish lens. (c) Overexpression of hsf4, p53 and fas could partially rescue the denucleation defect in hsf4null zebrafish lens. The hsf4, p53 and fas mRNA was microinjected into the hsf4null zebrafish embryos. The denucleation status was checked at 3 and 5d, respectively, through the DAPI staining. Obviously, decreased numbers of nuclei were presented in the lens injected with hsf4, p53 and fas mRNA at both 3 and 5d. Scale bar, 20 μm
To further test the functions of HSF4-controlled p53 and its target gene in regulating lens differentiation, we overexpressed these genes to determine whether they can rescue the denucleation defect in hsf4null lens. To do so, we collected hsf4null embryos and performed microinjection using hsf4, p53 and fas mRNA. As we have shown that the hsf4null zebrafish displayed obvious defects in degrading the nuclei of the lens fiber cells after 3 dpf, we collected the injected embryos at 3 and 5d. Through frozen section and DAPI staining, we found that hsf4, p53 and fas could partially rescue the denucleation defect in hsf4null lens (Figure 7c). These results demonstrated that hsf4 can regulate lens fiber cell differentiation by regulating p53 and its downstream apoptotic regulators.
