
Abstract
Later prehistoric woodland decline over most parts of Scotland is widely regarded as having been anthropogenic, via a range of mechanisms, to create farmland. Climatic causes are seen only to have driven the rapid expansion and then terminal decline of Pinus sylvestris around 2000 cal BC. Here we report radiocarbon dated analyses of pollen, microscopic charcoal, coprophilous fungal spores and peat humification from a small, water-shedding interfluve peat bog at 230 m elevation on the west-facing slope of the mountain Ben Lomond in west-central Scotland. The record spans the interval ca. 3450 − 200 cal BC. It shows marked and rapid changes in woodland composition before ca. 2600 cal BC, and from then to ca. 1940 cal BC a gradual decline of Betula woodland. This happened with no palaeoecological or archaeological evidence for anthropogenic activity. Woodland decline is interpreted at this site as climatically driven, perhaps through paludification or, more likely, exposure to wind, within a period of pronounced climatic deterioration. Anthropogenic activities are hinted at only after ca. 850 cal BC.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Although spatially patchy, palynology is well advanced in Scotland (Edwards et al. 2019) and has been central to models of Holocene vegetation change (Pennington 1974; Birks 1977, 1988, 1989; Edwards 1988; Lowe 1993; Bennett 1989, 1994; Tip** 1994, 1996). In the mid-Holocene, deciduous Quercus-Ulmus-Corylus woods covered the lowlands and coastal hinterland, Pinus and Pinus-Betula woods grew on higher ground and on acid soils north of the Midland Valley, and more open Betula-Corylus woods in the far north. The upland and montane expanses have been defined as agriculturally marginal because of persistent climatic and pedological constraints (Parry 1978), and Scotland was sensitive to periods of climate change (Bond et al.1997; Gilbertson et al. 1999; Charman et al. 2006; Tip** et al. 2013; Baker et al. 2015; Gallego-Sala et al. 2016; Stewart et al. 2017). Nevertheless, woodland decline and loss have almost always been interpreted as anthropogenic in origin because they were diachronous over short distances and small in scale until the late Iron Age ca. 300 cal BC. Woodland clearance was followed closely in time by the expansion of farmed land, through direct clearance of trees and seedling suppression by livestock grazing or vegetation burning. Until the late Iron Age, woodland decline was generally short-lived, followed by woodland regeneration (Turner 1965, 1975; Birks 1988; Buckland and Edwards 1984; Tip** 1994, 1997; Edwards and Whittington 2003; Edwards 2004; Edwards et al. 2019). Exposure to storminess has been used to explain woodland loss on islands around the northern mainland (Birks and Madsen 1979; Keatinge and Dickson 1979) but anthropogenic woodland loss is also argued for (Farrell 2015). Climatic changes related to abrupt hydrological shifts are more frequently asserted to have driven the abrupt expansion in range, and the equally abrupt contraction in range, of Pinus sylvestris woodland in the mountains and moors of central and northern Scotland between ca. 3400 and ca. 2200 cal BC (Bridge et al. 1990; Gear and Huntley 1991; Lowe 1993; Huntley et al. 1997).
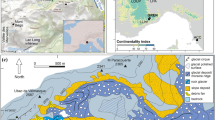
The Grampian Mountains are in central Scotland. They are sharply differentiated topographically, geologically and ecologically from the lowland trough of the Midland Valley to the south by the ‘Highland line’ (Whittow 1977; Fig. 1a, b). This division has resulted in significant long term agricultural and economic impoverishment of the Highlands (Forden 1380 in Smout 1969, p 39). In the west the fjord of Loch Lomond, close to sea level, pierces for 36 km into the Grampian Mountains, ringed to the west, north and east by hills rising above 600 m above sea level (a.s.l.) and formed of metamorphosed psammites and pelites. On its eastern side is Ben Lomond (974 m a.s.l.). Ramsay and Dickson (1997) summarised the vegetation history of central Scotland and described small-scale human impacts in lowland areas from ca. 2000 cal BC, becoming much more extensive, and often permanent, in the final centuries cal BC.

(a) The location of Scotland in north-west Europe, (b) the topography of Scotland (Robinson et al. 2014) marking the major divisions of the Grampian Mountains and the Midland Valley and the location of Fig. 1c; (c) the topography, drainage of central Loch Lomond (contour interval 10 m) and the pollen sites Coille Mhor Hill (this study), Dubh Lochan (Stewart et al. 1984) and Ptarmigan (Stewart 1979). Contains OS data © Crown copyright and database right (2021)
However, Stewart et al. (1984) described at Dubh Lochan on the shore of Loch Lomond, at the foot of Ben Lomond around 15 m a.s.l. (Fig. 1c) two pollen records, one from fen (DL) and one from lake sediment (DLM). They are strongly atypical of central–southern Scotland (Tip** 1994) in showing the persistence of deciduous woodland throughout the Holocene. The DLM core reflects changes in plant communities within several hundred metres of the lake (Stewart et al. 1984, p 538). From ca. 4000 cal BC until the BC/AD boundary, Alnus and Betula values are unchanging at around 20 and 15% (total land pollen (TLP)), respectively. Abruptly higher Quercus percentages, reaching 25% TLP at ca. 3300 cal BC are followed by a consistent and gradual decline to around 10% TLP at the cal BC/AD boundary, a pattern reflected also in Coryloid (Corylus avellana-type) pollen. Careful work at the DL site showed, however, that as Myrica became established on the fen, Corylus contributed < 10% TLP after ca. 4000 cal BC. Poaceae values rose after ca. 1250 cal BC when Plantago lanceolata (ribwort plantain) was more frequently recorded, but the highest values of Poaceae are small. The interpretation was that “Iron Age man did not have a large long-lasting effect on the vegetation around the Dubh Lochan” (Stewart et al. 1984, p 544). Lowland woodland is, to an extent, extant (Tittensor and Steele 1971).
Directly above Dubh Lochan, the steep slopes of Ben Lomond are treeless save for exclosures and recent conifer forest. To contextualise the unusual pollen record at Dubh Lochan, we sought to understand woodland decline on these slopes, providing better data than Stewart’s (1979) skeletal and undated pollen record at 500 m a.s.l. on Ben Lomond at Ptarmigan (NS 365 015; Fig. 1c). This paper reports new radiocarbon dated pollen, microscopic charcoal, fungal and humification data for an upland bog directly above Dubh Lochan and is designed to understand later Holocene vegetation dynamics and test the suggestion that prehistoric human impact was minimal on these slopes.
Coille Mhor Hill
Coille Mhor Hill is uphill and 2.5 km north of Dubh Lochan (Fig. 1c). It is a 30 ha rounded bedrock knoll on the western flank of Ben Lomond at 255 m a.s.l. East of Coille Mhor Hill rise the uniformly smooth and steep (18°) slopes of Beinn Urd (597 m a.s.l.). Two streams drain a col where peat has accumulated. The slopes around are formed in Ben Ledi Grit, an acid, nutrient poor metamorphosed sandstone. Soils are either skeletal, brown earths (brown forest soils in Scotland) or podsols (Tittensor and Steele 1971). They support, using Rodwell’s (1992) classification, species-poor Nardus stricta-Galium saxatile (U5) and Pteridium aquilinum-Galium saxatile (U20) grassland with Trichophorum- Eriophorum germanicum sedge bog (M17) and Juncus-rich Carex echinata-Sphagnum poor fen (M6d) around flushes and mosaics of Vaccinium myrtillus-rich Trichophorum germanicum-Erica tetralix grass-heath (M15), Carex echinata-Sphagnum poor fen (M6), Juncus and moss-rich (M17c) on shallow peat. Mean maximum and minimum temperatures 1961–1990 are around 10.7 and 3.3 °C, respectively, and mean precipitation around 1,540 mm (Harrison 1997).
Reconnaissance on the broad interfluve between the two streams located a bog, < 0.1 ha in area, around 60 m long north-west to south-west and little more than 20 m wide, with > 2 m deep peat at around 230 m a.s.l. (NS 37,432 99,068), draining south-east. The bog supports Erica tetralix-rich, Molinia caerulea-Potentilla erecta mire (M25). The pollen source area is hard to estimate. The basin is small and close to dryland soils, suggesting an extra-local pollen source area sensu Jacobson and Bradshaw (1981) but is at high altitude and barely sheltered from prevailing south-westerly winds such that the potential source of some pollen is best regarded as regional sensu Jacobson and Bradshaw (1981), and is increasingly so with woodland loss (cf. Bunting and Tip** 2004). It is assumed that woodland grew near and on the site when tree pollen (including Corylus avellana-type) reaches around 70% TLP (cf. Fossitt 1994) and that local plant communities contributed most pollen at these times.
The hillside has benefitted from detailed archaeological survey. Prehistoric structures are very rare, which contrasts with the frequent occurrence of Medieval and later farms and field systems, transhumant shielings and charcoal-burning iron-smelting sites (Boyle and MacInnes 2000). The bog is close to the generally accepted upper limit of sedentary prehistoric settlement in Scotland (Cowley 1998). Coille Mhor means ‘big wood’ in Gaelic, an early historic usage.
Sampling, materials and methods
After reconnaissance, survey and peat-stratigraphic description of the bog using a 1 m-long Eijelkamp peat gouge, the base of the bog was levelled by theodolite to an arbitrary datum to establish that the bog was free draining and sensitive to past water table fluctuations. A core was taken at the deepest point of the bog (259 cm) with a Russian-type corer of 1 m length and 6 cm internal diameter, placed in clean plastic guttering, wrapped, described in the laboratory and stored at 4 °C. The 32 samples taken for pollen and fungal analysis were each 5 mm thick. Subsamples were prepared by standard chemical methods (Moore et al. 1991). Mineral matter is absent above the basal sediment and hydrofluoric acid was not needed. Lycopodium spores (Stockmarr 1971) were added to calculate pollen concentrations and influx. Residues were embedded in silicone oil. Pollen identifications were made using an Olympus BX40 microscope. Counts were made at a magnification of ×400 with critical examinations and size measurements being made at a magnification of ×1,000 under oil immersion, sometimes using phase contrast. Pollen grains were identified with reference to Moore et al. (1991), the University of Stirling pollen reference collection, specialist keys and on-line images. Poaceae pollen grains with a-axes > 35 μm were not encountered. Counts were to a minimum of 300 TLP excluding Cyperaceae. The pollen sum is % TLP-Cyperaceae, excluding aquatics and spores. Pollen nomenclature follows Bennett (1994) and plant nomenclature follows Stace (2010). Pollen preservation was recorded in five categories (Cushing 1967). Microscopic charcoal was quantified in five size classes of a-axis lengths 10–25, 25–50, 50–75, 75–100 and > 100 μm. Six types of fungal spores were recorded from samples prepared for pollen analyses (cf. van Asperen et al. 2016) using photomicrographs in van Geel and Aptroot (2006) and van Geel et al. (2011). Values are given as % TLP + fungi. Most represent coprophilous (dung) fungi. Coniochaeta-type is found in both dung and wood, particularly Alnus and Fraxinus, Cercophora-type in both dung and wood, particularly Betula, and Gelasinospora-type is found in association with dung, charred wood and Vaccinium (Farr and Rossman 2009).
The pollen diagram is divided into six local pollen assemblage zones (LPAZ 1–6) with the aid of stratigraphically constrained cluster analysis (Grimm 1987). Humification by colorimetry was measured on contiguous 10 mm-thick sediment slices on a Jenway 6061 colorimeter at 540 nm (Blackford and Chambers 1993). Horizontally bedded, young, single entity roundwood samples from four depths were 14C dated by AMS. The 14C dates were calibrated using OxCal v4.3.2 r5 (Bronk Ramsey 2017) and the IntCal13 atmospheric curve (Reimer et al. 2013). An age-depth model of calibrated age estimates were generated using BACON v2.2 software (Blaauw and Christen 2011) with a step size of 5 cm.
Results
Table 1 describes the sediment stratigraphy at the sampled site. Table 2 lists the samples and depths from which AMS 14C assays were obtained. Figure 2 depicts the age-depth model generated by BACON. Linear regression through all depths has an r2 value of 0.99, suggesting a more-or-less constant peat accumulation rate of 12 ± 5 year cm− 1. In detail, however, peat accumulation falls from around 20 year cm− 1 to 8 year cm− 1 from the base to 125 cm depth and is constant at around 12 year cm− 1 above 125 cm depth. All age estimates in the text are calibrated and, unless stated, are age ranges modelled in BACON. Figure 3 is the complete percentage-based pollen, microscopic charcoal, fungal and humification record, plotted against depth. There is no evidence from the humification data for a long-term trend in decay, so the data are not detrended. The mean percentage humification is 16.8 ± 4.6%: higher percentages indicate less humified peat. The 1σ standard deviation around the mean (not plotted in Fig. 3 to retain clarity) is used to define significant changes in bog surface wetness. Figure 4a plots influx values (pollen accumulation rates) for major pollen taxa, used to infer local presence. Figure 4b presents the pollen data for major arboreal taxa on a % AP sum to allow direct comparison with the Dubh Lochan pollen record (Stewart et al. 1984). Figure 4c shows the percentages of well preserved (normal in Fig. 4c) and deteriorated pollen of four taxa common throughout the pollen record, calculated as % TLP. Pollen preservation is generally good. Interpretation focuses on fluctuations in proportions of corroded pollen because corrosion is closely linked to fluctuations in the peat water table (Havinga 1984).

Age-depth model for the peat on Coille Mhor Hill generated by BACON, showing (a) the positions of the calibrated 14C assays, the 2σ age range constrained within black dotted lines, all probable age-depth models in grey, darker areas with increasing probability and the best model based on the weighted mean average by the red dotted line, (b) the number of MCMC iterations in the model; (c) prior (curve) and posterior (histogram) distributions for accumulation rate estimates and (d) the ‘memory’ in accumulation rate estimations

Peat humification and percentage-based (% TLP) diagram at Coille Mhor Hill as follows: (a) pollen and spores (% TLP + spores), (b) coprophilous fungal spores (% TLP + fungi) and (c) microscopic charcoal (% TLP + charcoal), plotted against depth

(a) Influx values at Coille Mhor Hill of major taxa and charcoal fragments; (b) major taxa as % AP and (c) preservation characteristics of major taxa as % TLP. All data are plotted against depth
LPAZ 1: 251 to 233 cm depth (3525 − 3104 to 3272 − 2786 cal BC)
A young twig lying horizontally at the base of the peat is 14C dated to 3625 − 3372 cal BC, modelled to 3525 − 3104 cal BC, i.e. the early–mid Neolithic of the British Isles (ca. 4000 − 3300 cal BC; Bradley 2007). Peat accumulation above this was around 20 year cm− 1. Peat was significantly less humified than the mean until 3259 − 2771 cal BC (232 cm depth; Fig. 3). Corroded and degraded pollen grains are common (Fig. 4c), suggesting that the peat in LPAZ 1 was an aerated organic soil. The high influx values of Alnus glutinosa, Betula, Corylus avellana-type and Quercus in the basal pollen spectrum (Fig. 3) may include pollen from non-contemporary sources. Above this, Alnus pollen is recorded at percentages large enough to imply local presence before 3396 − 2924 cal BC (241 cm depth) (Tinsley and Smith 1974; Gearey and Gilbertson 1997) but influx values larger than 1,000 grains cm2 yr− 1 indicate local presence throughout LPAZ 1 (Abraham et al. 2021). It is likely that Alnus colonised the basin when soil was paludified. Betula and Salix accompanied Alnus, influx values for Betula indicating local presence by 232 cm depth (Tinsley 2001; Abraham et al. 2021). Percentages of Salix pollen underrepresent the plant. The community was probably close to the Juncus effusus subcommunity of W4 Betula pubescens-Molinia caerulea woodland (Rodwell 1991) where Alnus can outnumber Betula trees and Salix bushes in an open canopy. Corylus avellana-type pollen in this setting may more likely represent C. avellana but Myrica gale cannot be dismissed (Skene et al. 2000). If from Corylus, the low percentage representation and influx values < 2,000 grains cm2 yr− 1 in mid-zone are taken to mean the tree was not common on the hillside (Tinsley and Smith 1974; Lisitsyna et al. 2011; Abraham et al. 2021). Quercus, from its low influx, may not have been local (Tinsley 2001). Ulmus was rare if present. P. sylvestris was not present.
The wood may have been open, although total tree + shrub pollen percentages are very high, exceeding 80% TLP, or there were open areas. Less stable soils, perhaps gullied, supported plants producing Ulex-type pollen (probably Ulex europaeus). Proportions of Poaceae fluctuate, though average influx values around 5,000 grains cm2 yr− 1 imply local presence (Abraham et al. 2021). Grassland herb taxa were common, though their proportions are low, with Potentilla-type constant, two grains of Rhinanthus-type (perhaps Euphrasia), and Ranunculus, Rumex acetosa, Apiaceae and Melampyrum. Dwarf shrubs (Ericaceae, Empetraceae) were rare. Single spores of the coprophilous fungi Podospora- and Sordaria-type are recorded. Microscopic charcoal is very rare.
LPAZ 2: 233 − 207 cm depth (3272 − 2786 to 2860 − 2452 cal BC)
Alnus influx values > 5,000 grains cm2 yr− 1 continue to indicate its local abundance (Abraham et al. 2021) despite an abrupt fall in percentages at 233 cm depth (3272 − 2786 cal BC). Alnus may have been confined to the peat. Alnus was joined by Betula, probably rapidly in a few decades between 3388 − 2900 and 3312 − 2811 cal BC and later (225 cm depth; 3153 − 2657 cal BC) by an equally short-lived increase in Quercus values to around 50% TLP, and 25,000 grains cm2 yr− 1 , suggesting the tree was locally abundant (cf. Tinsley and Smith 1974; Tinsley 2001). A hiatus in deposition is not suggested by the highly resolved humification data. Although humification data suggest the peat surface became drier between 3259 − 2771 (232 cm depth) and 2924 − 2511 cal BC (212 cm depth) and was very dry (> 1σ) by 2951 − 2528 cal BC (214 cm depth), proportions of corroded pollen in major taxa fall, interpreted as meaning that reworked pollen grains from the organic soil were few. The shift in woodland trees is more likely to have been driven by the drying of the peat surface observed in the humification data (cf. McVean 1956a). In western Scotland today, the altitudinal limit of Quercus is around 150 m a.s.l. (McVean 1964) but Jones (1959, p 176) described Q. petraea woods up to 240 m a.s.l. in north Wales and, with Betula, in western Ireland up to 340 m a.s.l.
Corylus avellana-type pollen percentages remain comparatively low and unchanging. On very base-poor siliceous soils there can be few other canopy species with Betula and Quercus (McVean 1956b; McVean and Ratcliffe 1962; Steele 1974). The absence of Vaccinium-type pollen might indicate that Coille Mhor Hill was within the grass-rich Betula-herb nodum of Scottish Betula-Quercus woods (McVean and Ratcliffe 1962, p 16), or the species-poor W16 Quercus-Betula-Deschampsia flexuosa woodland of Rodwell (1991). Total tree + shrub pollen percentages are unchanging, and the woodland may have been closed. There are no other prominent palynological changes. Calluna (ling) is characteristic of upland and base-poor Betula-Quercus woodland but was rare at this time at Coille Mhor Hill. Coniochaeta-type spores are recorded in all counts. Microscopic charcoal is very rare.
LPAZ 3: 207 − 176 cm depth (2860 − 2452 to 2473 − 2092 cal BC)
Humification data (Fig. 3) suggest the bog surface was significantly wetter than the mean in this LPAZ. Despite this, Alnus values (% and influx) do not increase. Corroded Betula pollen grains are fewer because of this hydrological shift but pollen of other taxa do not show this. A second abrupt increase of Betula to 65% TLP and > 40,000 grains cm2 yr− 1 occurred at 2860 − 2452 cal BC (207 cm depth), an influx that probably implies dense woodland at the pollen site (Tinsley 2001). Betula may have become the only tree taxon at and around the pollen site. The peak in Betula percentages at 204 cm depth (2822 − 2404 cal BC) was followed by a gradual, consistent decline but influx values indicate a more temporally confined expansion and fall centred on 190 cm depth (2638 − 2227 cal BC). Quercus representation fell to around 10% TLP and 2,500 grains cm2 yr− 1, not necessarily indicative of local presence (Tinsley 2001; Abraham et al. 2021). Quercus may have persisted locally or elsewhere on the hillside or in the region; much of its pollen in this and later zones may have been regional in origin (cf. Lisitsyna et al. 2011) so that the woodland on Coille Mhor Hill may have remained floristically similar to that in LPAZ 2. Betula and Quercus are ordinarily intimately associated (McVean and Ratcliffe 1962, p 16) but Jermy et al. (1978, p 10.22) noted that Betula displaces Quercus through edaphic controls on steep slopes on thin, increasingly base poor, degraded brown earth soils that become with time more acid and increasingly podzolic. Quercus prefers deeper soils with accumulating nutrients at the base of slopes. Betula improves the base status of very poor soils (Miles 1981) but there are no changes in representation of C. avellana-type pollen to suggest that base status increased in soils on Coille Mhor Hill. Probably because of a dense cover of Betula, pollen of Ulmus, Calluna, Ranunculus-type, Rosaceae, Rumex acetosa, Apiaceae and Melampyrum (also Polypodiaceae, i.e. undifferentiated fern spores) are only infrequently recorded, as is Sphagnum even though it probably grew on the peat. Coniochaeta-type spores are recorded in all counts. Dung fungi are present, including Sordaria-type and Podospora-type. Microscopic charcoal is very rare.
LPAZ 4: 176-118 cm depth (2473 − 2092 to 1829 − 1625 cal BC)
From being wetter than the mean at 2473 − 2092 cal BC (176 cm depth), the humification data suggest that the bog surface, became very dry by 1853 − 1652 cal BC (120 cm depth) for around 50 years, and then continued to be drier than the mean. After 2527 − 2143 cal BC (180 cm depth), Betula and Alnus pollen grains are more corroded, particularly so between 2315 − 1943 (160 cm depth) and 1972 − 1718 cal BC (130 cm depth), supporting the interpretation of the humification data. Wood fragments in the peat are few above 140 cm depth (2097 − 1784 cal BC). Betula percentages continue a sustained fall to around 20% TLP by 140 cm depth (2097 − 1784 cal BC) and influx data show a decline after 140 cm depth. Both represent declining woodland extent, thinning of the canopy or pollen productivity, after which proportions and influx values are unchanged. Nevertheless, Betula persisted locally (cf. Tinsley 2001; Abraham et al. 2021). Percentages of Betula remain much higher than in surface pollen studies in Scotland where Betula is of long-distant origin (Gearey and Gilbertson 1997; Bunting 2002; Fossitt 1994).
Quercus pollen is slightly better represented, at around 15% TLP, except for a few decades in mid-zone (145 − 141 cm depth: 2156 − 1821 to 2106 − 1794 cal BC) where it is > 20% TLP, Influx values around 2,500 grains cm2 yr− 1 probably imply local presence (cf. Tinsley and Smith 1974; Tinsley 2001). An increasingly open canopy may have permitted the more consistent representation of trees of regional origin such as Pinus and perhaps Fraxinus. Salix pollen grains are slightly better represented. Cyperaceae are better represented at the upper zone boundary. Filipendula was probably more common after 145 cm depth (ca. 2000 cal BC), in moist grassland, and so probably was Rumex acetosa. P. lanceolata is recorded for the first time at 131 cm depth (1982 − 1728 cal BC), at the end of the phase of sustained woodland decline but at values no more than 1% TLP. P. lanceolata pollen is over-representative of the plant (Tinsley and Smith 1974; Hjelle 1998; Shaw and Whyte 2020) and its rarity in the pollen record at Coille Mhor Hill suggests its rarity also in plant communities. It is absent in acidic soils in uplands (Sagar and Harper 1964) and rare at pH < 4.5 (Grime et al. 1988, p 438). There are very few other grazing indicator herbs. Although percentages of Poaceae increase, influx values do not. Dung fungi (Sordaria-type, Podospora-type) continue to be represented, as is Cercophora-type above 150 cm depth (1859–2203 cal BC). Coniochaeta-type is less well represented. Microscopic charcoal is slightly more common.
Melampyrum pollen is markedly more common above 145 cm depth (2156 − 1821 cal BC). The high proportion at 121 cm depth (1862 − 1661 cal BC) may be due to incorporation in the pollen sample of an anther, but it nevertheless attests to local presence, as do the comparatively high pollen percentages (Moore et al. 1986). The pollen cannot be identified to species but either M. pratense or M. sylvaticum, or both, are possible from current distributions (Tippett 1974; Lee 1933; Rich et al. 1998). Both inhabit upland open Betula woodland (> 30% shade) and woodland edges of herb-rich W11/W17 Quercus petraea-Betula pubescens woodland (Rodwell 1991), as well as heaths and upland moors on shallow, often skeletal, acid (pH 3.9–4.7), continually moist free-draining soils Dalrymple 2007; Rich et al. 1998, p 34) comment that M. sylvaticum plants are “usually found in the same precise spots each year” despite being an annual plant and, for this reason, Melampyrum species are sometimes regarded as ancient woodland indicators (Rackham 2003, p 54; Miles 1988: see also ‘Discussion’ below). It became a persistent feature on Coille Mhor Hill into LPAZ 5, until at least 90 cm depth (1533 − 1222 cal BC).
LPAZ 5: 118-6 6 cm depth (2473 − 2092 to 1234 − 948 cal BC)
According to the peat humification data, the bog surface remained from dry or very dry, although Salix, Cyperaceae and Equisetum, and falling proportions of corroded Alnus and Betula pollen, suggest wetter conditions. Proportions of tree genera suggest no significant changes to the partly open Betula woodland. This zone sees the establishment above 111 cm depth (1752 − 1488 cal BC) of Calluna. Other dwarf shrubs like Empetrum (crowberry), Erica (heaths) and Ericales (undifferentiated Ericaceae or Empetraceae) are not recorded until 91 cm depth (1541 − 1235 cal BC). Microscopic charcoal was recorded more frequently, probably because heather species were more common. P. lanceolata is the only anthropogenic indicator, occurring mostly as single grains. All fungal spore taxa are absent above the basal spectrum save single occurrences of Sordaria-type.
LPAZ 6: 66 − 0 cm depth (1234 − 948 to 739 − 38 cal BC)
Modelled age ranges are increasingly imprecise above the youngest 14C assay at 73 cm depth. Extrapolation of the peat accumulation rate (~ 15 year cm− 1) above 73 cm depth suggests that the peat surface has an age of ca. 2400 cal BC. The peat has been truncated above this, probably by peat cutting. Quercus percentages abruptly fall to a mean 2.5% TLP at 66 cm depth (1234 − 948 cal BC); influx values are unchanging. Alnus, probably not common on the peat at Coille Mhor Hill above LPAZ 2 (above), declined to values < 5% TLP by 11 cm depth (821 − 187 cal BC). Betula proportions were maintained. Influx values increased to > 5,000 grains cm2 yr− 1, indicating the persistence of Betula near the pollen site. Salix is unrecorded between 91 and 51 cm depth (1541 − 1235 cal BC to 1127 − 730 cal BC) and above 51 cm depth was not common, if present. Calluna would have gained an advantage in grassland communities and under the open Betula wood. Vaccinium-type pollen is recorded consistently for the first time, and considerable diversity in heath taxa is indicated by consistent records of Ericales and Erica, and above 31 cm depth (974 − 445 cal BC) by Empetrum. Humification data indicate the bog surface became wetter, after 1049 − 566 cal BC (40 cm depth), and after 974 − 474 cal BC (31 cm depth), so do increases in Sphagnum and Cyperaceae percentages. However, proportions of corroded pollen do not reflect this. Local growth of Melampyrum may have ceased because of this hydrological shift. Charcoal fragments < 50 μm are abundant but it is not clear from these whether fires were local. Dung fungi (Sordaria- and Sporormiella-type for the first time) are found sporadically.
Discussion
Peat inception at Coille Mhor Hill at 3625 − 3372 cal BC coincided with evidence in southern Scotland for increased effective precipitation (Langdon et al. 2003, 2012). It may have been that peat initiation was climatically induced. Alnus colonised lowland soils along Loch Lomond around 4800 cal BC (Dickson et al. 1978; Stewart et al. 1984). It grew locally at Coille Mhor Hill from peat inception. On the slopes of Ptarmigan, Stewart (1979) recorded Alnus pollen in proportions high enough to suggest that the tree grew at much higher altitudes than is commonly recognised (McVean 1953).
Climatic factors may have resulted in partial losses of Alnus at Coille Mhor Hill at 3272 − 2786 cal BC when the bog surface became increasingly dry, approaching very dry at 3128 − 2647 cal BC. This shift may reflect the end of a wetter and stormier climate centred on ca. 3200 cal BC (Magny and Haas 2004; Caseldine et al. 2005; Magny et al. 2006; Leuschner et al. 2007; Moir et al. 2010; Roland et al. 2015; Stewart et al. 2017). As the bog surface and surrounding soils became drier, Alnus lost competitive advantage, although the numbers of Coniochaeta-type fungal spores, often associated with Alnus, suggest its presence at Coille Hhor Hill until LPAZ 5 at 2473 − 2092 cal BC. Alnus did not, however, respond when bog surface wetness increased between 2951 − 2528 and 2848 − 2435 cal BC, perhaps because seed sources were by then distant. Among tree taxa, Betula seems to have responded first to the loss of Alnus. Open Betula woodland already grew at Ptarmigan (Stewart 1979). The low proportions of C. avellana-type pollen at Coille Mhor Hill are explained by nutrient impoverishment of soils (Godwin 1975, p 272; Theuerkauf et al. 2014).
Pinus sylvestris woodland spread south from the Grampian Mountains (Fig. 1b) after ca. 3650 − 3350 cal BC (Bridge et al. 1990), into the deciduous woodland north and north-west of Loch Lomond after ca. 2900 − 2750 cal BC (Dickson et al. 1978; Stewart 1979; Wylie and Dickson 1998), less than 15 km north of Ben Lomond. This was part of a short-lived expansion of range in Pinus from core areas in the Scottish Highlands. Expansion has long been seen in Scotland as climatically driven (Bridge et al. 1990; Gear and Huntley 1991) as Pinus colonised peat surfaces made drier through lowered water tables. Pinus did not invade the Quercus woodland at Dubh Lochan (Stewart et al. 1984) or the Betula woodland on Coille Mhor Hill, or on Ptarmigan (Stewart 1979), at least sufficiently to be palynologically detected. Deciduous trees continued to out-compete Pinus.
Quercus replaced, for a time, a single generation of Betula (upland Betula may live for around 200 years (Kirby 1984)), though most die when less than a century old (Rackham 2003). If local to the peat at Coille Mhor Hill, which is likely, the establishment of Quercus occurred at a higher altitude than can be observed in western Scotland at the present day (150 m a.s.l.: McVean and Ratcliffe 1962, p 16; Stewart 1979), probably explained by warmer temperatures 5,000 years ago (Davis et al. 2003). The loss of Quercus on Coille Mhor Hill after 2860 − 2452 cal BC, perhaps again of a single generation of trees, may have a locale-specific explanation such as soil acidification, operating at altitudes and on slopes where Quercus had anyway a tenuous foothold (Miles 1985; Jermy et al. 1978). This change is not seen in the pollen ‘rain’ of the region (Dickson et al. 1978; Ramsay and Dickson 1997; Fyfe et al. 2013).
Because Quercus did not later regain a foothold around Coille Mhor Hill, edaphic factors (Leuschner et al. 2002; Wardle et al. 2004) are unlikely to have induced its decline. There is no evidence in the Coille Mhor Hill record from (a) anthropogenic indicator pollen taxa developed for north-west Europe (Behre 1981; cf. Deza-Araujo et al. 2020), (b) microscopic charcoal or (c) coprophilous fungi for anthropogenic activities to have changed woodland composition or caused the demise of locally growing Quercus (cf. Hellman et al. 2009) on Coille Mhor Hill. Indicator taxa are almost absent, and not all, like P. lanceolata, are poorly represented on acid soils. The only moderate increases in grassland communities at Coille Mhor Hill may have been within open Betula woodland. Melampyrum is often seen to be indicative of anthropogenic disturbance (Tinsley 1975; Moore et al. 1986; Mitchell 1988; Innes et al. 2013) but Tittensor and Steele (1971) found M. pratense most commonly in the undisturbed island oakwoods of Loch Lomond, and Dalrymple (2007) regarded M. sylvaticum as sensitive to grazing pressure. Fire, sometimes associated with Melampyrum (Innes and Simmons 2000, though see Moore et al. 1986, p 215 and Blackford et al. 2006, p 198), is not indicated at Coille Mhor Hill because proportions of microscopic charcoal are very low. Other forms of disturbance such as hydroseral change on bog surfaces can favour the plant (Pilcher 1973; Birks 1975). Melampyrum became established on Coille Mhor Hill from ca. 2000 cal BC when humification data show the bog surface became very wet. It then persisted for around 650 years, to at least ca. 1350 cal BC. Its persistence is taken to indicate a lack of disturbance of grassland communities near or on the peat surface. It is possible that the later, more prolonged phase of dry bog surface after 1606 − 1307 cal BC adversely affected its growth. Numbers of Cercophora-type fungal spores, associated with Betula wood, track the representation of Melampyrum for reasons that are unclear, but which might also imply a lack of vegetation disturbance. They are absent after LPAZ 4, after the decline of Betula.
Dung fungi were comparatively abundant when the woodland was locally present, but they decline to absence from ca. 1600 cal BC with Betula woodland decline, when this vegetation change would have favoured fungal spore dispersal. This pattern is unexpected had grazing animals been the cause of, or gained from, woodland decline (Blackford and Innes 2006; Cugny et al. 2010). It is not explained by changes in sample preparation methods, pollen preservation or change in sediment type, and although an absence of evidence, for which there are many explanations (cf. van Asperen et al. 2020), the simplest interpretation is that grazing animals did not have a role in woodland change or decline.
Anthropogenic disturbance is also hard to identify when Betula pollen percentages declined in LPAZ 3 between 2822 − 2404 and 2097 − 1784 cal BC. Interpretation is complicated because, as local woodland declined, arboreal pollen became increasingly regional in origin (above), but in the region Betula pollen percentages increased at this time (Dickson et al. 1978; Fyfe et al. 2013). It is likely, then, that the pollen record at Coille Mhor Hill describes a local decline. The linear decline in Betula pollen percentages between ca. 2600 and ca. 1950 cal BC (r2 = 0.98: n = 8) might, in its consistency, reflect natural change rather than anthropogenic interference. The period when Betula woodland declined at Coille Mhor Hill was one of major hemispheric scale climate change (Bond et al. 1997; Karlen and Larsson 2007; Walker et al. 2012). The tolerance of both Betula pendula and B. pubescens to very large temperature variations (Atkinson 1992) suggests that thermal change (McDermott et al. 2001; Taylor et al. 2018; McKeown et al. 2019) would not have affected upland Betula woodland at 230 m a.s.l. in western Scotland. The Betula woodland decline at Coille Mhor Hill occurred in a period of predominantly high local bog surface wetness 2909 − 2429 to 1955 − 1713 cal BC. The bog surface wetness record at Coille Mhor Hill need not relate to climate change because the peat receives, as well as sheds, water, but there is good agreement with data from raised mosses in central Scotland that bog surfaces became wetter from around 2000 cal BC and were very wet ca. 1850 cal BC (Charman et al. 2006). Although Swindles et al. (2013) and Roland et al. (2014) argued that there is little consistent peat-stratigraphic evidence in the Irish Sea region for major hydrological change, Barber et al. (1994) in north-west England regarded this deterioration as the most significant in the Holocene. Bog and soil surface wetness may have become inimical for a time to the growth of Betula (Crawford et al. 2003), as it did for Pinus populations further north in Scotland (Bridge et al. 1990; Gear and Huntley 1991; Lowe 1993; Huntley et al. 1997), but such an effect could be expected to have been recognised by earlier workers. Attrition by exposure to high winds on this west facing slope is perhaps more likely (Lamb 1964; Allen 1992; Quine and White 1994; Paus 1995). Ecological data on how winds affect Betula are few, but Atkinson (1992) drew on the northern limits of B. pubescens in Greenland and Iceland in indicating its vulnerability to cold and dry winds. Our study area lies on one of the principal Atlantic cyclone tracks (Lozano et al. 2004). Increased aeolian activity is commonly recorded in 14C-dated dune construction along the north-eastern Atlantic façade between 2500 and 2000 cal BC Gilbertson et al. 1999; Wilson et al. 2001, 2004; Sommerville et al. 2007; de Jong et al. 2009; Orme et al. 2015; Ballin-Smith 2018, p 19; Goslin et al. 2019), and Olsen et al. (2012) identified from a palaeo-limnological record in Greenland a strong, positive North Atlantic Oscillation before ca. 2350 cal BC.
After ca. 1950 cal BC the stresses imposed on Betula regeneration at Coille Mhor Hill appear to have eased. Betula pollen proportions do not decline further. Cercophora-type spores, if associated here with Betula, are not recorded after 1745 − 1464 cal BC, suggesting that the Betula record contains much long-distant pollen. The bog surface became very dry by 1853 − 1652 cal BC, then dry until 1606 − 1307 cal BC, before becoming wetter at 1533 − 1222 cal BC, tracking closely comparable changes in raised mosses in the region (Charman et al. 2006), and storminess subsided Wilson et al. 2001, 2004; de Jong et al. 2009; Orme et al. 2015).
Increases in Poaceae pollen in the region from ca. 1850 cal BC (Dickson et al. 1978; Fyfe et al. 2013) arise from the atmospheric mixing of pollen from many small and temporary anthropogenic activities that led to localised woodland clearance and settlement, and grazing-driven woodland losses (Turner 1965; Tip** 1994; Ramsay and Dickson 1997). Around Dubh Lochan at the foot of Ben Lomond, woodland decline on dry soils appears limited to Quercus, gradually from ca. 3300 cal BC and ceasing by the cal BC/AD boundary (Stewart et al. 1984). Further vegetation disturbance is seen at Coille Mhor Hill after 1054 − 580 cal BC, resulting in P. lanceolata being a component, though not common, of grassland. Betula pollen declines to values that suggest trees were not growing locally. Some dung fungi reappear sporadically after ca. 960 cal BC. If from domestic livestock, this is the first clear evidence in the pollen record at Coille Mhor Hill of human activity. It is not precisely dated but, as around Dubh Lochan (Stewart et al. 1984), later prehistoric people appear to have had little impact. There is a scarcity along Loch Lomond of archaeological sites from this period (Harding 2004; Alexander 2015; Lock and Ralston 2017). There is no indication that human activities at Coille Mhor Hill became more intense or extensive before truncation of the peat at ca. 2400 cal BC. The impact of Iron Age farming communities after ca. 500 cal BC elsewhere in central and southern Scotland, from similar anthropogenic indicator pollen taxa, was extraordinary (Turner 1965, 1979, 1981, 1983; Dumayne 1993a, b; Ramsay 1995; Ramsay and Dickson 1997; Tip** 1997, 2010; Dumayne-Peaty 1998, 1999; Dark 2000; Tip** and Tisdall 2006) as woodland clearance changed from small and temporary to large and, at some places, permanent, continuing into the period of Roman occupation after cal AD 80. Along the eastern shore of Loch Lomond, the palaeoecological data at Dubh Lochan (Stewart et al. 1984) and Coille Mhor Hill suggest agrarian impact was negligible.
Radiocarbon dated pollen records immediately north of Loch Lomond are few. In the Grampians, Bridge et al. (1990) describe Pinus-rich woodland which yielded in later prehistory to extensive paludification, blanket peat and wet heath spread with no evidence that these changes involved human impact. It is possible that the ‘Highland line’, a fundamental economic boundary in Scottish history, defined by sharply contrasting potentials for agricultural productivity through geological and climatic controls, had considerable significance for human ecology in later prehistory.
Conclusions
A 14C dated pollen, microscopic charcoal and coprophilous (dung) fungal record, supported by humification data, is described from a water-shedding peat, Coille Mhor Hill, on Ben Lomond, the most southerly slopes of the Grampian Mountains. The site is around 230 m a.s.l., on a steep and exposed west-facing slope. Mineral soils are very acid and nutrient poor. Analyses were developed to test the suggestion from Stewart et al. (1984) that later prehistoric people had almost no impact on the vegetation cover. Whilst later prehistoric human activities in southern Scotland are almost universally seen to have been causal in later Holocene woodland loss (Miles 1988; Tip** 1994; Edwards and Whittington 2003), we found that woodland decline on Coille Mhor Hill probably occurred without anthropogenic impacts. Because of this limited evidence for human interference, it has been necessary to explore in detail ecological and edaphic processes and relations, and to draw on palaeoclimatic data. We suggest that exposure to intense North Atlantic winds during climatic deterioration reduced the exposed west facing Betula wood at Coille Mhor Hill. How common this was in the uplands of Scotland needs to be explored. This interpretation does not replace the general model for central and southern Scotland of anthropogenic woodland decline (Tip** 1994; Edwards and Whittington 2003; Edwards et al. 2019); climatic impacts may have been restricted in extent. One implication, however, is that there may have been other areas of the Grampian Mountains that were almost empty of people in later prehistory. Another might be that the aphorism that “The crucial feature of upland vegetation within the natural forest zone in Britain is that it is largely man-made” (Miles 1988, p 57) needs to be re-visited.
Availability of data
on request.
References
Abraham V, Hicks S, Svobodová-Svitavská H et al (2021) Patterns in recent and Holocene pollen accumulation rates across Europe; the Pollen Monitoring Programme Database as a tool for vegetation reconstruction. Biogeosciences 18:4,511–534. https://doi.org/10.5194/bg-2020-217
Alexander D (2015) The Iron Age. In: MacAdam E (ed) Exploring our past: essays on the local history and archaeology of west-central Scotland. Glasgow Museums, Glasgow, pp 63–80
Allen JRL (1992) Trees and their response to wind: mid Flandrian strong winds, Severn Estuary and inner Bristol Channel, southwest Britain. Phil Trans R Soc Lond B 338:335–364
Atkinson MD (1992) Biological flora of the British Isles: Betula pendula Roth (B. verrucosa Ehrh.) and B. pubescens Ehrh. J Ecol 80:837–870
Baker A, Hellstrom JC, Kelly BFJ, Mariethoz G, Trouet V (2015) A composite annual-resolution stalagmite record of North Atlantic climate over the last three millennia. Sci Rep 5:10307. doi:https://doi.org/10.1038/srep10307
Ballin-Smith B (ed) (2018) Life on the edge: the Neolithic and Bronze Age of Iain Crawford’s Udal, North Uist. Archaeopress, Oxford
Barber KE, Chambers FM, Maddy D (1994) Sensitive high-resolution records of Holocene palaeoclimate from ombrotrophic bogs. In: Funnell BM, Kay RLF (eds) Palaeoclimate of the Last Glacial/Interglacial cycle. Special Publication 94/2. Natural Environment Research Council, Swindon, pp 57–60
Behre K-E (1981) The interpretation of anthropogenic indicators in pollen diagrams. Pollen Spores 23:225–245
Bennett KD (1989) A provisional map of forest types for the British Isles 5000 years ago. J Quat Sci 4:141–144
Bennett KD (1994) Annotated catalogue of pollen and pteridophyte spore types of the British Isles. Department of Plant Sciences, University of Cambridge, Cambridge https://www.researchgate.net/publication/245416986_Annotated_Catalogue_of_Pollen_and_Pteridophyte_Spore_Types_of_the_British_Isles. Accessed 12 January 2021
Birks HH (1975) Studies in the vegetational history of Scotland IV. Pine stumps in Scottish blanket peats. Phil Trans R Soc Lond B 270:181–226
Birks HJB (1977) The Flandrian forest history of Scotland: a preliminary synthesis. In: Shotton FW (ed) British Quaternary studies: recent advances. Clarendon Press, Oxford, pp 119–135
Birks HJB (1988) Long-term ecological change in the British uplands. In: Usher MB, Thompson DBA (eds) Ecological change in the uplands. Blackwell, Oxford, pp 37–56
Birks HJB (1989) Holocene isochrone maps and patterns of tree-spreading in the British Isles. J Biogeogr 16:503–540
Birks HJB, Madsen BJ (1979) Flandrian vegetational history of Little Loch Roag, Isle of Lewis, Scotland. J Ecol 67:825–842
Blaauw M, Christen JA (2011) Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal 6:457–474
Blackford JJ, Chambers FM (1993) Determining the degree of peat decomposition for peat-based palaeoclimatic studies. Int Peat J 5:7–24
Blackford JJ, Innes JB (2006) Linking current environments and processes to fungal spore assemblages: surface NPM data from woodland environments. Rev Palaeobot Palynol 141:179–187
Blackford JJ, Innes JB, Hatton JJ, Caseldine CJ (2006) Mid-Holocene environmental change at Black Ridge Brook, Dartmoor, SW England: a new appraisal based on fungal spore analysis. Rev Palaeobot Palynol 141:189–201
Bond G, Showers W, Cheseby M et al (1997) A pervasive millennial-scale cycle in North Atlantic Holocene and glacial climates. Science 278:1,257-1,266
Boyle SD, MacInnes L (2000) The historic landscape of Loch Lomond and the Trossachs. Royal Commission on the Ancient and Historical Monuments of Scotland, Edinburgh
Bradley R (2007) The prehistory of Britain and Ireland. Cambridge University Press, Cambridge
Bridge MC, Haggart BA, Lowe JJ (1990) The history and palaeoclimatic significance of subfossil remains of Pinus sylvestris in blanket peats from Scotland. J Ecol 78:77–99
Bronk Ramsey C (2017) OxCal ver. 4.3. https://c14.arch.ox.ac.uk/oxcal/OxCal.html. Accessed 9 September 2020
Buckland PC, Edwards KJ (1984) The longevity of pastoral episodes of clearance activity in pollen diagrams: the role of post-occupation grazing. J Biogeogr 11:243–249
Bunting MJ (2002) Detecting woodland remnants in cultural landscapes: modern pollen deposition around small woodlands in northwest Scotland. Holocene 12:291–301
Bunting MJ, Tip** R (2004) Complex hydroseral vegetation succession and ‘dryland’ pollen signals: a case study from northwest Scotland. Holocene 14:53–63
Caseldine C, Thompson G, Langdon C, Hendon D (2005) Evidence for an extreme climatic event on Achill Island, Co. Mayo, Ireland around 5200–5100 cal. yr BP. J Quat Sci 20:169–178
Charman DJ, Blundell A, Chiverrell RC, Hendon D, Langdon PG (2006) Compilation of non-annually resolved Holocene proxy climate records: stacked Holocene peatland palaeo-water table reconstructions from northern Britain. Quat Sci Rev 25:336–350
Cowley DC (1998) Identifying marginality in the first and second millennia BC in the Strath of Kildonan, Sutherland. In: Mills CM, Coles G (eds) Life on the edge: human settlement and marginality. Oxbow Books, Oxford, pp 165–171
Crawford RMM, Jeffree CE, Rees WG (2003) Paludification and forest retreat in northern oceanic environments. Ann Bot 91:213–226
Cugny C, Mazier F, Galop D (2010) Modern and fossil non-pollen palynomorphs from the Basque mountains (western Pyrenees, France): the use of coprophilous fungi to reconstruct pastoral activity. Veget Hist Archaeobot 19:391–408
Cushing EJ (1967) Evidence for differential pollen preservation in late quaternary sediments in Minnesota. Rev Palaeobot Palynol 4:87–101
Dalrymple SE (2007) Biological flora of the British Isles: Melampyrum sylvaticum L. J Ecol 95:583–597
Dark P (2000) The environment of Britain in the first millennium AD. Duckworth, London
Davis BAS, Brewer S, Stevenson AC, Guiot J (2003) The temperature of Europe during the Holocene reconstructed from pollen data.Quat Sci Rev 22:1,701-1,716
De Jong R, Björck S, Björkman L, Clemmensen LB (2006) Storminess variation during the last 6500 years as reconstructed from an ombrotrophic peat bog in Halland, southwest Sweden. J Quat Sci 21:905–919
Deza-Araujo M, Morales-Molino C, Tinner W et al (2020) A critical assessment of human-impact indices based on anthropogenic pollen indicators. Quat Sci Rev 236:106291. https://doi.org/10.1016/j.quascirev.2020.106291
Dickson JH, Stewart DA, Thompson R, Turner G, Baxter MS, Drndarsky ND, Rose J (1978) Palynology, palaeomagnetism and radiometric dating of Flandrian marine and freshwater sediments of Loch Lomond. Nature 274:548–553
Dumayne L (1993a) Iron Age and Roman vegetation clearance in northern Britain: further evidence. Bot J Scotl 46:385–392
Dumayne L (1993b) Invader or native? – vegetation clearance in northern Britain during Romano-British time. Veget Hist Archaeobot 2:29–36
Dumayne-Peaty L (1998) Human impact on the environment during the Iron Age and Romano-British times: palynological evidence from three sites near the Antonine Wall, Great Britain. J Archaeol Sci 25:203–214
Dumayne-Peaty L (1999) Continuity or discontinuity? Vegetation change in the Hadrianic-Antonine frontier zone of northern Britain at the end of the Roman occupation. J Biogeogr 26:643–655
Edwards KJ (1988) The hunter-gatherer/agricultural transition and the pollen record in the British Isles. In: Birks HH, Birks HJB, Kaland PE, Moe D (eds) The cultural landscape – past, present and future. Cambridge University Press, Cambridge, pp 255–266
Edwards KJ (2004) People, environmental impacts, and the changing landscapes of Neolithic and early Bronze Age times. In: Shepherd IAG, Barclay GJ (eds) Scotland in ancient Europe: The Neolithic and early Bronze Age of Scotland in their European context. Society of Antiquaries of Scotland, Edinburgh, pp 55–69
Edwards KJ, Whittington G (2003) Vegetation change. In: Edwards KJ, Ralston IBM (eds) Scotland: environment and archaeology, 8000 BC-AD 1000. Wiley, Chichester, pp 63–82
Edwards KJ, Bennett KD, Davies AL (2019) Palaeoecological perspectives on Holocene environmental change in Scotland. Earth Environ Sci Trans R Soc Edinb 110:199–217
Farr DF, Rossman AY (2009) Fungal databases. Systematic Mycology and Microbiology Laboratory, ARS, USDA, https://nt.ars-grin.gov/fungaldatabases/. Accessed 14 July 2021
Farrell M (2015) Later prehistoric vegetation dynamics and Bronze Age agriculture at Hobbister, Orkney, Scotland. Veget Hist Archaeobot 24:467–486
Fossitt JA (1994) Modern pollen rain in the northwest of the British Isles. Holocene 4:365–376
Fyfe RM, Twiddle C, Sugita S et al (2013) The Holocene vegetation cover of Britain and Ireland: overcoming problems of scale and discerning patterns of openness. Quat Sci Rev 73:132–148
Gallego-Sala AV, Charman DJ, Harrison SP, Li G, Prentice IC (2016) Climate-driven expansion of blanket bogs in Britain during the Holocene. Clim Past 12:129–136
Gear AJ, Huntley B (1991) Rapid changes in the range limits of Scots Pine 4000 years ago. Science 251:544–547
Gearey B, Gilbertson D (1997) Pollen taphonomy of trees in a windy climate: Northbay Plantation, Barra, Outer Hebrides. Scott Geogr Mag 113:113–120
Gilbertson DD, Schwenninger J-L, Kemp RA, Rhodes EJ (1999) Sand-drift and soil formation along an exposed North Atlantic coastline: 14,000 years of diverse geomorphological, climatic and human impacts. J Archaeol Sci 26:439–469
Godwin H (1975) History of the British flora, 2nd edn. Cambridge University Press, Cambridge
Goslin J, Gałka M, Sander L, Fruergaard M, Mokenbusch J, Thibault N, Clemmensen LB (2019) Decadal variability of north-eastern Atlantic storminess at the mid-Holocene: new inferences from a record of wind-blown sand, western Denmark. Glob Planet Chang 180:16–32. https://doi.org/10.1016/j.gloplacha.2019.05.010
Grime JP, Hodgson JG, Hunt R (1988) Comparative plant ecology: a functional approach to common British species. Unwin Hyman, London
Grimm EC (1987) CONISS: a FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput Geosci 13:13–35
Harding DW (2004) The Iron Age in northern Britain: Celts and Romans, Natives and Invaders. Routledge, London
Havinga AJ (1984) A 20-year experimental investigation into the differential corrosion susceptibility of pollen and spores in various soil types. Pollen Spores 26:541–558
Harrison J (1997) Central and southern Scotland. In: Wheeler D, Mayes J (eds) Regional climates of the British Isles. Routledge, London, pp 205–227
Hellman S, Bunting MJ, Gaillard M-J (2009) Relevant source area of pollen in patchy cultural landscapes and signals of anthropogenic landscape disturbance in the pollen record: a simulation approach. Rev Palaeobot Palynol 153:245–258. https://doi.org/10.1016/j.revpalbo.2008.08.006
Hjelle KL (1998) Herb pollen representation in surface moss samples from mown meadows and pastures in western Norway. Veget Hist Archaeobot 7:79–96
Huntley B, Daniell JRG, Allen JRM (1997) Scottish vegetation history: the Highlands. Bot J Scotl 49:163–175
Innes JB, Simmons IG (2000) Mid-Holocene charcoal stratigraphy, fire history and palaeoecology at North Gill, North York Moors, UK. Palaeogeogr Palaeoclimatol Palaeoecol 164:151–165
Innes JB, Blackford JJ, Rowley-Conwy PA (2013) Late Mesolithic and early Neolithic forest disturbance: a high-resolution palaeoecological test of human impact hypotheses. Quat Sci Rev 77:80–100
Jacobson GL Jr, Bradshaw RHW (1981) The selection of sites for paleovegetational studies. Quat Res 16:80–96
Jermy AC, James PW, Eddy A (1978) Terrestrial ecosystems. In: Jermy AC, Crabbe JA (eds) The island of Mull: a survey of its flora and environment. British Museum (Natural History), London. pp 10.1-10.75
Jones EW (1959) Biological flora of the British Isles: Quercus L. J Ecol 47:169–222
Karlén W, Larsson L (2007) Mid-Holocene climatic and cultural dynamics in northern Europe. In: Anderson DG, Maasch KA, Sandweiss DH (eds) Climate change and cultural dynamics: a global perspective on Mid-Holocene transitions. Elsevier, Amsterdam, pp 407–434
Keatinge TH, Dickson JH (1979) Mid-Flandrian changes in vegetation on Mainland Orkney. New Phytol 82:585–612
Kirby J (1984) Scottish birchwoods and their conservation — a review. Trans Bot Soc Edinb 44:205–218
Lamb HH (1964) Trees and climatic history in Scotland. Quat J R Meteorol Soc 90:382–394
Langdon PG, Barber KE, Hughes PDM (2003) A 7500-year peat-based palaeoclimatic reconstruction and evidence for an 1100-year cyclicity in bog surface wetness from Temple Hill Moss, Pentland Hills, southeast Scotland. Quat Sci Rev 22:259–274
Langdon PG, Brown AG, Caseldine CJ, Blockley SPE, Stuijts I (2012) Regional climate change from peat stratigraphy for the mid- to late Holocene in central Ireland. Quat Int 268:145–155
Lee JR (1933) The flora of the Clyde area. John Smith, Glasgow
Leuschner HH, Bauerochse A, Metzler A (2007) Environmental change, bog history and human impact around 2900 B.C. in NW Germany – preliminary results from a dendroecological study of a sub-fossil pine woodland at Campemoor, Dümmer Basin. Veget Hist Archaeobot 16:183–195
Leuschner HH, Sass-Klaassen U, Jansma E, Baillie MGL, Spurk M (2002) Subfossil European bog oaks: population dynamics and long-term growth depressions as indicators of changes in the Holocene hydro-regime and climate. Holocene 12:695–706
Lisitsyna OV, Giesecke T, Hicks S (2011) Exploring pollen percentage threshold values as an indication for the regional presence of major European trees. Rev Palaeobot Palynol 166:311–324
Lock G, Ralston I (2017) Atlas of hillforts of Britain and Ireland. https://hillforts.arch.ox.ac.uk. Accessed 7 January 2021
Lowe JJ (1993) Isolating the climatic factors in early- and mid-Holocene palaeobotanical records from Scotland. In: Chambers FM (ed) Climate change and human impact on the landscape. Chapman & Hall, London, pp 67–82
Lozano I, Devoy RJN, May W, Andersen U (2004) Storminess and vulnerability along the Atlantic coastlines of Europe: analysis of storm records and of a greenhouse gases induced climate scenario. Mar Geol 210:205–225
Magny M, Haas JN (2004) A major widespread climate change around 5300 cal. yr BP at the time of the Alpine Iceman. J Quat Sci 19:423–430
Magny M, Leuzinger U, Bortenschlager S, Haas NN (2006) Tripartite climate reversal in central Europe 5600 – 5300 years ago. Quat Res 65:3–19
McDermott F, Mattey DP, Hawkesworth C (2001) Centennial-scale Holocene climate variability revealed by a high-resolution speleothem δ18O record from SW Ireland. Science 294:1,328–331
McKeown MM, Caseldine CJ, Thompson G et al (2019) Complexities in interpreting chironomid-based temperature reconstructions over the Holocene from a lake in Western Ireland. Quat Sci Rev 222:105908. https://doi.org/10.1016/j.quascirev.2019.105908
McVean DN (1953) Biological flora of the British Isles: Alnus glutinosa (L.) Gaertn. J Ecol 41:447–466
McVean DN (1956a) Ecology of Alnus glutinosa (L.) Gaertn. III. Seedling establishment. J Ecol 44:195–218
McVean DN (1956b) Ecology of Alnus glutinosa (L.) Gaertn. V. Notes on some British alder populations. J Ecol 44:321–330
McVean DN (1964) Woodland and scrub. In: Burnett JH (ed) The vegetation of Scotland. Oliver & Boyd, Edinburgh, pp 144–167
McVean DN, Ratcliffe DA (1962) Plant communities of the Scottish Highlands: a study of Scottish mountain, moorland and forest vegetation. HMSO, London
Miles J (1981) Effect of birch on moorlands. Institute of Terrestrial Ecology, Cambridge
Miles J (1985) The pedogenic effects of different species and vegetation types and the implications of succession. J Soil Sci 36:571–584
Miles J (1988) Vegetation and soil change in the uplands. In: Usher MB, Thompson DBA (eds) Ecological change in the uplands. Blackwell, Oxford, pp 57–70
Mitchell FJG (1988) The vegetational history of the Killarney oakwoods, SW Ireland: evidence from fine spatial resolution pollen analysis. J Ecol 76:415–436
Moir AK, Leroy SAG, Brown D, Collins PEF (2010) Dendrochronological evidence for a lower water-table on peatland around 3200 – 3000 BC from subfossil pine in northern Scotland. Holocene 20:931–942
Moore PD, Evans AT, Chater M (1986) Palynological and stratigraphic evidence for hydroseral changes in mires associated with human activity. In: Behre K-E (ed) Anthropogenic indicators in pollen diagrams. Balkema, Rotterdam, pp 209–220
Moore PD, Webb JA, Collinson ME (1991) Pollen analysis, 2nd edn. Blackwell, Oxford
Olsen J, Anderson NJ, Knudsen MF (2012) Variability of the North Atlantic Oscillation over the past 5,200 years. Nat Geosci 5:808–812
Orme LC, Davies SJ, Duller GAT (2015) Reconstructed centennial variability of late Holocene storminess from Cors Fochno, Wales, UK. J Quat Sci 30:478–488
Parry ML (1978) Climate change, agriculture and settlement. Dawson, Folkestone
Paus A (1995) The late Weichselian and early Holocene history of tree birch in south Norway and the Bølling Betula time-lag in northwest Europe. Rev Palaeobot Palynol 85:243–262
Pennington W (1974) The history of British vegetation. English Universities Press, London
Pilcher JR (1973) Pollen analysis and radiocarbon dating of a peat on Slieve Gallion, Co. Tyrone, N. Ireland. New Phytol 72:681–689
Quine CP, White IMS (1994) Using the relationship between rate of tatter and topographical variables to predict site windiness in upland Britain. Forestry 67:245–256
Rackham O (2003) Ancient woodland: its history, vegetation and uses in England. Castlepoint Press, Kirkcudbrightshire
Ramsay S (1995) Woodland clearance in west-central Scotland during the past 3000 years. PhD Dissertation, University of Glasgow, Glasgow
Ramsay S, Dickson JH (1997) Vegetation history of central Scotland. Bot J Scotl 49:141–150
Reimer PJ, Bard E, Bayliss A et al (2013) IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 55:1,869-1,887
Robinson N, Regetz J, Guralnick RP (2014) EarthEnv-DEM90: A nearly-global, void-free, multi-scale smoothed, 90m digital elevation model from fused ASTER and SRTM data. ISPRS J Photogramm Remote Sens 87:57–67. http://www.sciencedirect.com/science/article/pii/S0924271613002360
Rich TCG, Fitzgerald R, Sydes C (1998) Distribution and ecology of Small Cow-Wheat (Melampyrum sylvaticum L. Scrophulariaceae) in the British Isles. Bot J Scot 50:29–46
Rodwell JS (1991) British plant communities, Vol 1: Woodlands and scrub. Cambridge University Press, Cambridge
Rodwell JS (1992) British plant communities. Grasslands and montane communities, vol 3. Cambridge University Press, Cambridge
Roland TP, Caseldine CJ, Charman DJ, Turney CSM, Amesbury MJ (2014) Was there a ‘4.2 ka event’ in Great Britain and Ireland? Evidence from the peatland record. Quat Sci Rev 83:11–27
Roland TP, Daley TJ, Caseldine CJ et al (2015) The 5.2 ka climate event: evidence from stable isotope and multi-proxy palaeoecological peatland records in Ireland. Quat Sci Rev 124:209–223
Sagar GR, Harper JL (1964) Biological flora of the British Isles: Plantago major L., P. media L. and P. lanceolata L. J Ecol 52:189–221
Shaw H, Whyte I (2020) Interpretation of the herbaceous pollen spectra in paleoecological reconstructions: a spatial extension of indices of association and determination of individual pollen source areas from binary data. Rev Palaeobot Palynol 279:104238. https://doi.org/10.1016/j.revpalbo.2020.104238
Skene KR, Sprent JI, Raven JA, Herdman L (2000) Biological flora of the British Isles. Myrica gale L. J Ecol 88:1,079–1,094
Smout TC (1969) A history of the Scottish people 1560–1830. Collins, London
Sommerville AA, Hansom JD, Housley RA, Sanderson DCW (2007) Optically stimulated luminescence (OSL) dating of coastal aeolian sand accumulation in Sanday, Orkney Islands. Scotl Holocene 17:627–637
Stace C (2010) New flora of the British Isles, 3rd edn. Cambridge University Press, Cambridge
Steele RC (1974) Variation in oakwoods in Britain. In: Morris MG, Perring FH (eds) The British oak. Its history and natural history. Botanical Society of the British Isles, Faringdon, pp 130–140
Stewart DA (1979) The Flandrian vegetational history of the Loch Lomond area. PhD thesis, University of Glasgow, Glasgow
Stewart DA, Walker A, Dickson JH (1984) Pollen diagrams from Dubh Lochan, near Loch Lomond. New Phytol 98:531–549
Stewart H, Bradwell T, Bullard J, Davies SJ, Golledge N, McCulloch RD (2017) 8000 years of North Atlantic storminess reconstructed from a Scottish peat record: implications for Holocene atmospheric circulation patterns in Western Europe. J Quat Sci 32:1,075–1,084
Stockmarr J (1971) Tablets with spores used in absolute pollen analysis. Pollen Spores 13:615–621
Swindles GT, Lawson IT, Matthews IP et al (2013) Centennial-scale climate change in Ireland during the Holocene. Earth-Sci Rev 126:300–320
Taylor KJ, McGinley S, Potito AP, Molloy K, Beilman DW (2018) A mid to late Holocene chironomid-inferred temperature record from northwest Ireland. Palaeogeogr Palaeoclimtol Palaeoecol 505:274–286. https://doi.org/10.1016/j.palaeo.2018.06.006
Theuerkauf M, Bos JAA, Jahns S, Janke W, Kuparinen A, Stebich M, Joosten H (2014) Corylus expansion and persistent openness in the early Holocene vegetation of northern central Europe. Quat Sci Rev 90:183–198
Tinsley HM (1975) The former woodland of the Nidderdale Moors (Yorkshire) and the role of early man in its decline. J Ecol 63:1–26
Tinsley HM (2001) Modern pollen deposition in traps on a transect across an anthropogenic tree-line on Exmoor, southwest England: a note summarising the first three years of data. Rev Palaeobot Palynol 117:153–158
Tinsley HM, Smith RT (1974) Surface pollen studies across a woodland/heath transition and their application to the interpretation of pollen diagrams. New Phytol 73:547–565
Tippett R (1974) Natural history of Loch Lomond. University of Glasgow Press, Glasgow
Tip** R (1994) The form and the fate of Scottish woodlands. Proc Soc Antiq Scot 124:1–54
Tip** R (1996) Microscopic charcoal records, inferred human activity and climate change in the Mesolithic of northernmost Scotland. In: Pollard A, Morrison A (eds) The early prehistory of Scotland. Edinburgh University Press, Edinburgh, pp 39–61
Tip** R (1997) Pollen analysis, late Iron Age and Roman agriculture around Hadrian’s Wall. In: Gwilt A, Haselgrove C (eds) Reconstructing Iron Age societies. Oxbow Books, Oxford, pp 239–247
Tip** R (2010) Bowmont: An environmental history of the Bowmont Valley and the Northern Cheviot Hills. Society of Antiquaries of Scotland, Edinburgh
Tip** R, Tisdall E (2006) The landscape context of the Antonine Wall: a review of the literature. Proc Soc Antiq Scot 135:443–470
Tip** R, Bradley R, McCulloch R, Sanders J, Wilson R (2013) Moments of crisis: from coincidence to hypothesis testing in defining the impacts of abrupt climate change in Scottish prehistory. Proc Soc Antiq Scot 142:1–17
Tittensor RM, Steele RC (1971) Plant communities of the Loch Lomond oakwoods. J Ecol 59:561–582
Turner J (1965) A contribution to the history of forest clearance. Proc R Soc Lond B 161:343–354
Turner J (1975) The evidence for land use by prehistoric farming communities: the use of three-dimensional pollen diagrams. In: Evans JG, Limbrey S, Cleere H (eds) The effect of man on the landscape: the highland zone. CBA Research Reports 11. Council of British Archaeology, London, pp 86–95
Turner J (1979) The environment of north east England during Roman times as shown by pollen analysis. J Arch Sci 6:285–290
Turner J (1981) The Iron Age. In: Simmons IG, Tooley MJ (eds) The environment in British prehistory. Duckworth, London, pp 250–281
Turner J (1983) Some pollen evidence for the environment of northern Britain 1000 B.C. to A.D. 1000. In: Chapman JC, Mytum HC (eds) Settlement in North Britain 1000 BC-AD 1000: Papers Presented to George Jobey, Newcastle Upon Tyne, December 1982. BAR British Series 118, Oxford, pp 3–27
Van Asperen EN, Kirby JR, Hunt CO (2016) The effect of preparation methods on dung fungal spores: implications for recognition of megafaunal populations. Rev Palaeobot Palynol 229:1–8
Van Asperen EN, Kirby JR, Shaw HE (2020) Relating dung fungal spore influx rates to animal density in a temperate environment: implications for palaeoecological studies. Holocene 30:218–232
Van Geel B, Aptroot A (2006) Fossil ascomycetes in Quaternary deposits. Nova Hedwigia 82:313–329
Van Geel B, Gelorini V, Lyaruu A et al (2011) Diversity and ecology of tropical African fungal spores from a 25,000-year palaeoenvironmental record in southeastern Kenya. Rev Palaeobot Palynol 164:174–190
Walker MJC, Berkelhammer M, Björck S et al (2012) Formal subdivision of the Holocene Series/Epoch: a discussion paper by a working group of INTIMATE (Integration of ice-core, marine and terrestrial records) and the Subcommission on Quaternary Stratigraphy (International Commission on Stratigraphy). J Quat Sci 27:649–659
Wardle DA, Walker LR, Bardgett RD (2004) Ecosystem properties and forest decline in contrasting long-term chronosequences. Science 305:509–513
Whittow JB (1977) Geology and scenery in Scotland. Penguin Books, London
Wilson P, McGourty J, Bateman MD (2004) Mid- to late-Holocene coastal dune event stratigraphy for the north coast of Ireland. Holocene 14:406–416
Wilson P, Orford JD, Knight J, Braley SM, Wintle AG (2001) Late Holocene (post-4000 years BP) coastal dune development in Northumberland, northeast England. Holocene 11:215–229
Wylie DD, Dickson JH (1998) The Holocene history of Scots pine Pinus sylvestris L. at Loch Sloy, Scottish Highlands. Glasg Nat 23:16–23
Acknowledgements
We thank Alasdair Eckersall for the invitation to work on the NTS Ben Lomond Estate and for assistance with fieldwork, NTS Foundation USA for funding, Althea Davies (St. Andrews University) for assistance with pollen identifications and Charlie Perfect for cartographic help. Michael O’Connell and two referees are thanked for their comments on drafts.
Funding
National Trust for Scotland (NTS) Foundation USA.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Fieldwork and project management were undertaken by Eileen Tisdall and Richard Tip**, Rebecca Barclay was the pollen analyst, Carla Ferreira recorded fungal spores and Emma Ballantyne undertook peat humification analyses. The first draft of the manuscript was written by Richard Tip** and all authors commented on all versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflicts of interest/competing interests:
none.
Additional information
Communicated by M. O’Connell.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Barclay, R., Ferreira, C., Ballantyne, E. et al. Possible climatically driven, later prehistoric woodland decline on Ben Lomond, central Scotland. Veget Hist Archaeobot 32, 1–15 (2023). https://doi.org/10.1007/s00334-022-00871-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-022-00871-4