
Abstract
Background
Fusobacterium nucleatum is a gram-negative anaerobic species residing in the oral cavity and implicated in several inflammatory processes in the human body. Although F. nucleatum abundance is increased in inflammatory bowel disease subjects and is prevalent in colorectal cancer patients, the causal role of the bacterium in gastrointestinal disorders and the mechanistic details of host cell functions subversion are not fully understood.
Results
We devised a computational strategy to identify putative secreted F. nucleatum proteins (FusoSecretome) and to infer their interactions with human proteins based on the presence of host molecular mimicry elements. FusoSecretome proteins share similar features with known bacterial virulence factors thereby highlighting their pathogenic potential. We show that they interact with human proteins that participate in infection-related cellular processes and localize in established cellular districts of the host–pathogen interface. Our network-based analysis identified 31 functional modules in the human interactome preferentially targeted by 138 FusoSecretome proteins, among which we selected 26 as main candidate virulence proteins, representing both putative and known virulence proteins. Finally, six of the preferentially targeted functional modules are implicated in the onset and progression of inflammatory bowel diseases and colorectal cancer.
Conclusions
Overall, our computational analysis identified candidate virulence proteins potentially involved in the F. nucleatum—human cross-talk in the context of gastrointestinal diseases.
Similar content being viewed by others


Background
Fusobacterium nucleatum is a gram-negative anaerobic bacterium best known as a component of the oral plaque and a key pathogen in gingivitis and periodontitis [1]. It has also been isolated in several inflammatory processes in distinct body sites (e.g., endocarditis, septic arthritis, liver and brain abscesses) and implicated in adverse pregnancy outcomes (reviewed in [2]). Moreover, it has been demonstrated that F. nucleatum can adhere to and invade a variety of cell types, thereby inducing a pro-inflammatory response [3,4,5,6,7,8]. Recent work showed that (i) F. nucleatum is prevalent in colorectal cancer (CRC) patients [9,10,11] and (ii) its abundance is increased in new-onset Crohn’s disease (CD) subjects [12]. Interestingly, follow-up studies suggested a potential role of this bacterium in CRC tumorigenesis and tumor-immune evasion [13,14,15,16].
Despite these findings, a large fraction of F. nucleatum gene products are still uncharacterized. Moreover, to date, only a handful of pathogenic factors has been experimentally identified [17, 18] and protein interaction data between these factors and human proteins, which could inform on the molecular details underlying host-cell subversion mechanisms, are sparse [4, 16, 19]. Altogether, this underlines that a comprehensive view of the molecular details of the F. nucleatum—human cross-talk is currently missing.
How could F. nucleatum hijack human cells? Pathogens employ a variety of molecular strategies to reach an advantageous niche for survival. One of them consists of subverting host protein interaction networks. Indeed, they secrete and deliver factors such as toxic compounds, small peptides, and even proteins to target the host molecular networks. To achieve this, virulence factors often display structures resembling host components in form and function [20,21,22] to interact with host proteins, thus providing a benefit to the pathogen [23]. Such “molecular mimics” (e.g., targeting motifs, enzymatic activities, and protein–protein interaction elements) allow pathogens to enter the host cell and perturb cell pathways (e.g., [24,25,26]).
Over the years, several experimental approaches have been applied to identify protein–protein interactions (PPIs) between pathogens and their hosts providing new insights on the pathogen’s molecular invasion strategies. However, the vast majority of these systematic studies focused on viruses (e.g., [27,28,29]) and, to a lesser extent, on bacteria [30,31,32,33] and eukaryotic parasites [33, 34]. Indeed, as cellular pathogens have large genomes and complex life cycles, the experimental identification of virulence proteins and the large-scale map** of host-pathogen PPIs require a lot of effort and time [35, 36]. In this context, computational approaches have proved to be instrumental for the identification of putative pathogenic proteins (e.g., [37, 38]), the characterization of molecular mimics [23, 39, 40], and the inference of their interactions with host proteins (for a review see [41]).
Here, in order to gain new insights on the molecular cross-talk between F. nucleatum and the human host, we devised a computational strategy combining secretion prediction, protein–protein interaction inference, and protein interaction network analyses (Fig. 1). Doing so, we defined a secretome of the bacterium and the human proteins with which they interact based on the presence of mimicry elements. We identified the host cellular pathways that are likely perturbed by F. nucleatum including immune and infection response, homeostasis, cytoskeleton organization, and gene expression regulation. Interestingly, our results identify candidate virulence proteins, including the established Fap2 adhesin, and provide new insights underlying the putative causative role of F. nucleatum in colorectal cancer and inflammatory bowel diseases.
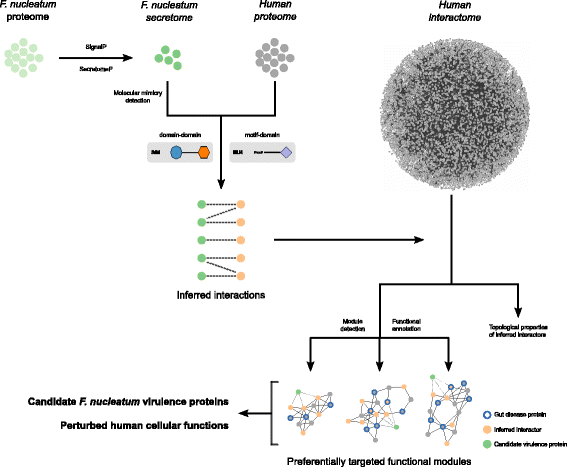
Flow strategy of our computational approach
Results
Prediction of F. nucleatum secreted proteome
Previous computational analyses highlighted that F. nucleatum has a reduced repertoire of secretion machinery [42, 43] meaning that it might exploit alternative “non-classical” translocation mechanisms to unleash virulence proteins. Thus, we sought to identify putative F. nucleatum secreted proteins by analyzing the 2046 protein sequences of the type species F. nucleatum subsp. nucleatum (strain ATCC 25586) proteome using two distinct algorithms: SignalP [44] for peptide-triggered secretion and SecretomeP [45] for leaderless protein secretion. While the SignalP algorithm predicted 61 F. nucleatum sequences being secreted via classical/regular secretion pathways, SecretomeP found 176 proteins as possibly secreted through non-classical routes. In total, we identified 237 putative secreted proteins in the F. nucleatum proteome (herein called “FusoSecretome”) (see Additional file 1: Table S1). Notably, we were able to correctly predict as secreted all the F. nucleatum virulence proteins known so far, namely FadA (FN0264), Fap2 (FN1449), RadD (FN1526), and the recently identified Aid1 adhesin (FN1253) [46]. This result underlines the relevance of secretion prediction to identify novel putative virulence proteins in the F. nucleatum proteome.
It has been shown that disorder propensity is an emerging hallmark of pathogenicity [47, 48]. As SecretomeP exploits protein disorder as a predicting feature, we analyzed the intrinsic disorder content of the FusoSecretome proteins identified by the SignalP algorithm only. We indeed observed a significantly higher disorder propensity of these proteins compared to the non-secreted proteins (P value = 1.9 × 10−4, Kolgomorov–Smirnov test, two-sided) (Fig. 2; Additional file 2: Figure S1; Additional file 3: Table S2), further reinforcing the possible role of the FusoSecretome in the infection/invasion process.
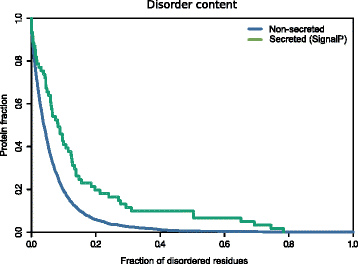
Disorder propensity of the FusoSecretome. SignalP-secreted proteins show a significantly higher fraction of disordered residues compared to non-secreted proteins (P value = 1.9 × 10−4, Kolgomorov–Smirnov test, two-sided)
To detect functional elements that can further contribute to F. nucleatum pathogenicity, we sought for the presence of globular domains in the FusoSecretome. We observed an enrichment of domains mainly belonging to the outer membrane beta-barrel protein superfamily (Table 1). Six out of the eight over-represented domains among the FusoSecretome proteins are also found in known virulence proteins of gram-negative bacteria [49] and are involved in adhesion, secretion, transport, and invasion. Altogether, these findings suggest that FusoSecretome proteins display features of known virulence proteins and can likely be involved in the cross-talk with the human host.
Inference of the FusoSecretome—human interaction network
Generally, pathogens employ a variety of molecular strategies to interfere with host-cell networks, controlling key functions such as plasma membrane and cytoskeleton dynamics, immune response, and cell death/survival. In particular, their proteins often carry a range of mimics, which resemble structures of the host at the molecular level, to “sneak” into host cells [20,21,22, 50].
Here, we focused on putative molecular mimicry events that can mediate the interaction with host proteins: (i) globular domains that occur in both FusoSecretome and the human proteome and (ii) known eukaryotic short linear motifs (SLiMs) found in FusoSecretome proteins. SLiMs are short stretches of 3–10 contiguous amino acids residues that often mediate transient PPIs and tend to bind with low affinity [51].
We first scanned the sequences of the FusoSecretome and human proteins for the presence of domains as defined by Pfam [52]. We identified 55 “host-like” domains in 50 FusoSecretome proteins out of 237, including several domains related to ribosomal proteins, aminopeptidases, and tetratricopeptide repeats (TPR) (Additional file 4: Table S3). Interestingly, 29 of these domains are also found in known bacterial binders of human proteins [30].
We next detected the occurrence of experimentally identified SLiMs gathered from the Eukaryotic Linear Motif (ELM) database [53]. As linear motifs are short and degenerate in sequence, SLiM detection is prone to over-prediction [54]. To reduce the number of false positives, we kept occurrences falling in conserved and disordered protein sequences (see the “Methods” section). Indeed, known functional SLiMs show a higher degree of conservation compared to surrounding residues [51] and are located in unstructured regions [55, 56]. In this way, we identified at least one putative mimicry SLiM in 139 FusoSecretome proteins. Most of the 57 different detected SLiMs represents binding sites such as motifs recognized by PDZ, SH3, and SH2 domains (Additional file 4: Table S3).
We exploited these putative mimicry events to infer the interaction with human proteins by using templates of domain–domain and SLiM–domain interactions (see the “Methods” section for further details). Doing so, we obtained 3744 interactions (1544 domain- and 2201 SLiM-mediated interactions, respectively) between 144 FusoSecretome, which we designated as “candidate virulence proteins,” and 934 human proteins (Additional file 5: Table S4 and Additional file 6: Table S5) designated as “human inferred interactors.”
In order to assess the reliability of the inferences, we evaluated the biological relevance of the putative human interactors by performing enrichment analyses of different orthogonal datasets using as a reference background all the proteins encoded by the human genome.
First, human proteins experimentally identified as binders or targets of bacterial and viral proteins are over-represented among the 934 inferred human interactors of the FusoSecretome proteins (415 proteins, 1.3-fold, P value = 1.61 × 10−11). Notably, the over-representation holds when bacterial and viral binders are considered separately (176 bacterial interactors, 1.1-fold, P value = 3.5 × 10−3 and 338 viral interactors, 1.5-fold, P value <2.2 × 10−16). This result is consistent with current knowledge on convergent targeting of host proteins by distinct pathogens [30, 33, 57, 58]. Second, according to the Human Proteins Atlas (see the “Methods” section), the vast majority of the inferred human interactors has been detected either in small intestine (652, 70%) or colorectal (671 proteins, 72%) tissues as well as in the saliva (673, 72%), confirming their presence in human body sites hosting F. nucleatum. Third, we assessed whether the inferred human interactors are implicated in gastrointestinal disorders by seeking for an over-representation of genes associated to such diseases (see the “Methods” section). Indeed, the human interactors of the FusoSecretome are enriched in (i) proteins identified in the human colon secretomes of colorectal cancer (CRC) tissue samples (3.5-fold, P value <2.2 × 10−16), (ii) proteins encoded by genes whose expression correlates with F. nucleatum abundance in CRC patients [13] (twofold, P value = 4 × 10−4), and (iii) genes associated with inflammatory bowel diseases (IBDs) (twofold, P value = 8 × 10−4). We obtained very similar enrichments by using a reduced statistical background corresponding to the interaction inference space (see the “Methods” section and Additional file 7: Supplementary Results).
Altogether, the results of these analyses highlight the relevance of the inferred human interactors as putative binders of FusoSecretome proteins and their potential implication in gut diseases, therefore validating the undertaken inference approach.
Functional role of the human proteins targeted by F. nucleatum
Globally, the inferred FusoSecretome human interactors are involved in several processes related to pathogen infection such as immune response and inflammation, response to stress, endocytosis as shown by the 137 significantly enriched Biological Processes Gene Ontology (GO) terms among their annotations (Table 2, Additional file 8: Table S6A). Similarly, the targeted human proteins are over-represented in 125 pathways (54 from KEGG and 71 from Reactome databases [59, 60]) involved in cell adhesion and signaling, extracellular matrix remodeling, immunity, response to infection, and cancer-related pathways (Table 2, Additional file 8: Table S6A). These human proteins are mainly localized in the extracellular space, plasma membrane, and at cell-cell junctions that represent the main districts involved in the initial encounter between a pathogen and the host, as indicated by the over-representation of 30 Cellular Component GO terms (Table 2, Additional file 8: Table S6A). A substantial fraction of these enriched functional categories is significantly over-represented when using the reduced statistical background as well (see the “Methods” section and Additional file 8: Table S6B). Overall, this indicates that our inferred interactions can participate in the F. nucleatum—human cross-talk.
F. nucleatum targets topologically important proteins in the host network
To gain a broader picture of the inferred interactions in the cellular context, we mapped the FusoSecretome human interactors on a binary human interactome built by gathering protein interactions data from both small-scale experiments and systematic screens reported in the literature (see the “Methods” section and Additional file 9: Table S7). Around 70% of the inferred human interactors (i.e., 663 proteins) are present in the human binary interactome. Interestingly, the human targeted proteins occupy topologically important positions in the interactome as shown by their significantly higher number of interactions and higher values of betweenness centrality compared to other network proteins (number of interactions: mean = 23 vs. 11, P value = 1.9 × 10−10; betweeness centrality: mean = 0.00078 vs. 0.00018, P value = 6.2 × 10−12; two-sided Mann–Whitney U test) (Fig. 3).
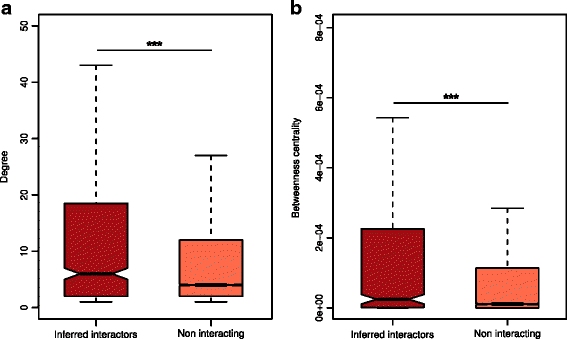
Topological properties of inferred human interactors in the human interactome. a Inferred human interactors have more interaction partners and b higher values of betweenness centrality compared to non-interacting proteins in the human interactome
The human interactome is composed of functional network modules, defined as group of proteins densely connected through their interactions and involved in the same biological process [61] (see the “Methods” section). We thus next investigated the 855 functional modules that we previously detected [62] using the OCG algorithm that decomposes a network into overlap** modules, based on modularity optimization [63] (Additional file 10: Table S8). A significant number of interactors participate in 2 or more of these functional units (259 proteins, 1.3-fold enrichment, P value = 1.4 × 10−7), indicating that the FusoSecretome tends to target multifunctional proteins in the human interactome [63]. Moreover, among the multifunctional inferred human interactors, we found an enrichment of extreme multifunctional proteins (52 interactors, twofold enrichment, P value = 1.0 × 10−5), which are defined as proteins involved in unrelated cellular functions and may represent candidate moonlighting proteins [64]. This suggests that the FusoSecretome might perturb multiple cellular pathways simultaneously by targeting preferentially a whole range of multifunctional proteins.
Functional subnetworks of the human interactome perturbed by F. nucleatum and identification of the main candidate virulence proteins
Based on their enrichment in inferred human interactors, 31 network modules (~4% of the 855 detected modules) are preferentially targeted by 138 distinct proteins of the FusoSecretome (Table 3). Targeted modules are involved in relevant processes such as immune response, cytoskeleton organization, cancer, and infection-related pathways (Table 3 and Additional file 11: Table S9). Moreover, proteins belonging to these modules are mainly localized in the extracellular space or in membranous structures (Table 3 and Additional file 11: Table S9), which represent important districts of the microbe-host interface. Interestingly, the enrichment of functional categories related to gene expression regulation (Additional file 11: Table S9) in several modules suggests novel potential host subversion mechanisms by F. nucleatum.
These modules are targeted on average by 50 FusoSecretome proteins (ranging from 2 to 104 per module) and the number of inferred host–pathogen interactions for each module varies considerably (Table 3). What are the main network perturbators among the FusoSecretome proteins? To quantify their impact on network modules based on the number of interactions, they have with each of them, we computed a Z score (see the “Methods” section, Additional file 12: Table S10). We considered the 26 FusoSecretome proteins having a perturbation Z score >2 in at least one module as main candidate virulence proteins. They consist in outer membrane proteins, enzymes, iron-binding proteins, and protein involved in transport (Table 4). Ten of them (38%) can perturb at least two distinct modules (Fig. 4a). Notably, we identified among the candidates, the known virulence protein Fap2 (FN1449) (Fig. 4b) that targets 4 modules, and a protein containing the MORN_2 domain (FN2118) (Fig. 4c) recently identified as a key element in actively invading F. nucleatum species [65], which perturbs 6 modules. On the other hand, 25 preferentially targeted modules are perturbed by at least two candidate virulence proteins, Module 78 involved in immune response being the most potentially subverted (Fig. 4a).
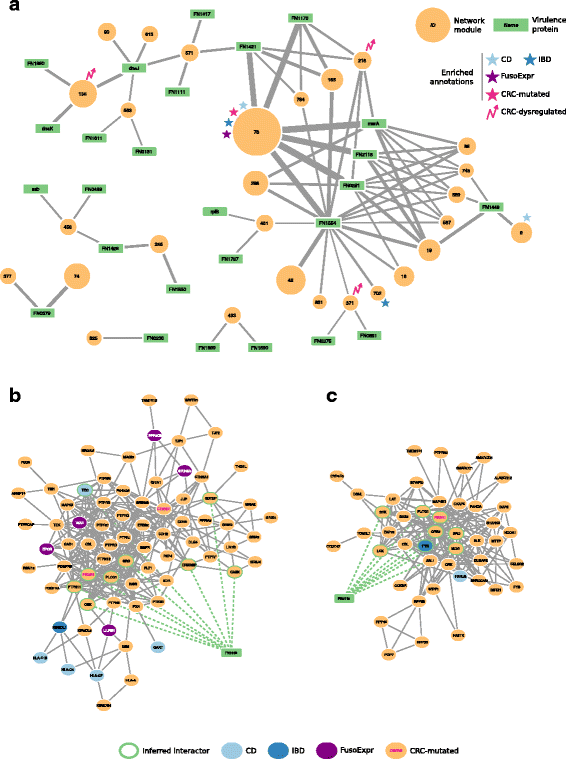
Interaction network between FusoSecretome candidate virulence proteins and preferentially targeted modules. a Candidate virulence proteins are depicted as green rectangular nodes labeled with respective gene symbol, whereas network modules as orange circles, whose size is proportional to the number of proteins belonging to each module and are labeled with the corresponding identifier. Edge width is proportional to the number of inferred interactions of a virulence protein with a given module. Network modules enriched in gut-related disease gene sets are labeled with symbols of different colors (i.e., light blue star: Crohn’s disease, CD; dark blue star: Inflammatory bowel disease, IBD; violet star: genes whose expression correlates with F. nucleatum abundance in colorectal cancer patients, FusoExpr; rose star: genes mutated in colorectal cancer, CRC-mutated; rose zig-zag arrow: dysregulated expression during colorectal cancer progression, CRC-dysregulated). b The protein Fap2 (FN1449) interacts with 9 proteins (nodes with a green border) of Module 9 and c the MORN2 domain containing protein (FN2118) interacts with 8 proteins in Module 89
F. nucleatum and gut diseases from a network perspective
Among the 855 network modules detected in the human interactome, 38 are enriched in genes involved in at least one gut disease (i.e., CRC and IBDs, see the “Methods” section). Interestingly, 27 of them (i.e., 71%) are targeted by at least one FusoSecretome protein, among which 3 contain a statistically significant fraction of inferred human interactors (Fig. 5). Notably, Module 78, involved in immune response, is enriched in genes associated to inflammatory bowel diseases (IBDs) (28 proteins, 5.2-fold enrichment, P value = 4.78 × 10−4) as well as in CD-specific (9 proteins, 13.4-fold enrichment, P value = 2.52 × 10−3) and CRC-mutated (11 proteins, fourfold enrichment, P value = 1.46 × 10−2) genes. Moreover, it is enriched in genes whose expression correlates with F. nucleatum abundance in CRC patients (24 proteins, 3.7-fold enrichment, P value = 3.35 × 10−4). This module is targeted by several main candidate virulence proteins, including a hemolysin (FN0291), an outer membrane protein (FN1554) and the MORN_2 domain containing protein (FN21118) (Fig. 4a), which therefore, may play critical roles in these diseases. IBD genes are also enriched in Module 702 (5 proteins, 11-fold enrichment, P value = 2.13 × 10−2), whose proteins participate in Jak-STAT signaling, whereas CD-specific genes are over-represented in Module 9 implicated in immunity (5 proteins, 28-fold enrichment, P value = 2.52 × 10−3). Interestingly, Module 9 is specifically perturbed by Fap2 (FN1449) (Fig. 4b), which is known to modulate the host immune response.
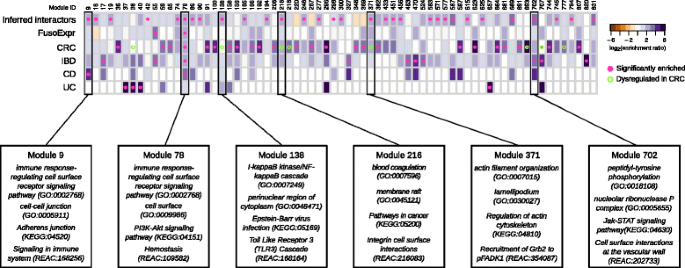
Enrichment of FusoSecretome inferred human interactors and gut disease related proteins in network modules. Each column of the heatmap represents a module. The color of the cells corresponds to the log-transformed enrichment ratio. Pink circles indicate enriched sets. Modules showing a significant dysregulation in CRC progression are highlighted by an empty circle with green border. For the six modules showing an enrichment in inferred interactions and at least in one of gut disease related proteins, the most representative functions are reported. FusoExpr: genes whose expression correlates with F. nucleatum abundance in CRC patients; CRC: genes mutated in colorectal cancer samples; IBD: genes associated to inflammatory bowel disease; CD and UC: genes specifically associated to Crohn’s disease and ulcerative colitis respectively
Three other modules enriched in inferred human interactors show a significant dysregulation of the expression of their constituent proteins during CRC progression [66] and are implicated in infection-response pathways and cytoskeleton organization (Fig. 5). In particular, two of these modules (Modules 138 and 216) show significant and specific upregulation in stage II, whereas the third (Module 371) is significantly upregulated in normal and stage II samples. Overall, these results indicate that F. nucleatum could contribute to the onset and progression of IBDs and CRC by perturbing some of the underlying network modules.
Comparison with additional bacterial strains
We applied our computational approach on the recently released proteomes of 6 actively invading Fusobacteria strains isolated from biopsy tissues [8, 65] (i.e., 4 F. nucleatum subspecies and 2 F. periodonticum strains), and the proteome of E. coli K-12 as a “control strain” (see Additional file 7: Supplementary Results, Table S12). We found that the secretomes of these 7 bacteria share common features (i.e., disorder propensity, enriched domains, host-like domain and mimicry SLiM content) with the FusoSecretome (Additional file 7: Table S13–S15 and Figure. S2–S8). However, we observed a moderate overlap in terms of inferred interactors, enriched functions and preferentially targeted network modules (Additional File 7: Table S16–S18), and a modest concordance in term of network module perturbators (Additional File 7: Table S19).
The results of these analyses suggest that, on the one hand, actively invading Fusobacteria species share common mechanisms to interact with host cell and, on the other hand, are consistent with the fact that F. nucleatum is an unusual heterogeneous species both at the genotypic and phenotypic level [8, 65, 67]. Finally, the commonalities between the FusoSecretome and the E. coli K-12 secreted proteins are not surprising, since previous work showed that E. coli K-12 carries cryptic genes coding for virulence factors [68], whose expression is activated by mutations in the histone-like protein HU, which convert this established commensal strain to an invasive species in intestinal cells [69].
Discussion
Over the years, it has been shown that F. nucleatum can adhere and invade human cells triggering a pro-inflammatory response. Nevertheless, the current knowledge on the molecular players underlying the F. nucleatum—human cross-talk is still limited.
For this reason, we carried out a computational study to identify F. nucleatum putative secreted factors (FusoSecretome) that can interact with human proteins.
The originality of our study is manifold compared to previous work. First, we used secretion prediction to identify potential F. nucleatum proteins that can be present at the microbe–host interface. Second, we exploited both domain–domain and domain–motif templates to infer interactions with human proteins. Earlier works, including one on F. nucleatum, chiefly applied homology-based methods for interaction inference with host proteins (e.g., [70,71,72,73]). To our knowledge, domain–motif templates have been only exploited so far to infer or to resolve human–virus protein interaction networks [105]). We kept only likely direct (i.e., binary) interactions according to the experimental detection method [106] and mapped protein identifiers to UniprotKB IDs. Given the redundancy among SwissProt and TrEMBL entries, protein sequences were clustered using the CD-HIT algorithm [107]. SwissProt/TrEMBL pairs at 95% identity were considered as the same protein: interactions of TrEMBL protein were assigned to the SwissProt protein. As a result, we obtained a human binary interactome containing 74,388 interactions between 12,865 proteins (February 2013).
We detected 855 network modules detected using the Overlap** Cluster Generator algorithm [63]. Modules were functionally annotated by assessing the enrichment of Gene Ontology (GO) biological process and cellular component terms [103], and cellular pathways from KEGG [59] and Reactome [60]. Enrichment P values were computed using the R package gProfileR [102] and corrected for multiple testing with the Benjamini–Hochberg procedure (significance threshold = 0.025) and annotated proteins in the human interactome were used as statistical background. Similarly, the over-representation of inferred human interactors and gut disease gene sets in network modules of the human interactome was assessed using a one-sided Fisher’s exact test followed by Benjamini–Hochberg multiple testing correction (significance threshold = 0.025).
Network module perturbation Z score
We devised a score to quantify the contribution of F. nucleatum secreted proteins to the perturbation of a network module through their inferred interactions. We defined the perturbation Z score for each F. nucleatum protein f interacting with at least one protein in module m as follows:
Where x f , m is the number of inferred interactions of the protein f with module m, Z f,m is the perturbation Z score of the protein f in the module m, μm, and σ m are the mean of the inferred interaction values and their standard deviation in the module m, respectively.
Network modules significantly dysregulated during CRC progression
The 77 network modules showing a significant dysregulation during CRC progression were taken from our previous work [66], in which we devised a computational method that combines quantitative proteomic profiling of TCGA CRC samples, protein interaction network, and statistical analysis to identify significantly dysregulated cellular functions during cancer progression.
References
Moore WE, Moore LV. The bacteria of periodontal diseases. Periodontol. 1994;5:66–77.
Han YW. Fusobacterium nucleatum: a commensal-turned pathogen. Curr Opin Microbiol. 2015;23C:141–7.
Dharmani P, Strauss J, Ambrose C, Allen-Vercoe E, Chadee K. Fusobacterium nucleatum infection of colonic cells stimulates MUC2 mucin and tumor necrosis factor alpha. Infect Immun. 2011;79:2597–607.
Fardini Y, Wang X, Témoin S, Nithianantham S, Lee D, Shoham M, et al. Fusobacterium nucleatum adhesin FadA binds vascular endothelial cadherin and alters endothelial integrity. Mol Microbiol. 2011;82:1468–80.
Gursoy UK, Könönen E, Uitto V-J. Intracellular replication of fusobacteria requires new actin filament formation of epithelial cells. APMIS Acta Pathol Microbiol Immunol Scand. 2008;116:1063–70.
Han YW, Shi W, Huang GT, Kinder Haake S, Park NH, Kuramitsu H, et al. Interactions between periodontal bacteria and human oral epithelial cells: Fusobacterium nucleatum adheres to and invades epithelial cells. Infect Immun. 2000;68:3140–6.
Quah SY, Bergenholtz G, Tan KS. Fusobacterium nucleatum induces cytokine production through Toll-like-receptor-independent mechanism. Int Endod J. 2014;47:550–9.
Strauss J, Kaplan GG, Beck PL, Rioux K, Panaccione R, Devinney R, et al. Invasive potential of gut mucosa-derived Fusobacterium nucleatum positively correlates with IBD status of the host. Inflamm Bowel Dis. 2011;17:1971–8.
Castellarin M, Warren RL, Freeman JD, Dreolini L, Krzywinski M, Strauss J, et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012;22:299–306.
Kostic AD, Gevers D, Pedamallu CS, Michaud M, Duke F, Earl AM, et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012;22:292–8.
McCoy AN, Araújo-Pérez F, Azcárate-Peril A, Yeh JJ, Sandler RS, Keku TO. Fusobacterium is associated with colorectal adenomas. PLoS One. 2013;8:e53653.
Gevers D, Kugathasan S, Denson LA, Vázquez-Baeza Y, Van Treuren W, Ren B, et al. The treatment-naive microbiome in new-onset Crohn’s disease. Cell Host Microbe. 2014;15:382–92.
Kostic AD, Chun E, Robertson L, Glickman JN, Gallini CA, Michaud M, et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe. 2013;14:207–15.
Mima K, Nishihara R, Qian ZR, Cao Y, Sukawa Y, Nowak JA, et al. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut. 2016;65(12):1973–1980.
Mima K, Sukawa Y, Nishihara R, Qian ZR, Yamauchi M, Inamura K, et al. Fusobacterium nucleatum and T cells in colorectal carcinoma. JAMA Oncol. 2015;1:653–61.
Rubinstein MR, Wang X, Liu W, Hao Y, Cai G, Han YW. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe. 2013;14:195–206.
Han YW, Ikegami A, Rajanna C, Kawsar HI, Zhou Y, Li M, et al. Identification and characterization of a novel adhesin unique to oral fusobacteria. J Bacteriol. 2005;187:5330–40.
Kaplan CW, Ma X, Paranjpe A, Jewett A, Lux R, Kinder-Haake S, et al. Fusobacterium nucleatum outer membrane proteins Fap2 and RadD induce cell death in human lymphocytes. Infect Immun. 2010;78:4773–8.
Gur C, Ibrahim Y, Isaacson B, Yamin R, Abed J, Gamliel M, et al. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity. 2015;42:344–55.
Elde NC, Malik HS. The evolutionary conundrum of pathogen mimicry. Nat Rev Microbiol. 2009;7:787–97.
Davey NE, Travé G, Gibson TJ. How viruses hijack cell regulation. Trends Biochem Sci. 2011;36:159–69.
Via A, Uyar B, Brun C, Zanzoni A. How pathogens use linear motifs to perturb host cell networks. Trends Biochem Sci. 2015;40:36–48.
Ludin P, Nilsson D, Mäser P. Genome-wide identification of molecular mimicry candidates in parasites. PLoS One. 2011;6:e17546.
Baxt LA, Garza-Mayers AC, Goldberg MB. Bacterial subversion of host innate immune pathways. Science. 2013;340:697–701.
Rudel T, Kepp O, Kozjak-Pavlovic V. Interactions between bacterial pathogens and mitochondrial cell death pathways. Nat Rev Microbiol. 2010;8:693–705.
Haglund CM, Welch MD. Pathogens and polymers: microbe-host interactions illuminate the cytoskeleton. J Cell Biol. 2011;195:7–17.
Uetz P, Dong Y-A, Zeretzke C, Atzler C, Baiker A, Berger B, et al. Herpesviral protein networks and their interaction with the human proteome. Science. 2006;311:239–42.
de Chassey B, Navratil V, Tafforeau L, Hiet MS, Aublin-Gex A, Agaugué S, et al. Hepatitis C virus infection protein network. Mol Syst Biol. 2008;4:230.
Jäger S, Cimermancic P, Gulbahce N, Johnson JR, McGovern KE, Clarke SC, et al. Global landscape of HIV-human protein complexes. Nature. 2012;481:365–70.
Dyer MD, Neff C, Dufford M, Rivera CG, Shattuck D, Bassaganya-Riera J, et al. The human-bacterial pathogen protein interaction networks of Bacillus anthracis, Francisella tularensis, and Yersinia pestis. PLoS One. 2010;5:e12089.
Blasche S, Arens S, Ceol A, Siszler G, Schmidt MA, Häuser R, et al. The EHEC-host interactome reveals novel targets for the translocated intimin receptor. Sci Rep. 2014;4:7531.
Mirrashidi KM, Elwell CA, Verschueren E, Johnson JR, Frando A, Von Dollen J, et al. Global map** of the inc-human interactome reveals that retromer restricts chlamydia infection. Cell Host Microbe. 2015;18:109–21.
Weßling R, Epple P, Altmann S, He Y, Yang L, Henz SR, et al. Convergent targeting of a common host protein-network by pathogen effectors from three kingdoms of life. Cell Host Microbe. 2014;16:364–75.
Ahn H-J, Kim S, Kim H-E, Nam H-W. Interactions between secreted GRA proteins and host cell proteins across the paratitophorous vacuolar membrane in the parasitism of Toxoplasma gondii. Korean J Parasitol. 2006;44:303–12.
Wu H-J, Wang AH-J, Jennings MP. Discovery of virulence factors of pathogenic bacteria. Curr Opin Chem Biol. 2008;12:93–101.
Vidal M, Cusick ME, Barabási A-L. Interactome networks and human disease. Cell. 2011;144:986–98.
McDermott JE, Corrigan A, Peterson E, Oehmen C, Niemann G, Cambronne ED, et al. Computational prediction of type III and IV secreted effectors in gram-negative bacteria. Infect Immun. 2011;79:23–32.
Wang Y, Wei X, Bao H, Liu S-L. Prediction of bacterial type IV secreted effectors by C-terminal features. BMC Genomics. 2014;15:50.
Garamszegi S, Franzosa EA, **a Y. Signatures of pleiotropy, economy and convergent evolution in a domain-resolved map of human-virus protein-protein interaction networks. PLoS Pathog. 2013;9:e1003778.
Ruhanen H, Hurley D, Ghosh A, O’Brien KT, Johnston CR, Shields DC. Potential of known and short prokaryotic protein motifs as a basis for novel peptide-based antibacterial therapeutics: a computational survey. Front Microbiol. 2014;5:4.
Arnold R, Boonen K, Sun MGF, Kim PM. Computational analysis of interactomes: current and future perspectives for bioinformatics approaches to model the host-pathogen interaction space. Methods. 2012;57:508–18.
Kapatral V, Anderson I, Ivanova N, Reznik G, Los T, Lykidis A, et al. Genome sequence and analysis of the oral bacterium Fusobacterium nucleatum strain ATCC 25586. J Bacteriol. 2002;184:2005–18.
Desvaux M, Khan A, Beatson SA, Scott-Tucker A, Henderson IR. Protein secretion systems in Fusobacterium nucleatum: genomic identification of Type 4 piliation and complete type V pathways brings new insight into mechanisms of pathogenesis. Biochim Biophys Acta. 2005;1713:92–112.
Petersen TN, Brunak S, von Heijne G, Nielsen H. SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat Methods. 2011;8:785–6.
Bendtsen JD, Kiemer L, Fausbøll A, Brunak S. Non-classical protein secretion in bacteria. BMC Microbiol. 2005;5:58.
Kaplan A, Kaplan CW, He X, McHardy I, Shi W, Lux R. Characterization of aid1, a novel gene involved in Fusobacterium nucleatum interspecies interactions. Microb Ecol. 2014;68:379–87.
Marín M, Uversky VN, Ott T. Intrinsic disorder in pathogen effectors: protein flexibility as an evolutionary hallmark in a molecular arms race. Plant Cell. 2013;25:3153–7.
Xue B, Blocquel D, Habchi J, Uversky AV, Kurgan L, Uversky VN, et al. Structural disorder in viral proteins. Chem Rev. 2014;114:6880–911.
Chen L, **ong Z, Sun L, Yang J, ** Q. VFDB 2012 update: toward the genetic diversity and molecular evolution of bacterial virulence factors. Nucleic Acids Res. 2012;40:D641–5.
Dean P. Functional domains and motifs of bacterial type III effector proteins and their roles in infection. FEMS Microbiol Rev. 2011;35:1100–25.
Davey NE, Van Roey K, Weatheritt RJ, Toedt G, Uyar B, Altenberg B, et al. Attributes of short linear motifs. Mol BioSyst. 2012;8:268–81.
Punta M, Coggill PC, Eberhardt RY, Mistry J, Tate J, Boursnell C, et al. The Pfam protein families database. Nucleic Acids Res. 2012;40:D290–301.
Dinkel H, Michael S, Weatheritt RJ, Davey NE, Van Roey K, Altenberg B, et al. ELM–the database of eukaryotic linear motifs. Nucleic Acids Res. 2012;40:D242–51.
Edwards RJ, Palopoli N. Computational prediction of short linear motifs from protein sequences. Methods Mol Biol. 2015;1268:89–141.
Fuxreiter M, Tompa P, Simon I. Local structural disorder imparts plasticity on linear motifs. Bioinformatics. 2007;23:950–6.
Edwards RJ, Davey NE, O’Brien K, Shields DC. Interactome-wide prediction of short, disordered protein interaction motifs in humans. Mol BioSyst. 2012;8:282–95.
Mukhtar MS, Carvunis A-R, Dreze M, Epple P, Steinbrenner J, Moore J, et al. Independently evolved virulence effectors converge onto hubs in a plant immune system network. Science. 2011;333:596–601.
Durmuş Tekir S, Cakir T, Ulgen KÖ. Infection strategies of bacterial and viral pathogens through pathogen-human protein-protein interactions. Front Microbiol. 2012;3:46.
Kanehisa M, Goto S, Sato Y, Furumichi M, Tanabe M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res. 2012;40:D109–14.
Croft D, O’Kelly G, Wu G, Haw R, Gillespie M, Matthews L, et al. Reactome: a database of reactions, pathways and biological processes. Nucleic Acids Res. 2011;39:D691–7.
Hartwell LH, Hopfield JJ, Leibler S, Murray AW. From molecular to modular cell biology. Nature. 1999;402:C47–52.
Chapple CE, Robisson B, Spinelli L, Guien C, Becker E, Brun C. Extreme multifunctional proteins identified from a human protein interaction network. Nat Commun. 2015;6:7412.
Becker E, Robisson B, Chapple CE, Guénoche A, Brun C. Multifunctional proteins revealed by overlap** clustering in protein interaction network. Bioinformatics. 2012;28:84–90.
Chapple CE, Brun C. Redefining protein moonlighting. Oncotarget. 2015;6:16812–3.
Manson McGuire A, Cochrane K, Griggs AD, Haas BJ, Abeel T, Zeng Q, et al. Evolution of invasion in a diverse set of Fusobacterium species. mBio. 2014;5:e01864.
Zanzoni A, Brun C. Integration of quantitative proteomics data and interaction networks: Identification of dysregulated cellular functions during cancer progression. Methods. 2016;93:103–9.
Strauss J, White A, Ambrose C, McDonald J, Allen-Vercoe E. Phenotypic and genotypic analyses of clinical Fusobacterium nucleatum and Fusobacterium periodonticum isolates from the human gut. Anaerobe. 2008;14:301–9.
Kar S, Edgar R, Adhya S. Nucleoid remodeling by an altered HU protein: Reorganization of the transcription program. Proc Natl Acad Sci U S A. 2005;102:16397–402.
Koli P, Sudan S, Fitzgerald D, Adhya S, Kar S. Conversion of commensal Escherichia coli K-12 to an invasive form via expression of a mutant histone-like protein. MBio. 2011;2(5). doi:10.1128/mBio.00182-11.
Tyagi N, Krishnadev O, Srinivasan N. Prediction of protein-protein interactions between Helicobacter pylori and a human host. Mol BioSyst. 2009;5:1630–5.
Doolittle JM, Gomez SM. Map** protein interactions between Dengue virus and its human and insect hosts. PLoS Negl Trop Dis. 2011;5:e954.
Schleker S, Garcia-Garcia J, Klein-Seetharaman J, Oliva B. Prediction and comparison of Salmonella-human and Salmonella-Arabidopsis interactomes. Chem Biodivers. 2012;9:991–1018.
Kumar A, Thotakura PL, Tiwary BK, Krishna R. Target identification in Fusobacterium nucleatum by subtractive genomics approach and enrichment analysis of host-pathogen protein-protein interactions. BMC Microbiol. 2016;16:84.
Evans P, Dampier W, Ungar L, Tozeren A. Prediction of HIV-1 virus-host protein interactions using virus and host sequence motifs. BMC Med Genet. 2009;2:27.
Hagai T, Azia A, Babu MM, Andino R. Use of host-like peptide motifs in viral proteins is a prevalent strategy in host-virus interactions. Cell Rep. 2014;7:1729–39.
Christie-Oleza JA, Piña-Villalonga JM, Bosch R, Nogales B, Armengaud J. Comparative proteogenomics of twelve Roseobacter exoproteomes reveals different adaptive strategies among these marine bacteria. Mol Cell Proteomics. 2012;11:M111.013110.
Tjalsma H, Lambooy L, Hermans PW, Swinkels DW. Shedding & shaving: disclosure of proteomic expressions on a bacterial face. Proteomics. 2008;8:1415–28.
Pérez-Cruz C, Delgado L, López-Iglesias C, Mercade E. Outer-inner membrane vesicles naturally secreted by gram-negative pathogenic bacteria. PLoS One. 2015;10:e0116896.
Chodavarapu S, Felczak MM, Kaguni JM. Two forms of ribosomal protein L2 of Escherichia coli that inhibit DnaA in DNA replication. Nucleic Acids Res. 2011;39:4180–91.
Jostins L, Ripke S, Weersma RK, Duerr RH, McGovern DP, Hui KY, et al. Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature. 2012;491:119–24.
Cusick ME, Yu H, Smolyar A, Venkatesan K, Carvunis A-R, Simonis N, et al. Literature-curated protein interaction datasets. Nat Methods. 2009;6:39–46.
Koegl M, Uetz P. Improving yeast two-hybrid screening systems. Brief Funct Genomic Proteomic. 2007;6:302–12.
Navratil V, de Chassey B, Combe CR, Lotteau V. When the human viral infectome and diseasome networks collide: towards a systems biology platform for the aetiology of human diseases. BMC Syst Biol. 2011;5:13.
Bierne H, Hamon M, Cossart P. Epigenetics and bacterial infections. Cold Spring Harb Perspect Med. 2012;2:a010272.
Zhang B, Wang J, Wang X, Zhu J, Liu Q, Shi Z, et al. Proteogenomic characterization of human colon and rectal cancer. Nature. 2014;513:382–7.
Sadanandam A, Lyssiotis CA, Homicsko K, Collisson EA, Gibb WJ, Wullschleger S, et al. A colorectal cancer classification system that associates cellular phenotype and responses to therapy. Nat Med. 2013;19:619–25.
Desousaemelo F, Wang X, Jansen M, Fessler E, Trinh A, de Rooij LPMH, et al. Poor-prognosis colon cancer is defined by a molecularly distinct subtype and develops from serrated precursor lesions. Nat Med 2013;19:614–618.
UniProt Consortium. Reorganizing the protein space at the Universal Protein Resource (UniProt). Nucleic Acids Res. 2012;40:D71–5.
Ward JJ, McGuffin LJ, Bryson K, Buxton BF, Jones DT. The DISOPRED server for the prediction of protein disorder. Bioinformatics. 2004;20:2138–9.
Dosztányi Z, Csizmok V, Tompa P, Simon I. IUPred: web server for the prediction of intrinsically unstructured regions of proteins based on estimated energy content. Bioinformatics. 2005;21:3433–4.
Linding R, Jensen LJ, Diella F, Bork P, Gibson TJ, Russell RB. Protein disorder prediction: implications for structural proteomics. Structure. 2003;11:1453–9.
Li W, Cowley A, Uludag M, Gur T, McWilliam H, Squizzato S, et al. The EMBL-EBI bioinformatics web and programmatic tools framework. Nucleic Acids Res. 2015;43:W580–4.
Davey NE, Haslam NJ, Shields DC, Edwards RJ. SLiMSearch 2.0: biological context for short linear motifs in proteins. Nucleic Acids Res. 2011;39:W56–60.
Davey NE, Shields DC, Edwards RJ. SLiMDisc: short, linear motif discovery, correcting for common evolutionary descent. Nucleic Acids Res. 2006;34:3546–54.
Stein A, Céol A, Aloy P. 3did: identification and classification of domain-based interactions of known three-dimensional structure. Nucleic Acids Res. 2011;39:D718–23.
Weatheritt RJ, Luck K, Petsalaki E, Davey NE, Gibson TJ. The identification of short linear motif-mediated interfaces within the human interactome. Bioinforma Oxf Engl. 2012;28:976–82.
Weatheritt RJ, Jehl P, Dinkel H, Gibson TJ. iELM—a web server to explore short linear motif-mediated interactions. Nucleic Acids Res. 2012;40:W364–9.
Navratil V, de Chassey B, Meyniel L, Delmotte S, Gautier C, André P, et al. VirHostNet: a knowledge base for the management and the analysis of proteome-wide virus-host interaction networks. Nucleic Acids Res. 2009;37:D661–8.
Uhlén M, Fagerberg L, Hallström BM, Lindskog C, Oksvold P, Mardinoglu A, et al. Tissue-based map of the human proteome. Science. 2015;347:1260419.
de Wit M, Kant H, Piersma SR, Pham TV, Mongera S, van Berkel MPA, et al. Colorectal cancer candidate biomarkers identified by tissue secretome proteome profiling. J Proteome. 2014;99:26–39.
An O, Pendino V, D’Antonio M, Ratti E, Gentilini M, Ciccarelli FD. NCG 4.0: the network of cancer genes in the era of massive mutational screenings of cancer genomes. Database J Biol Databases Curation. 2014;2014:bau015.
Reimand J, Arak T, Adler P, Kolberg L, Reisberg S, Peterson H, Vilo J. g:Profiler-a web server for functional interpretation of gene lists (2016 update). Nucleic Acids Res. 2016;44(W1):W83–9.
The Gene Ontology Consortium. The gene ontology in 2010: extensions and refinements. Nucleic Acids Res. 2010;38:D331–5.
Aranda B, Blankenburg H, Kerrien S, Brinkman FSL, Ceol A, Chautard E, et al. PSICQUIC and PSISCORE: accessing and scoring molecular interactions. Nat Methods. 2011;8:528–9.
Rolland T, Taşan M, Charloteaux B, Pevzner SJ, Zhong Q, Sahni N, et al. A proteome-scale map of the human interactome network. Cell. 2014;159:1212–26.
Rual J-F, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, et al. Towards a proteome-scale map of the human protein-protein interaction network. Nature. 2005;437:1173–8.
Fu L, Niu B, Zhu Z, Wu S, Li W. CD-HIT: accelerated for clustering the next-generation sequencing data. Bioinformatics. 2012;28:3150–2.
Acknowledgements
The authors would like to thank the members of the TAGC laboratory for fruitful discussion, Anaïs Baudot (I2M, CNRS, France) for critically reading the first draft of the manuscript, Aleksandar Kostic (Broad Institute, USA) for kindly providing the list of human genes whose expression correlates with F. nucleatum abundance in colorectal cancer patients, and Henrik Nielsen (DTU Bioinformatics, Denmark) for assistance in running SecretomeP predictions. AZ is grateful to Coralie, Olivia, and Claire for their constant support.
Funding
The project leading to this publication has received funding from Excellence Initiative of Aix-Marseille University—A*MIDEX, a French “Investissements d’Avenir” program, to CB, and was partially supported by the French ‘Plan Cancer 2009–2013’ program (Systems Biology call, A12171AS). The funding organizations had no role in the design of the study and collection, analysis, interpretation of data, and in writing the manuscript.
Availability of data and materials
All data generated or analyzed on the ATCC 25586 strain are included in this published article (and its Additional files). All other data are available from the corresponding author on reasonable request.
Author information
Authors and Affiliations
Contributions
AZ conceived the study, designed and performed the experiments, analyzed the data, and wrote the manuscript. LS performed the experiments and analyzed the data. SB performed the experiments. CB designed the experiments, analyzed the data, and wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study is based on publicly available datasets only. Thus, no ethical approval is needed/applicable nor is consent from any participants.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1: Table S1.
List of the F. nucleatum proteins predicted as secreted (FusoSecretome). (XLSX 61 kb)
Additional file 2: Figure S1.
Assessment of the disorder propensity of the FusoSecretome proteins (SignalP prediction) with additional prediction algorithms. (PDF 120 kb)
Additional file 3: Table S2.
Disorder content of FusoSecretome proteins (SignalP prediction) as determined by different algorithms. (XLSX 52 kb)
Additional file 4: Table S3.
List of detected mimicry elements used for protein interaction inference between FusoSecretome and human proteins. (XLSX 53 kb)
Additional file 5: Table S4.
Inferred interactions between FusoSecretome and human proteins. (XLSX 116 kb)
Additional file 6: Table S5.
List of the FusoSecretome inferred human interactors and their annotations. (XLSX 77 kb)
Additional file 7:
Supplementary Information. Supplementary results, tables and figures. (PDF 1670 kb)
Additional file 8: Table S6.
Functional annotations significantly enriched among inferred interactions. (XLSX 51 kb)
Additional file 9: Table S7.
The human binary interactome used in this study. (XLSX 1174 kb)
Additional file 10: Table S8.
List of network modules detected by the OCG algorithm. (XLSX 471 kb)
Additional file 11: Table S9.
Functional annotations for the 31 network modules preferentially targeted by the FusoSecretome. (XLSX 140 kb)
Additional file 12: Table S10.
Perturbation scores of the FusoSecretome proteins for the 31 preferentially targeted modules. (XLSX 65 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Zanzoni, A., Spinelli, L., Braham, S. et al. Perturbed human sub-networks by Fusobacterium nucleatum candidate virulence proteins. Microbiome 5, 89 (2017). https://doi.org/10.1186/s40168-017-0307-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40168-017-0307-1