
Abstract
The novel human coronavirus, Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2), which results in the coronavirus disease 2019 (COVID-19), has caused a serious threat to global public health. Therefore, many studies are performed on the causes and prevalence of this disease and the possible co-occurrence of the infection with other viral and bacterial pathogens is investigated. Respiratory infections predispose patients to co‐infections and these lead to increased disease severity and mortality. Numerous types of antibiotics have been employed for the prevention and treatment of bacterial co‐infection and secondary bacterial infections in patients with a SARS-CoV-2 infection. Although antibiotics do not directly affect SARS‐CoV‐2, viral respiratory infections often result in bacterial pneumonia. It is possible that some patients die from bacterial co‐infection rather than virus itself. Therefore, bacterial co‐infection and secondary bacterial infection are considered critical risk factors for the severity and mortality rates of COVID‐19. In this review, we will summarize the bacterial co‐infection and secondary bacterial infection in some featured respiratory viral infections, especially COVID‐19.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Special Features of Coronavirus
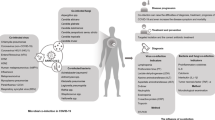
The pandemic of severe acute respiratory syndrome coronavirus 2 raised the attention toward bacterial co-infection and its role in coronavirus disease 2019 (COVID-19) disease. The coronavirus belongs to the Coronaviridae virus family, infecting birds and mammals through respiratory infections [1]. This virus can cause common cold to more severe illnesses, such as SARS, MERS, and COVID-19 [2, 3]. The most prominent clinical symptoms of this disease are dry cough, shortness of breath, fever, and fluid accumulation in the interstitial space of the lungs and chest cavity [4, 5]. Respiratory viruses, such as coronaviruses and influenza viruses, can cause acute damage to lung epithelial cells, allowing other pathogens to infiltrate the affected area. [6, 7]. The history of COVID-19 infection is as follows: the first cases were reported in December 2019, from December 18 to 29, 2019, five patients were admitted with acute respiratory symptoms, one of whom died [2]. On January 2, 2020, a total of 41 patients with infectious symptoms were admitted to Wuhan Hospital in China and confirmed positive for COVID-19 by molecular tests [2]. Diabetes, hypertension, cardiovascular disease, cancer, lung disease, and other medical conditions were present in these patients [2]. It was assumed that these patients probably acquired nosocomial infections, which might be a reason for the severity of their disease [2]. Fever, cough, muscle, or body aches are common symptoms of coronavirus disease, while other symptoms include sputum, headache, bleeding, diarrhea, indigestion, and lymphopenia (decreased lymphocyte count) [8]. Lung ultrasound (LUS) is proven to be a valuable tool to detect specific findings in COVID-19 patients such as pneumonia [8]. Shortness of breath and muscle pain have also been reported in cases of systematic and severe implications [9, 10]. Abnormal features such as acute respiratory syndrome, acute heart failure, and fibrotic pulmonary lesions that lead to death have also been observed in these patients [62, 71]. In addition to the 6D3, 4A8 anti SARS-CoV-2 spike (NTD) antibody can also be attached to similar sections, blocking the S1/S2 Section [62]. The proximity of the viral SAg-like site to the S1/S2 proteolytic cleavage site (peptide bond 685R-S686) suggests that an anti-SEB mAb that cross-react with SARS-CoV-2 may have the potential to block the incision area of the virus. It also modulates cytokine storms and subsequent pro-inflammatory reactions [72]. Three SEB-related mAbs called 14G8, 6D3, and 20B1 have been proposed as the effective SAg toxin B activity inhibitors in animal models. These antibodies bind to different parts of the SEB. Of these antibodies, only 6D3 is structurally and persistently similar to the SARS-CoV-2 S SAg-like motif [73]. Computational analysis studies by Cheng et al. indicate that 6D3 antibody has a stronger tendency to bind to the SAg polybasic site than TMPRSS2 and furin [62, 71]. Based on the results of immunological studies on SARS-CoV-2, it has been shown that the 6D3 antibody can prevent COVID-19 viral infection (Fig. 4).

Sequence alignment of SARS-CoV-2 and multiple SARS-related strains near the insertion of PRRA. Sequence alignment of SARS-CoV-2 is near the S1/S2 cleavage site against multiple bat and pangolin SARS-related strains. Note that the polybasic insert PRRA of SARS-CoV-2 S is not found in closely related SARS-like CoVs, but exists in MERS and HCoVs HKU1 and OC43 (Red fonts). The furin-like cleavage site is indicated by the blue-shaded box (Color figure online)
Conclusion
Distinct differences in microbial composition are observed between patients infected with COVID-19 having low or severe virulence in their respiratory and gastrointestinal systems. One reason for these differences is the use of antibiotics and, on the other hand, the development of antibiotic resistance by some bacterial species. Bacterial populations of BCC and S. epidermidis and Mycoplasma spp. have been identified as opportunistic respiratory pathogens in patients with acute COVID-19 and patients with long-term admission to ICU intensive care unit. These bacteria have even been mentioned as significant cause of death in these patients. On the other hand, the genera Veillonella, Neisseria, Streptococcus, and Prevotella in a mild form are the dominant bacteria in the respiratory tract of patients [6]. The results of various studies show that the exposure to COVID-19 reduces microbial diversity but increases the pathogen population [14]. Therefore, the abundant respiratory pathogens are the bacteria including A. baumannii and S. pneumonia bacteria and fungi, such as C. albicans and C. tropicalis. The presence of C. albicans together with pathogenic bacteria in a synergistic interaction helps the fungal species readily absorb iron and contribute to further pathogenesis [14]. As a result, the presence of nosocomial opportunistic respiratory pathogens during the colonization process in the host body expresses and secretes a wide range of virulence factors, leading to the secretion of cytokines and the development of an inflammatory response [14]. In the early stages of SARS-CoV-2 infection, mainly type 2 pneumocytes become infected in the lungs, causing pneumonia, which can progress to ARDS and increase the risk of secondary bacterial infections with decreased lung immune response. The virus can enter the bloodstream and cause viremia, affecting various organs in the body, including the intestines, with high ACE2 expression. The SARS-CoV-2 infection causes an inflammatory response in the gastrointestinal tract and changes in the intestinal microenvironment. These changes include epithelial hyperpermeability, localized immunosuppression, and microbiome dysbiosis. These disorders allow pathogenic bacteria to enter the bloodstream from the intestinal lumen. Therefore, the sources of endotoxin isolated from COVID-19 patients are known to be the pathogenic bacteria in the intestinal tract causing secondary bacterial infections in the respiratory system. Immunological studies in patients with COVID-19 have shown that 6D3 antibody can block the S protein protease cleavage site of SARS-CoV-2 and prevent virus entry. On the other hand, virus S protein is structurally similar to SAg-like SEB. Therefore, the 6D3 antibody has the dual potential to block SARS-CoV-2 infection as well as SAg associated with T-cell activity and pro-inflammatory responses.
References
Abolghasemi H, Bashash D, Jafari R, Naseri P, Farzanehpour M, Bolandian M et al (2021) A comparative study of laboratory findings in PCR-positive and PCR-negative COVID-19 hospitalized patients. Irish J Med Sci 191:1–8
Rothan HA, Byrareddy SN (2020) The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J Autoimmun 109:102433
Akbariqomi M, Hosseini MS, Rashidiani J, Sedighian H, Biganeh H, Heidari R et al (2020) Clinical characteristics and outcome of hospitalized COVID-19 patients with diabetes: a single-center, retrospective study in Iran. Diabetes Res Clin Pract 169:108467
Sohrabi C, Alsafi Z, O’Neill N, Khan M, Kerwan A, Al-Jabir A et al (2020) World Health Organization declares global emergency: a review of the 2019 novel coronavirus (COVID-19). Int J Surg (London, England) 76:71–76
Fallah A, Sedighian H, Behzadi E, Havaei SA, Kachuei R, Fooladi AAI (2022) The role of serum circulating microbial toxins in severity and cytokine storm of COVID positive patients. Microb Pathog 174:105888
Zhong H, Wang Y, Shi Z, Zhang L, Ren H, He W et al (2021) Characterization of respiratory microbial dysbiosis in hospitalized COVID-19 patients. Cell Discov 7(1):23
de Wit E, van Doremalen N, Falzarano D, Munster VJ (2016) SARS and MERS: recent insights into emerging coronaviruses. Nat Rev Microbiol 14(8):523–534
Liu F, Xu A, Zhang Y, Xuan W, Yan T, Pan K et al (2020) Patients of COVID-19 may benefit from sustained lopinavir-combined regimen and the increase of eosinophil may predict the outcome of COVID-19 progression. Int J Infect Dis 95:183–191
Hedayati MA (2020) Ambient temperature interferes with COVID-19. Int J Prev Med 1:109
Hedayati MA (2021) Viral respiratory coinfections enhance the spread of SARS-CoV-2. Infect Dis Clin Pract (Baltimore, Md) 29(6):e488
Xu J, Zhao S, Teng T, Abdalla AE, Zhu W, **e L et al (2020) Systematic comparison of two animal-to-human transmitted human coronaviruses: SARS-CoV-2 and SARS-CoV. Viruses 12(2):244
Lei J, Li J, Li X, Qi XJR (2020) CT imaging of the 2019 novel coronavirus (2019-nCoV) pneumonia. Radiology 295(1):18
Sirivongrangson P, Kulvichit W, Payungporn S, Pisitkun T, Chindamporn A, Peerapornratana S et al (2020) Endotoxemia and circulating bacteriome in severe COVID-19 patients. Intensiv Care Med Exp 8(1):1–15
de Carvalho FM, Lemos LN, Ciapina LP, Moreira RG, Gerber A, Guimarães APC et al (2020) Prevalence of bacterial pathogens and potential role in COVID-19 severity in patients admitted to intensive care units in Brazil. Medrxiv. https://doi.org/10.1101/2020.12.22.20248501
Peiris JSM, Chu C-M, Cheng VC-C, Chan K, Hung I, Poon LL et al (2003) Clinical progression and viral load in a community outbreak of coronavirus-associated SARS pneumonia: a prospective study. Lancet 361(9371):1767–1772
Morris DE, Cleary DW, Clarke SCJ (2017) Secondary bacterial infections associated with influenza pandemics. Front Microbiol 8:1041
Meng L, Qiu H, Wan L, Ai Y, Xue Z, Guo Q et al (2020) Intubation and ventilation amid the COVID-19 outbreak: Wuhan’s experience. Anesthesiology 132(6):1317–1332
Zhou F, Yu T, Du R, Fan G, Liu Y, Liu Z et al (2020) Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. Lancet (London, England) 395(10229):1054–1062
Dickson RP, Martinez FJ, Huffnagle GB (2014) The role of the microbiome in exacerbations of chronic lung diseases. Lancet (London, England) 384(9944):691–702
Kim D, Quinn J, Pinsky B, Shah NH, Brown IJJ (2020) Rates of co-infection between SARS-CoV-2 and other respiratory pathogens. JAMA 323(20):2085–2086
Rawson TM, Moore LS, Zhu N, Ranganathan N, Skolimowska K, Gilchrist M et al (2020) Bacterial and fungal coinfection in individuals with coronavirus: a rapid review to support COVID-19 antimicrobial prescribing. Clin Infect Dis 71(9):2459–2468
Lee JY, Monk IR, da Silva AG, Seemann T, Chua KY, Kearns A et al (2018) Global spread of three multidrug-resistant lineages of Staphylococcus epidermidis. Nat Microbiol 3(10):1175–1185
Miragaia M, Couto I, Pereira SF, Kristinsson KG, Westh H, Jarløv JO et al (2002) Molecular characterization of methicillin-resistant Staphylococcus epidermidis clones: evidence of geographic dissemination. J Clin Microbiol 40(2):430–438
Yildiz H, Sünnetçioğlu A, Ekin S, Baran Aİ, Özgökçe M, Aşker S et al (2019) Delftia acidovorans pneumonia with lung cavities formation. Colomb Med (Cali) 50(3):215–221
Patel D, Iqbal AM, Mubarik A, Vassa N, Godil R, Saad M et al (2019) Delftia acidovorans: a rare cause of septic pulmonary embolism from catheter-related infection: case report and literature review. Med Case Rep 27:100835
Man WH, de Steenhuijsen Piters WA, Bogaert DJNRM (2017) The microbiota of the respiratory tract: gatekeeper to respiratory health. Nat Rev Mocrobiol 15(5):259–270
Huffnagle G, Dickson R, Lukacs NW (2017) The respiratory tract microbiome and lung inflammation: a two-way street. Mucosal Immunol 10(2):299–306
Mishra D, Satpathy G, Wig N, Fazal F, Ahmed NH, Panda SKJJoi, et al (2020) Evaluation of 16S rRNA broad range PCR assay for microbial detection in serum specimens in sepsis patients. J Infect Public Health 13(7):998–1002
Mehta P, McAuley DF, Brown M, Sanchez E, Tattersall RS, Manson JJ (2020) COVID-19: consider cytokine storm syndromes and immunosuppression. Lancet 395(10229):1033–1034
Harding CM, Hennon SW, Feldman MF (2018) Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat Rev Microbiol 16(2):91–102
Kadioglu A, Weiser JN, Paton JC, Andrew PWJNRM (2008) The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat Rev Microbiol 6(4):288–301
Bien J, Sokolova O, Bozko P (2011) Characterization of virulence factors of Staphylococcus aureus: novel function of known virulence factors that are implicated in activation of airway epithelial proinflammatory response. J Pathog 2011:601905
Adair CG, Gorman SP, Feron BM, Byers LM, Jones DS, Goldsmith CE et al (1999) Implications of endotracheal tube biofilm for ventilator-associated pneumonia. Intensive Care Med 25(10):1072–1076
Wolcott R, Costerton JW, Raoult D, Cutler SJ (2013) The polymicrobial nature of biofilm infection. Clin Microbiol Infect 19(2):107–112
Hanke ML, Kielian T (2012) Deciphering mechanisms of staphylococcal biofilm evasion of host immunity. Front Cell Infect Microbiol 2:62
Hirschfeld J (2014) Dynamic interactions of neutrophils and biofilms. J Oral Microbiol 6:26102
Doran KS, Chang JC, Benoit VM, Eckmann L, Nizet V (2002) Group B streptococcal beta-hemolysin/cytolysin promotes invasion of human lung epithelial cells and the release of interleukin-8. J Infect Dis 185(2):196–203
Deorukhkar SC, Saini S, Mathew S (2014) Virulence factors contributing to pathogenicity of Candida tropicalis and Its antifungal susceptibility profile. Int J Microbiol 2014:456878
Hurley BP, McCormick BA (2008) Multiple roles of phospholipase A2 during lung infection and inflammation. Infect Immun 76(6):2259–2272
Smith AW, Roche H, Trombe MC, Briles DE, Håkansson A (2002) Characterization of the dihydrolipoamide dehydrogenase from Streptococcus pneumoniae and its role in pneumococcal infection. Mol Microbiol 44(2):431–448
Datta V, Myskowski SM, Kwinn LA, Chiem DN, Varki N, Kansal RG et al (2005) Mutational analysis of the group A streptococcal operon encoding streptolysin S and its virulence role in invasive infection. Mol Microbiol 56(3):681–695
Ghoneim HE, Thomas PG, McCullers JA (2013) Depletion of alveolar macrophages during influenza infection facilitates bacterial superinfections. J Immunol (Baltimore) 191(3):1250–1259
Sodhi CP, Nguyen J, Yamaguchi Y, Werts AD, Lu P, Ladd MR et al (2019) A dynamic variation of pulmonary ACE2 is required to modulate neutrophilic inflammation in response to Pseudomonas aeruginosa lung infection in mice. J Immunol 203(11):3000–3012
Aggarwal S, Ahmad I, Lam A, Carlisle MA, Li C, Wells JM et al (2018) Heme scavenging reduces pulmonary endoplasmic reticulum stress, fibrosis, and emphysema. JCI Insight. https://doi.org/10.1172/jci.insight.120694
Raval M, C, J Lee PJ, (2010) Heme oxygenase-1 in lung disease. Curr Drug Target 11(12):1532–1540
Su WL, Lin CP, Hang HC, Wu PS, Cheng CF, Chao YC (2021) Desaturation and heme elevation during COVID-19 infection: a potential prognostic factor of heme oxygenase-1. J Microbiol Immunol Infect 54(1):113–116
Wagener F, Pickkers P, Peterson SJ, Immenschuh S, Abraham NG (2020) Targeting the heme-heme oxygenase system to prevent severe complications following COVID-19 infections. Antioxidants (Basel, Switzerland) 9(6):540
Petruk G, Puthia M, Petrlova J, Samsudin F, Strömdahl A-C, Cerps S et al (2020) SARS-CoV-2 Spike protein binds to bacterial lipopolysaccharide and boosts proinflammatory activity. J Mol Cell Biol 12(12):916–932
Wan Y, Shang J, Graham R, Baric RS, Li F (2020) Receptor recognition by the novel coronavirus from Wuhan: an analysis based on decade-long structural studies of SARS coronavirus. J Virol. https://doi.org/10.1128/JVI.00127-20
Rittirsch D, Flierl MA, Ward PA (2008) Harmful molecular mechanisms in sepsis. Nat Rev 8(10):776–787
Liu X, Yin S, Chen Y, Wu Y, Zheng W, Dong H et al (2018) LPS-induced proinflammatory cytokine expression in human airway epithelial cells and macrophages via NF-κB, STAT3 or AP-1 activation. Mol Med Rep 17(4):5484–5491
Puthia M, Butrym M, Petrlova J, Strömdahl A-C, Andersson MÅ, Kjellström S et al (2020) A dual-action peptide-containing hydrogel targets wound infection and inflammation. Sci Transl Med. https://doi.org/10.1126/scitranslmed.aax6601
Luo S, Zhang X, Xu HJCG (2020) Don’t overlook digestive symptoms in patients with 2019 novel coronavirus disease (COVID-19). Clin Gastroenterol Hepatol 18(7):1636
Opal S (2007) The host response to endotoxin, antilipopolysaccharide strategies, and the management of severe sepsis. Int J Med Microbiol 297(5):365–377
Leelahavanichkul A, Worasilchai N, Wannalerdsakun S, Jutivorakool K, Somparn P, Issara-Amphorn J et al (2016) Gastrointestinal leakage detected by serum (1→ 3)-β-D-glucan in mouse models and a pilot study in patients with sepsis. Shock 46(5):506–518
Netea MG, Joosten LA, Van Der Meer JW, Kullberg B-J, Van De Veerdonk FL (2015) Immune defence against Candida fungal infections. Nat Rev Immun 15(10):630–642
Koch R, Diavatopoulos D, Ferwerda G, Pickkers P, de Jonge M, Kox M (2018) The endotoxin-induced pulmonary inflammatory response is enhanced during the acute phase of influenza infection. Intensiv Care Med Exp 6(1):1–10
Huang C, Wang Y, Li X, Ren L, Zhao J, Hu Y et al (2020) Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 395(10223):497–506
** X, Lian J-S, Hu J-H, Gao J, Zheng L, Zhang Y-M et al (2020) Epidemiological, clinical and virological characteristics of 74 cases of coronavirus-infected disease 2019 (COVID-19) with gastrointestinal symptoms. Gut 69(6):1002–1009
Lin L, Jiang X, Zhang Z, Huang S, Zhang Z, Fang Z et al (2020) Gastrointestinal symptoms of 95 cases with SARS-CoV-2 infection. Gut 69(6):997–1001
Wang Q, Boshoff HI, Harrison JR, Ray PC, Green SR, Wyatt PG et al (2020) PE/PPE proteins mediate nutrient transport across the outer membrane of Mycobacterium tuberculosis. Science 367(6482):1147–1151
Cheng MH, Zhang S, Porritt RA, Noval Rivas M, Paschold L, Willscher E et al (2020) Superantigenic character of an insert unique to SARS-CoV-2 spike supported by skewed TCR repertoire in patients with hyperinflammation. Proc Natl Acad Sci 117(41):25254
Rivas MN, Porritt RA, Cheng MH, Bahar I, Arditi M (2021) COVID-19–associated multisystem inflammatory syndrome in children (MIS-C): a novel disease that mimics toxic shock syndrome—the superantigen hypothesis. J Allergy Clin immunol 147(1):57–59
Benton DJ, Wrobel AG, Xu P, Roustan C, Martin SR, Rosenthal PB et al (2020) Receptor binding and priming of the spike protein of SARS-CoV-2 for membrane fusion. Nature 588(7837):327–330
Coutard B, Valle C, de Lamballerie X, Canard B, Seidah NG, Decroly E (2020) The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antivir Res 176:104742
Cui J, Li F, Shi ZL (2019) Origin and evolution of pathogenic coronaviruses. Nat Rev Microbiol 17(3):181–192
Yan R, Zhang Y, Li Y, **a L, Guo Y, Zhou Q (2020) Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science (New York, NY) 367(6485):1444–1448
Matsuyama S, Nao N, Shirato K, Kawase M, Saito S, Takayama I et al (2020) Enhanced isolation of SARS-CoV-2 by TMPRSS2-expressing cells. Proc Natl Acad Sci 117(13):7001–7003
Walls AC, Park YJ, Tortorici MA, Wall A, McGuire AT, Veesler D (2020) Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 181(2):281–92.e6
Shang J, Wan Y, Luo C, Ye G, Geng Q, Auerbach A et al (2020) Cell entry mechanisms of SARS-CoV-2. Proc Natl Acad Sci 117(21):11727–11734
Min L, Sun QJ (2021) Antibodies and vaccines target RBD of SARS-CoV-2. Front Mol Biosci 8:247
Krakauer TJT (2019) Staphylococcal superantigens: pyrogenic toxins induce toxic shock. Toxin 11(3):178
Dutta K, Varshney AK, Franklin MC, Goger M, Wang X, Fries BC (2015) Mechanisms mediating enhanced neutralization efficacy of staphylococcal enterotoxin B by combinations of monoclonal antibodies. J Biol Chem 290(11):6715–6730
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or non-profit sectors.
Author information
Authors and Affiliations
Contributions
PF, HS, and AI wrote the manuscript with support from all authors. PF draw the figures. EB, HS, and RK revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Fazel, P., Sedighian, H., Behzadi, E. et al. Interaction Between SARS-CoV-2 and Pathogenic Bacteria. Curr Microbiol 80, 223 (2023). https://doi.org/10.1007/s00284-023-03315-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00284-023-03315-y