
Abstract
Background
Innate immunity activator (INAVA) has been shown to be elevated in lung adenocarcinoma. However, its expression pattern and function in papillary thyroid cancer (PTC) are unknown. This study aimed to identify the clinical, biological, and mechanistic impacts of INAVA on PTC.
Methods
Using The Cancer Genome Atlas dataset, real time PCR, and immunohistochemistry, the expression of INAVA in PTC was analyzed. Gain- and loss-of-function assays were performed to investigate the role of INAVA in PTC cell invasion, migration, and metastasis. We explored the molecular mechanisms underlying the roles of INAVA in PTC cells using transcriptome resequencing, real time PCR, western blotting and immunohistochemistry.
Results
We found that INAVA expression was significantly upregulated in PTC and was significantly associated with lymph node metastasis. Loss- and gain-of-function experiments demonstrated that INAVA promoted the aggressive phenotype of PTC cells in vitro and in vivo. Mechanistic study suggested that upregulation of INAVA resulted in elevated fibroblast growth factor 1 (FGF1), which in turn increased the expression level of matrix metalloproteinases 9 (MMP9). We further identified that the level of INAVA was positively correlated with the levels of FGF1 and MMP9 in clinical PTC specimens.
Conclusion
These data establish a novel role for INAVA in promoting PTC progression and suggest that INAVA may represent a therapeutic target for the disease.
Similar content being viewed by others


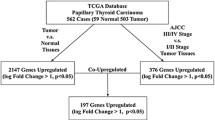
Background
Thyroid cancer is the most common type of endocrine-related malignancy and shows rapidly increasing incidences during the past decades [1]. Papillary thyroid cancer (PTC) is the most common form of thyroid cancer, representing 70–80% of all thyroid cancer [2]. In most cases, patients with PTC follow an indolent course and have excellent prognosis [3]. However, extrathyroidal extension and distant metastasis are extremely important risk factors for PTC and are still major clinical challenges [4, 5]. Thus, a good understanding of the molecular alterations responsible for aggressive behavior of PTC is still imperative.
Tumor metastasis is a multistep process during which a subset of cancer cells disseminate from the site of a primary tumor and establish secondary tumors at distant sites [6]. Matrix metalloproteinases (MMPs) are proteolytic enzymes responsible for remodeling extracellular matrix (ECM) in many physiological and pathological processes [7]. Moreover, besides its role in degradation of ECM molecules, MMPs play important roles in multiple aspects of cancer progression, including tumor growth, almost all metastatic steps, and angiogenesis [8, 9]. MMP9, one of the best characterized members in MMP family, is well known to degrade ECM and is closely related to the cancer invasion and metastasis [10]. In PTC, Ahmed et al. showed that silencing of FoxM1 inhibited migration and invasion of PTC cells via decreasing the expression levels of MMP9 and MMP2 [11]. Moreover, the expression level of arachidonate 5-lipoxygenase (ALOX5) was upregulated in PTC and it promoted PTC cell invasion through increased secretion of MMP9 [Full size image
Overexpression of INAVA promotes PTC cells invasion, migration and metastasis
Given the association of INAVA with more aggressive PTC, we next explored whether the function of INAVA in PTC could explain this association. We ectopically overexpressed INAVA in two PTC cell lines (TPC1 and K1) using empty vector as a control to determine its effect on the aggressive phenotype of the cells (Fig. 2a). As shown in Fig. 2b, c, in vitro cell invasion and migration assays showed that overexpression of INAVA significantly enhanced both the invasion and migration abilities of TPC1 and K1 cells. Scratch wound healing assay was performed to assess the effect of INAVA on cellular migration and the results showed that overexpression of INAVA significantly promoted wound closure in TPC1 and K1 cells (Fig. 2d). In an in vivo experimental metastasis assay, overexpression of INAVA promoted mouse lung colonization by tail vein-injected K1 cells (Fig. 2e and Additional file 1: Figure S1a).

Overexpression of INAVA promotes cell invasion, migration and metastasis. a Overexpression of INAVA in PTC cell lines (TPC1 and K1) was assessed by WB. α-Tubulin was used as a loading control. b Representative images (left) and quantification (right) of transwell migration assays in TPC1, TPC1-INAVA, K1, and K1-INAVA cells. c Representative images (upper) and quantification (lower) of Transwell invasion assays in indicated cells. d Scratch wound healing assays were performed in TPC1-Vector, TPC1-INAVA, K1-Vector, and K1-INAVA cells (left). Quantification analyses for the wound healing assays were shown (right). e Representative bioluminescence images in mice with tail vein injection of the K1-Vector and K1-INAVA cells (left). Representative histopathology of lung metastasis developed in the indicated animals stained with HE (right). For b–d, data are quantified as mean ± SD of 3 independent experiments in the bar graphs. *P < 0.05
Knockdown of INAVA inhibits PTC cells invasion, migration and metastasis
We next established pools of TPC1 and K1 cell lines with stable depletion of INAVA using retroviral-based shRNA vectors (Fig. 3a). As shown in Fig. 3b–e, Transwell assay with or without Matrigel coating showed that silencing of INAVA significantly decreased the invasiveness and migration of the TPC1 and K1 cell lines. Moreover, wound healing assay showed that knockdown of INAVA inhibited the migratory speed of TPC1 and K1 cells in comparison with that of the vector controls (Fig. 3f). In addition, knockdown of INAVA in K1 cells significantly decreased pulmonary metastatic nodule development (Fig. 3g and Additional file 1: Figure S1b). These data, together with results of Fig. 2, suggest that INAVA greatly contributes to the development of PTC invasion and metastasis.

Silencing of INAVA inhibits cell invasion, migration and metastasis. a Protein expression levels of INAVA in INAVA-silencing and vector-control cells were analyzed by WB. α-Tubulin was used as a loading control. Representative images (b) and quantification (c) of transwell migration assays in indicated cells. Representative images (d) and quantification (e) of transwell invasion assays in indicated cells. f Scratch wound healing assays were performed in indicated cells (left) and quantification analyses for the assays were shown (right). g Representative bioluminescence images in mice with tail vein injection of the indicated cells (left) and representative histopathology of lung metastasis stained with HE was shown (right). For b–f, data are quantified as mean ± SD of 3 independent experiments in the bar graphs. *P < 0.05
MMP9 is the key molecule mediating INAVA-induced invasion and migration ability of PTC cells
To explore the molecular mechanisms by which INAVA regulates the invasion and metastasis ability of PTC cells, transcriptome resequencing was performed on INAVA-overexpressing TPC1 cells and vector control cells. Notably, many invasion/metastasis related genes were significantly increased in INAVA overexpressing TPC1 cells as compared with that in vector control (Fig. 4a). Importantly, among these genes, MMP9 was the most elevated gene in response to ectopic overexpression of INAVA. Next, real time RT-PCR assays were performed to confirm the results of deep sequencing analysis. As shown in Fig. 4b, the MMP9 mRNA expression levels were significantly increased and decreased in INAVA overexpressing and knockdown cell lines, respectively, suggesting that transcriptional efficiency of MMP9 is regulated by INAVA in PTC cells. The protein levels of MMP9 in indicated cells were in line with the changes of mRNA (Fig. 4c). As shown in Fig. 4d, the results of gelatin zymography showed that the secretion of MMP9 into culture medium was significantly increased in cells overexpressing INAVA and was significantly decreased in cells with INAVA knockdown. Moreover, knockdown of MMP9 using specific siRNAs in INAVA overexpressing TPC1 and K1 cells (Fig. 4e) suppressed the phenotype caused by INAVA overexpression (Fig. 4f). These results indicate that MMP9 plays a vital role in INAVA-induced cell invasion and migration.

The effect of INAVA on PTC cells is mediated by MMP9. a Transcriptome resequencing was performed on TPC1-Vector and TPC1-INAVA cells and mRNA expression of the indicated genes related to invasion/metastasis is shown. b Altered cellular mRNA levels of MMP9 in the indicated cells were detected by real time RT-PCR assays. GAPDH served as a house kee** gene control. c WB assays were conducted to analyze the protein levels of MMP9 in response to deregulated INAVA expression in TPC1 and K1 cells. α-Tubulin acted as a loading control. d MMP9 activities in indicated cells were analyzed using gelatin zymography. Knockdown of MMP9 in INAVA overexpressing cells (e), attenuated phenotypic characteristics induced by INAVA overexpression (f). For b, f, data represents mean ± SD of 3 independent experiments in the bar graphs. *P < 0.05
FGF1 mediates the regulation of INAVA on MMP9
Given that MMP9 has been demonstrated to be regulated by FGF1 [19], together with that the results of deep sequencing analysis showed FGF1 was significantly upregulated in INAVA-overexpressing cells (Fig. 4a), we hypothesized that FGF1 was involved in the role of INAVA on regulation of MMP9. As expected, the mRNA and protein levels of FGF1 were elevated by overexpression of INAVA but were decreased by depletion of INAVA in PTC cells (Fig. 5a–d). Consistently, ELISA assays showed that INAVA-overexpressing cells secreted increased amount of FGF1, whereas INAVA silencing reduced the secretion of FGF1 (Fig. 5e). To further confirm the involvement of FGF1 in INAVA-mediated effects, the specific siRNAs were used to knockdown FGF1 in INAVA overexpressing cells. As shown in Fig. 5f, the upregulation of MMP9 induced by INAVA were inhibited by FGF1 silencing. As anticipated, invasive phenotype caused by INAVA overexpression was also suppressed by depletion of FGF1 (Fig. 5g). Taken together, these data indicate FGF1 play a vital role in INAVA-induced MMP9 expression and cell invasion.

INAVA modulates MMP9 via regulating FGF1. mRNA levels of FGF1 were examined in INAVA-overexpressing (a) or -knockdown cells (b). c, d Protein levels of FGF1 were assessed in indicated cells. e ELISA assays were performed to assess the effect of INAVA on FGF1 release in TPC1 and K1 cells. f Knockdown of FGF1 by siRNAs in INAVA-overexpressing cells reversed increased level of MMP9. g Invasiveness caused by INAVA was abrogated by silencing FGF1 expression in INAVA-overexpressing cells. For a, b, e, g, results derived from three independent experiments are expressed as mean ± SD. *P < 0.05
INAVA expression correlates with FGF1 and MMP9 expression in primary PTC
To evaluate the correlations between INAVA expression and the levels of FGF1 and MMP9 in clinical specimens, IHC analyses were performed. As shown in Fig. 6a, b, 64.9% (37 cases) and 61.4% (35 cases) of samples with low INAVA expression (57 cases), respectively, exhibited low levels of FGF1 and MMP9, whereas 30.9% (17 cases) and 20.0% (11 cases) of samples with high INAVA expression (55 cases) showed low expression of FGF1 and MMP9, respectively (P < 0.05). Furthermore, we sorted patient cohort into low and high INAVA expression using RNAseq data mined from TCGA (INAVA low and high, n = 248 respectively). As shown in Fig. 6c, INAVA expression significantly correlated with the levels of FGF1 and MMP9 in PTC specimens. These results suggest that INAVA overexpression in PTC is associated with FGF1–MMP9 axis.

INAVA positively correlates with FGF1 and MMP9 in PTC clinical specimens. a The expression of INAVA is associated with FGF1 and MMP9 in clinical PTC specimens. Two representative cases are shown. b Percentage of cases showing low (n = 57) or high (n = 55) INAVA expression in relation to the FGF1 and MMP9 expression levels. c The association between INAVA and FGF1 as well as MMP9 in 496 cases of PTC specimens using RNAseq data deposited on TCGA. The expression level of INAVA was divided into low (n = 248) and high (n = 248) groups according to median distribution. For b, c, *P < 0.05
