
Abstract
Escherichia coli, one of the most efficient expression hosts for recombinant proteins (RPs), is widely used in chemical, medical, food and other industries. However, conventional expression strains are unable to effectively express proteins with complex structures or toxicity. The key to solving this problem is to alleviate the host burden associated with protein overproduction and to enhance the ability to accurately fold and modify RPs at high expression levels. Here, we summarize the recently developed optimization strategies for the high-level production of RPs from the two aspects of host burden and protein activity. The aim is to maximize the ability of researchers to quickly select an appropriate optimization strategy for improving the production of RPs.
Similar content being viewed by others


Introduction
Since the last century, the emergence of recombinant protein (RP) expression systems has revolutionized biotechnology. Excitingly, with the advancement of biotechnology, the yield of RPs has increased from the gram to the kilogram scale, and the range of applications has expanded from traditional food and chemical industries to biopharmaceuticals [1, 2]. For example, it is projected that the industrial enzyme market will grow from USD 6.6 billion in 2021 to USD 9.1 billion by 2026 [3], illustrating the enormous market value and growth potential of RPs. Similarly, a variety of protein drugs have been successfully marketed, including monoclonal antibodies (mAbs), recombinant vaccines, and hormones, demonstrating that RPs already play a significant role in the biopharmaceutical field [4].
Due to its inexpensive fermentation requirements, rapid proliferation ability and stable high-level expression, Escherichia coli (hereafter E. coli) has become the mainstay of RP expression among prokaryotic expression hosts [5]. As early as the 1970s, E. coli was applied in the production of clinical drugs, such as the hormones somatostatin [6] and insulin [7], which were commercialized early on. As a gold standard for expressing RPs, E. coli BL21(DE3) and the pET expression system are widely used in research and commercial production. This is primarily attributed to the T7 RNA polymerase (RNAP) from λ prophage in the genome of BL21(DE3), which can specifically recognize the T7 promoter (PT7) on the pET plasmid and transcribe at eightfold the speed of the E. coli native RNAP [8, 9]. In recent years, several BL21(DE3)-derived strains have been widely used to produce various types of RPs, including C41/C43(DE3) (for the production of membrane proteins) [10], BL21(DE3)-pLysS (for reduction of T7 RNAP expression intensity) [11], BL21Star(DE3) (for improvement of mRNA stability) [12], and SixPack (for codon bias correction) [13]. Such efficient production capacity has given it an unassailable position in structural research, new enzyme mining and industrial production [14, 15].
Despite the availability of so many alternative expression systems, there is no guarantee that every type of protein will have a high yield or catalytic/functional activity. The occurrence of these phenomena can be attributed to two main aspects: (i) the host burden caused by the massive production of RPs [16] and (ii) the limited post-translational modification (PTM) capacity and generation of inclusion bodies (IBs) [17]. In fact, any production of RPs, especially toxic proteins, will inevitably compete with the host for resources, which are mainly reflected in the additional DNA replication burden, competition for transcription- and translation-related elements (RNAP, ribosomes, tRNA, and amino acids), and the additional energy and substrates consumed by PTMs [18]. For instance, high-level expression of membrane proteins can lead to the saturation of the Sec translocator-dependent transport pathway, affecting electron transport in the respiratory chain and inhibiting the expression of key enzymes of the tricarboxylic acid cycle [19]. Similarly, glucose dehydrogenase (GDH, an industrial enzyme) leads to significant autolysis of the bacterial cell during the later stages of fermentation [20]. To solve this problem, various means of genetic engineering and synthetic biology have been applied to alleviate host burden, including optimization of the expression intensity of T7 RNAP and pET expression systems (Fig. 1A) [21, 22], as well as balancing or decoupling the cell growth and RP production [23,24,25]. These optimization strategies effectively relieve or even remove the metabolic burden and increase the capacity of unit cell production. However, when proteins are synthesized at high rates, limited PTMs and molecular chaperones can lead to protein misfolding and the formation of a large number of IBs, affecting the functional activity and solubility of certain proteins. Therefore, the production of highly active RPs is also an important optimization aim, which can be achieved by strengthening or supplementing PTMs, increasing proteolysis and overexpressing suitable molecular chaperones [26]. This review summarizes different classes of optimization strategies developed in recent years from the two main aspects of alleviating host burden and optimizing protein activity, providing a reference for increasing the production of different RPs and discusses the future development direction of related optimization strategies.

The optimization expression strategies for T7 RNAP and pET plasmids. A Illustration of protein expression of recombinant protein genes on pET plasmids. B Optimization of T7 RNAP transcription and translation level, including substitutions of different promoters, and mutations in promoter functional region and RBS sequence. C regulation of T7 RNAP activity. The conventional approach is to utilize lysozyme or light-induction to regulate. D Optimization of pET plasmids based on expression intensity and copy numbers. Among them, the expression intensity was optimized by constructing an ITR library to screen for optimal expression results. The degree of binding of RNA-i to RNA-p determines the replication intensity of the plasmid to control the copy numbers. By constructing a promoter library for RNA-p, replacing the inducible promoter, and using dCas9 to regulate expression intensity, the copy numbers can be controlled
Optimization of target protein expression rate based on the gold standard T7 RNAP platform
When T7 RNAP is sufficiently induced, its powerful transcriptional capacity enables the rapid production of large amounts of mRNA, bringing the yield of RPs to 50% of the total cellular protein in just a few hours [27]. However, a strong production capacity is a double-edged sword, especially in the expression of toxic proteins. Numerous studies have shown that growth inhibition during RP production is mainly attributed to excessively strong gene transcription, and translation further exacerbates the host burden [21, 28, 29]. Therefore, the ability to precisely balance the intensity of RP transcription and translation levels is key to reducing host burden and increasing production. This is usually optimised in two aspects as follows: T7 RNAP and pET plasmid.
Regulation of the target protein expression rate-T7 RNAP
The easiest way to control the expression intensity of RPs is to regulate the amount and activity of T7 RNAP, which is often achieved by optimizing transcription or translation levels. In the BL21(DE3) genome, the T7 RNAP gene is controlled by the lacUV5 promoter (PlacUV5), which is a strongly inducible promoter that ensures rapid expression and accumulation after induction (induced by Isopropyl-beta-d-thiogalactopyranoside (IPTG)) [30]. However, high levels of expression are not compatible with some RPs, especially toxic proteins. Accordingly, many studies increased the production of toxic proteins by reducing the transcript level of T7 RNAP. For example, the membrane protein expression host C41(DE3) was obtained by stress screening, while the autolysin expression host BL21(DE3-lac1G) was constructed by recombining PlacUV5 with Plac sequences [10, 20, 31]. Furthermore, the PlacUV5 is independent of CRP, which makes it leakier than Plac [32]. Replacing the promoter of T7 RNAP with other kinds of inducible promoters is an effective way to regulate transcription levels and reduce leakage (Fig. 1B). Du et al. [32] tested the effects of three inducible promoters (ParaBAD, PrhaBAD and Ptet) on the transcriptional intensity and leaky expression of T7 RNAP, respectively. It was found that all three promoters were suitable for prolonged fermentation of toxic proteins, whereby PrhaBAD and Ptet were able to regulate T7 RNAP transcription more rigorously, providing additional options for the expression of various RPs, especially toxic proteins. Similarly, enhancing the ability to block proteins is also an effective way to reduce leaky expression. In addition to the conversion of PlacUV5 to Plac, the study found that the lac repressor gene (lacI) was also mutated (V192F, referred to as mLacI hereafter) in the membrane protein expression host (C41/C43(DE3)) [33]. Excitingly, mLacI can specifically bind to the lac operator site, but the blocking effect cannot be removed by the addition of IPTG. Based on this phenomenon, Kim et al. [31] developed an anti-leakage expression system for the overproduction of membrane proteins. Among them, mLacI expression is regulated by the rhamnose inducible promoter PrhaBAD. When trace amounts of L-rhamnose were added, T7 RNAP leakage expression could be inhibited during host growth, reducing growth burden. With the increasing concentration of L-rhamnose, mLacI is abundantly produced and thus reduces the transcription intensity of T7 RNAP, even in the presence of IPTG. This approach makes it possible to control the rate of protein production.
Unlike the transcriptional level, which is controlled by the promoter and RNAP, the strength of translation is mainly determined by the nucleotide sequence and arrangement of the ribosome binding site (RBS) (Fig. 1B). Liang et al. [34] designed 10 RBS sequences with different expression intensities for expressing T7RNAP using an RBS calculator, which was successfully implemented in five Gram-negative and one Gram-positive bacteria. To further extend the regulatory range, Li et al. [35] constructed a more extensive RBS library of T7 RNAP using CRISPR/Cas9 and cytosine base editor, with expression levels ranging from 28 to 220% of the wild-type strain. Using this library, the authors obtained customized hosts for eight difficult-to-express proteins in just three days. The tested model RPs included an autolytic protein, membrane protein, antimicrobial peptide, and insoluble protein, while the production of the industrial enzyme GDH was increased 298-fold. These results show that optimizing the expression intensity of T7 RNAP can effectively improve the RP production, and regulation of the translational level makes it easier to construct screening libraries and rapidly obtain optimized hosts for individual RPs.
Since it is an enzyme, the catalytic activity of T7 RNAP is also a key factor affecting the rate and efficiency of transcription. Mutations of key amino acid residues in T7 RNAP are one of the most effective methods to tune its activity, whose mechanisms are divided into two categories: weakening the binding ability to PT7 or generating code-shifting mutations to reduce the catalytic activity [36,37,38]. For example, Baumgarten et al. [37] found a single amino acid mutation (A102D) of T7 RNAP in the membrane protein expression host Mt56(DE3), which reduced the ability to bind to the PT7 and decreased the RP production rate. In addition, the addition of T7 RNAP inhibitors is also a way to effectively regulate T7 RNAP activity, and various derivative hosts including BL21(DE3)-pLysS, BL21(DE3)-pLysE, and Lemo21(DE3) have been developed based on this principle [39,40,41] (Fig. 1C). With the development of synthetic biology, researchers hope to change the strength of T7 RNAP activity in logic gates to precisely and dynamically regulate the process of growth and production. A variety of T7 RNAP expression systems regulated by light induction have been developed successively, achieving dynamic regulation of RP production [42,43,44]. For example, the Opto-T7RNAPs system splits the T7RNAP into two fragments and expresses them in tandem with a light-sensitive dimerization domain. When the fragments are expressed and irradiated by the light of a specific wavelength, T7 RNAP can resume its transcriptional activity, with up to 80-fold change in activity between blue light and darkness [43]. Regrettably, these studies have only been validated with fluorescent proteins or lycopene, and have not been applied to RP production.
Regulation of the target protein expression rate-pET plasmid
Another key factor affecting the expression rate of RPs depends on the combination of different elements on the pET plasmid, including sequences of relevant functional regions near PT7 (-35/-10 region, translation initiation region (TIR) and operator sequence) and replicon [45]. As the core region of the pET plasmid, various functional regions near the PT7 determine the rigor of basal expression before induction and the appropriate transcription rate after induction.
To reduce the host burden of leaky expression, several more rigorous inducible systems have been combined with PT7 to increase the yield of toxic or structurally complex proteins, such as the cumate operator [46], inducible translational ON orthogonal riboswitch [47], and temperature-regulated self-induction [48]. After solving the leaky expression problem, an urgent task is to quickly screen the appropriate expression intensity of various RPs. In contrast to complex genomic manipulations, the combination of degenerate primers and MEGAWHOP PCR or enzymatic digestion and ligation allows rapid access to very large libraries of various functional sequences, including promoter mutation and TIR libraries [22, 84, 85]. Among them, Liu et al. [84] utilized phage-assisted continuous evolution technology for rapid optimization of 16S rRNA by screening pressure. After multiple rounds of directed evolution, the mutant o-ribosome achieved faster translation, resulting in 6.3-fold higher RP production than the wild-type. Most importantly, this ribosome can introduce ncAAs into the protein with high efficiency, which is 9.08-fold higher than that of the native ribosome, improving the application of orthogonal translation systems in RP production. In brief, whether it is to inhibit or block the expression of growth-essential genes or to use o-ribosomes to express RPs, the aim is to ensure normal growth of the host during the growth phase (Fig. 2D).
Optimizing protein activity—another key to the production
In addition to ensuring the quantity of RPs, the functional activity of the protein at high yields is also a key focus of RP production. When the expression rate or quantity of RPs exceeds the capacity of the host cell, it will result in a large number of proteins that misfold and aggregate, eventually producing IBs [17]. This phenomenon has greatly hindered the use of E. coli in various fields, especially the expression of protein-based drugs. The key reason for the generation of IBs is the limited PTM capacity and folding efficiency, which are the top priorities for optimizing the functional activity of RPs.
Enhancement of post-translational modifications
Most proteins with complex structures contain multiple disulfide bonds (DSBs) that maintain their normal conformation, including insulin [7] and epidermal growth factor [86]. As an oxidative process, the natural DSB formation is completed in the periplasmic space of E. coli and not in the reductive environment of the cytoplasm, which requires the protein to be localized and translocated to the appropriate location for modification [87]. The common protein translocation pathways are divided into three main categories: SecB-dependent, SRP-mediated and TAT translocation pathways [88]. Among them, SecB-dependent and SRP-mediated pathways both complete the translocation process by binding to SecA, and genetic fusion of signal peptides to RPs can enable them to utilize these pathways to translocate. Commonly used signal peptides include pelB, OmpA and DsbA [89, 90], but each signal peptide triggers a different mechanism that greatly affects the effectiveness of RP transport. In contrast to SRP-mediated DsbA, SecB-dependent OmpA drives the synthesis of endogenous secreted and membrane proteins, preventing Sec translocator saturation [89]. In recent years, the TAT translocation pathway has attracted the interest of researchers due to its natural "quality control" system, which can prioritize the output of correctly folded proteins [91]. The "TatExpress" strain was successfully developed and applied for the gram-level production of human growth hormone, proving its great potential [92]. In addition to the above translocation pathways, a signal peptide based on the N-terminal sequence of penicillin-binding protein 2 (PBP2) was shown to anchor the fusion protein to the cytoplasmic membrane. Interestingly, the high expression of PBP2 affects morphological changes in E. coli (rods to spheres) and interacts with lysis transglycosylase leading to host lysis [93]. This phenomenon has the potential to be developed into a self-cleaving transport system for rapidly accumulating RPs production.
Compared to the narrow periplasmic space, the cytoplasm has enough space to accomplish more protein folding and increase productivity. By blocking the natural reduction pathway in a Δgor/ΔtrxB strain, the reductive cytoplasmic environment becomes oxidative, which facilitates the formation of DSBs [94]. The earliest commercial DSB-forming E. coli strain, Origami from Novagen, was developed based on this principle. By overexpressing sulfhydryl oxidase from the yeast mitochondria and disulfide bond isomerase from human cells, a host called CyDisCo was developed for the production of RPs with high DSB content, and was able to produce even perlecan with 44 DSBs (Fig. 3A) [95, 96]. Apart from the above, other means of optimization, including replacement of sulfhydryl oxidases from other sources [97], inversion or development of a periplasmic transmembrane disulfide bond-forming enzyme DsbB [98, 99], were also used to improve the efficiency and capacity of DSB formation.

The optimization strategies to enhance PTMs. A Principle of disulfide bond formation in the cytoplasm using the CyDisCo system. B Modification process of phosphorylation and acetylation. P: phosphonate; AC: acetyl. C Modification process of glycosylation by overexpression of a heterologous N/O-glycosylase. D Introduction of PTMs via ncAA. The figure shows the principle of phosphoserine introduction
In addition to the formation of DSBs, the efficiency of other PTMs also affects the functional activity of RPs, such as phosphorylation, acetylation (Fig. 3B), glycosylation and many other modifications that are often found in mAbs and functional proteins [100,101,102]. Among them, glycosylation is one of the most abundant and complex PTMs [103]. By linking monosaccharides, oligosaccharides or polysaccharides to proteins, the variety of protein functional activities is greatly expanded. Currently, over 70% of therapeutic proteins are modified by glycosylation, and precision glycosylation can effectively enhance the use of glycoproteins in the medical industry [102]. Compared to eukaryotes, E. coli does not have a natural mechanism for glycosylation of encoded proteins. Therefore, it can be used as a suitable chassis cell to develop bottom-up glycoengineering for different types of glycoproteins [104]. The first N-glycosylation expression system was successfully developed in E. coli by introducing genes related to N-glycosylation of Campylobacter jejuni, opening the curtain on the glycoprotein synthesis in E. coli [105] (Fig. 3C). Over the last two decades, many efforts have conferred the potential to produce a wide range of N/O-glycoproteins from E. coli or cell-free extracts, including optimization of glycosyltransferase substrate identification and orthogonality [102, 106,107,108], exploration of glycosylase function from multiple sources [107,108,109] and optimization of host environment, metabolic pathways and culture conditions [110,111,112,113]. Based on these studies, a variety of medically relevant products are in production and in the clinical phase, such as recombinant vaccine exotoxin A [114], therapeutic protein O-glycosylated interferon-α2b [115] and N-glycosylated mannose3-N-acetylglucosamine2 [116]. In a similar way to DSB, the glycosylation process in the above systems is mostly completed in the periplasmic space. In recent years, several studies have identified cytoplasmic glycosylation systems in various bacteria, laying the foundation for the development of novel glycosylation systems in E. coli [117,118,119]. Among them, the asparagine (N)-glucosyltransferase from Actinobacillus pleuropneumoniae (ApNGT) can be actively expressed in the E. coli cytoplasm and transfer glucose residues to the naturally N-terminal glycosylation site of the protein (e.g. recombinant human EPO) [117]. Based on this discovery, Tytgat et al. [120] developed an N-glycosylation system in E. coli cytoplasm. Using ApNGT in combination with various oligosaccharide synthesis pathways (e.g. human milk oligosaccharides and glycosphingolipids), glycosylation modifications of various glycoproteins (glycoconjugate vaccines and multivalent glycopolymers) have been achieved. Surprisingly, the system can complete the glycosylation of megadalton protein assemblies, which can be used as customized carriers for delivery of drugs and vaccines.
It is worth mentioning that the orthogonality of ncAAs with specific codons can be used to introduce various types of modified amino acids more directly and precisely. Park et al. [121] successfully introduced phosphorylated serine residues into RPs at specific sites by orthogonal pairing of SepRS/tRNASep (Fig. 3D). Similarly, phosphor-threonine [122] and phospho-tyrosine [123] were utilized for RP modification. In addition to phosphorylation, acetylation, methylation and ubiquitination have been successfully introduced into various RPs [124]. In conclusion, the introduction of PTMs using ncAAs has the potential to once again make E. coli a "star host" for biopharmaceuticals.
Elimination of inclusion bodies
In addition to limited PTMs, a variety of factors such as misfolding, low solubility, and host burden also contribute to IB formation. Three strategies are usually used to solve the problems: (i) enhancing solubility; (ii) improving correct folding efficiency; (iii) optimizing the appropriate expression intensity. Among them, the relevant aspects of (iii) have been described above.
The use of peptide tags is the most direct and effective means to enhance the solubility of RPs. Common tags include maltose binding protein (MBP), glutathione-S-transferase (GST), carbohydrate-binding module (CBM), thioredoxin, and NusA, which have been reviewed by Ki et al. [125]. Notably, a novel CBM (CBM66) was shown to have a pro-solubilizing effect on several types of RPs and to increase production titer [126]. For example, the combination of poly (ethylene terephthalate) hydrolase and CBM resulted in a 3.7-fold improvement compared to the other commercial labels (MBP and GST), without affecting protein bioactivity. However, if the molecular weight of the peptide tag is close to or larger than that of the RP, it will override the solubility of the RP itself. Furthermore, the subsequent label removal can negatively affect the solubility and stability of RPs. Conversely, the use of peptide tags with smaller molecular weights allows more reliable evaluation and optimization of the solubility of RPs. In recent years, a variety of low-molecular-weight protein tags have contributed to the solubilization and yield enhancement of various RPs, including the NEXT tag [127], low-molecular-weight protamine [128], and 6HFh8 [129]. Kim et al. utilized 6HFh8 [129] to express a variety of growth factor proteins. Among them, 6HFh8-aFGF and 6HFh8-VEGF165 obtained high respective yields of 9.7 and 3.4 g/L in a 5-L batch supplement fermentation, with a purity of more than 99%. The removal of the small peptide tags does not significantly affect the solubility and functional activity, which is suitable for the purification of small RPs.
Molecular chaperones are a class of auxiliary proteins that facilitate the folding and assembly of peptide structures, ensuring proper folding and preventing the aggregation of newly translated peptides [130]. E. coli possesses several molecular chaperone systems, such as GroES/EL and DnaK-DnaJ-GrpE, all with different functions [131]. Among them, DnaK-DnaJ-GrpE not only helps correctly fold newly translated peptides, but also functions during co- and post-translational modification. By contrast, the GroES/EL system associates with peptides only post-translationally, powering the repair of misfolded proteins [127]. It is easy to understand that the folding efficiency can be effectively enhanced by overexpression of molecular chaperones, which is usually done in three combinations: GroES/GroEL, DnaK-DnaJ-GrpE, and co-expression. However, co-expression is usually not better than expressing a single factor, and only some chaperones can have a beneficial effect on protein folding [132]. Huang et al. [133] expressed distinct combinations of molecular chaperones to enhance the solubility and activity of polyunsaturated fatty acid isomerase (PAI). The results showed that overexpression of GroES/EL increased the solubility of PAI from 29 to 97% and improved its specific activity by 57.8%. By contrast, the co-expression of DnaK-DnaJ-GrpE or GroES/EL had a weakening effect, resulting in only an 11.9% increase in activity.
Conclusion and outlook
Different types of RPs from different origins have highly specific characteristics, and there can be no single optimization strategy that applies to all proteins. This review summarizes the recently developed optimization strategies from the two major aspects of alleviating the host burden and optimizing functional activity, which helps researchers quickly select an appropriate expression strategy for their protein of interest (Table 1, Fig. 4). Encouragingly, with the continued development of synthetic biology, systems biology, and various gene editing tools, it is becoming less difficult to rapidly develop a customized host. Multiple in vivo mutagenesis strategies facilitate adaptive laboratory evolution for rapid screening of strongly tolerant expression hosts, including DNA replication proteins, RNAP and T7 RNAP fused with base deaminases [134,135,136,137]. Construction of artificial organelles allows for E. coli compartmentalization, which has the potential to accomplish precise PTMs [138, 139]. In addition, researchers are updating the BL21(DE3) genome annotation, as well as combining mathematical modeling, statistical analysis, and computer aided design to achieve precise optimization [140, 141]. In conclusion, we have reason to believe that E. coli will remain one of the brightest stars among RP production hosts.

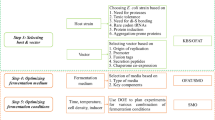
The routine workflow for expression optimization based on protein properties
References
Puetz J, Wurm FM. Recombinant proteins for industrial versus pharmaceutical purposes: a review of process and pricing. Processes. 2019;7:476–84.
Deckers M, Deforce D, Fraiture M-A, Roosens NH. Genetically modified micro-organisms for industrial food enzyme production: an overview. Foods. 2020;9:326–45.
Industrial Enzymes Market. https://www.marketsandmarkets.com/Market-Reports/industrial-enzymes-market-237327836.html. Accessed Jan 2022.
Walsh G. Biopharmaceutical benchmarks 2018. Nat Biotechnol. 2018;36:1136–45.
Deo S, Turton KL, Kainth T, Kumar A, Wieden H-J. Strategies for improving antimicrobial peptide production. Biotechnol Adv. 2022:107968–84.
Itakura K, Hirose T, Crea R, Riggs AD, Heyneker HL, Bolivar F, Boyer HW. Expression in Escherichia coli of a chemically synthesized gene for the hormone somatostatin. Science. 1977;198:1056–63.
Williams DC, Van Frank RM, Muth WL, Burnett JP. Cytoplasmic inclusion bodies in Escherichia coli producing biosynthetic human insulin proteins. Science. 1982;215:687–9.
Studier FW, Moffatt BA. Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J Mol Biol. 1986;189:113–30.
Iost I, Guillerez J, Dreyfus M. Bacteriophage T7 RNA polymerase travels far ahead of ribosomes in vivo. J Bacteriol. 1992;174:619–22.
Miroux B, Walker JE. Over-production of proteins in Escherichia coli: mutant hosts that allow synthesis of some membrane proteins and globular proteins at high levels. J Mol Biol. 1996;260:289–98.
Studier FW. Use of bacteriophage T7 lysozyme to improve an inducible T7 expression system. J Mol Biol. 1991;219:37–44.
Lopez PJ, Marchand I, Joyce SA, Dreyfus M. The C-terminal half of RNase E, which organizes the Escherichia coli degradosome, participates in mRNA degradation but not rRNA processing in vivo. Mol Microbiol. 1999;33:188–99.
Lipinszki Z, Vernyik V, Farago N, Sari T, Puskas LG, Blattner FR, Posfai G, Gyorfy Z. Enhancing the translational capacity of E. coli by resolving the codon bias. ACS Synth Biol. 2018;7:2656–64.
Chapman J, Ismail AE, Dinu CZ. Industrial applications of enzymes: recent advances, techniques, and outlooks. Catalysts. 2018;8:238–63.
Sørensen HP, Mortensen KK. Advanced genetic strategies for recombinant protein expression in Escherichia coli. J Biotechnol. 2005;115:113–28.
Weber J, Li Z, Rinas U. Recombinant protein production provoked accumulation of ATP, fructose-1, 6-bisphosphate and pyruvate in E coli K12 strain TG1. Microb Cell Fact. 2021;20:1–8.
Tripathi NK, Shrivastava A. Recent developments in bioprocessing of recombinant proteins: expression hosts and process development. Front Bioeng Biotechnol. 2019;7:420–54.
Rugbjerg P, Sommer MO. Overcoming genetic heterogeneity in industrial fermentations. Nat Biotechnol. 2019;37:869–76.
Wagner S, Baars L, Ytterberg AJ, Klussmeier A, Wagner CS, Nord O, Nygren P-A, van Wijk KJ, de Gier J-W. Consequences of membrane protein overexpression in Escherichia coli. Mol Cell Proteomics. 2007;6:1527–50.
Sun XM, Zhang ZX, Wang LR, Wang JG, Liang Y, Yang HF, Tao RS, Jiang Y, Yang JJ, Yang S. Downregulation of T7 RNA polymerase transcription enhances pET-based recombinant protein production in Escherichia coli BL21(DE3) by suppressing autolysis. Biotechnol Bioeng. 2021;118:153–63.
Tan S-I, Ng I-S. New insight into plasmid-driven T7 RNA polymerase in Escherichia coli and use as a genetic amplifier for a biosensor. ACS Synth Biol. 2020;9:613–22.
Shilling PJ, Mirzadeh K, Cumming AJ, Widesheim M, Köck Z, Daley DO. Improved designs for pET expression plasmids increase protein production yield in Escherichia coli. Commun Biol. 2020;3:1–8.
Lozano Terol G, Gallego-Jara J, Sola Martínez RA, Cánovas Díaz M, de Diego PT. Engineering protein production by rationally choosing a carbon and nitrogen source using E. coli BL21 acetate metabolism knockout strains. Microb Cell Fact. 2019;18:1–19.
Kumar J, Chauhan AS, Shah RL, Gupta JA, Rathore AS. Amino acid supplementation for enhancing recombinant protein production in E. coli. Biotechnol Bioeng. 2020;117:2420–33.
Stargardt P, Feuchtenhofer L, Cserjan-Puschmann M, Striedner G, Mairhofer J. Bacteriophage inspired growth-decoupled recombinant protein production in Escherichia coli. ACS Synth Biol. 2020;9:1336–48.
Mital S, Christie G, Dikicioglu D. Recombinant expression of insoluble enzymes in Escherichia coli: a systematic review of experimental design and its manufacturing implications. Microb Cell Fact. 2021;20:1–20.
Graumann K, Premstaller A. Manufacturing of recombinant therapeutic proteins in microbial systems. Biotechnol J. 2006;1:164–86.
Mittal P, Brindle J, Stephen J, Plotkin JB, Kudla G. Codon usage influences fitness through RNA toxicity. Proc Natl Acad Sci USA. 2018;115:8639–44.
Li Z, Rinas U. Recombinant protein production associated growth inhibition results mainly from transcription and not from translation. Microb Cell Fact. 2020;19:1–11.
Jeong H, Barbe V, Lee CH, Vallenet D, Yu DS, Choi S-H, Couloux A, Lee S-W, Yoon SH, Cattolico L. Genome sequences of Escherichia coli B strains REL606 and BL21(DE3). J Mol Biol. 2009;394:644–52.
Kim SK, Lee D-H, Kim OC, Kim JF, Yoon SH. Tunable control of an Escherichia coli expression system for the overproduction of membrane proteins by titrated expression of a mutant lac repressor. ACS Synth Biol. 2017;6:1766–73.
Du F, Liu Y-Q, Xu YS, Li ZJ, Wang YZ, Zhang ZX, Sun XM. Regulating the T7 RNA polymerase expression in E coli BL21(DE3) to provide more host options for recombinant protein production. Microb Cell Fact. 2021;20:1–10.
Kwon S-K, Kim SK, Lee D-H, Kim JF. Comparative genomics and experimental evolution of Escherichia coli BL21 (DE3) strains reveal the landscape of toxicity escape from membrane protein overproduction. Sci Rep. 2015;5:1–13.
Liang X, Li C, Wang W, Li Q. Integrating T7 RNA polymerase and its cognate transcriptional units for a host-independent and stable expression system in single plasmid. ACS Synth Biol. 2018;7:1424–35.
Li ZJ, Zhang ZX, Xu Y, Shi TQ, Ye C, Sun XM, Huang H. CRISPR-Based Construction of a BL21 (DE3)-derived variant strain library to rapidly improve recombinant protein production. ACS Synth Biol. 2022;11:343–52.
Temme K, Hill R, Segall-Shapiro TH, Moser F, Voigt CA. Modular control of multiple pathways using engineered orthogonal T7 polymerases. Nucleic Acids Res. 2012;40:8773–81.
Baumgarten T, Schlegel S, Wagner S, Löw M, Eriksson J, Bonde I, Herrgård MJ, Heipieper HJ, Nørholm MH, Slotboom DJ. Isolation and characterization of the E coli membrane protein production strain Mutant56 (DE3). Sci Rep. 2017;7:1–14.
Tan S-I, Hsiang C-C, Ng I-S. Tailoring genetic elements of the plasmid-driven T7 system for stable and robust one-step cloning and protein expression in broad Escherichia coli. ACS Synth Biol. 2021;10:2753–62.
Huang J, Villemain J, Padilla R, Sousa R. Mechanisms by which T7 lysozyme specifically regulates T7 RNA polymerase during different phases of transcription. J Mol Biol. 1999;293:457–75.
Zhang X, Studier FW. Mechanism of inhibition of bacteriophage T7 RNA polymerase by T7 lysozyme. J Mol Biol. 1997;269:10–27.
Schlegel S, Löfblom J, Lee C, Hjelm A, Klepsch M, Strous M, Drew D, Slotboom DJ, de Gier J-W. Optimizing membrane protein overexpression in the Escherichia coli strain Lemo21(DE3). J Mol Biol. 2012;423:648–59.
Han T, Chen Q, Liu H. Engineered photoactivatable genetic switches based on the bacterium phage T7 RNA polymerase. ACS Synth Biol. 2017;6:357–66.
Baumschlager A, Aoki SK, Khammash M. Dynamic blue light-inducible T7 RNA polymerases (Opto-T7RNAPs) for precise spatiotemporal gene expression control. ACS Synth Biol. 2017;6:2157–67.
Raghavan AR, Salim K, Yadav VG. Optogenetic control of heterologous metabolism in E. coli. ACS Synth Biol. 2020;9:2291–300.
Rosano GL, Ceccarelli EA. Recombinant protein expression in Escherichia coli: advances and challenges. Front Microbiol. 2014;5:172–88.
Chaudhary AK, Lee EY. Tightly regulated and high level expression vector construction for Escherichia coli BL21(DE3). J Ind Eng Chem. 2015;31:367–73.
Horga LG, Halliwell S, Castiñeiras TS, Wyre C, Matos CF, Yovcheva DS, Kent R, Morra R, Williams SG, Smith DC. Tuning recombinant protein expression to match secretion capacity. Microb Cell Fact. 2018;17:1–18.
Anilionyte O, Liang H, Ma X, Yang L, Zhou K. Short, auto-inducible promoters for well-controlled protein expression in Escherichia coli. Appl Microbiol Biotechnol. 2018;102:7007–15.
Nie Z, Luo H, Li J, Sun H, **ao Y, Jia R, Liu T, Chang Y, Yu H, Shen Z. High-throughput screening of T7 promoter mutants for soluble expression of cephalosporin C acylase in E. coli. Appl Biochem Biotechnol. 2020;190:293–304.
Mirzadeh K, Martinez V, Toddo S, Guntur S, Herrgard MJ, Elofsson A, Nørholm MH, Daley DO. Enhanced protein production in Escherichia coli by optimization of cloning scars at the vector–coding sequence junction. ACS Synth Biol. 2015;4:959–65.
Mirzadeh K, Shilling PJ, Elfageih R, Cumming AJ, Cui HL, Rennig M, Nørholm MH, Daley DO. Increased production of periplasmic proteins in Escherichia coli by directed evolution of the translation initiation region. Microb Cell Fact. 2020;19:1–12.
Rennig M, Martinez V, Mirzadeh K, Dunas F, Rojsater B, Daley DO, Nørholm MH. TARSyn: tunable antibiotic resistance devices enabling bacterial synthetic evolution and protein production. ACS Synth Biol. 2018;7:432–42.
Rouches MV, Xu Y, Cortes LBG, Lambert G. A plasmid system with tunable copy number. Nat Commun. 2022;13:1–12.
Ajikumar PK, **ao W-H, Tyo KE, Wang Y, Simeon F, Leonard E, Mucha O, Phon TH, Pfeifer B, Stephanopoulos G. Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli. Science. 2010;330:70–4.
Minton NP. Improved plasmid vectors for the isolation of translational lac gene fusions. Gene. 1984;31:269–73.
Nordström K. Plasmid R1—replication and its control. Plasmid. 2006;55:1–26.
Li C, Zou Y, Jiang T, Zhang J, Yan Y. Harnessing plasmid replication mechanism to enable dynamic control of gene copy in bacteria. Metab Eng. 2022;70:67–78.
Segall-Shapiro TH, Meyer AJ, Ellington AD, Sontag ED, Voigt CA. A ‘resource allocator’for transcription based on a highly fragmented T7 RNA polymerase. Mol Syst Biol. 2014;10:742–56.
Yari K, Fatemi SS-A, Tavallaei M. High level expression of recombinant BoNT/A-Hc by high cell density cultivation of Escherichia coli. Bioproc Biosyst Eng. 2012; 35:407–14.
Ceroni F, Algar R, Stan G-B, Ellis T. Quantifying cellular capacity identifies gene expression designs with reduced burden. Nat Methods. 2015;12:415–8.
Jung H-M, Im D-K, Lim JH, Jung GY, Oh M-K. Metabolic perturbations in mutants of glucose transporters and their applications in metabolite production in Escherichia coli. Microb Cell Fact. 2019;18:1–14.
Fuentes LG, Lara AR, Martínez LM, Ramírez OT, Martínez A, Bolívar F, Gosset G. Modification of glucose import capacity in Escherichia coli: physiologic consequences and utility for improving DNA vaccine production. Microb Cell Fact. 2013;12:1–11.
Cheng L, Yang X, Li S, Fu Q, Fu S, Wang J, Li F, Lei L, Shen Z. Impact of gene modification of phosphotransferase system on expression of glutamate dehydrogenase protein of Streptococcus suis in Escherichia coli. Biotechnol Biotec EQ. 2017;31:612–8.
Sharma AK, Shukla E, Janoti DS, Mukherjee KJ, Shiloach J. A novel knock out strategy to enhance recombinant protein expression in Escherichia coli. Microb Cell Fact. 2020;19:1–10.
Guleria R, Jain P, Verma M, Mukherjee KJ. Designing next generation recombinant protein expression platforms by modulating the cellular stress response in Escherichia coli. Microb Cell Fact. 2020;19:1–17.
Gustafsson C, Govindarajan S, Minshull J. Codon bias and heterologous protein expression. Trends Biotechnol. 2004;22:346–53.
Lipońska A, Ousalem F, Aalberts DP, Hunt JF, Boël G. The new strategies to overcome challenges in protein production in bacteria. Microb Biotechnol. 2019;12:44–7.
Rahmen N, Schlupp CD, Mitsunaga H, Fulton A, Aryani T, Esch L, Schaffrath U, Fukuzaki E, Jaeger K-E, Büchs J. A particular silent codon exchange in a recombinant gene greatly influences host cell metabolic activity. Microb Cell Fact. 2015;14:1–14.
Zhong C, Wei P, Zhang YHP. Enhancing functional expression of codon-optimized heterologous enzymes in Escherichia coli BL21(DE3) by selective introduction of synonymous rare codons. Biotechnol Bioeng. 2017;114:1054–64.
Ye Z, Li S, Hennigan JN, Lebeau J, Moreb EA, Wolf J, Lynch MD. Two-stage dynamic deregulation of metabolism improves process robustness & scalability in engineered E. coli. Metab Eng. 2021;68:106–18.
Faust G, Stand A, Weuster-Botz D. IPTG can replace lactose in auto-induction media to enhance protein expression in batch-cultured Escherichia coli. Eng Life Sci. 2015;15:824–9.
Nocadello S, Swennen EF. The new pLAI (lux regulon based auto-inducible) expression system for recombinant protein production in Escherichia coli. Microb Cell Fact. 2012;11:1–10.
Menacho-Melgar R, Ye Z, Moreb EA, Yang T, Efromson JP, Decker JS, Wang R, Lynch MD. Scalable, two-stage, autoinduction of recombinant protein expression in E coli utilizing phosphate depletion. Biotechnol Bioeng. 2020;117:2715–27.
Briand L, Marcion G, Kriznik A, Heydel J-M, Artur Y, Garrido C, Seigneuric R, Neiers F. A self-inducible heterologous protein expression system in Escherichia coli. Sci Rep. 2016;6:1–11.
Shariati FS, Keramati M, Valizadeh V, Cohan RA, Norouzian D. Comparison of E. coli based self-inducible expression systems containing different human heat shock proteins. Sci Rep. 2021; 11:1–10.
Menacho-Melgar R, Moreb EA, Efromson JP, Yang T, Hennigan JN, Wang R, Lynch MD. Improved two-stage protein expression and purification via autoinduction of both autolysis and auto DNA/RNA hydrolysis conferred by phage lysozyme and DNA/RNA endonuclease. Biotechnol Bioeng. 2020;117:2852–60.
Izard J, Gomez Balderas CD, Ropers D, Lacour S, Song X, Yang Y, Lindner AB, Geiselmann J, de Jong H. A synthetic growth switch based on controlled expression of RNA polymerase. Mol Syst Biol. 2015;11:840–55.
Stargardt P, Striedner G, Mairhofer J. Tunable expression rate control of a growth-decoupled T7 expression system by L-arabinose only. Microb Cell Fact. 2021;20:1–17.
Galindo Casas M, Stargardt P, Mairhofer J, Wiltschi B. Decoupling protein production from cell growth enhances the site-specific incorporation of noncanonical amino acids in E coli. ACS Synth Biol. 2020;9:3052–66.
Kasari M, Kasari V, Kärmas M, Jõers A. Decoupling growth and production by removing the origin of replication from a bacterial chromosome. ACS Synth Biol. 2022. doi: https://doi.org/10.1021/acssynbio.1c00618.
Li S, Jendresen CB, Grünberger A, Ronda C, Jensen SI, Noack S, Nielsen AT. Enhanced protein and biochemical production using CRISPRi-based growth switches. Metab Eng. 2016;38:274–84.
Li S, Jendresen CB, Landberg J, Pedersen LE, Sonnenschein N, Jensen SI, Nielsen AT. Genome-wide CRISPRi-based identification of targets for decoupling growth from production. ACS Synth Biol. 2020;9:1030–40.
Wan X, Pinto F, Yu L, Wang B. Synthetic protein-binding DNA sponge as a tool to tune gene expression and mitigate protein toxicity. Nat Commun. 2020;11:1–12.
Liu F, Bratulić S, Costello A, Miettinen TP, Badran AH. Directed evolution of rRNA improves translation kinetics and recombinant protein yield. Nat Commun. 2021;12:1–14.
Kolber NS, Fattal R, Bratulic S, Carver GD, Badran AH. Orthogonal translation enables heterologous ribosome engineering in Ecoli. Nat Commun. 2021;12:1–12.
Yadwad V, Wilson S, Ward O. Production of human epidermal growth factor by an ampicillin resistant recombinant Escherichia coli strain. Biotechnol Lett. 1994;16:885–90.
De Marco A. Strategies for successful recombinant expression of disulfide bond-dependent proteins in Escherichia coli. Microb Cell Fact. 2009;8:1–18.
McElwain L, Phair K, Kealey C, Brady D. Current trends in biopharmaceuticals production in Escherichia coli. Biotechnol Lett. 2022;89:1–15.
Ytterberg AJ, Zubarev RA, Baumgarten T. Posttranslational targeting of a recombinant protein promotes its efficient secretion into the Escherichia coli periplasm. Appl Environ Microbiol. 2019;85:e00671-e719.
Gawin A, Ertesvåg H, Hansen SAH, Malmo J, Brautaset T. Translational regulation of periplasmic folding assistants and proteases as a valuable strategy to improve production of translocated recombinant proteins in Escherichia coli. BMC Biotechnol. 2020;20:1–11.
Alanen HI, Walker KL, Suberbie MLV, Matos CF, Bönisch S, Freedman RB, Keshavarz-Moore E, Ruddock LW, Robinson C. Efficient export of human growth hormone, interferon α2b and antibody fragments to the periplasm by the Escherichia coli Tat pathway in the absence of prior disulfide bond formation. BBA-Mol Cell Res. 2015;1853:756–63.
Guerrero Montero I, Richards KL, Jawara C, Browning DF, Peswani AR, Labrit M, Allen M, Aubry C, Davé E, Humphreys DP. Escherichia coli “TatExpress” strains export several g/L human growth hormone to the periplasm by the Tat pathway. Biotechnol Bioeng. 2019;116:3282–91.
Legaree BA, Adams CB, Clarke AJ. Overproduction of penicillin-binding protein 2 and its inactive variants causes morphological changes and lysis in Escherichia coli. J Bacteriol. 2007;189:4975–83.
Derman AI, Prinz WA, Belin D, Beckwith J. Mutations that allow disulfide bond formation in the cytoplasm of Escherichia coli. Science. 1993;262:1744–7.
Matos CF, Robinson C, Alanen HI, Prus P, Uchida Y, Ruddock LW, Freedman RB, Keshavarz-Moore E. Efficient export of prefolded, disulfide-bonded recombinant proteins to the periplasm by the Tat pathway in Escherichia coli CyDisCo strains. Biotechnol Prog. 2014;30:281–90.
Sohail AA, Gaikwad M, Khadka P, Saaranen MJ, Ruddock LW. Production of extracellular matrix proteins in the cytoplasm of E coli: making giants in tiny factories. Int J Mol Sci. 2020;21:688–702.
Zhang W, Zheng W, Mao M, Yang Y. Highly efficient folding of multi-disulfide proteins in superoxidizing Escherichia coli cytoplasm. Biotechnol Bioeng. 2014;111:2520–7.
Hatahet F, Ruddock LW. Topological plasticity of enzymes involved in disulfide bond formation allows catalysis in either the periplasm or the cytoplasm. J Mol Biol. 2013;425:3268–76.
Mizrachi D, Robinson M-P, Ren G, Ke N, Berkmen M, DeLisa MP. A water-soluble DsbB variant that catalyzes disulfide-bond formation in vivo. Nat Chem Biol. 2017;13:1022–8.
Walsh G, Jefferis R. Post-translational modifications in the context of therapeutic proteins. Nat Biotechnol. 2006;24:1241–52.
Lapteva YS, Vologzhannikova AA, Sokolov AS, Ismailov RG, Uversky VN, Permyakov SE. In Vitro N-Terminal Acetylation of Bacterially Expressed Parvalbumins by N-Terminal Acetyltransferases from Escherichia coli. Appl Biochem Biotechnol. 2021;193:1365–78.
Natarajan A, Jaroentomeechai T, Cabrera-Sánchez M, Mohammed JC, Cox EC, Young O, Shajahan A, Vilkhovoy M, Vadhin S, Varner JD. Engineering orthogonal human O-linked glycoprotein biosynthesis in bacteria. Nat Chem Biol. 2020;16:1062–70.
Eichler J, Koomey M. Sweet new roles for protein glycosylation in prokaryotes. Trends Microbiol. 2017;25:662–72.
Harding CM, Feldman MF. Glycoengineering bioconjugate vaccines, therapeutics, and diagnostics in E coli. Glycobiology. 2019;29:519–29.
Wacker M, Linton D, Hitchen PG, Nita-Lazar M, Haslam SM, North SJ, Panico M, Morris HR, Dell A, Wren BW. N-linked glycosylation in Campylobacter jejuni and its functional transfer into E coli. Science. 2002;298:1790–3.
Silverman JM, Imperiali B. Bacterial N-glycosylation efficiency is dependent on the structural context of target sequons. J Biol Chem. 2016;291:22001–10.
Ollis AA, Zhang S, Fisher AC, DeLisa MP. Engineered oligosaccharyltransferases with greatly relaxed acceptor-site specificity. Nat Chem Biol. 2014;10:816–22.
Kightlinger W, Warfel KF, DeLisa MP, Jewett MC. Synthetic glycobiology: parts, systems, and applications. ACS Synth Biol. 2020;9:1534–62.
Keys TG, Wetter M, Hang I, Rutschmann C, Russo S, Mally M, Steffen M, Zuppiger M, Müller F, Schneider J. A biosynthetic route for polysialylating proteins in Escherichia coli. Metab Eng. 2017;44:293–301.
Yates LE, Mills DC, DeLisa MP. Bacterial glycoengineering as a biosynthetic route to customized glycomolecules. Adv Biochem Eng Biot. 2018;175:167–200.
Yates LE, Natarajan A, Li M, Hale ME, Mills DC, DeLisa MP. Glyco-recoded Escherichia coli: Recombineering-based genome editing of native polysaccharide biosynthesis gene clusters. Metab Eng. 2019;53:59–68.
Strutton B, Jaffé SR, Pandhal J, Wright PC. Producing a glycosylating Escherichia coli cell factory: the placement of the bacterial oligosaccharyl transferase pglB onto the genome. Biochem Biophys Res Commun. 2018;495:686–92.
Pratama F, Linton D, Dixon N. Genetic and process engineering strategies for enhanced recombinant N-glycoprotein production in bacteria. Microb Cell Fact. 2021;20:1–25.
Ihssen J, Kowarik M, Dilettoso S, Tanner C, Wacker M, Thöny-Meyer L. Production of glycoprotein vaccines in Escherichia coli. Microb Cell Fact. 2010;9:1–13.
Valderrama-Rincon JD, Fisher AC, Merritt JH, Fan Y-Y, Reading CA, Chhiba K, Heiss C, Azadi P, Aebi M, DeLisa MP. An engineered eukaryotic protein glycosylation pathway in Escherichia coli. Nat Chem Biol. 2012;8:434–6.
Du T, Buenbrazo N, Kell L, Rahmani S, Sim L, Withers SG, DeFrees S, Wakarchuk W. A bacterial expression platform for production of therapeutic proteins containing human-like O-linked glycans. Cell Chem Biol. 2019;26:203–12.
Naegeli A, Neupert C, Fan Y-Y, Lin C-W, Poljak K, Papini AM, Schwarz F, Aebi M. Molecular analysis of an alternative N-glycosylation machinery by functional transfer from Actinobacillus pleuropneumoniae to Escherichia coli. J Biol Chem. 2014;289:2170–9.
Rempe KA, Spruce LA, Porsch EA, Seeholzer SH, Nørskov-Lauritsen N, Geme JWS. Unconventional N-Linked Glycosylation Promotes Trimeric Autotransporter Function in Kingella kingae and Aggregatibacter aphrophilus. MBio. 2015;6:e01206-e1215.
Keys TG, Aebi M. Engineering protein glycosylation in prokaryotes. Curr Opin Syst Biol. 2017;5:23–31.
Tytgat HL, Lin C-W, Levasseur MD, Tomek MB, Rutschmann C, Mock J, Liebscher N, Terasaka N, Azuma Y, Wetter M. Cytoplasmic glycoengineering enables biosynthesis of nanoscale glycoprotein assemblies. Nat Commun. 2019;10:1–10.
Park H-S, Hohn MJ, Umehara T, Guo L-T, Osborne EM, Benner J, Noren CJ, Rinehart J, Söll D. Expanding the genetic code of Escherichia coli with phosphoserine. Science. 2011;333:1151–4.
Zhang MS, Brunner SF, Huguenin-Dezot N, Liang AD, Schmied WH, Rogerson DT, Chin JW. Biosynthesis and genetic encoding of phosphothreonine through parallel selection and deep sequencing. Nat Methods. 2017;14:729–36.
Hoppmann C, Wong A, Yang B, Li S, Hunter T, Shokat KM, Wang L. Site-specific incorporation of phosphotyrosine using an expanded genetic code. Nat Chem Biol. 2017;13:842–4.
Davis L, Chin JW. Designer proteins: applications of genetic code expansion in cell biology. Nat Rev Mol Cell Bio. 2012;13:168–82.
Ki M-R, Pack SP. Fusion tags to enhance heterologous protein expression. Appl Microbiol Biotechnol. 2020;104:2411–25.
Ko H, Kang M, Kim M-J, Yi J, Kang J, Bae J-H, Sohn J-H, Sung BH. A novel protein fusion partner, carbohydrate-binding module family 66, to enhance heterologous protein expression in Escherichia coli. Microb Cell Fact. 2021;20:1–12.
Jo BH. An intrinsically disordered peptide tag that confers an unusual solubility to aggregation-prone proteins. Appl Environ Microbiol. 2022;88:e00097-e122.
Choi SW, Pangeni R, Jung DH, Kim SJ, Park JW. Construction and characterization of cell-penetrating peptide-fused fibroblast growth factor and vascular endothelial growth factor for an enhanced percutaneous delivery system. J Nanosci Nanotechno. 2018;18:842–7.
Kim YS, Lee H-J, Han M-H, Yoon N-K, Kim Y-C, Ahn J. Effective production of human growth factors in Escherichia coli by fusing with small protein 6HFh8. Microb Cell Fact. 2021;20:1–16.
Schlieker C, Bukau B, Mogk A. Prevention and reversion of protein aggregation by molecular chaperones in the E coli cytosol: implications for their applicability in biotechnology. J Biotechnol. 2002;96:13–21.
Fatima K, Naqvi F, Younas H. A review: Molecular chaperone-mediated folding, unfolding and disaggregation of expressed recombinant proteins. Cell Biochem Biophys. 2021;79:153–74.
Yao D, Fan J, Han R, **ao J, Li Q, Xu G, Dong J, Ni Y. Enhancing soluble expression of sucrose phosphorylase in Escherichia coli by molecular chaperones. Protein Expression Purif. 2020;169:105571–80.
Huang MN, Lu XY, Zong H, Bin ZG, Shen W. Bioproduction of trans-10, cis-12-Conjugated Linoleic Acid by a Highly Soluble and Conveniently Extracted Linoleic Acid Isomerase and an Extracellularly Expressed Lipase from Recombinant Escherichia coli Strains. J Microbiol Biotechnol. 2018;28:739–47.
Eom G, Lee H, Kim S. Development of a genome-targeting mutator for the adaptive evolution of microbial cells. Nucleic Acids Res. 2022;50:e38.
Moore CL, Papa LJ III, Shoulders MD. A processive protein chimera introduces mutations across defined DNA regions in vivo. J Am Chem Soc. 2018;140:11560–4.
Álvarez B, Mencía M, de Lorenzo V, Fernández LÁ. In vivo diversification of target genomic sites using processive base deaminase fusions blocked by dCas9. Nat Commun. 2020;11:1–14.
Pan Y, **a S, Dong C, Pan H, Cai J, Huang L, Xu Z, Lian J. Random base editing for genome evolution in Saccharomyces cerevisiae. ACS Synth Biol. 2021;10:2440–6.
Wei S-P, Qian Z-G, Hu C-F, Pan F, Chen M-T, Lee SY, **a X-X. Formation and functionalization of membraneless compartments in Escherichia coli. Nat Chem Biol. 2020;16:1143–8.
Wang Y, Liu M, Wei Q, Wu W, He Y, Gao J, Zhou R, Jiang L, Qu J, **a J. Phase-Separated Multienzyme Compartmentalization for Terpene Biosynthesis in a Prokaryote. Angew Chem Int Ed. 2022;8:61–9.
Kim S, Jeong H, Kim E-Y, Kim JF, Lee SY, Yoon SH. Genomic and transcriptomic landscape of Escherichia coli BL21(DE3). Nucleic Acids Res. 2017;45:5285–93.
Packiam KAR, Ramanan RN, Ooi CW, Krishnaswamy L, Tey BT. Stepwise optimization of recombinant protein production in Escherichia coli utilizing computational and experimental approaches. Appl Microbiol Biotechnol. 2020;104:3253–66.
Acknowledgements
This work was supported by the Nature Science Foundation of Jiangsu Province (No. BK20202002).
Author information
Authors and Affiliations
Contributions
ZXZ wrote the manuscript. FTN and YZW helped with preparation of the manuscript. CXY reviewed and edited the manuscript. PS and XMS conceptualized, reviewed and edited the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, ZX., Nong, FT., Wang, YZ. et al. Strategies for efficient production of recombinant proteins in Escherichia coli: alleviating the host burden and enhancing protein activity. Microb Cell Fact 21, 191 (2022). https://doi.org/10.1186/s12934-022-01917-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-022-01917-y