
Abstract
Background
Resveratrol is a natural polyphenol found in berries, roots and wine that is well known to have anti-inflammatory and anti-oxidative properties. The anti-inflammatory effect has been reported for both immune cells and connective tissues, but only few studies have investigated effects on immune mediated inflammatory arthritis. None of which have studied this effect when combining resveratrol with methotrexate or adalimumab, two major drugs in the treatment of immune mediated inflammatory arthritis.
We therefore aimed to investigate the anti-inflammatory effect of resveratrol alone and in combination with methotrexate or adalimumab in ex vivo models of immune mediated inflammatory arthritis. We furthermore aimed to describe any variations in this effect based on disease activity and cellular composition of the synovial fluid infiltrate.
Methods
Synovial fluid mononuclear cells from patients with rheumatoid arthritis (n = 7) and spondyloarthritis (n = 7) were cultured for either 48 h or 21 days. In both models, synovial fluid mononuclear cells were treated with resveratrol alone or in combination with methotrexate or adalimumab. Monocyte chemoattractant protein 1, matrix metalloproteinase 3 and tartrate resistant acidic phosphatase were measured to quantify inflammation, enzymatic degradation and osteoclast differentiation, respectively.
Results
Resveratrol reduced monocyte chemoattractant protein 1 production by synovial fluid mononuclear cells significantly (p = 0.005) compared to untreated controls. The effect of resveratrol was greatest in cultures from patients with low disease activity, i.e. DAS28CRP ≤ 3.2 (p = 0.022), and in cultures dominated by lymphocytes (p = 0.03). Further, the combination of methotrexate and resveratrol significantly reduced monocyte chemoattractant protein 1 levels compared with methotrexate alone in cultures from patients with low disease activity (p = 0.016), and in cultures with high lymphocyte count (p = 0.011). Resveratrol did not significantly affect matrix metalloproteinase 3 and tartrate resistant acidic phosphatase production.
Conclusion
Resveratrol has anti-inflammatory properties in our ex vivo model of immune mediated inflammatory arthritis. Results show an additive effect of resveratrol, when combined with methotrexate in samples dominated by lymphocytes and samples from patients with low disease activity. This suggests further investigations in vitro and whether this effect may also be present in a clinical setting.
Similar content being viewed by others
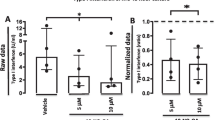


Background
The natural polyphenol resveratrol (RSV) is found in certain berries, grapes and plants [1] were it exerts anti-fungal and anti-oxidative effects [2]. In humans, RSV is also shown to possess anti-inflammatory and anti-oxidative properties [1]. The anti-inflammatory effect has been shown in both immune cells [3,4, 48 h- and 21 days SFMC cultures were assessed under a microscope for viability after the incubation period. Fig. 1 shows images of untreated and RSV treated cultures. Viable cells were present in similar densities for both. No visible cell toxicity was detected in medium with dimethyl sulfoxide compared to medium alone. Furthermore, 21 days SFMC cultures showed presence of multinucleated cells, as shown previously [21]. Enzyme analysis on untreated 21 days SFMC cultures showed TRAP activity in 8 cultures. An additional file shows similar cell viability in 48 h and 21 days SFMC cultures that was untreated or treated with RSV, MTX and adalimumab; Additional file 1. Cell culture viability assessed in microscope (× 10) of 48 h- and 21 days SFMC cultures after incubation period. No immediate cell loss after RSV treatment (25 μM) compared to untreated (UT) cultures. 21 days SFMC cultures showed forming of multinucleated cells The measured concentrations/activity of MCP-1, MMP3 and TRAP in culture supernatants are presented in Fig. 2. RSV significantly decreased the concentration of MCP-1 in 48 h SFMC cultures (Fig. 2a, p = 0.005), and showed no significant difference when comparing MCP-1 ratios in samples from patients with RA and SpA (Fig. 3, p = 0.815). RSV did not change MMP3 concentration in 48 h SFMC cultures or MCP-1 concentration and TRAP activity in 21 days SFMC cultures (Fig. 2b-d). Concentrations of MCP-1 (a, n = 14 and b, n = 8), MMP3 (c, n = 9) and TRAP activity (d, n = 8) in untreated (UT) and resveratrol (RSV) treated cultures measured by ELISA and enzyme activity assay in their respective supernatants. UT is presented as a negative control to its in-patient corresponding RSV treated culture, * p < 0.05. RSV concentration was 25 μM Ratio of MCP-1 in resveratrol (RSV) treated cultures (n = 14) divided in groups of patients with rheumatoid arthritis (RA) and spondyloarthritis (SpA). MCP-1 was measured by ELISA. RSV concentration was 25 μM The effect of RSV on MCP-1 concentration in 48 h SFMC cultures was assessed in the subgroup analyses as described in the methods section. In short, subgroup changes in MCP-1 concentrations were compared based on DAS28CRP and the lymphocyte/monocyte count in the SFMC cultures (Fig. 4). Ratio of MCP-1 in resveratrol (RSV) treated cultures (n = 11) plotted against DAS28CRP (a), and the percentage of monocytes (b) and lymphocytes (c) in synovial fluid of the donor. d-f depicts group comparisons as median and interquartiles. *p < 0.05 vs. negative control. MCP-1 was measured by ELISA. RSV concentration was 25 μM In the subgroup analysis, RSV induced a significant MCP-1 decrease only in cultures from patients with DAS28CRP ≤ 3.2, p = 0.028, low monocytes count (≤ 27%, p = 0.019), and high lymphocyte count (> 63%, p = 0.002). The median MCP-1 ratio was significantly decreased in patients with DAS28CRP ≤ 3.2 vs. > 3.2 (p = 0.022) and when lymphocyte count was > 63% compared with patients with ≤ 63% (p = 0.03). In 48 h SFMC cultures similar visual tendency was present for MMP3 ratios but no statistical significant correlation. Thus, the effect of RSV was most potent in cultures from patients with low DAS28CRP and a high percentage of lymphocytes in the synovial fluid. However, the limited number of MMP3 results made subgroup analysis inappropriate. The 21 days SFMC cultures showed no statistically significant correlation of MCP-1 or TRAP ratios, when plotted against DAS28CRP, monocyte count or lymphocyte count. Subgroup analysis was again inappropriate due to the small number of data. To assess effects of different treatments, MCP-1 ratios (treated/untreated) from 48 h and 21 days SFMC culture were calculated. These ratios from RSV, MTX, MTX + RSV, adalimumab and adalimumab+RSV treatment are presented in Fig. 5. The ratios were then compared to identify differences in treatment effects. Ratios of MCP-1 (a, n = 14 and b, n = 8 (RSV and MTX + RSV) or n = 7 (MTX, ADA and ADA + RSV)) and MMP3 (c, n = 9) concentrations, and TRAP activity (d, n = 8) in supernatants from cultures with the noted substance or a combination of resveratrol (RSV), Ebetrex (MTX) and Adalimumab (ADA). Concentrations were: RSV; 25 μM, MTX; 0.5 μg/ml, Adalimumab; 5 μg/ml. MCP-1 and MMP3 were measured by ELISA and TRAP with an enzyme activity assay. Data is presented as a dot for each measurement; box and bars represent median and interquartile range. *p < 0.05 vs. negative control In the 48 h SFMC cultures, MCP-1 ratios were significantly decreased by all the above-mentioned treatment conditions compared to no treatment (Fig. 5a). MMP3 ratios were not decreased significantly by any treatment (Fig. 5c), though RSV treatment resulted in the largest decrease (MMP3 ratio median = 0.77). MTX and adalimumab treatment did not result in a significantly different response compared to RSV in 48 h SFMC cultures and no additive effect of RSV was detected when including all cultures in the comparative analysis. In 21 days SFMC cultures, MTX, MTX + RSV and adalimumab+RSV decreased median MCP-1 ratios, but only MTX + RSV produced a significant decrease. This was both when compared to the negative control (p = 0.008) and RSV alone (p = 0.0025). Adalimumab treatment produced a significant decrease in TRAP activity with and without RSV compared to no treatment (p = 0.023 and p = 0.008, respectively). No other treatment affected TRAP activity. In subgroup analysis of the 48 h SFMC cultures, MTX significantly decreased MCP-1 ratios in patients with high lymphocyte count (> 63%). RSV + MTX significantly lowered MCP-1 in patients with DAS28CRP ≤ 3.2, DAS28CRP > 3.2, low monocyte count (≤ 27%) and high lymphocyte count (> 63%); Fig. 6a-c. MCP-1 ratios as median and interquartiles in cultures treated with ebetrex (MTX) alone or in combination with resveratrol (RSV). Group comparisons were based on a) Disease Activity Score 28 (DAS28CRP), b) the percentage of monocytes and c) the percentage of lymphocytes in synovial fluid of the donor. *p < 0.05 vs. negative control. MCP-1 was measured by ELISA. Concentrations were: RSV = 25 μM and MTX = 0.5 μg/ml Moreover, the addition of RSV to MTX treatment produced a significant lower MCP-1 ratio than MTX alone in the subgroup with DAS28CRP ≤ 3.2, p = 0.016, DASCRP > 3.2, p = 0.036, low monocyte count (≤ 27%, p = 0.04) and in cultures with high lymphocyte count (> 63%, p = 0.011). Figure 7a-c presents subgroup MCP-1 ratio results (48 h SFMC cultures) for treatment with adalimumab alone or in combination with RSV. Both treatments produced significantly lower MCP-1 ratios in several subgroups (Fig. 7a-c) compared to no treatment. The greatest decrease in MCP-1 ratio was produced by adalimumab in combination with RSV in subgroups with DAS28CRP ≤ 3,2, low monocyte count (≤ 27%) and high lymphocyte count (> 63%). MCP-1 ratios as median and interquartiles in cultures treated with adalimumab (ADA) alone or in combination with resveratrol (RSV). Group comparisons were based on a) Disease Activity Score 28 (DAS28CRP), b) the percentage of monocytes and c) the percentage of lymphocytes in synovial fluid of the donor. *p < 0.05 vs. negative control. MCP-1 was measured by ELISA. Concentrations were: RSV = 25 μM and ADA = 5 μg/ml None of these ratios were, however, significantly different compared with adalimumab treatment alone. Subgroup analysis was not carried out for MMP3 ratios in 48 h SFMC cultures and for MCP-1 and TRAP ratios from 21 days SMFC cultures due to the smaller number of cell cultures. A combination of two in vitro arthritis models was used to investigate the effects of RSV. RSV inhibited MCP-1 production in the 48 h inflammation model to a similar extent as MTX and adalimumab. This well-known anti-inflammatory property of RSV in vitro is in accord with previous reports [29, 34, 35]. Our results are to the author’s knowledge, however, the first to suggest that RSV’s anti-inflammatory effect may be influenced by the level of disease activity in patients with arthritis and composition of immune cells present in the cell cultures. Thus, RSV seemed to exert the greatest relative inhibition of MCP-1 in cultures from patients with low disease activity i.e. a DAS28CRP ≤ 3.2 or a synovial fluid infiltration dominated by lymphocytes. RSV also resulted in a decrease of MMP3 ratio medians in 48 h SFMC cultures. However, this change was not significant as otherwise reported previously in fibroblast-like synoviocyte cultures from RA patients [8, 9]. Several parameters may offer an explanation to this. 1) The cellular composition of our models, i.e. SFMC may differ significantly from a fibroblast-like synoviocyte mono culture. 2) The mentioned studies only showed significant inhibition when cultures were additionally stimulated with TNF-α [8] or IL-1β [9], whereas our cultures received no additional stimulation in vitro. 3) The concentration of RSV may influence the effective inhibition of MMP3; Glehr et al. [9] used 100 μM versus our 25 μM, and Tian et al. [8] reported significant inhibition at 12.5, 25 and 50 μM. Despite differences, the two previous studies together with our data suggest an inhibitory effect of RSV on MMP3 production, and thereby a possible cartilage protective effect in relation to inflammatory arthritis. RSV had no measurable effect on TRAP activity. This result is somewhat in conflict with previous reports were RSV is shown to diminish TRAP production in canine osteoclasts [36], and inhibit canine and murine osteoclastogenesis through various mechanisms [36, 37]. In contrast, our in vitro model received no additional stimulation, which may have limited our models’ ability to detect subtle changes in TRAP activity. Osteoclasts are, however, not the only cell line of interest when evaluating RSV effects on arthritis. RSV has also been shown to attenuate pro-inflammatory processes in chondrocytes [30] and stimulates osteoblast differentiation [38]. Results that may give explanation for chondro-protective effects seen in osteoarthritic rabbits treated with intraarticular injections of RSV [39, 40]. Subgroup analysis of 48 h SFMC cultures showed RSV anti-inflammatory effect was most pronounced in cultures from patients with low disease activity and in patients with a synovial fluid dominated by lymphocytes. Our study was not designed to explain these differences. We therefore suggest further studies that investigate whether greater levels of systemic inflammation induce a different array of inflammatory pathways in these SFMCs, and whether pathways are affected differently by RSV at different levels of disease activity/inflammatory burden. It would also be interesting to see detailed investigations of the cellular subset, i.e. whether different lymphocyte subtypes are affected differently by RSV. RSV has, to the author’s knowledge not been tested against or together with MTX or adalimumab in vitro or in vivo before. It is of great interest that the addition of RSV to MTX treatment showed significant additive effect especially in subgroups with low disease activity, or when the synovial fluid was dominated by lymphocytes. The effect in patients with DAS28CRP ≤ 3.2, is interesting because patients with low disease activity can still experience pain and discomfort. This encourages in vivo studies of the addition of RSV to MTX in patients with low disease activity. It should be mentioned that we did not have information about DAS28CRP in 4 patients which explains the differences in Fig. 5 (n = 14) and Fig. 6 (n = 10). In addition to this, a previous study has reported that RSV protects against MTX induced oxidative stress in the small intestines of rats [41]. An interesting protective effect, since gastro-intestinal side effects from MTX are reported as a major reason for treatment discontinuation in RA patients [42]. Results from adalimumab treatment in the 48 h SFMC model, showed no significant additive effect of RSV, but did show same tendencies as with MTX. Despite RSVs lipophilic nature and quick absorption from the small intestines, only relatively low amounts are found in the bloodstream after oral ingestion [43]. Studies explain this by a high rate of hepatic metabolism, which conjugates RSV with sulfates and glucuronides [1, 43]. Thus, a big challenge when discussing RSV as a possible drug candidate is drug delivery. Possible solutions could be a specific inhibition of the hepatic RSV metabolism or a delivery matrix that allows passage through the liver without major metabolism. Another possible delivery method is intraarticular injection of RSV. Animal studies have so far shown that intraarticular injected RSV reduce synovial hyperplasia and inflammation in an induced arthritis model [44], and attenuate if not prevent osteoarthritic progress in mice [45] and rabbits [39, 40]. As with all in vitro studies, the models used here have the fundamental limitation of only being a proxy to the in vivo situation, it seeks to investigate. This issue was addressed here by abstaining from additional stimulation of cultures, which in return may have diminished the study’s ability to detect subtle effects of RSV. The fact that this study in general have been limited to show trending effects of RSV and only few significant changes, may also be influenced by the relative small study population and unfortunate missing data from some cultures. This study included a very heterogenous patient population with both RA and SpA patients on different disease modifying anti rheumatic drugs. RA and SpA both affect the synovial joints but also show very different clinical and immunological features. RSV treatment effect was not different when comparing SpA and RA in this study. Treatment with anti-TNFα or other disease modifying anti rheumatic drugs may affect immune cells even after isolation and thereby skew results from cultures. The potential influence of different treatments could not be studied due to the low sample size. The study does, however, also possess strengths in its design and execution. First and foremost, the study combines both synovial fluid cells and osteoclasts involved in the osteoimmunological processes of inflammatory arthritis. The combination of models together with the different treatment conditions could allow interesting arguments across the pathological spectrum of inflammatory arthritis. The design furthermore permits the discovery of possible interactions between RSV, MTX and adalimumab, and enables it to suggest questions for future studies regarding RSVs effect at different levels of disease activity, inflammatory burden and on various cellular subsets. To conclude, this study confirms RSV as an anti-inflammatory agent in our ex vivo model of immune mediated inflammatory arthritis. An effect which our data suggest can work synergistic with MTX; especially in cases of low disease activity making RSV a possible supplemental treatment. As a result, we suggest future investigations into interactions and potential additive effects of RSV in MTX treatment of patients with immune mediated inflammatory arthritis.Results
Viability and differentiation of cells

Resveratrol significantly decreased MCP-1 concentration in 48 h SFMC cultures without differences comparing response in RA and SpA patients


Resveratrol’s effect may be influenced by the level of disease activity and the cellular subset in synovial fluid

Resveratrol, methotrexate and adalimumab show similar MCP-1 lowering effects in 48 h SFMC cultures, but not 21 days SFMC cultures

Potential additive effects of RSV may be influenced by the level of disease activity and cellular subset in synovial fluid


Discussion
Conclusion
Abbreviations
- DAS28CRP:
-
Disease activity score 28
- ELISA:
-
Enzyme-linked immunosorbent assay
- IL-1β:
-
Interleukin 1 beta
- MCP-1:
-
Monocyte chemoattractant protein 1
- MMP3:
-
Matrix metalloproteinase 3
- MTX:
-
Methotrexate
- RA:
-
Rheumatoid arthritis
- RSV:
-
Resveratrol
- SFMC:
-
Synovial fluid mononuclear cells
- SpA:
-
Spondyloarthritis
- TNF-α:
-
Tumor necrosis factor alfa
- TRAP:
-
Tartrate resistant acid phosphatase
References
Gambini J, Ingles M, Olaso G, Lopez-Grueso R, Bonet-Costa V, Gimeno-Mallench L, et al. Properties of resveratrol: in vitro and in vivo studies about metabolism, bioavailability, and biological effects in animal models and humans. Oxidative Med Cell Longev. 2015;2015:837042.
Jeandet P, Douillet-Breuil AC, Bessis R, Debord S, Sbaghi M, Adrian M. Phytoalexins from the Vitaceae: biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. J Agric Food Chem. 2002;50:2731–41.
Vasamsetti SB, Karnewar S, Gopoju R, Gollavilli PN, Narra SR, Kumar JM, et al. Resveratrol attenuates monocyte-to-macrophage differentiation and associated inflammation via modulation of intracellular GSH homeostasis: relevance in atherosclerosis. Free Radic Biol Med. 2016;96:392–405.
Park SY, Lee SW, Kim HY, Lee SY, Lee WS, Hong KW, et al. SIRT1 inhibits differentiation of monocytes to macrophages: amelioration of synovial inflammation in rheumatoid arthritis. J Mol Med (Berl). 2016;94:921–31.
Zou T, Yang Y, **a F, Huang A, Gao X, Fang D, et al. Resveratrol inhibits CD4+ T cell activation by enhancing the expression and activity of Sirt1. PLoS One. 2013;8:e75139.
Gao X, Xu YX, Janakiraman N, Chapman RA, Gautam SC. Immunomodulatory activity of resveratrol: suppression of lymphocyte proliferation, development of cell-mediated cytotoxicity, and cytokine production. Biochem Pharmacol. 2001;62:1299–308.
Siriwardhana N, Kalupahana NS, Cekanova M, LeMieux M, Greer B, Moustaid-Moussa N. Modulation of adipose tissue inflammation by bioactive food compounds. J Nutr Biochem. 2013;24:613–23.
Tian J, J-w C, Gao J-s, Li L, **e X. Resveratrol inhibits TNF-α-induced IL-1β, MMP-3 production in human rheumatoid arthritis fibroblast-like synoviocytes via modulation of PI3kinase/Akt pathway. Rheumatol Int. 2013;33:1829–35.
Glehr M, Breisach M, Walzer S, Lohberger B, Furst F, Friesenbichler J, et al. The influence of resveratrol on the synovial expression of matrix metalloproteinases and receptor activator of NF-kappaB ligand in rheumatoid arthritis fibroblast-like synoviocytes. Z Naturforsch. 2013;68:336–42.
Wuertz KP, Quero LM, Sekiguchi MMDP, Klawitter MM, Nerlich AMD, Konno S-IMD, et al. The red wine polyphenol resveratrol shows promising potential for the treatment of nucleus pulposus-mediated pain in vitro and in vivo. Spine. 2011;36:E1373–84.
Cottart CH, Nivet-Antoine V, Laguillier-Morizot C, Beaudeux JL. Resveratrol bioavailability and toxicity in humans. Mol Nutr Food Res. 2010;54:7–16.
Rudwaleit M, van der Heijde D, Landewe R, Akkoc N, Brandt J, Chou CT, et al. The assessment of SpondyloArthritis International Society classification criteria for peripheral spondyloarthritis and for spondyloarthritis in general. Ann Rheum Dis. 2011;70:25–31.
Aletaha D, Neogi T, Silman AJ, Funovits J, Felson DT, Bingham CO 3rd, et al. 2010 Rheumatoid arthritis classification criteria: an American College of Rheumatology/European league against rheumatism collaborative initiative. Arthritis Rheum. 2010;62:2569–81.
Scott DL, Wolfe F, Huizinga TW. Rheumatoid arthritis. Lancet. 2010;376:1094–108.
Feldmann M, Brennan FM, Maini RN. Role of cytokines in rheumatoid arthritis. Annu Rev Immunol. 1996;14:397–440.
Dawson J, Miltz W, Mir AK, Wiessner C. Targeting monocyte chemoattractant protein-1 signalling in disease. Expert Opin Ther Targets. 2003;7:35–48.
Ellingsen T, Buus A, Stengaard-Pedersen K. Plasma monocyte chemoattractant protein 1 is a marker for joint inflammation in rheumatoid arthritis. J Rheumatol. 2001;28:41–6.
Johnson LL, Dyer R, Hupe DJ. Matrix metalloproteinases. Curr Opin Chem Biol. 1998;2:466–71.
Maksymowych WP, Landewe R, Conner-Spady B, Dougados M, Mielants H, van der Tempel H, et al. Serum matrix metalloproteinase 3 is an independent predictor of structural damage progression in patients with ankylosing spondylitis. Arthritis Rheum. 2007;56:1846–53.
Houseman M, Potter C, Marshall N, Lakey R, Cawston T, Griffiths I, et al. Baseline serum MMP-3 levels in patients with rheumatoid arthritis are still independently predictive of radiographic progression in a longitudinal observational cohort at 8 years follow up. Arthritis Res. Ther. 2012;14:R30.
Greisen SR, Einarsson HB, Hvid M, Hauge EM, Deleuran B, Kragstrup TW. Spontaneous generation of functional osteoclasts from synovial fluid mononuclear cells as a model of inflammatory osteoclastogenesis. Acta Pathol Microbiol Immunol Scand. 2015;123:779–86.
Svajger U, Jeras M. Anti-inflammatory effects of resveratrol and its potential use in therapy of immune-mediated diseases. Int Rev Immunol. 2012;31:202–22.
Manna SK, Mukhopadhyay A, Aggarwal BB. Resveratrol suppresses TNF-induced activation of nuclear transcription factors NF-kappa B, activator protein-1, and apoptosis: potential role of reactive oxygen intermediates and lipid peroxidation. J Immunol. 2000;164:6509–19.
Tome-Carneiro J, Gonzalvez M, Larrosa M, Yanez-Gascon MJ, Garcia-Almagro FJ, Ruiz-Ros JA, et al. One-year consumption of a grape nutraceutical containing resveratrol improves the inflammatory and fibrinolytic status of patients in primary prevention of cardiovascular disease. Am J Cardiol. 2012;110:356–63.
Tome-Carneiro J, Larrosa M, Yanez-Gascon MJ, Davalos A, Gil-Zamorano J, Gonzalvez M, et al. One-year supplementation with a grape extract containing resveratrol modulates inflammatory-related microRNAs and cytokines expression in peripheral blood mononuclear cells of type 2 diabetes and hypertensive patients with coronary artery disease. Pharmacol Res. 2013;72:69–82.
Timmers S, Konings E, Bilet L, Houtkooper RH, van de Weijer T, Goossens GH, et al. Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab. 2011;14:612–22.
Bo S, Ciccone G, Castiglione A, Gambino R, De Michieli F, Villois P, et al. Anti-inflammatory and antioxidant effects of resveratrol in healthy smokers a randomized, double-blind, placebo-controlled, cross-over trial. Curr Med Chem. 2013;20:1323–31.
Poulsen MM, Fjeldborg K, Ornstrup MJ, Kjaer TN, Nohr MK, Pedersen SB. Resveratrol and inflammation: challenges in translating pre-clinical findings to improved patient outcomes. Biochim Biophys Acta. 2015;1852:1124–36.
Glehr M, Fritsch-Breisach M, Lohberger B, Walzer SM, Moazedi-Fuerst F, Rinner B, et al. Influence of resveratrol on rheumatoid fibroblast-like synoviocytes analysed with gene chip transcription. Phytomedicine. 2013;20:310–8.
Csaki C, Keshishzadeh N, Fischer K, Shakibaei M. Regulation of inflammation signalling by resveratrol in human chondrocytes in vitro. Biochem Pharmacol. 2008;75:677–87.
Chen X, Lu J, An M, Ma Z, Zong H, Yang J. Anti-inflammatory effect of resveratrol on adjuvant arthritis rats with abnormal immunological function via the reduction of cyclooxygenase-2 and prostaglandin E2. Mol Med Rep. 2014;9:2592–8.
Riveiro-Naveira RR, Valcarcel-Ares MN, Almonte-Becerril M, Vaamonde-Garcia C, Loureiro J, Hermida-Carballo L, et al. Resveratrol lowers synovial hyperplasia, inflammatory markers and oxidative damage in an acute antigen-induced arthritis model. Rheumatology. 2016;55:1889–900.
Kragstrup TW, Greisen SR, Nielsen MA, Rhodes C, Stengaard-Pedersen K, Hetland ML, et al. The interleukin-20 receptor axis in early rheumatoid arthritis: novel links between disease-associated autoantibodies and radiographic progression. Arthritis Res. Ther. 2015;18:61.
Olholm J, Paulsen SK, Cullberg KB, Richelsen B, Pedersen SB. Anti-inflammatory effect of resveratrol on adipokine expression and secretion in human adipose tissue explants. Int J Obes. 2010;34:1546–53.
Wakabayashi I, Takeda Y. Inhibitory effects of resveratrol on MCP-1, IL-6, and IL-8 production in human coronary artery smooth muscle cells. Naunyn Schmiedeberg's Arch Pharmacol. 2013;386:835–9.
Shakibaei M, Buhrmann C, Mobasheri A. Resveratrol-mediated SIRT-1 interactions with p300 modulate receptor activator of NF-kappaB ligand (RANKL) activation of NF-kappaB signaling and inhibit osteoclastogenesis in bone-derived cells. J Biol Chem. 2011;286:11492–505.
He X, Andersson G, Lindgren U, Li Y. Resveratrol prevents RANKL-induced osteoclast differentiation of murine osteoclast progenitor RAW 264.7 cells through inhibition of ROS production. Biochem Biophys Res Commun. 2010;401:356–62.
Ornstrup MJ, Harslof T, Sorensen L, Stenkjaer L, Langdahl BL, Pedersen SB. Resveratrol increases osteoblast differentiation in vitro independently of inflammation. Calcif Tissue Int. 2016;99:155–63.
Wang J, Gao JS, Chen JW, Li F, Tian J. Effect of resveratrol on cartilage protection and apoptosis inhibition in experimental osteoarthritis of rabbit. Rheumatol Int. 2012;32:1541–8.
Elmali N, Esenkaya I, Harma A, Ertem K, Turkoz Y, Mizrak B. Effect of resveratrol in experimental osteoarthritis in rabbits. Inflammation. 2005;54:158–62.
Arslan A, Ozcicek F, Keskin Cimen F, Altuner D, Yarali O, Kurt N, et al. Protective effect of resveratrol against methotrexate-induced oxidative stress in the small intestinal tissues of rats. Int J Clin Exp Med. 2015;8:10491–500.
Nikiphorou E, Negoescu A, Fitzpatrick JD, Goudie CT, Badcock A, Ostor AJ, et al. Indispensable or intolerable? Methotrexate in patients with rheumatoid and psoriatic arthritis: a retrospective review of discontinuation rates from a large UK cohort. Clin Rheumatol. 2014;33:609–14.
Walle T, Hsieh F, DeLegge MH, Oatis JE Jr, Walle UK. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab Dispos. 2004;32:1377–82.
Elmali N, Baysal O, Harma A, Esenkaya I, Mizrak B. Effects of resveratrol in inflammatory arthritis. Inflammation. 2007;30:1–6.
Li W, Cai L, Zhang Y, Cui L, Shen G. Intra-articular resveratrol injection prevents osteoarthritis progression in a mouse model by activating SIRT1 and thereby silencing HIF-2alpha. J Orthop Res. 2015;33:1061–70.
Acknowledgements
The authors would like to acknowledge and thank professor Bent Deleuran (Deleuran lab, Aarhus University) for his intellectual and technical support of the study.
Funding
The study received unrestricted grants from the Danish Rheumatism Association, Aage Bang Foundation, Augustinus Foundation and Nyegaard Foundation.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author information
Authors and Affiliations
Contributions
TWK, SL and SBP helped design the study. TWK, SL, AM and MBI collected the samples, carry out experiments, which TWK furthermore supervised. SL analyzed and interpreted the data and drafted the manuscript. All authors helped to critically review analysis and interpretation of the data, were involved in revising the manuscript, and read and approved the final version.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Samples were collected at the rheumatology outpatient clinic, Aarhus University Hospital. Written consent was obtained from all subjects in accord with the Helsinki Declaration. Collection and culturing were, furthermore, approved by the Research Ethics Committee of Central Jutland (20050046 and 20121329) and reported to the Danish Data Protection Agency.
Consent for publication
Not applicable.
Competing interests
TWK is a member of the BMC Rheumatology editorial board but was not involved in the editorial process.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
48 h SFMC cultures – 21 days SFMC cultures. Photographic images showing similar cell density in cultures that were untreated or treated with resveratrol, methotrexate or adalimumab, and showing formation of multinucleated cells in 21 days SFMC cultures. (PDF 443 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Lomholt, S., Mellemkjaer, A., Iversen, M.B. et al. Resveratrol displays anti-inflammatory properties in an ex vivo model of immune mediated inflammatory arthritis. BMC Rheumatol 2, 27 (2018). https://doi.org/10.1186/s41927-018-0036-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41927-018-0036-5