
Abstract
Background
Bogs are unique ecosystems inhabited by distinctive, coevolved assemblages of organisms, which play a global role for carbon storage, climate stability, water quality and biodiversity. To understand ecology and plant–microbe co-occurrence in bogs, we selected 12 representative species of bryophytes and vascular plants and subjected them to a shotgun metagenomic sequencing approach. We explored specific plant–microbe associations as well as functional implications of the respective communities on their host plants and the bog ecosystem.
Results
Microbial communities were shown to be functionally adapted to their plant hosts; a higher colonization specificity was found for vascular plants. Bryophytes that commonly constitute the predominant Sphagnum layer in bogs were characterized by a higher bacterial richness and diversity. Each plant group showed an enrichment of distinct phylogenetic and functional bacterial lineages. Detailed analyses of the metabolic potential of 28 metagenome-assembled genomes (MAGs) supported the observed functional specification of prevalent bacteria. We found that novel lineages of Betaproteobacteria and Actinobacteria in the bog environment harboured genes required for carbon fixation via RuBisCo. Interestingly, several of the highly abundant bacteria in both plant types harboured pathogenicity potential and carried similar virulence factors as found with corresponding human pathogens.
Conclusions
The unexpectedly high specificity of the plant microbiota reflects intimate plant–microbe interactions and coevolution in bog environments. We assume that the detected pathogenicity factors might be involved in coevolution processes, but the finding also reinforces the role of the natural plant microbiota as a potential reservoir for human pathogens. Overall, the study demonstrates how plant–microbe assemblages can ensure stability, functioning and ecosystem health in bogs. It also highlights the role of bog ecosystems as a playground for plant–microbe coevolution.
Video abstract
Similar content being viewed by others
Introduction
Bog ecosystems are one of the oldest vegetation forms in the world; they play a central role in the global carbon cycle and storage as well as a source for fresh water [1,2,3]. This widespread ecosystem is commonly dominated by Sphagnum mosses due to their unique biochemical and morphological adaptation to acidic, water-saturated and nutrient-poor environments [4]. As a keystone species group, Sphagnum mosses engineer this ecosystem, i.e. by acidification due to cation exchange and by forming a deep anaerobic peat layer. Well-adapted, non-vascular (acidophytic liverworts and mosses) and vascular plants (graminoids and dwarf shrubs) commonly co-occur in low abundances embedded in the Sphagnum layer [5,6,7]; therefore, they have different strategies regarding their growth form and root architecture [6]. Interactions between Sphagnum mosses and the other plants are important for the formation of the hummock topology and nutrient cycling, which facilitates their growth in the bog ecosystem [5, 7]. The bog microbiome plays a crucial role for bog ecosystem functioning, nutrient cycling and protection towards biotic and abiotic stress [8,9,10]. Previously, a core microbiome or meta-community of bog plants was identified [11], but knowledge about plant-specific distribution patterns and functions is still missing.
Bacteria are known to be the main constituent of the bog microbiome, co-occurring and interacting with archaea, fungi and other members of the microbiome [8, 12, 13]. Moreover, bog ecosystems were found to be natural hotspots for a high diversity of bacterial species and antagonists [12] and microbial genes [11] and interestingly also for antibiotic resistance genes [4, 11, 14]. A vast majority of bog bacteria were found to engage in a plant-associated, often endophytic life style [4]. The plant microbiota is involved in plant growth, productivity, adaptation, diversification and health [15,16,17], which is of special importance under extreme conditions in bog environments. Specifically, Sphagnum mosses were shown to contain a specific bacterial community which fulfils important roles for ecosystem functioning and health [1, 8, 12, 18,19,20,21]. A previous study indicated that Acidobacteria, Actinobacteria and Proteobacteria are highly responsive towards substrate-specific changes in the bog ecosystem [22]. Interestingly, studies indicate an extraordinary high degree of plant-specific microorganisms [11, 18, 23]; this can be explained by plant–microbe coevolution [17]. Coevolution was especially studied for microbial symbionts and pathogens, but the degree to which specialization and coevolution between plant species and microbes in ecosystems occur is relatively understudied [24]. For bogs, our hypothesis is that vascular plants with vascular tissues, which allow transport and diversification and which have more developed defence mechanisms against pathogens [25], harbour a more specific but less diverse microbiota than the phylogenetically older bryophytes.
Therefore, we studied bacterial community structures and functions in typical circumpolar bog vegetation of two Austrian bogs (Rotmoos and Pürgschachen Moor). We selected representative plants that belong to two separate phylogenetic branches, bryophytes, i.e. Sphagnum magellanicum, Sphagnum angustifolium and Sphagnum fuscum (peat mosses), Pleurozium schreberi (Brid.) Mitt. (Schreber’s big red stem moss), Polytrichum strictum (bog haircap moss) and Mylia anomala (anomalous flapwort; liverwort), and vascular plants, i.e. Vaccinium myrtillus (European blueberry), Vaccinium oxycoccus (bog cranberry), Calluna vulgaris (heather), Andromeda polifolia (bog rosemary), Eriophorum vaginatum (tussock cottongrass) and Pinus mugo (dwarf mountain pine), and analysed them using a shotgun metagenomic sequencing approach. In combination with targeted bioinformatics, such data can be used for functional microbiota profiling and to capture detailed taxonomic diversity of the entire microbiome [26]. Specifically, we addressed the following questions: (i) Are there general differences in bacterial communities between bryophytes and vascular plants? (ii) Do bacteria harbour functions that are specific for the different plant host interactions? and (iii) Do different bacterial taxa have distinct roles to support bog ecosystem functioning? Overall, this study provides key insights into the ecosystem function of bog-associated bacteria that are at present largely underrepresented in global surveys.
Material and methods
Sample collection and shotgun metagenomic sequencing
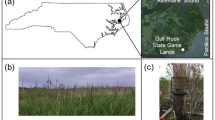
The samples for the analyses were obtained from northern Styria, Austria, in November 2012 within two geographically distinct peat bogs, Rotmoos (N47 41.029 E15 09.284 and N47 41.059 E1509.269) and Pürgschachen Moor (N47 34.835 E14 20.390 and N47 34.815 E14 20.482). In the past, the microbiome in these two bogs was intensively studied with a comprehensive experimental design [23, 27]. Based on these results, which showed markedly high species specificity of microbial communities, we developed a specific design for metagenomics approaches, which were already used to describe ecosystem functioning under extreme conditions as well as the role of archaea [11, 28]. At both sites, we selected randomly two plots (each of 1 m2) that represent the two typical plant communities dominated by either Sphagnum magellanicum or Sphagnum angustifolium. Bryophytes and vascular plants included in this study represent exemplary species that grow naturally in peat bogs; plant cover per plot together with environmental variables are detailed in Table S1. The metagenomic datasets (Table S2) were generated as previously described [11, 28]. In brief, total genomic DNA was extracted using the FastDNA Spin kit for soil (MP Biomedical, USA) following the manufacturer’s protocol. Shotgun metagenomic sequencing was performed by the sequencing provider Eurofins MWG Operon (Ebersberg, Germany). DNA library preparation was performed using a TruSeq DNA library kit (Illumina, USA) before the sequencing. The extracted DNA was then sequenced using an Illumina HiSeq 2500 system PE 2 × 150 bp (Illumina, USA) resulting in a range of 22.7–40.9 million reads per individual metagenome (Table S2). The datasets were previously used to explore archaeal community structure and function [28]. However, they remained entirely unexplored in terms of bacterial diversity which was previously one of the main constituents of bog microbiome [8, 27]. In the present study, the available raw data was subjected to a targeted bioinformatic approach to address this important component of the microbiome.
Bacterial community structure and diversity analysis
Kraken2 v2.0.9 and Bracken v2.6.0 were used to measure the bacterial taxonomic composition and estimate the diversity of the short-read sequencing samples [29, 30]. Kraken2 implements a k-mer-based approach to classify individual metagenome reads by map** all k-mer to the lowest common ancestor (LCA) of all genomes containing the given k-mer. The standard Kraken2 database was used to classify individual reads. Following Kraken2 taxonomic assignment, Bracken was used to estimate species abundances. To account for uneven sequencing depth, the datasets were normalized by rarefying to the lowest number of reads (2,271,071) and using MetagenomeSeq’s cumulative sum scaling (CSS; [31]) for subsequent alpha (species richness) and beta (variation in species composition) diversity analysis, respectively. Phyloseq, MicrobiomeAnalyst and vegan R packages implemented in RStudio v 1.3.1093 were used to analyse bacterial community diversity and composition [32,50].
Detection of nitrogen cycling genes in metagenomic data
Reads from the metagenomes were aligned against NCycDB, a manually curated integrative database for nitrogen cycle gene profiling from shotgun metagenome sequencing data [51] using the blast algorithm in DIAMOND v0.9.35.136 [36]. The generated distance matrix was subjected for PERMANOVA to investigate if MAG phylogeny explains the composition of functional genes within the MAGs. The distance matrix was further visualized by using principal coordinate analysis (PCoA) plots. A heatmap plot was used to visualize the functional feature abundance profiles. Venn diagram was generated using InteractiVenn [64].
Results
Bacterial diversity with the bog vegetation
Targeted diversity analyses were conducted in order to resolve if bacterial community richness and structure differ between vascular plants and bryophytes although they grew in close proximity and all individuals were embedded into Sphagnum mosses. Alpha diversity analysis indicated that plant types (vascular plants and bryophytes) significantly affected bacterial richness and community structure (Fig. 1a). Based on the Kraken2 taxonomic classifier, a total of 5699 bacterial species was identified within the metagenome sequence library. Respective rarefaction curves indicate the sampling size was sufficient to capture overall bacterial diversity (Figure S1). A significantly higher number of species (P = 0.025) and Shannon index (P = 0.016) was observed in bryophytes (S = 5533, H’ = 7.0) in comparison to vascular plants (S = 5451, H’ = 6.7; Fig. 1a). A total of 37 core taxa with a relative abundance > 0.01% were exclusively detected in bryophytes. Beta diversity analysis revealed significantly different clustering between bacterial community structures of vascular plants and bryophytes (R2 = 0.229, P = 0.004; Fig. 1b). Additional beta diversity analyses were performed by analysing bacterial communities on phylum, order and genus levels (Figure S2). At higher taxonomic levels (phylum and class), the bacterial community composition was relatively similar between bryophytes and vascular plants (R2 = 0.175, P = 0.140; R2 = 0.130, P = 0.124; Figure S2a & b). However, a distinct clustering between the two groups was observed at lower taxonomic levels (order and genus) when subjected to the same analysis (R2 = 0.250, P = 0.015; R2 = 0.218, P = 0.004; Figure S2c & d).

Bacterial richness, community clustering, composition and identified taxa that are either specific for bryophytes or vascular plants. Bacterial richness was assessed with the Shannon index (a). Community clustering of bacteria in bryophytes and vascular plants is visualized in a two-dimensional PCoA plot based on a Bray–Curtis matrix (b). Bacterial composition is shown at the class level (c). LEfSe analysis indicated specific biomarkers at bacterial order for both plant groups. Only bacterial taxa with LDA scores > 1 and Padjusted < 0.1 according to LEfSe analysis are shown (d)
Among bacterial phyla, Proteobacteria (74.1%) was the predominant taxonomic group in the bog vegetation followed by Actinobacteria (14.7%) and Acidobacteria (4.2%). Specific differences in bacterial composition were observed at lower taxonomic levels. A relatively higher proportion of Alphaproteobacteria and Gammaproteobacteria was observed for vascular plants (38.2 and 20.4%, respectively; Fig. 1c) in comparison to bryophytes (36.5 and 16.4%). In contrast, the bacterial class Acidobacteria was relatively more abundant in bryophytes (4.9%) compared to vascular plants (3.4%). A complementary LEfSe analysis indicated that several bacterial lineages within Proteobacteria (i.e. Rhizobiales, Pseudomonadales, Xanthomonadales and Pasteurellales) and Cyanobacteria (i.e. Synechococcales, Nostocales, Synechococcales, Chroococcales) were significantly enriched in vascular plants whereas bacterial lineages that belong to Verrucomicrobia (i.e. Verrucomicrobiales and Opitutales) and Planctomycetes (i.e. Planctomycetales) were significantly enriched in bryophytes (Fig. 1d).
Profiling of bacterial functions in the bog microbiome
Due to the observed differences in the bacterial community structure, we expected that the overall functional profiles of the respective communities would be also different. Following filtering of metagenomic data, 8.56 × 106– 27.9 × 106 reads were annotated as bacterial genes (Table S2). Functional annotation assigned the reads into 5852 KOs. A high proportion of them (5522 KOs; 94.3%) was detected in all samples indicating a large functional core in the bog microbiome. The KOs were mainly associated with carbohydrate, amino acid and energy metabolism (22.1, 18.8 and 12.3%, respectively; Fig. 2a). Moreover, specific KOs were associated with functions known to be prevalent in plant-associated microbes, i.e. ABC transporters (n = 220, 3.8%), quorum sensing (n = 63, 1.1%), nitrogen metabolism (n = 41, 0.7%) and biosynthesis of siderophores (n = 20, 0.34%). Methane oxidizing bacteria are important to reduce methane emissions from peat bogs and provide CO2 for photosynthesis [1]. We also detected genes involved in methane oxidation, i.e. pmoA, pmoB, pmoC and mmoX that belong to Alphaproteobacteria. Altogether, beta diversity analysis indicated a clear clustering for microbiota’s functioning in bryophytes and vascular plants (R2 = 22.9%, P = 0.004; Fig. 2b).

Comparison of functional assignment of the bog-associated microbiome, functional community clustering (a, b) and number of bacterial KEGG Orthology (KO) that enriched or depleted between bryophytes and vascular plants (c). Annotated genes were assigned to corresponding KEGG Orthology (KO) functional categories (a). A Bray–Curtis distance matrix between samples was visualized using a principal coordinate analysis (PCoA) plot (b). c Numbers of bacterial genes that were statistically enriched/depleted according to differential gene abundance analysis were obtained by using edgeR (log2 fold change > 1 and Padjusted values below 0.1). These genes were grouped according to KEGG pathways (third tier) and visualized in a bar plot (c)
A pairwise comparison using edgeR revealed that 290 KOs (log2 fold change > 1, Padjusted < 0.1, Table S4) were significantly different between bryophytes and vascular plants. A total of 216 KOs was significantly enriched in vascular plants, whereas 74 KOs were reduced (Fig. 2c). Among the enriched KOs, 33 were assigned to ABC transporter pathways in vascular plants. Moreover, four KOs that were involved in oligosaccharide (i.e. galactose, raffinose and sorbitol), six KOs that were involved in monosaccharide (i.e. L-arabinose and xylitol) and six KOs that were involved in biotin and amino acid transporters (i.e. octopine and cystine) were enriched in vascular plants whereas two KOs assigned to bicarbonate transport system substrate-binding protein and cobalt/nickel transport system permease protein were enriched in bryophytes. A higher abundance of KOs (n = 6) that are related to the biosynthesis of other secondary metabolites (i.e. pinoresinol/lariciresinol reductase and antibiotic kanosamine) was also observed in vascular plants. In addition, a total of three KOs related to the biosynthesis of vitamins, i.e. vitamin K2, B1 and B12, were also enriched in vascular plants. The siderophore group represented by aerobacterin (K03897 and K03894), yersiniabactin (K04784) and mycobactin (K04789 and K04792) was also more abundant in vascular plants compared to bryophytes, whereas the siderophore enterobactin (k02361) showed the opposite trend. Within nitrogen metabolism, six KOs annotated as nitrogenase and nitrate reductase (nifH-K02588, nifK-K02591, narG-K00370, narH-K00371, nifD-KO2586 and napA-K02567) were enriched in vascular plants. Contigs that were assigned to nifH originated from Alphaproteobacteria and Betaproteobacteria. In contrast, KOs involved in degradation of lysine (n = 3), dioxin (n = 2) and aromatic compounds, i.e. aminobenzoate (n = 3), ethylbenzene (n = 2), naphthalene (n = 1), chlorocyclohexane (n = 1) and chlorobenzene (n = 1), were enriched in bryophytes. Notably, specific CRISPR-associated proteins (n = 6) were also enriched in the bryophytes indicating potentially higher selection pressures from bacteriophages in these plants (Fig. 2c). The overall results indicate that despite a high share of functional features, the plant hosts enrich not only certain taxonomic groups (Fig. 1d), but also distinct functional groups to support host functioning.
Profiling of nitrogen cycling genes in the bog microbiome
As the nitrogen cycle is an important biogeochemical pathway in the bog ecosystem, we further explored if the respective communities (associated to bryophytes and vascular plants) were associated to distinct nitrogen cycling profiles. Using a high-stringency approach (90% similarity and < 10−5E-value), a total of 234,505 reads (ranging from 6417 to 36,098 reads per plant) was annotated as nitrogen cycling genes (Table S2). Beta diversity analysis indicated a distinct cluster between bryophytes and vascular plants (R2 = 47.6%, P = 0.003; Fig. 3a). Furthermore, a pairwise comparison using edgeR indicated that the abundance of genes that are involved in nitrogen fixation and denitrification was significantly higher in bryophytes compared to vascular plants (Fig. 3b, Table S5). However, the differences in log-fold change were only 0.7 and 0.4, respectively. When abundance profiles were analysed at the gene level, we observed that 11 genes that are involved in nitrogen cycling (Table S5) significantly differed between bryophytes and vascular plants. The abundance of the nifH gene was significantly higher in bryophytes. This result was in contrast to the abundance analysis using the eggNOG database (Table S4, see K02588-nifH) that indicated that the abundance of nifH was higher in vascular plants. A set of genes encoding particulate methane monooxygenase (pmoB and pmoC) which also have a shared high sequence similarity with the amo gene family [51] were enriched in vascular plants (Fig. 3). Genes encoding nitrate reductase (NAD(P)H) (NR), glutamate synthase [NADH] (GLT1) and nitrate reductase (narH) were also enriched in the vascular plants.

Clustering of nitrogen cycling gene profiles in vascular plants and bryophytes (a) and bar plots showing pathways (b) and genes (c) involved in nitrogen cycling that were enriched or reduced. A Bray–Curtis distance matrix between samples was visualized using a principal coordinate analysis (PCoA) plot (a). Pathways that were statistically enriched/reduced according to edgeR (Padjusted < 0.1) were included (b). Nitrogen cycling genes that were statistically enriched/reduced according to edgeR (log2 fold change > 1 and Padjusted < 0.1) were included (c). Negative log2 fold change values indicate that the respective genes are more abundant in bryophytes whereas positive values indicate that the genes are more abundant in vascular plants
Genome-centric analysis of prevalent bacteria in the bog vegetation
In total, 28 MAGs with completeness levels more than 80% and contamination levels lower than 10% were reconstructed from bog metagenome datasets representing the most abundant members of the bacterial communities. Taxonomical assignment (Fig. 4a, Table S6) MAGs (completeness > 80%) resulted in the classification to six phyla, i.e. Proteobacteria (n = 11), Acidobacteria (n = 6), Actinobacteria (n = 6), Verrucomicrobia (n = 2), Bacteroidetes (n = 2) and Chlamydiae (n = 1). One MAG was assigned to Candidatus Eremiobacteraeota which was previously known as Candidate division WPS-2 (bin ID: MAG_22). Despite high completeness (> 90%), several MAGs were only assignable at the class level, i.e. Gammaproteobacteria (MAG_2, MAG_99) and Betaproteobacteria (MAG_119, MAG_129, MAG_110), indicating a possible occurrence of novel bacterial lineages in the bog microbiome. Among MAGs, two MAGs (MAG_178 and MAG_201) that belong to Verrucomicrobia and Acidobacteria, respectively, were more abundant in non-vascular plants whereas four MAGs that belong to Proteobacteria (MAG_154, MAG_111, MAG_2 and MAG_119) were more abundant in vascular plants (Padjusted < 0.1, Table S7). This result indicated that despite similar abundances from the majority of MAGs, the plant host seems to allow specific bacterial taxa to enrich as shown previously (Fig. 1d).

Phylogenetic tree of reconstructed metagenome-assembled genomes (MAGs) and clustering based on KEGG Orthology (KO) counts from each MAG. Different bacterial phylum and proteobacterial classes that are presented in the phylogenetic tree constructed MAGs are highlighted with different colours (a). Dots represent MAGs that were generated with metagenomic data from the bog ecosystem. Principal coordinate analysis (PCoA) of MAG clusters is based on KO counts in the genomes (b)
A broad phylogenetic representation of bacteria and high genome completeness facilitated detailed assessments of the functional potential of different taxonomic groups in the bog vegetation. MAGs belonging to Chlamydiae (MAG_12) and Candidatus Eremiobacteraeota (Candidate division WPS-2; MAG_22) were excluded from the PERMANOVA analysis due to insufficient replicates for these groups (Fig. 4, Table S6). Function-based clustering in a PCoA plot indicated the occurrence of specific clusters was determined by MAG phylogeny (Fig. 3b). This was confirmed by a complementary statistical assessment with PERMANOVA (P < 0.001, R2 = 0.471). Three distinct clusters were observed, i.e. cluster 1 (Actinobacteria MAGs), cluster 2 (Alpha-, Beta- and Gammaproteobacteria) and cluster 3 (Acidobacteria, Bacteroidetes and Verrucomicrobia). This observation indicates that various functional groups are present within naturally occurring bacterial communities in the bog vegetation. In a subsequent step, functions that differed between MAGs (Fig. 5) and that might be involved in plant–microbe interactions were explored in detail. Alpha-, Beta-, Gammaproteobacteria and Actinobacteria encoded a higher number of genes involved in carbon fixation pathways, i.e. atoB (acetyl-CoA C-acetyltransferase) and sdhD. In fact, the majority of MAGs that belong to Alpha-, Beta-, Gammaproteobacteria and Actinobacteria had a complete gene set of sdhABCD/fdrABCD. The latter taxa also had a higher number of genes involved in carbon fixation pathways, i.e. mcl (citramalyl-CoA lyase), than other MAG groups whereas Alpha-, Beta- and Gammaproteobacteria harboured a higher number of pta gene encode phosphate acetyltransferase. Moreover, we also detected small and large subunits of RuBisCO genes and aceA which encodes isocitrate lyase in three MAGs (MAG_39, MAG_129 and MAG_194) that were assigned to Alpha-, Betaproteobacteria and Actinobacteria. A set of genes that are involved in oxidative phosphorylation, especially cytochrome o ubiquinol oxidase (cyo) and cytochrome aa3 quinol oxidase (qox), were found to be more prevalent in MAGs of the Proteobacteria phylum compared to others (Fig. 5). Proteobacteria and Actinobacteria might play a crucial role in carbon fixation in the bog ecosystem (Fig. 5). The latter taxa also had a set of genes that are involved in F420 biosynthesis, which plays a central role in methanogenesis due to the presence of cofC, cofD, cofE and fbiC. Genes involved in assimilatory sulfate reduction were evenly distributed (cysN, cysD, cysNC, cysI and cysJ) among MAGs. However, Acidobacteria and Verrucomicrobia harboured a higher number of genes related to sulfur metabolism through the Sox system such as soxB and soxX, respectively (Fig. 5). Genes that are involved in assimilatory/dissimilatory nitrate reduction such as narB, nirA, nirB and nirD were detected in MAGs that belong to Actinobacteria (MAG_194 and MAG_172), Alphaproteobacteria (MAG_111), Betaproteobacteria (MAG_129 and MAG_110) and Gammaproteobacteria (MAG_2).

Heatmap plots showing abundance profiles for each bacterial group based on KEGG Orthology (KO) counts in MAGs. Different bacterial phyla and proteobacterial classes are highlighted with different colours. Act Actinobacteria, Al Alphaproteobacteria, Be Betaproteobacteria, Ga Gammaproteobacteria, Aci Acidobacteria, Ba Bacteroidetes, Ve Verrucomicrobia. The colour gradient from green to red represents a number of KO counts present in each MAG from low to high
In addition to genes from the central carbohydrate and energy metabolism, also, genes that are related to vitamin biosynthesis, metabolism of secondary metabolites and abiotic stress responses and thus potentially important for host-microbe interactions were detected. A set of genes (bioA, bioB, bioC and bioF) that is involved in biotin metabolism was detected in MAGs that belong to Actinobacteria, Gammaproteobacteria, Bacteroidetes and Verrucomicrobia. Specific genes involved in menaquinone biosynthesis (menA, menB, menC, mend and menF) were prevalent in Actinobacteria. A collection of genes involved in the MEP/DOXP pathway (dxr, ispA, ispB, ispD, ispE, ispF, gcpE, ispH and idsA) were detected in the majority of MAGs, except from those that belong to Gammaproteobacteria. Interestingly, Gammaproteobacteria may also play an important role in the production of terpenoid but through the mevalonate pathway due to the presence of idi, mvaA, mvaD, mvk and mvkA2 genes (Fig. 5). Several molecular chaperones, i.e. hslO, ibpA, ibpB and pccA, were more prevalent in MAGs that belong to Proteobacteria, which indicates their enhanced ability to mitigate environmental stress.
Among the MAGs, two MAGs (MAG_178 and MAG_201) that were assigned to Verrucomicrobia and Acidobacteria, respectively, were more abundant in bryophytes. In contrast, four MAGs that belong to Proteobacteria (MAG_154, MAG_111, MAG_2 and MAG_119) were more abundant in vascular plants (Padjusted < 0.1; Table S7). A key feature that was found in MAG_178 and MAG_201. was that these MAGs harboured a complete set of enzymes that are involved in riboflavin (B2) biosynthesis, i.e. GTP cyclohydrolase II and riboflavin synthase. On the other hand, all of the MAGs that were more prevalent in vascular plants harboured genes that encode urea transport system permease protein (urtA, urtB, urtC, urtD and urtE). In particular, MAG_154 also harboured genes that encode multiple sugar transport system ATP-binding proteins. These results indicate that despite similar abundances of the majority of analysed MAGs, that plant host likely facilitates colonization of bacterial taxa with specific functions (Fig. 1) to support plant host physiology.
Previous studies provided first indications that the keystone species in the bog ecosystem, Sphagnum, is a reservoir for potentially opportunistic human pathogens [14, 65]. In this study, we also identified a list of potential human pathogens, i.e. Clamydia and Mycobacterium spec. div. Therefore, we searched for potential virulence factor homologs (≥ 50% identity and < 1e − 10 E-value) that are prevalent among the MAGs (Table S8). Calculation of the average distance of the MAGs and clinical isolate genomes indicated cluster formation that was mainly attributable to bacterial class level (Figure S3). This result was strengthened by PERMANOVA analysis which indicated that the origin of isolates (clinical and environment) only explained a small variation virulence gene composition (R2 = 2.4%) while taxonomical identity explained 29.6% of the variation. There was no clear differentiation in the virulence gene composition between the genomes of closely related clinical and environmental isolates within the clusters. Nevertheless, we identified a number of genes that were exclusively detected in the bog MAGs in comparison to the closely related clinical isolate genomes. For instance, when MAG 172 was compared with a clinical Mycobacterium, MAG12 with clinical Chlamydia isolates and MAG_111 with clinical Sphingomonas isolates, bog-specific differences were found (Figure S4; Table S9). When analysed in detail, a gene that encodes catalase peroxidase (katA) was detected in MAGs 110 and 172 but was not present in the clinical isolates. Specific genes that are involved in iron uptake, i.e. ccm, ybtQ and mbtJ, were also only present in MAGs generated from the bog ecosystem when compared to their closely related clinical taxa. Genes encoding the phytotoxin phaseolotoxin were exclusively detected in MAGs 111 and 139 that belong to Sphingomonadaceae and Betaproteobacteria, respectively.
Discussion
In the present study, we showed that vascular plants and bryophytes enrich certain bacterial taxa with distinct functional properties reflecting plant–microbe coevolution. The differences in community composition were statistically significant despite the fact that all analysed plant species co-occur in close proximity and are connected by a bacterial meta-community [11]. Plant–microbe coevolution was already reported for distinct plants, interactions and processes [17, 24]; here, we show complementary implications at the ecosystem level by employing a shotgun metagenomic sequencing approach using a novel bioinformatic pipeline and detailed genome-centric analysis. Compared with previous studies, we found concordance in terms of the prevalence of Proteobacteria, but also observed differences in the relative abundance of less abundant taxa [11]. This is commonly observed when studies based on the amplification of marker genes are compared with such based on shotgun metagenomics [66,67,68]. In this context, it is noteworthy to mention that using shotgun metagenomics we detected a higher number of bacterial taxa (5699 vs. 4898 bacterial OTUs/species). Bogs are important native models to study general rules of plant–microbe coevolution and specificity, because these ecosystems have been stable for centuries and are shaped by extreme but similar abiotic factors around the world [69,70,71]. In addition to confirming previous findings with complementary approaches, we further illustrated that the bog microbiome is collectively involved in beneficial functions, which are referred to as microbiome services, i.e. nutrient supply, plant growth promotion and vitamin biosynthesis to support bog ecosystem functioning (Fig. 6).

Schematic illustration of bacterial groups associated with vascular plants and bryophytes contributing to bog ecosystem functioning. Different bacterial groups involved in bog ecosystem functioning are indicated by different colours (red: Alphaproteobacteria, green: Betaproteobacteria, purple: Gammaproteobacteria, gold: Actinobacteria, blue: Acidobacteria, black: Bacteroidetes and grey: Verrucomicrobia). Overall microbiome services and plant-specific (for bryophytes and vascular plants) traits were included in the lower part of the figure
Bryophytes were found to be associated with a significantly higher bacterial richness and diversity in comparison to vascular plants. Nevertheless, the bacterial alpha diversity was found to be in a similar range for both plant systems. We argue that the spatial proximity of plants facilitates the transfer of microorganisms, leading to an increase in overall diversity while maintaining plant group specificity. Bryophytes, including Sphagnum mosses, provide an exceptional habitat for plant-associated bacteria and their richness is tightly connected with the bog ecosystem functioning [27]. A recent study showed that a decline in the microbial diversity of Sphagnum mosses was correlated with a decrease in N2 fixation activity [72], which highlights the importance of bacterial richness for the overall bog ecosystem functioning. The community composition was more similar among different bryophytes, and more different between the vascular plant species. The latter are known to recruit a specific fraction of bacterial taxa from their surrounding environment due to secretory activities in their root system [16], which is also suggested as an entry point for bacteria to colonize above-ground plant compartments [73]. In vascular plants, bacterial diversity is commonly reduced from soil to rhizosphere to endosphere [16]. In contrast, bryophytes, which lack functional root structures, do not rely on their belowground compartments to recruit bacteria. Processes of water and mass transfer run over the entire surface of the plants. They can maintain a high bacterial diversity within the whole lifecycle, independent of their respective environment [23]. A foregoing study indicated that the liverwort Marchantia paleacea harboured a higher bacterial diversity than its soil substrate [74]. Vascular plants are more influenced by host-environmental interactions especially in the bog ecosystem with its extreme conditions, i.e. low pH, nutrient and high water saturation [4, 23].
When bacterial community composition was explored in the present study, a subset of differing bacterial features was detected between vascular plants and bryophytes indicating that the analysed plant hosts harboured functionally adapted bacterial communities. In the bog ecosystem, vascular plants and bryophytes grow in close proximity and are mostly embedded in the predominant Sphagnum plants. Therefore, it was to be expected that a majority of the prevalent bacteria and their functions, i.e. ABC transporters, quorum sensing, nitrogen metabolism and biosynthesis of siderophores, are widely shared by hosts as already shown before by marker gene analyses [8, 12, 28]. Nevertheless, plant-specific differences in the bacterial community, especially at the bacterial order level and below, as well as their function were observed. Previous studies have provided concordant results for specific functional groups in these environments [23, 75, 76]; however, they did not assess the entire taxonomic and functional diversity in typical bog ecosystems. Coevolution between microbes and hosts has apparently led to nuanced functional specificity in different plant groups. We detected a higher abundance of bacterial genes involved in aromatic compound degradation in bryophytes that might be explained by the typical accumulation of the aromatic compound in Sphagnum bogs that can be used as a carbon source by bacteria [77, 78]. In nutrient-poor environments, bryophytes such as Sphagnum mosses can absorb and accumulate mineral nutrients all over their surface highly effective even in extreme nutrient-poor environments, which gives them a competitive advantage over the vascular plant [79]. In contrast, root systems of typical vascular plants in bog ecosystems, e.g. Eriophorum vaginatum, Calluna vulgaris, Vaccinium myrtillus and Vaccinium oxycoccus that were included in the present study, can penetrate into deeper portions of the bog with their root systems [6, 28]. ABC transporter genes that are involved in the uptake of oligosaccharides and amino acids were enriched in the vascular plants. Enrichment of bacteria with this function might be more important for microbes of vascular plants growing in bogs to increase nutrient uptake from the environment and support their growth under nutrient-limited conditions.
Bacterial communities of bryophytes and vascular plants were shown to harbour different nitrogen cycling gene profiles. In nitrogen-poor environments, bacteria play a crucial role in nitrogen acquisition for the growth and development of plants (reviewed in [80, 81]). Nitrogen-fixing bacteria with nifH genes were shown to be more prevalent in bryophytes in comparison to vascular plants when an extended reference database was used. It should be noted that when solely using the eggNOG database, the abundance of nifH genes was found to be higher in vascular plants. However, the read number that was identified by using the eggNOG database was substantially lower (898 reads) when compared to NCycDB database hits (7608 reads). NCycDB was constructed by integrating nitrogen cycling genes from various databases (COG, KEGG, SEED and eggNOG) and thus provides a better reference set for assignments. This result also highlighted that bryophytes, especially Sphagnum, contained a high diversity of bacterial genes involved in nitrogen fixation. It is in agreement with previous findings describing the potential of nitrogen-fixing bacteria in bogs and possible occurrence of novel nifH variants [27]. Hence, this observation corroborates the importance of nitrogen-fixing bacteria which were mainly assigned to Alphaproteobacteria, for Sphagnum mosses [4, 8, 12]. In contrast to the observed abundance of nifH genes, a higher number of genes encoding nitrate reductases for denitrification processes were observed in metagenomes of vascular plants. The results are in accordance with previous findings that different plant species harbour specific microbial communities with specific functions even when grown on the same environment [82, 83].
By including a detailed genome-centric analysis of microbial communities, we were able to identify the metabolic potential of individual MAGs and highlight the specific roles of each functional guild. Alpha-, Beta- and Actinobacteria likely play an important role in carbon fixation in this ecosystem. Previous studies have shown that autotrophic bacteria within the bacterial phyla Proteobacteria and Actinobacteria significantly contribute to carbon fixation in various environments [84, 102]. However, these genes can become pathogenicity factors when transferred to other environments. In the present study, we could even show that some environmental isolates can harbour more virulence factors than their clinical counterparts, which further confirms that many human pathogens likely originated in the environment.
Conclusion
Differences in the abundance of specific bacterial taxa with distinct functions between vascular plants and bryophytes indicate coevolution with their host plants. By employing genome-centric analyses, we could identify a highly complex metabolic potential within the bog microbiome and expand our understanding of the role of microorganisms in bog vegetation. The extraordinary high functional microbial diversity discovered in bog ecosystems allows understanding of stability, functioning and ecosystem health in bogs. However, this includes a high diversity of antimicrobial resistance as well as virulence genes, which indicates not only a reservoir for multi-resistant pathogens but also a source for studying suppression of them under natural conditions and emergence under anthropogenic influence.
Availability of data and materials
This shotgun metagenome project has been deposited in the European Nucleotide Archive (ENA) database under the study number PRJEB39100 and accession numbers ERR4298333 and ERR4298344.
References
Raghoebarsing AA, Smolders AJ, Schmid MC, Rijpstra WIC, Wolters-Arts M, Derksen J, et al. Methanotrophic symbionts provide carbon for photosynthesis in peat bogs. Nature. 2005;436:1153–6.
Hopple A, Wilson R, Kolton M, Zalman CA, Chanton JP, Kostka J, et al. Massive peatland carbon banks vulnerable to rising temperatures. Nat Commun. 2020;11:1–7.
Dise NB. Peatland response to global change. Science. 2009;326:810–1.
Kostka JE, Weston DJ, Glass JB, Lilleskov EA, Shaw AJ, Turetsky MR. The Sphagnum microbiome: new insights from an ancient plant lineage. New Phytol. 2016;211:57–64.
Groeneveld EV, Masse A, Rochefort L. Polytrichum strictum as a nurse-plant in peatland restoration. Restor Ecol. 2007;15:709–19.
Malmer N, Svensson BM, Wallén B. Interactions between Sphagnum mosses and field layer vascular plants in the development of peat-forming systems. Folia Geobot Phytotaxon. 1994;29:483–96.
Pouliot R, Rochefort L, Karofeld E, Mercier C. Initiation of Sphagnum moss hummocks in bogs and the presence of vascular plants: is there a link? Acta Oecologica. 2011;37:346–54.
Bragina A, Oberauner-Wappis L, Zachow C, Halwachs B, Thallinger GG, Müller H, et al. The Sphagnum microbiome supports bog ecosystem functioning under extreme conditions. Mol Ecol. 2014;23:4498–510.
Larmola T, Leppänen SM, Tuittila E-S, Aarva M, Merilä P, Fritze H, et al. Methanotrophy induces nitrogen fixation during peatland development. Proc Natl Acad Sci. 2014;111:734–9.
Hausmann B, Knorr K-H, Schreck K, Tringe SG, Del Rio TG, Loy A, et al. Consortia of low-abundance bacteria drive sulfate reduction-dependent degradation of fermentation products in peat soil microcosms. ISME J. 2016;10:2365–75.
Bragina A, Berg C, Berg G. The core microbiome bonds the Alpine bog vegetation to a transkingdom metacommunity. Mol Ecol. 2015;24:4795–807.
Opelt K, Chobot V, Hadacek F, Schönmann S, Eberl L, Berg G. Investigations of the structure and function of bacterial communities associated with Sphagnum mosses. Environ Microbiol. 2007;9:2795–809.
Woodcroft BJ, Singleton CM, Boyd JA, Evans PN, Emerson JB, Zayed AA, et al. Genome-centric view of carbon processing in thawing permafrost. Nature. 2018;560:49–54.
Obermeier MM, Wicaksono WA, Taffner J, Bergna A, Poehlein A, Cernava T, et al. Plant resistome profiling in evolutionary old bog vegetation provides new clues to understand emergence of multi-resistance. ISME J. 2021;15:921–37.
Berg G, Rybakova D, Grube M, Köberl M. The plant microbiome explored: implications for experimental botany. J Exp Bot. 2016;67:995–1002.
Bulgarelli D, Schlaeppi K, Spaepen S, Van Themaat EVL, Schulze-Lefert P. Structure and functions of the bacterial microbiota of plants. Annu Rev Plant Biol. 2013;64:807–38.
Cordovez V, Dini-Andreote F, Carrión VJ, Raaijmakers JM. Ecology and evolution of plant microbiomes. Annu Rev Microbiol. 2019;73:69–88.
Bragina A, Maier S, Berg C, Müller H, Chobot V, Hadacek F, et al. Similar diversity of Alphaproteobacteria and nitrogenase gene amplicons on two related Sphagnum mosses. Front Microbiol. 2012;2:275.
Opelt K, Berg C, Schönmann S, Eberl L, Berg G. High specificity but contrasting biodiversity of Sphagnum-associated bacterial and plant communities in bog ecosystems independent of the geographical region. ISME J. 2007;1:502–16.
Shcherbakov A, Bragina A, Kuzmina EY, Berg C, Muntyan A, Makarova N, et al. Endophytic bacteria of Sphagnum mosses as promising objects of agricultural microbiology. Microbiology. 2013;82:306–15.
Pankratov TA, Ivanova AO, Dedysh SN, Liesack W. Bacterial populations and environmental factors controlling cellulose degradation in an acidic Sphagnum peat. Environ Microbiol. 2011;13:1800–14.
Dedysh SN, Ivanova AA. Planctomycetes in boreal and subarctic wetlands: diversity patterns and potential ecological functions. FEMS Microbiol Ecol. 2019;95:fiy227.
Bragina A, Berg C, Cardinale M, Shcherbakov A, Chebotar V, Berg G. Sphagnum mosses harbour highly specific bacterial diversity during their whole lifecycle. ISME J. 2012;6:802–13.
Baltrus DA. Adaptation, specialization, and coevolution within phytobiomes. Curr Opin Plant Biol. 2017;38:109–16.
Ponce de León I, Montesano M. Activation of defense mechanisms against pathogens in mosses and flowering plants. Int J Mol Sci. 2013;14:3178–200.
Fricker AM, Podlesny D, Fricke WF. What is new and relevant for sequencing-based microbiome research? A mini-review. J Adv Res. 2019;19:105–12.
Bragina A, Berg C, Müller H, Moser D, Berg G. Insights into functional bacterial diversity and its effects on Alpine bog ecosystem functioning. Sci Rep. 2013;3:1–8.
Taffner J, Erlacher A, Bragina A, Berg C, Moissl-Eichinger C, Berg G. What is the role of Archaea in plants? New insights from the vegetation of alpine bogs. MSphere. 2018;3(3):e00122-18.
Lu J, Breitwieser FP, Thielen P, Salzberg SL. Bracken: estimating species abundance in metagenomics data. PeerJ Comput Sci. 2017;3:e104.
Wood DE, Lu J, Langmead B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019;20:257.
Paulson JN, Stine OC, Bravo HC, Pop M. Differential abundance analysis for microbial marker-gene surveys. Nat Methods. 2013;10:1200–2.
Allaire J. RStudio: integrated development environment for R. Boston MA. 2012;770:394.
Chong J, Liu P, Zhou G, **a J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat Protoc. 2020;15:799–821.
Core Team R. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing; 2013.
Dhariwal A, Chong J, Habib S, King IL, Agellon LB, **a J. MicrobiomeAnalyst: a web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017;45:W180–8.
McMurdie PJ, Holmes S. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One. 2013;8(4):e61217.
Oksanen J, Kindt R, Legendre P, O’Hara B, Stevens MHH, Oksanen MJ, et al. The vegan package. Community Ecol Package. 2007;10:631–7.
Segata N, Izard J, Waldron L, Gevers D, Miropolsky L, Garrett WS, et al. Metagenomic biomarker discovery and explanation. Genome Biol. 2011;12:1–18.
Andrews S. FastQC: a quality control tool for high throughput sequence data. 2010.
Li D, Liu C-M, Luo R, Sadakane K, Lam T-W. MEGAHIT: an ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics. 2015;31:1674–6.
Hyatt D, Chen G-L, LoCascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics. 2010;11:119.
Li W, Godzik A. Cd-hit: a fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics. 2006;22:1658–9.
Buchfink B, **e C, Huson DH. Fast and sensitive protein alignment using DIAMOND. Nat Methods. 2015;12:59.
Huerta-Cepas J, Forslund K, Coelho LP, Szklarczyk D, Jensen LJ, Von Mering C, et al. Fast genome-wide functional annotation through orthology assignment by eggNOG-mapper. Mol Biol Evol. 2017;34:2115–22.
Huerta-Cepas J, Szklarczyk D, Heller D, Hernández-Plaza A, Forslund SK, Cook H, et al. eggNOG 5.0: a hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019;47:D309-14.
Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, et al. The sequence alignment/map format and SAMtools. Bioinformatics. 2009;25:2078–9.
Li H, Durbin R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics. 2010;26:589–95.
Jonsson V, Österlund T, Nerman O, Kristiansson E. Statistical evaluation of methods for identification of differentially abundant genes in comparative metagenomics. BMC Genomics. 2016;17:78.
Robinson MD, McCarthy DJ, Smyth GK. edgeR: a bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics. 2010;26:139–40.
Kanehisa M, Sato Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020;29:28–35.
Tu Q, Lin L, Cheng L, Deng Y, He Z. NCycDB: a curated integrative database for fast and accurate metagenomic profiling of nitrogen cycling genes. Bioinformatics. 2019;35:1040–8.
Zhang J, Buhe C, Yu D, Zhong H, Wei Y. Ammonia stress reduces antibiotic efflux but enriches horizontal gene transfer of antibiotic resistance genes in anaerobic digestion. Bioresour Technol. 2020;295:122191.
Alneberg J, Bjarnason BS, De Bruijn I, Schirmer M, Quick J, Ijaz UZ, et al. Binning metagenomic contigs by coverage and composition. Nat Methods. 2014;11:1144–6.
Kang DD, Li F, Kirton E, Thomas A, Egan R, An H, et al. MetaBAT 2: an adaptive binning algorithm for robust and efficient genome reconstruction from metagenome assemblies. PeerJ. 2019;7:e7359.
Wu Y-W, Simmons BA, Singer SW. MaxBin 2.0: an automated binning algorithm to recover genomes from multiple metagenomic datasets. Bioinformatics. 2016;32:605–7.
Wicaksono WA, Cernava T, Berg C, Berg G. Reconstruction of bacterial metagenome-assembled genome sequences from Alpine bog vegetation. Microbiol Resour Announc. 2020;9(35):e00821-20.
Sieber CM, Probst AJ, Sharrar A, Thomas BC, Hess M, Tringe SG, et al. Recovery of genomes from metagenomes via a dereplication, aggregation and scoring strategy. Nat Microbiol. 2018;3:836–43.
Parks DH, Imelfort M, Skennerton CT, Hugenholtz P, Tyson GW. CheckM: assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015;25:1043–55.
Bowers RM, Kyrpides NC, Stepanauskas R, Harmon-Smith M, Doud D, Reddy T, et al. Minimum information about a single amplified genome (MISAG) and a metagenome-assembled genome (MIMAG) of bacteria and archaea. Nat Biotechnol. 2017;35:725–31.
von Meijenfeldt FB, Arkhipova K, Cambuy DD, Coutinho FH, Dutilh BE. Robust taxonomic classification of uncharted microbial sequences and bins with CAT and BAT. Genome Biol. 2019;20:217.
Chen L, Zheng D, Liu B, Yang J, ** Q. VFDB 2016: hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 2016;44:D694–7.
Zhang H, Zhang Q, Song J, Zhang Z, Chen S, Long Z, et al. Tracking resistomes, virulence genes, and bacterial pathogens in long-term manure-amended greenhouse soils. J Hazard Mater. 2020;396:122618.
Asnicar F, Thomas AM, Beghini F, Mengoni C, Manara S, Manghi P, et al. Precise phylogenetic analysis of microbial isolates and genomes from metagenomes using PhyloPhlAn 3.0. Nat Commun. 2020;11:1–10.
Heberle H, Meirelles GV, da Silva FR, Telles GP, Minghim R. InteractiVenn: a web-based tool for the analysis of sets through Venn diagrams. BMC Bioinformatics. 2015;16:1–7.
Opelt K, Berg C, Berg G. The bryophyte genus Sphagnum is a reservoir for powerful and extraordinary antagonists and potentially facultative human pathogens. FEMS Microbiol Ecol. 2007;61:38–53.
Durazzi F, Sala C, Castellani G, Manfreda G, Remondini D, De Cesare A. Comparison between 16S rRNA and shotgun sequencing data for the taxonomic characterization of the gut microbiota. Sci Rep. 2021;11:1–10.
Taffner J, Cernava T, Erlacher A, Berg G. Novel insights into plant-associated archaea and their functioning in arugula (Eruca sativa Mill.). J Adv Res. 2019;19:39–48.
Regalado J, Lundberg DS, Deusch O, Kersten S, Karasov T, Poersch K, et al. Combining whole-genome shotgun sequencing and rRNA gene amplicon analyses to improve detection of microbe–microbe interaction networks in plant leaves. ISME J. 2020;14:2116–30.
Pedrotti E, Rydin H, Ingmar T, Hytteborn H, Turunen P, Granath G. Fine-scale dynamics and community stability in boreal peatlands: revisiting a fen and a bog in Sweden after 50 years. Ecosphere. 2014;5:1–24.
Chaudhary N, Miller PA, Smith B. Biotic and abiotic drivers of peatland growth and microtopography: a model demonstration. Ecosystems. 2018;21:1196–214.
Robroek BJ, Jassey VE, Payne RJ, Martí M, Bragazza L, Bleeker A, et al. Taxonomic and functional turnover are decoupled in European peat bogs. Nat Commun. 2017;8:1–9.
Carrell AA, Kolton M, Glass JB, Pelletier DA, Warren MJ, Kostka JE, et al. Experimental warming alters the community composition, diversity, and N2 fixation activity of peat moss (Sphagnum fallax) microbiomes. Glob Change Biol. 2019;25:2993–3004.
Compant S, Clément C, Sessitsch A. Plant growth-promoting bacteria in the rhizo-and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil Biol Biochem. 2010;42:669–78.
Alcaraz LD, Peimbert M, Barajas HR, Dorantes-Acosta AE, Bowman JL, Arteaga-Vázquez MA. Marchantia liverworts as a proxy to plants’ basal microbiomes. Sci Rep. 2018;8:1–12.
Lebeis SL, Paredes SH, Lundberg DS, Breakfield N, Gehring J, McDonald M, et al. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science. 2015;349:860–4.
Turner TR, Ramakrishnan K, Walshaw J, Heavens D, Alston M, Swarbreck D, et al. Comparative metatranscriptomics reveals kingdom level changes in the rhizosphere microbiome of plants. ISME J. 2013;7:2248–58.
Belova S, Pankratov T, Dedysh S. Bacteria of the genus Burkholderia as a typical component of the microbial community of Sphagnum peat bogs. Microbiology. 2006;75:90–6.
Kracht O, Gleixner G. Isotope analysis of pyrolysis products from Sphagnum peat and dissolved organic matter from bog water. Org Geochem. 2000;31:645–54.
Malmer N. Mineral nutrients in vegetation and surface layers of Sphagnum-dominated peat-forming systems. Adv Bryol. 1993;5:223–48.
Kraiser T, Gras DE, Gutiérrez AG, González B, Gutiérrez RA. A holistic view of nitrogen acquisition in plants. J Exp Bot. 2011;62:1455–66.
Moreau D, Bardgett RD, Finlay RD, Jones DL, Philippot L. A plant perspective on nitrogen cycling in the rhizosphere. Funct Ecol. 2019;33:540–52.
Berendsen RL, Pieterse CM, Bakker PA. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012;17:478–86.
Berg G, Smalla K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol Ecol. 2009;68:1–13.
Hügler M, Sievert SM. Beyond the Calvin cycle: autotrophic carbon fixation in the ocean. Annu Rev Mar Sci. 2011;3:261–89.
Lynn TM, Ge T, Yuan H, Wei X, Wu X, **ao K, et al. Soil carbon-fixation rates and associated bacterial diversity and abundance in three natural ecosystems. Microb Ecol. 2017;73:645–57.
Yuan H, Ge T, Chen C, O’Donnell AG, Wu J. Significant role for microbial autotrophy in the sequestration of soil carbon. Appl Environ Microbiol. 2012;78:2328–36.
Holland-Moritz H, Stuart J, Lewis LR, Miller S, Mack MC, McDaniel SF, et al. Novel bacterial lineages associated with boreal moss species. Environ Microbiol. 2018;20:2625–38.
Singleton CM, McCalley CK, Woodcroft BJ, Boyd JA, Evans PN, Hodgkins SB, et al. Methanotrophy across a natural permafrost thaw environment. ISME J. 2018;12:2544–58.
Fritz C, Lamers LP, Riaz M, van den Berg LJ, Elzenga TJ. Sphagnum mosses-masters of efficient N-uptake while avoiding intoxication. PLoS One. 2014;9:e79991.
Liu X, Koba K, Makabe A, Li X, Yoh M, Liu C. Ammonium first: natural mosses prefer atmospheric ammonium but vary utilization of dissolved organic nitrogen depending on habitat and nitrogen deposition. New Phytol. 2013;199:407–19.
Frigaard N-U, Dahl C. Sulfur metabolism in phototrophic sulfur bacteria. Adv Microb Physiol. 2008;54:103–200.
Widdel F. Growth of methanogenic bacteria in pure culture with 2-propanol and other alcohols as hydrogen donors. Appl Environ Microbiol. 1986;51:1056–62.
Zhu X, Campanaro S, Treu L, Seshadri R, Ivanova N, Kougias PG, et al. Metabolic dependencies govern microbial syntrophies during methanogenesis in an anaerobic digestion ecosystem. Microbiome. 2020;8:1–14.
Cenci U, Qiu H, Pillonel T, Cardol P, Remacle C, Colleoni C, et al. Host-pathogen biotic interactions shaped vitamin K metabolism in Archaeplastida. Sci Rep. 2018;8:1–14.
Hale MB, Blankenship RE, Fuller RC. Menaquinone is the sole quinone in the facultatively aerobic green photosynthetic bacterium Chloroflexus aurantiacus. Biochim Biophys Acta BBA-Bioenerg. 1983;723:376–82.
Krewing M, Stepanek JJ, Cremers C, Lackmann J-W, Schubert B, Müller A, et al. The molecular chaperone Hsp33 is activated by atmospheric-pressure plasma protecting proteins from aggregation. J R Soc Interface. 2019;16:20180966.
LeThanh H, Neubauer P, Hoffmann F. The small heat-shock proteins IbpA and IbpB reduce the stress load of recombinant Escherichia coli and delay degradation of inclusion bodies. Microb Cell Factories. 2005;4:6.
Berg G, Eberl L, Hartmann A. The rhizosphere as a reservoir for opportunistic human pathogenic bacteria. Environ Microbiol. 2005;7:1673–85.
Pinski A, Betekhtin A, Hupert-Kocurek K, Mur LA, Hasterok R. Defining the genetic basis of plant–endophytic bacteria interactions. Int J Mol Sci. 2019;20:1947.
Casadevall A. The pathogenic potential of a microbe. Msphere. 2017;2(1):e00015-17.
Lira F, Berg G, Martínez JL. Double-face meets the bacterial world: the opportunistic pathogen Stenotrophomonas maltophilia. Front Microbiol. 2017;8:2190.
Reinhold-Hurek B, Bünger W, Burbano CS, Sabale M, Hurek T. Roots sha** their microbiome: global hotspots for microbial activity. Annu Rev Phytopathol. 2015;53:403–24.
Acknowledgements
We thank Barbara Fetz, Anastasia Bragina and Melanie Obermeier (all Graz) for the technical support during the initial processing of the samples.
Funding
This study was financially supported by a grant to G.B. (grant no. E-1653100183) from the Austrian Science Foundation FWF. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
C.B. and G.B. designed and performed the sampling, and G.B. and T.C. designed the study. W.A.W. performed the research and analysed the bioinformatic data. W.A.W., T.C. and G.B. wrote the manuscript. All authors discussed results and commented on the manuscript at all stages. The author(s) read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
Rarefaction curves showing the number of metagenomic reads that were classified as bacterial sequences according to the Kraken2 classifier. Figure S2. Bacterial community clustering at phylum (a), class (b), order (c) and genus (d) level was visualized in two-dimensional Bray Curtis PCoA plots. Figure S3. Analysis of virulence gene composition of clinical and environmental bog isolates. Hierarchical clustering of clinical and environmental strains is based in the presence/absence of virulence genes in the genome. Figure S4. Venn diagrams showing the numbers of shared and unique virulence genes detected in clinical and environmental bog isolate genomes. The graph was generated using InteractiVenn [61]. Table S1. Description of vegetation field plots regarding habitat characteristics and plant coverage per plot. Table S2. List of the 12 plant species that were included to represent the vegetation of a Sphagnum-dominated bog ecosystem. Table S3. Detailed taxonomical classification, source of origin and NCBI accessions of clinical and environmental isolate genomes. Table S4. List of the KEGG orthologs that were significantly enriched between bryophytes and vascular plants. Table S5. List of the pathways and genes involved in nitrogen cycling that were significantly enriched in either bryophytes or vascular plants. Table S6. Detailed taxonomic classification, completeness, contamination values, and genome sizes of bacterial MAGs. Table S7. Abundance estimation of metagenome assembled genomes in each plant sample. Table S8. Prevalence of putative virulence factors as predicted for metagenome-assembled genomes (MAGs) using the virulence factor database (VFDB). Table S9. Putative virulence factors that were uniquely present in the MAGs generated from the bog ecosystem when compared to their closely related clinical isolates.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wicaksono, W.A., Cernava, T., Berg, C. et al. Bog ecosystems as a playground for plant–microbe coevolution: bryophytes and vascular plants harbour functionally adapted bacteria. Microbiome 9, 170 (2021). https://doi.org/10.1186/s40168-021-01117-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40168-021-01117-7