
Abstract
Background
Glaesserella parasuis is the causative agent of Glässer’s disease in pigs. Seroty** is the most common method used to type G. parasuis isolates. However, the high number of non-typables (NT) and low discriminatory power make seroty** problematic. In this study, 218 field clinical isolates and 15 G. parasuis reference strains were whole-genome sequenced (WGS). Multilocus sequence types (MLST), serotypes, core-genome phylogeny, antimicrobial resistance (AMR) genes, and putative virulence gene information was extracted.
Results
In silico WGS seroty** identified 11 of 15 serotypes. The most frequently detected serotypes were 7, 13, 4, and 2. MLST identified 72 sequence types (STs), of which 66 were novel. The most predominant ST was ST454. Core-genome phylogeny depicted 3 primary lineages (LI, LII, and LIII), with LIIIA sublineage isolates lacking all vtaA genes, based on the structure of the phylogenetic tree and the number of virulence genes. At least one group 1 vtaA virulence genes were observed in most isolates (97.2%), except for serotype 8 (ST299 and ST406), 15 (ST408 and ST552) and NT (ST448). A few group 1 vtaA genes were significantly associated with certain serotypes or STs. The putative virulence gene lsgB, was detected in 8.3% of the isolates which were predominantly of serotype 5/12. While most isolates carried the bcr, ksgA, and bacA genes, the following antimicrobial resistant genes were detected in lower frequency; blaZ (6.9%), tetM (3.7%), spc (3.7%), tetB (2.8%), bla-ROB-1 (1.8%), ermA (1.8%), strA (1.4%), qnrB (0.5%), and aph3''Ia (0.5%).
Conclusion
This study showed the use of WGS to type G. parasuis isolates and can be considered an alternative to the more labor-intensive and traditional seroty** and standard MLST. Core-genome phylogeny provided the best strain discrimination. These findings will lead to a better understanding of the molecular epidemiology and virulence in G. parasuis that can be applied to the future development of diagnostic tools, autogenous vaccines, evaluation of antibiotic use, prevention, and disease control.
Similar content being viewed by others
Background
Glaesserella parasuis (G. parasuis) is a Gram-negative bacterium that causes Glässer’s disease in pigs. This disease is characterized by arthritis, meningitis, and polyserositis, commonly observed in 4 to 8-week-old pigs, but it can also sporadically occur in older pigs [1]. Clinical signs are abdominal breathing or coughing, lameness, paddling, and septicemia with acute death [2, 3]. While the agent was first described in 1910 [1], it continues to challenge the health and productivity of swine production systems today. In fact, over the last ten years, disease due to G. parasuis increased substantially in swine cases received at the Iowa State University Veterinary Diagnostic Laboratory (ISU-VDL) [4].
The bacterium is considered endemic in all swine populations, colonizing the upper respiratory tract of pigs. Since the pathogenicity of G. parasuis strains varies quite significantly, from highly pathogenic, such as the well-described Nagasaki strain (serotype 5 ST24), to non-pathogenic strains, such as strains of serotype 3, 6, 9, and 11 [5, 6], effective control hinges upon the correct identification and ty** of the disease-causing strains. Seroty** has been the most commonly used ty** method, with 15 serotypes based on capsular polysaccharides described to date [5, 7]. The major pitfall of seroty** is its low discriminatory power and typeabilty. Multilocus sequence ty** (MLST) [8, 9] is an alternative ty** technique. Although MLST provides better strain discrimination and portability compared to seroty**, it is labor-intensive and time-consuming, hindering adoption. A plethora of other GPS genoty** methods have been described previously for instance pulsed field gel electrophoresis (PFGE), amplified fragment length polymorphisms (AFLP), multilocus variable number of tandem repeat analysis (MLVA), random amplified polymorphic (RAPD), enterobacterial repetitive insertion consensus PCR (ERIC-PCR) [10,11,12,13,14,15]. Some of these methods have been compared previously [16]. Although most of these methods have relatively good discriminatory power, the results are hard to compare between laboratories, interpretation of the banding pattern can be ambiguous, and do not in general provide information on the virulence potential of a given strain.
Pathoty** is also available for G. parasuis. The most important virulence factors described for G. parasuis are the virulence-associated trimeric autotransporter (vtaA) genes [6]. There is increasing evidence that group 1 vtaA genes are associated with virulent isolates [17, 18]. While other putative virulence-associated genes, [19,20,21,22] have been described, these genes are not targeted frequently in diagnostic settings to assess the virulence potential of strains.
The reduction in sequencing costs and advances in bioinformatic pipelines have made whole genome sequencing (WGS) more feasible for diagnostic use. WGS offers higher discrimination between isolates by allowing the extraction of relevant information related to phylogeny, sequence type, putative virulence factors, and antimicrobial resistance (AMR) genes. Current knowledge about the molecular epidemiology of disease-associated G. parasuis strains circulating in North American pig populations is lacking. The objective of this study was to perform genetic characterization of G. parasuis strains from disease-associated cases using WGS. In-depth characterization using WGS could vastly improve on farm control and prevention programs.
Results
Phylogenetic analysis
At least 47,061,190 of the paired-end reads processed had a Phred score of 30, suggestive of high quality. The overall GC content was between 39.06 and 40.26. A total of 71,368 core genome SNPs were identified from all strains by kSNP3. The phylogenetic tree based on the core-genome SNPs of these isolates indicated high genetic diversity, and were clustered into three primary lineages (LI, LII, LIII) (threshold = 0.1) based on the structure of the phylogenetic tree and the number of virulence genes (Fig. 1). The largest primary lineage was LIII, containing 49.5% (n = 108) of the study isolates. LI and LII contained 13.8% (n = 30) and 36.2% (n = 79) of the isolates, respectively. Each lineage contained sub-lineages with only LIIIA sublineage being highlighted due to its unique characteristics (Fig. 2).

Core-genome phylogenetic tree of 218 sequenced genomes and 15 reference strains of G. parasuis isolates. A phylogenetic tree was constructed based on core-genome SNPs. Color shades indicate the primary lineages; LI, LII, and LIII. One isolate between LI and LII is an outlier

Distribution of selected virulence-associated genes. The phylogenetic tree was constructed based on core-genome SNPs. The red and green dots indicate the relative distribution of the vtaA genes and other putative virulence genes
Serotype, sequence type, and clonal complex
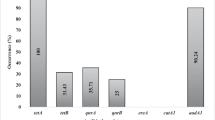
Among field isolates (n = 218), in silico whole-genome seroty** pipeline identified 11 of the 15 known G. parasuis serotypes (Fig. 3) with serotypes 9, 10, and 11 not being detected. A total of 6.4% (14/218) of isolates were untypable. The Simpson’s index of diversity for seroty** was estimated as 0.82. The most frequently detected serotypes were 7 (32.6%, 71/218), 13 (16.1%, 35/218), 2 (14.7%, 32/218), 4 (14.2%, 31/218), and 5/12 (7.3%, 16/218). Each lineage had unique predominant serotype (s). Within LI, LII, and LIII isolates, serotype 5/12 (50%, 15/30), 7 (75.9%, 60/79), and 13 (27.8%, 30/108), were the most frequently detected serotypes, respectively (Table 1). Consistent with the above distribution of serotypes within lineages, statistical analysis showed that serotype 5/12 was associated with LI, serotype 7 was associated with LII, and serotypes 2, 4, and 13 were associated with LIII (p < 0.05). In addition, LI and LII isolates were significantly associated with respiratory and systemic isolates, respectively (p < 0.05). A total of 80 isolates were isolated from tissues for which histopathological data was available. Of these, 8 did not have detectable lesions and were of serotypes 4, 7 and 13 with majority of pigs having lesions in other sites in which Streptococcus suis isolated. Among the 72 remaining isolates, the following lesions and serotypes were detected: polyserositis (n = 27) bronchopneumonia/pneumonia (n = 26) epicarditis/myocarditis (n = 9); meningitis/encephalitis (n = 3) arthritis/synovitis (n = 4), pleuritis (n = 2) splenitis (n = 1) (Additional file 3).

Distribution of sequence types (STs) and serotypes of Glaesserella parasuis (G. parasuis) isolates. The colour codes represent the sequence type diversity within in a given serotype. Frequently detected STs ST454, ST478, and ST6 are highlighted
Through multilocus sequence ty** analysis, we uncovered a total of 72 sequence types (STs) within the collection of 218 field isolates. Remarkably, among these, 66 STs were entirely new discoveries, representing 91.7% of the total STs identified, with ST454 emerging as the most commonly encountered one (Fig. 3, Additional file 3). The Simpson’s index of diversity (SDI) for MLST was 0.95. All reference strains typed as previously known STs, except serotype 10 (Additional file 3). In contrast with seroty** data, the majority of the STs clustered within the same lineage based on whole-genome SNP phylogenetic analysis. The majority of isolates belonged to a limited number of clonal complexes (CCs), which included CC157 (24.7%, n = 54), CC478 (8.3%, n = 18), CC413 (4.6%, n = 10), CC417 (3.7%, n = 8), CC56 (2.8%, n = 6), CC92 (2.8%, n = 6), CC452 (2.8%, n = 6), CC442 (2.3%, n = 5), CC438 (1.4%, n = 3), CC245 (0.9%, n = 2),), CC96 (0.9%, n = 2), CC433 (0.9%, n = 2), and CC246 (0.5%, n = 1). The most common CCs were CC157 and CC478 with CC157 comprising of STs, ST416, ST454, ST458, ST462, ST526, and ST157. CC478 was the second most frequently detected clone containing ST459, ST478, and ST624 (Additional file 1). ST478 is a single locus variant (SLV) of ST157 but formed its own clone (Additional file 1). Based on the group definition criterion, 43.4% (95/218) of the STs were considered singletons. ST6, ST454, and ST478 were associated with serotypes 5/12, 7, and 2, respectively (P < 0.05). However, ST6 strains were also from serotypes 13, ST454 of serotypes 4, 13 and 14, and ST478 of serotypes 7. A high ST diversity within serotypes was observed. For example, within serotype 7 strains, a total of 19 different STs were identified. Similar findings were observed with serotype 13 and 2. The untypable strains were of different sequence types.
Within the dataset, the three dominant STs were ST454 (19.3%, 42/218), ST478 (5.5%, 12/218), and ST6 (5%, 11/218). The remaining STs were each represented by ≤ 7 isolates, and 33/72 STs were represented by a single isolate. The two novel STs (454 and 478) are single locus variants (SLVs) of each other at the infB locus. Within LI and LII, ST6 (36.7%, 11/30) and ST454 (51.9%, 41/79) were the dominant STs, respectively, while LIII showed high diversity with 49 different STs. All six isolates in the LIII A sublineage (Fig. 2) were identified as novel STs, except for two serotype 8 isolates that were both ST299. ST6 was associated with LI (p < 0.05), and ST454 and ST478 were significantly associated with LII (p < 0.05). From cases with histopathology data, most ST454 and ST478 isolates originated from polyserositis cases.
Distribution of virulence-associated genes
A total of 212 (97.2%) isolates possessed at least one of the group 1 vtaA genes. Within group 1 vtaA genes, vtaA5 (88.1%, 192/218), vtaA6 (84.7%,185/218), vtaA3 (67.4%, 147/218), vtaA8 (66.5%, 145/218), and vtaA2 (65.1%, 142/218) were the most frequently detected (Additional file 3). The remaining group 1 vtaA genes were vtaA1, vtaA9, vtaA4 and vtaA7, with 58.7% (128/218), 53.2% (116/218), 43.1% (94/218) and 30.7% (67/218) presence, respectively. On average, LI isolates carried 6.8 (min: 2, max: 9) group 1 vtaA genes, LII isolates carried on average 6.2 (min: 4 max: 9) group 1 vtaA genes, and LIII isolates carried an average 5.02 (min: 1, max: 9) group 1 vtaA genes, excluding LIIIA. Sublineage LIIIA isolates lacked all vtaA genes (Fig. 2). VtaA3, vtaA8, and vtaA9 were significantly associated with LI and LII (p < 0.05). VtaA1, vtaA4, and vtaA7 were significantly associated with LI, and vtaA6 was associated with LIII.
A total of 23 isolates, predominantly within LI and LII, carried all 9 group 1 vtaA genes and were of serotypes 7 ST454, ST624, and ST632, serotype 5/12 ST6, ST427, and ST474, serotype 2 ST478 and ST548, serotype 4 ST549 and ST629, serotype 13 ST6, serotype 14 ST442, and NT ST427. From these 23 isolates, six also carried all the group 2 and 3 vtaA genes, and were of serotype 7 ST454 and ST624 and serotype 14 ST442. Some isolates represented by the same ST, carried a varied number of group 1 vtaA genes (Fig. 4).

Number of Group 1 vtaA genes by ST. Black line represents the range of Group 1 vtaA genes with blue dot identifying the median
Group 2 vtaA genes were found in the following proportions: vtaA10 (34.4%, 74/218) and vtaA11 (23.4% 51/218). Finally, within group 3 vtaA genes, vtaA12 was found in 56.4% (123/218) and vtaA13 in 61.5% of isolates, respectively (134/218).
Within serotypes, vtaA1 was associated with serotype 5/12, vtaA2 and vtaA7 were associated with serotype 2, and vtaA5, vtaA6 and vtaA8 were associated with serotype 7 (p < 0.05). Among the dominant STs, vtaA1, vtaA4, vtaA7, and vtaA9 were associated with ST6, and vtaA8 and vtaA9 were associated with ST454 (p < 0.05).
The prevalence of other putative virulence genes was further determined using whole-genome sequencing analysis. All isolates carried the porin protein ompP5 gene, and 56.9% (124/218) of the isolates were also positive for the porin protein gene ompP2. Fimbrial genes (pilA, pilB, and pilC) and siaB were present in 54.6% (119/218), 69.3% (151/218), 51.8% (113/218), and 84.4% (184/218) isolates, respectively. Among the monomeric autotransporter genes, bmaA5 (78.4%, 171/218), bmaA4 (51.3%,112/218), and bmaA6 (51.3% (112/218), were the most prevalent while bmaA1 (10.6%, 23/218) was the least prevalent. The lsgB gene was the least prevalent gene among the selected putative virulence genes, with a proportion of only 8.3% (18/218) of isolates being positive; all isolates in LIII lacked this gene. Almost all lsgB positive isolates (15/18) were part of L1 (Table 1), and were predominantly serotype 5/12.
Identification of antimicrobial resistance genes
Fourteen AMR genes were detected among the 218 genomes, encoding resistance to antibiotics of nine different classes including 2,5-diketopiperazines, aminoglycosides, beta-lactams, liconsamides, macrolides, lincosamide streptogramins B (MLS), polypeptides, quinolones, sulfonamides, and tetracyclines. Among these, bcr, ksgA, bacA, sul2, and aph (3'')-Ib were detected in almost all isolates included in the collection. The remaining genes were present at the following proportions; blaZ (6.9%, 15/218, tetM (3.7%, 8/218), spc (3.7%, 8/218), tetB (2.8%, 6/218), bla-ROB-1 (1.8%, 4/218), ermA (1.8%, 4/218), strA (1.4%, 3/218)), qnrB (0.5%, 1/218), and aph3''Ia (0.5%, 1/215).
Relationships between strains and geographical location, tissue, and disease
The distribution of G. parasuis isolates was further analyzed geographically. A total of 76% of isolates (19/25) from Mexico, 48% from the USA (74/154), and 35% from Canada (7/20) were grouped into LIII; 20% (5/25), 37% (57/154), and 30% (6/20) of the isolates from Mexico, USA, and Canada, respectively, clustered in LII. Finally, 4% (1/25), 14.2% (22/154), and 35% (7/20) of Mexico, the USA, and Canada isolates, respectively, were grouped into LI. A few isolates from Chile, Peru, and Denmark clustered in specific lineages (Additional file 3). Lung (n = 86) and systemic strains (n = 89) accounted for 80.3% of the isolates. The remaining were from URT (n = 8) or unknown (n = 35). Generally, there was no observed tissue-specific distribution within the lineages. In several isolates (n = 80) from tissues with reported histopathological lesions, serotypes associated with disease were 7, 13, 4, 5/12 and 2.
Discussion
G. parasuis continues to challenge the productivity, health, and well-being of post-weaning pig populations, and represents a driver of antimicrobial use on farms [1]. Effective control of G. parasuis requires a multifaceted approach based on the judicious use of antimicrobials, vaccination, minimizing viral co-infections and environmental triggers, improving husbandry and characterization of the disease-causing strains. Whole-genome sequencing analysis provides multiple layers of information that aid in determining the clinical relevance of the isolates and providing epidemiological insight.
In this study, WGS was performed on 218 G. parasuis isolates predominantly from North America. The size and GC content were within range with other published G. Parasuis genomes [23]. Phylogenetic analysis revealed three primary lineages, LI, LII and LIII. This is in contrast with a recently published study from China [23] that showed G. parasuis isolates clustering into two main primary lineages comprised of STs not found in the present study, highlighting the high genetic variation found within the species.
Whole-genome in silico seroty** revealed 11 of the 15 known serotypes, with serotypes 7, 13, 4, 2, and 5/12, as the most common serotypes detected. The WGS in silico seroty** scheme was based on the DNA sequence fragment for each serotype published in Howell et al.[24]. The scheme is unable to discriminate serotype 5 and 12. However, future comparative genomic analysis studies using complete genomes could be used to determine whether these two serotypes can be discriminated. In this study, only 6.4% of the isolates were untypable via in silico seroty**. This percentage was lower compared to 39% and 9.7% that utilized traditional and PCR based seroty** methods [10, 25], respectively, emphasizing the potential advantage of sequence-based methods for bacterial ty**. Furthermore, a real-time PCR seroty** scheme recently developed was able to assign a serotype to all 40 isolates that were previously untypable by conventional seroty** [26].
The most frequently detected serotypes observed in this study have been previously associated with disease and are commonly detected from clinical cases [10, 25, 27, 28]. However, in most studies serotypes 4 and 5/12 were the most frequently detected [10, 29,30,31]. This study further supports the increasing evidence for serotype 7 in disease [25, 31]. In a recent study, serotype 7 strains from clinical cases were frequently detected in USA, Canada, Europe, China, and Vietnam, and some were associated with virulence [25]. Its widespread nature could be attributed to trade between these countries or companies moving pigs between them.
Multilocus sequence ty** identified 72 sequence types within 218 isolates, of which 91.7% (66/72) were novel. Specifically, this study highlighted the emergence of ST454, a strain detected in outbreaks of high mortality, characterized by per acute disease and polyserositis. The eBurst analysis showed that ST454 belonged to the CC157 with its predicted founder ST157, a sequence type first identified in Brazil from a pig with bronchopneumonia (https://pubmlst.org/organisms/glaesserella-parasuis). Surprisingly, ST157 was among the least prevalent STs in this study. In contrast, a clonal complex’s ancestral ST (founder) is typically the most prevalent ST in a population due to the fitness advantage or random genetic drift [32]. It is likely that as the number of isolates in the database increases, the founder ST might change to ST454.
The eBurst analysis revealed that almost half of the isolates had singleton STs, suggesting high heterogeneity and instability in the population structure, as previously described [11, 17, 33]. The ST diversity observed in this study has also been reported in other studies. Olvera et al. identified 122 STs within 150 strains (17), Turni et al., identified 54 STs (41 novel) within 75 isolates from Australia [11]. In this study, only 5 of the 72 STs identified had been previously reported, further highlighting the significant heterogeneity of the species on a global scale. While a high diversity of STs was observed, only a few STs, including ST454, ST478, and ST6 were the most frequently detected in the current study. The high detection frequency of these STs could reflect unique production systems submitting samples to the diagnostic laboratory and not a true reflection of the overall prevalence. Still, the data shows the clear role of these STs in disease cases in the North America. U.S, but also their distribution in different countries and flows. For example, ST454 was detected in 24 distinct flows and 5 countries. Targeted control and elimination procedures, such as inclusion of these STs in autogenous vaccine products or depopulation of herds carrying such STs could potentially reduce the G. parasuis disease burden for a significant number of production systems. While the intensity of sampling varied across production systems, multiple flows had at least 2 STs (Additional file 2), supporting previous knowledge that within a production system multiple G. parasuis strains can be detected [34] and could be contributing to disease. Therefore, disease control should rely on the characterization of the clinically relevant strains within a flow to determine the appropriate vaccine candidates for autogenous vaccine production or to evaluate the potential efficacy of commercially available vaccines, as well as to improve the sourcing of replacement animals and pig flow management. In this study, while multiple serotypes and STs were detected within most flows, several flows were represented by a predominant ST or serotype (Additional file 1), particularly genotypes with apparent higher pathogenicity, such as with ST454.
Unlike serotypes, isolates of the same STs were mostly lineage specific, as revealed in the WGS-based phylogenetic analysis (Fig. 2). Furthermore, isolates of the same serotypes had multiple different STs but the opposite was observed less frequently, highlighting the higher discriminatory power of MLST compared to seroty**. Furthermore, core-genome SNP phylogenetic analysis showed evidence of genetic variation in isolates within the same STs e.g. ST6 and ST454 (Fig. 2), demonstrating the higher resolution offered by whole-genome sequencing compared to MLST and seroty**. Still, whether these genomic differences between strains of the same STs impact the clinical outcome is yet to be elucidated.
Among the virulence-associated genes, group 1 vtaA genes are good predictors of virulence, and some have been shown experimentally to play a role in disease [17, 18, 35, 36]. Approximately 97% of isolates from this study carried at least 1 group 1 vtaA gene. Based on previous research [25, 36], this suggests that these strains could have pathogenic potential. Phylogenetic SNP analysis revealed three primary lineages with varied number of average group 1 vtaA genes present. LI and LII isolates carried a numerically higher number of group 1 vtaA genes on average than LIII, whereas isolates from the LIIIA sublineage (Fig. 2) lacked all the vtaA genes. It is not entirely known if there is a correlation between the number of group 1 vtaA genes and increased virulence or if certain patterns of vtaA gene carriage can be a predictor of virulence. However, we observed some group 1 vtaA genes were significantly associated with known serotypes associated with disease for example 5/12 or 7, suggesting that these could serve as predictors of virulence. Isolates of serotype 8 and 15 in LIIIA lacked all vtaA genes and are serotypes that have been predominantly detected from the nasal cavity of pigs, confirming previous findings [25]. Variability in carriage of group 1 vtaA genes was noticed within isolates of the same ST or genetically similar based on SNP phylogenetic analysis (Fig. 2). For instance, group 1 vtaA gene carriage for 39 serotype 7 ST454 isolates ranged from 4–9. Similarly, all CC157 isolates (n = 54) carried between 4 and 9 group 1 vtaA genes. This shows how variable vtaA carriage can be even between strains showing high similarity at the core -genome level. The lack of detection of group 3 vtaA genes 12 and 13 in 44% and 33% of isolates, respectively, was an unexpected finding since previous studies have identified these genes among all G. parasuis strains previously examined but this was based on the translocator domain PCR (17). Some VtaA12 and vtaA13 might be divergent and therefore not identified as vtaA12 or vtaA13 homologs. In this study, genes were only reported if their sequence identity was above 70% and the gene coverage was above 50%. Previous studies have shown divergence within these genes which could have contributed to a lack of detection given our stringent definition for gene reporting (V. Aragon, unpublished data).
Monomeric autotransporters, porin proteins, and fimbria are involved in G. parasuis pathogenesis [21, 37, 38]. At least 51% of the isolates were positive for bmaA4, bmaA5, bmaA6, ompP2, ompP5, pilA, pilB, pilC and siaB in the current study. In fact, even isolates that were negative for all vtaA genes were positive for some of these, such as ompP5 and siaB. This may suggest that some of these putative virulence genes are conserved in both virulent and nonvirulent strains and may not be good predictors of virulence compared to group 1 vtaA genes. However, this notion should be interpreted with caution since the study isolates were predominantly from clinical cases. lsgB, which has been associated with virulent strains [20], was detected in only 8.3% of the isolates which were predominantly of serotype 5/12, suggesting a potential marker for this serotype.
Control and prevention of Glässer’s disease often rely on the strategic use of antimicrobials. Thus, detection of antimicrobial resistance genes could provide value in antimicrobial use decision-making. Among the genes, bcr (bicyclomycin resistance), ksgA (kasugamycin resistance), bacA (bacitracin resistance), sul2 (sulfonamide resistance, and aph (3’’)-Ib (streptomycin resistance) were detected in almost 100% of the isolates. Consistent with this study, bcr, ksgA, and bacA genes were highly prevalent in G. parasuis isolates in a recent study [23]. Some of these highly prevalent genes encode resistance to antibiotics not commonly used in swine production. A recent study showed that tetracyclines, lincosamides, and beta-lactams are among the top three antibiotic classes used in the USA [39]. Furthermore, susceptibility data from Iowa State University shows most 2021 G. parasuis strains (n = 908) as mostly susceptible to ampicillin, ceftiofur, enrofloxacin, florfenicol, tiamulin and tilmicosin (https://vetmed.iastate.edu/sites/default/files/VDL/pdf/Susceptibility-Summary-Porcine-2021.pdf). The remaining genes; aph3’’Ia, strA, blaROB-1, blaZ, spc, ermA, tetB, tetM and qnrB were detected in lower frequencies. Some studies showed varying frequencies of these genes in G. parasuis [40, 41]. Due to the low prevalence of some of these AMR genes a definitive conclusion on their distribution in phylogenetic lineages could not be determined. However, they were detected more frequently in LIII than LI and LII isolates (Table 1). For instance, 73.3% (11/15) and 75% (6/8) of the isolates positive for blaZ and tetM, respectively, were in LIII, and were of serotypes 1 ST417 and ST420, 2 ST476, 4 ST418, 7 ST471 and ST473, 13 ST415 and ST421, and NT ST415. Further studies are needed to correlate if the genotypic antimicrobial resistance profile can predict phenotypic resistance in this species.
Although the findings of this study were consistent with the published literature to date, caution is needed when interpreting the results. This is because the G. parasuis isolates included in the current study were from disease diagnosis cases submitted to the ISU VDL, and thus only captured production systems that submit samples to this laboratory. However, the contributing production systems represented the top pork producers in North America.
Conclusion
While WGS is computational demanding, requires high technical skill and is currently more expensive than other ty** tools, the value obtained with the information far exceeds these limitations. Still, it is expected that seroty** and virulence marker PCRs will continue to be used to monitor G. parasuis strain variation. Results herein will lead to a better understanding of the molecular epidemiology and virulence potential in G. parasuis that can be applied to the development of improved diagnostic tools, evaluation of antibiotic use, tracking of outbreak strains and identification of vaccine candidates for improved management of Glässer’s disease in swine production systems.
Methods
Source of isolates
A total of 218 G. parasuis isolates were obtained from porcine cases submitted to the Iowa State University Veterinary Diagnostic Laboratory (ISU VDL) from 2015 through 2022 (Additional file 3). While strains mostly originated from the USA (n = 154), they were also obtained from Canada (n = 20), Chile (n = 9), Denmark (n = 4), Peru (n = 6), and Mexico (n = 25). (Additional file 3). For each isolate, metadata was obtained, including; the year of isolation, tissue type, flow and farm name, and pig age. Strains from the USA originated from at least 16 different states. Histopathological data was obtained for 80/218 isolates (Additional file 3).
In addition, a total of 15 reference strains were provided by Dr. Marcelo Gottschalk, each representing one of the 15 G. parasuis serotypes [5]. All presumptive G. parasuis strains isolated at the ISU VDL or submitted as pure cultures from VDL clients were subjected to matrix-assisted laser desorption ionization-time-of flight mass spectrometry (MALDI-TOF MS) for definitive species identification prior to sequencing.
Whole genome sequencing
Initial isolation of G. parasuis from samples was achieved by plating clinical samples onto 5% sheep blood (Hardy Diagnostics, Santa Maria, CA) with Staphylococcus hyicus nurse streak and incubating overnight at 370C. Presumptive G. parasuis isolates were subcultured onto chocolate agar (Thermo scientific™), confirmed using MALDI-TOF MS, and frozen at -800C in brain heart infusion broth with 30% glycerol. Two days before submission for sequencing, bacterial strains were retrieved from the freezer and streaked onto chocolate agar. An overnight pure growth culture lawn was inoculated into 2 mL phosphate-buffered saline (PBS). Genomic DNA was extracted using ChargeSwitch gDNA mini bacterial kit (Life Technologies Carlsbad, CA) according to the manufacturer’s instructions [42].
The DNA quality (A280/A260) was assessed using a Nanodrop (Thermo scientific™) and quantified using a Qubit fluorometer dsDNA HS kit (Life Technologies). Multiplex genome libraries were prepared using the Nextera XT DNA library preparation kit (Illumina, San Diego, CA). The genomic library was quantified using a Qubit fluorometer dsDNA HS kit (Life Technologies Carlsbad, CA) and normalized to the recommended amplification concentrations. The pooled libraries were sequenced on an Illumina Miseq sequencer using Miseq Reagent V3 for 600 cycles (Illumina, San Diego, CA). Raw reads were demultiplexed automatically on the Miseq.
Raw sequence reads were pre-processed using Trimmomatic VO.36 [43] to remove adaptors, trim poor quality ends, and delete short sequences (< 36nt). Raw and pre-processed reads were assessed for quality to ensure cleaning efficiency using FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc). Pre-processed reads were assembled using SPAdes Genome Assembler version 3.11.1-Linus [44] using assembly options for paired-end reads and Burrows-Wheeler Aligner mismatch correction. Small (< 500nt) and low (< 2) average kmer coverage contigs were removed from the SPAdes assembly results from further analysis using custom scripts to determine N50, longest contigs, and total length of the contigs.
Bioinformatic analysis
Virulence-associated genes in each genome were detected using SRST2 [45] against a custom virulence gene database using raw reads. Blastn was used to detect highly divergent virulence-associated genes. Antimicrobial resistance genes in each isolate were detected using SRST2 [45] with default parameters (–min_coverage, 90%, –min_depth 5) by map** reads from each isolate against the acquired antimicrobial resistance gene database (/data/ARGannot_r3.fasta) provided by SRST2. In addition, the assembled contigs were also blasted against the same antimicrobial resistance gene database, virulence gene database, and candidate hits with identity above 70% and gene coverage above 50% were also reported and merged with the SRST2 results. Serotype was determined by detecting serotype-specific capsule loci [24]. Single nucleotide polymorphisms (SNPs) of G. parasuis isolates were identified by running kSNP3 with standard mode. The optimal k-mers size was calculated by the Kchooser program, and the whole-genome phylogeny was analyzed based on identified core genome SNPs [46].
Multilocus sequence ty** was performed using SRST2 with an integrated MLST database and definitions. Sequence types (STs) were further confirmed by querying the assembled fasta files using the G. parasuis pubMLST ty** platform [9, 47]. Assembled fasta files of isolates with novel alleles or allelic profiles were submitted to the PubMLST database. Novel alleles and allelic profiles were extracted from the assembled files and assigned a number using the G. parasuis ty** database. Novel ST were added to the pubMLST G. parasuis database with their corresponding metadata. Determination of clonal complexes was done using the goeburst algorithm [48], a refinement of eBURST [32], implemented in PHYLOViZ (http://www.phyloviz.net), using the stringent six of 7 alleles identical with the founder ST [49]. The global pubMLST G. parasuis dataset and the study isolates were subjected to the goeBURST analysis to determine the study isolates' evolutionary descent and clonal complexes. A clonal complex (CC) was defined if it contained at least three STs, including the founder ST. Sequence types that did not meet this criterion were considered singleton STs.
A core-genome single nucleotide polymorphism (SNP) phylogenetic tree was generated by Parsnp V1.2 [50] and visualized by the online tool iTOL [51].
Data analysis
Data management, descriptive, and inferential analyses of G. parasuis serotypes, STs, AMR, and virulence genes were performed in Microsoft Excel® and R (R program version 4.0.0, R core team 2020).
Simpson's index of diversity [52] was used to measure the diversity of genetic type distributions, which included serotype and ST. The calculations followed the formula:
where N was the total number of isolates, and n was the total number of isolates per serotype or ST.
Tissues of isolation, when reported, were classified as lung, upper respiratory tract (URT), systemic tissues or unknown. Likewise, reported lesions were categorized by affected body system, including nervous, respiratory, systemic, and unknown.Associations between serotype/ST, lineages, vtaA genes (1 – 9) versus tissue of isolation, and ST/serotypes versus lineages and vtaA genes (1 – 9) were evaluated using logistic regression. To account for the statistical model variation of logistic regression, only serotypes and STs containing at least ten isolates, representing at least 10% of the isolates, were selected for association analyses, which included serotypes 7, 13, 4, 5/12, 2, and NT, and STs 6, 454 and 478. Using Poisson Regression, the total number of Group 1 VtaA genes (1—9) was compared across serotypes (or ST).
Availability of data and materials
The genome assembly data and raw read sequences were deposited in the National Center for Biotechnology Information (NCBI) and Sequence Read Archive (SRA), respectively, under the Bio project number PRJNA749326. Other data is available in the additional files –-.
References
Aragon V, Segales J, Tucker AW. Glässer’s disease. In: Zimmerman JJ, Karriker LA, Ramirez A, Schwartz K, Stevenson GW, Zhang J, editors. Diseases of Swine. Ames: Wiley-Blackwell; 2019. p. 844–52.
Vahle JL, Haynes JS, Andrews JJ. Experimental reproduction of Haemophilus parasuis infection in swine: clinical, bacteriologic, and morphologic findings. J Vet Diagn Invest. 1995;7(4):476–80. https://doi.org/10.1177/104063879500700409.
Brockmeier SL, Register KB, Kuehn JS, Nicholson TL, Loving CL, Bayles DO, Shore SM, Phillips GJ. Virulence and draft genome sequence overview of multiple strains of the swine pathogen Haemophilus parasuis. PloS one. 2014;9(8):e103787. https://doi.org/10.1371/journal.pone.0103787.
Poeta Silva, AP, Schwartz K, Arruda B, Burrough E, Santos J, Macedo N, Sahin O, Harmon K, Siepker C, Gauger P, Sitthicharoenchai P, Rahe M, Magstadt D, Michael A, Pineyro P, Derscheid R, Main R, Fano E, Clavijo M. Diagnostic trends of five swine endemic bacterial pathogens using data from the Iowa State University Veterinary Diagnostic Laboratory (2010–2019). In proceedings: 2021; 52nd Annual Meeting of the American Association of Swine Veterinarians (AASV), Atlanta, Georgia, USA., San Francisco, California, USA (Virtual meeting).
Kielstein P, Rapp-Gabrielson VJ. Designation of 15 serovars of Haemophilus parasuis on the basis of immunodiffusion using heat-stable antigen extracts. J Clin Microbiol. 1992;30(4):862–5. https://doi.org/10.1128/jcm.30.4.862-865.1992.
Pina S, Olvera A, Barceló A, Bensaid A. Trimeric autotransporters of Haemophilus parasuis: generation of an extensive passenger domain repertoire specific for pathogenic strains. J Bacteriol. 2009;191(2):576–87. https://doi.org/10.1128/JB.00703-08.
Tad**e M, Mittal KR, Bourdon S, Gottschalk M. Development of a new serological test for seroty** Haemophilus parasuis isolates and determination of their prevalence in North America. J Clin Microbiol. 2004;42(2):839–40. https://doi.org/10.1128/JCM.42.2.839-840.2004.
Oliveira A, Cerdà-Cuéllar M, Aragon V. Study of the population structure of Haemophilus parasuis by multilocus sequence ty**. Microbiology. 2006;152(12):3683–90. https://doi.org/10.1099/mic.0.29254-0.
Mullins MA, Register KB, Brunelle BW, Aragon V, Galofré-Mila N, Bayles DO, Jolley KA. A curated public database for multilocus sequence ty** (MLST) and analysis of Haemophilus parasuis based on an optimized ty** scheme. Vet Microbiol. 2013;162(2–4):899–906. https://doi.org/10.1016/j.vetmic.2012.11.019.
Castilla KS, de Gobbi DD, Moreno LZ, Paixão R, Coutinho TA, dos Santos JL, Moreno AM. Characterization of Haemophilus parasuis isolated from Brazilian swine through seroty**. AFLP and PFGE Res Vet Sci. 2012;92(3):366–71. https://doi.org/10.1016/j.rvsc.2011.04.006.
Turni C, Singh R, Blackall PJ. Virulence-associated gene profiling, DNA fingerprinting and multilocus sequence ty** of Haemophilus parasuis isolates in Australia. Aust Vet J. 2018;96(6):196–202.
Boerlin P, Poljak Z, Gallant J, Chalmers G, Nicholson V, Soltes GA, MacInnes JI. Genetic diversity of Haemophilus parasuis from sick and healthy pigs. Vet Microbiol. 2013;167(3–4):459–67. https://doi.org/10.1016/j.vetmic.2013.07.028.
Zehr ES, Lavrov DV, Tabatabai LB. Comparison of Haemophilus parasuis reference strains and field isolates by using random amplified polymorphic DNA and protein profiles. BMC Microbiol. 2012;12(1):1–4. https://doi.org/10.1186/1471-2180-12-108.
Rafiee M, Bara M, Stephens CP, Blackall PJ. Application of ERIC PCR for the comparison of isolates of Haemophilus parasuis. Aust Vet J. 2000;78(12):846–9. https://doi.org/10.1111/j.1751-0813.2000.tb10507.x.
Zhang J, Xu C, Guo L, Ke B, Ke C, Zhang B, Deng XL, Liao M. A rapid pulsed-field gel electrophoresis method of genoty** Haemophilus parasuis isolates. Lett Appl Microbiol. 2011;52(6):589–95. https://doi.org/10.1111/j.1472-765X.2011.03048.x.
Olvera A, Segalés J, Aragón V. Update on the diagnosis of Haemophilus parasuis infection in pigs and novel genoty** methods. Vet J. 2007;174(3):522–9. https://doi.org/10.1016/j.tvjl.2006.10.017.
Olvera A, Pina S, Macedo N, Oliveira S, Aragon V, Bensaid A. Identification of potentially virulent strains of Haemophilus parasuis using a multiplex PCR for virulence-associated autotransporters (vtaA). Vet J. 2012;191(2):213–8. https://doi.org/10.1016/j.tvjl.2010.12.014.
Galofré-Milà N, Correa-Fiz F, Lacouture S, Gottschalk M, Strutzberg-Minder K, Bensaid A, Pina-Pedrero S, Aragon V. A robust PCR for the differentiation of potential virulent strains of Haemophilus parasuis. BMC Vet Res. 2017;13(1):1–5.
Mullins MA, Register KB, Bayles DO, Loving CL, Nicholson TL, Brockmeier SL, Dyer DW, Phillips GJ. Characterization and comparative analysis of the genes encoding Haemophilus parasuis outer membrane proteins P2 and P5. J Bacteriol. 2009;191(19):5988–6002.
Martínez-Moliner V, Soler-Llorens P, Moleres J, Garmendia J, Aragon V. Distribution of genes involved in sialic acid utilization in strains of Haemophilus parasuis. Microbiol. 2012;158(8):2117–24. https://doi.org/10.1099/mic.0.056994-0.
Costa-Hurtado M, Aragon V. Advances in the quest for virulence factors of Haemophilus parasuis. Vet J. 2013;198(3):571–6. https://doi.org/10.1016/j.tvjl.2013.08.027.
Wang H, Liu L, Cao Q, Mao W, Zhang Y, Qu X, Cai X, Lv Y, Chen H, Xu X, Wang X. Haemophilus parasuis α-2, 3-sialyltransferase-mediated lipooligosaccharide sialylation contributes to bacterial pathogenicity. Virulence. 2018;9(1):1247–62. https://doi.org/10.1080/21505594.2018.1502606.
Wan X, Li X, Osmundson T, Li C, Yan H. Whole-genome sequence analyses of Glaesserella parasuis isolates reveals extensive genomic variation and diverse antibiotic resistance determinants. PeerJ. 2020;8:e9293. https://doi.org/10.7717/peerj.9293.
Howell KJ, Peters SE, Wang J, Hernandez-Garcia J, Weinert LA, Luan SL, Chaudhuri RR, Angen Ø, Aragon V, Williamson SM, Parkhill J. Development of a multiplex PCR assay for rapid molecular seroty** of Haemophilus parasuis. J Clin Microbiol. 2015;53(12):3812–21. https://doi.org/10.1128/JCM.01991-15.
Macedo N, Gottschalk M, Strutzberg-Minder K, Van CN, Zhang L, Zou G, Zhou R, Marostica T, Clavijo MJ, Tucker A, Aragon V. Molecular characterization of Glaesserella parasuis strains isolated from North America, Europe and Asia by seroty** PCR and LS-PCR. Vet Res. 2021;52(1):1–10. https://doi.org/10.1186/s13567-021-00935-9.
Cui Y, Guo F, Cai X, Cao X, Guo J, Wang H, Yang B, Zhou H, Su X, Blackall PJ, Xu F. Ct value-based real time PCR seroty** of Glaesserella parasuis. Vet Microbiol. 2021;254:109011. https://doi.org/10.1016/j.vetmic.2021.109011.
Jia A, Zhou R, Fan H, Yang K, Zhang J, Xu Y, Wang G, Liao M. Development of serotype-specific PCR assays for ty** of Haemophilus parasuis isolates circulating in southern China. J Clin Microbiol. 2017;55(11):3249–57. https://doi.org/10.1128/JCM.00688-17.
Ma L, Wang L, Chu Y, Li X, Cui Y, Chen S, Zhou J, Li C, Lu Z, Liu J, Liu Y. Characterization of Chinese Haemophilus parasuis isolates by traditional seroty** and molecular seroty** methods. PLoS One. 2016;11(12):e0168903. https://doi.org/10.1371/journal.pone.0168903.
Turni C, Blackall PJ. Serovar profiling of Haemophilus parasuis on Australian farms by sampling live pigs. Aust Vet J. 2010;88(7):255–9. https://doi.org/10.1111/j.1751-0813.2010.00592.x.
Wang Z, Zhao Q, Wei H, Wen X, Cao S, Huang X, Wu R, Yan Q, Huang Y, Wen Y. Prevalence and seroepidemiology of Haemophilus parasuis in Sichuan province. China PeerJ. 2017;5:e3379. https://doi.org/10.7717/peerj.3379.
Schuwerk L, Hoeltig D, Waldmann KH, Strutzberg-Minder K, Valentin-Weigand P, Rohde J. Seroty** and pathoty** of Glaesserella parasuis isolated 2012–2019 in Germany comparing different PCR-based methods. Vet Res. 2020;51(1):1–4. https://doi.org/10.1186/s13567-020-00862-1.
Feil EJ, Li BC, Aanensen DM, Hanage WP, Spratt BG. eBURST: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence ty** data. J Bacteriol. 2004;186(5):1518–30. https://doi.org/10.1128/JB.186.5.1518-1530.2004.
Wang L, Ma L, Liu Y, Gao P, Li Y, Li X, Liu Y. Multilocus sequence ty** and virulence analysis of Haemophilus parasuis strains isolated in five provinces of China. Infect Genet Evol. 2016;44:228–33. https://doi.org/10.1016/j.meegid.2016.07.015.
Cerdà-Cuéllar M, Naranjo JF, Verge A, Nofrarias M, Cortey M, Olvera A, Segalés J, Aragon V. Sow vaccination modulates the colonization of piglets by Haemophilus parasuis. Vet microbiol. 2010;145(3–4):315–20.
Costa-Hurtado M, Ballester M, Galofré-Milà N, Darji A, Aragon V. VtaA8 and VtaA9 from Haemophilus parasuis delay phagocytosis by alveolar macrophages. Vet Res. 2012;43(1):1–10. https://doi.org/10.1186/1297-9716-43-57.
Olvera A, Pina S, Pérez-Simó M, Oliveira S, Bensaid A. Virulence-associated trimeric autotransporters of Haemophilus parasuis are antigenic proteins expressed in vivo. Vet Res. 2010;41(3):26. https://doi.org/10.1051/vetres/2009074.
Zhang NZ, Chu YF, Gao PC, Zhao P, He Y, Lu ZX. Immunological identification and characterization of extracellular serine protease-like protein encoded in a putative espP2 gene of Haemophilus parasuis. J. Vet. Med. 2012a; 1203180795. https://doi.org/10.1292/jvms.11-0260
Zhang B, Feng S, Xu C, Zhou S, He Y, Zhang L, Zhang J, Guo L, Liao M. Serum resistance in Haemophilus parasuis SC096 strain requires outer membrane protein P2 expression. FEMS Microbiol Lett. 2012;326(2):109–15. https://doi.org/10.1111/j.1574-6968.2011.02433.x.
Davies PR, Singer RS. Antimicrobial use in wean to market pigs in the United States assessed via voluntary sharing of proprietary data. Zoonoses Public Health. 2020;67:6–21. https://doi.org/10.1111/zph.12760.
Zhao Y, Guo L, Li J, Huang X, Fang B. Characterization of antimicrobial resistance genes in Haemophilus parasuis isolated from pigs in China. PeerJ. 2018;6:e4613. https://doi.org/10.7717/peerj.4613.
Van CN, Zhang L, Thanh TV, Son HP, Ngoc TT, Huang Q, Zhou R. Association between the phenotypes and genotypes of antimicrobial resistance in Haemophilus parasuis isolates from Swine in Quang Binh and Thua Thien Hue Provinces. Vietnam Eng. 2020;6(1):40–8. https://doi.org/10.1016/j.eng.2019.10.014.
Chen X, Resende-De-Macedo N, Sitthicharoenchai P, Sahin O, Burrough E, Clavijo M, Derscheid R, Schwartz K, Lantz K, Robbe-Austerman S, Main R. Genetic characterization of Streptococcus equi subspecies zooepidemicus associated with high swine mortality in the United States. Transbound Emerg Dis. 2020;67(6):2797–808. https://doi.org/10.1111/tbed.13645.
Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinform. 2014;30(15):2114–20. https://doi.org/10.1093/bioinformatics/btu170.
Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, Lesin VM, Nikolenko SI, Pham S, Prjibelski AD, Pyshkin AV. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012;19(5):455–77. https://doi.org/10.1089/cmb.2012.0021.
Inouye M, Dashnow H, Raven LA, Schultz MB, Pope BJ, Tomita T, Zobel J, Holt KE. 2014. SRST2: rapid genomic surveillance for public health and hospital microbiology labs. Genome Med. 2014;6(11):1–6.
Gardner SN, Slezak T, Hall BG. kSNP3. 0: SNP detection and phylogenetic analysis of genomes without genome alignment or reference genome. Bioinform. 2015;31(17):2877–8. https://doi.org/10.1093/bioinformatics/btv271.
Jolley KA, Bray JE, Maiden MC. Open-access bacterial population genomics: BIGSdb software, the PubMLST. org website and their applications. Wellcome Open Res. 2018;3:24. https://doi.org/10.12688/wellcomeopenres.14826.1.
Francisco AP, Bugalho M, Ramirez M, Carriço JA. Global optimal eBURST analysis of multilocus ty** data using a graphic matroid approach. BMC Bioinform. 2009;10(1):1–15. https://doi.org/10.1186/1471-2105-10-152.
Spratt BG, Hanage WP, Li B, Aanensen DM, Feil EJ. Displaying the relatedness among isolates of bacterial species–the eBURST approach. FEMS Microbiol Lett. 2004;241(2):129–34. https://doi.org/10.1016/j.femsle.2004.11.015.
Treangen TJ, Ondov BD, Koren S, Phillippy AM. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014;15(11):1–5.
Letunic I, Bork P. Interactive tree of life (iTOL) v3: an online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016;44(W1):W242–5. https://doi.org/10.1093/nar/gkw290.
Hunter PR, Gaston MA. Numerical index of the discriminatory ability of ty** systems: an application of Simpson’s index of diversity. J Clin Microbiol. 1988;26(11):2465–6. https://doi.org/10.1128/jcm.26.11.2465-2466.1988.
Acknowledgements
We acknowledge the Iowa State University Veterinary Diagnostic Laboratory Bacteriology section for their invaluable work carrying out the bacteriological culture of all study isolates. We thank the veterinarians and swine production systems for the sharing of isolates.
Funding
This study was funded in part by PIC North America.
Author information
Authors and Affiliations
Contributions
Conceptualization: M.C, Robert Mugabi R.M, A.P, G.L, V.A, M.G, N.M, O.S, P.H, A.T. Methodology and design: M.J, Robert Mugabi R.M, M.G, V.A, G.L, A.T, Rodger Main R.Main. Data collection: Robert Mugabi R.M, M.C, M.G, P.H, G.L, X.H. Statistical analysis: A.P, Robert Mugabi R.M, M.C, X.H. Writing original draft: Robert Mugabi R.M, M.C. Writing Review and Editing: All authors. All authors read and approved the final manuscript
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Isolates used in this study were from submitted clinical cases to ISU-VDL, so IACUC approval was unnecessary. Farm information and any identifying information were coded or assigned unique IDs to not be traced to the farm or flow.
Consent for publication
Not application.
Competing interests
The authors state no conflict of interest, with the following exceptions: PIC North America provided partial funding for the study and employs Drs. Clavijo (part-time) and Harms. Dr. Tucker serves as a consultant for PIC.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Mugabi, R., Silva, A.P.S.P., Hu, X. et al. Molecular characterization of Glaesserella parasuis strains circulating in North American swine production systems. BMC Vet Res 19, 135 (2023). https://doi.org/10.1186/s12917-023-03698-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12917-023-03698-x