
Abstract
Litter decomposition produces labile and recalcitrant forms of dissolved organic matter (DOM) that significantly affect soil carbon (C) sequestration. Chemical analysis of this DOM can provide important knowledge for understanding soil DOM dynamics, but detailed molecular analyses on litter derived DOM are scarce. Here we use ultrahigh resolution mass spectrometry (FT-ICR MS) to characterize the molecular composition of DOM from fresh and progressively decomposed litter samples. We compared high reactive (HR) and low reactive (LR) litter sources with regard to changes in the chemistry and bioavailability of leachates throughout the early phase of litter decay. We show that litter reactivity is a driver of chemical changes in the leached DOM of litter species. Birch, alder and Vaccinium (i.e. HR) litter initially produced more DOM with a higher lability than that of spruce, pine and wood (i.e. LR) litter. Labile oxidized phenolic compounds were abundant in leachates produced during the initial HR litter decay stages, indicating litter lignin degradation. However, the similarity in chemistry between HR and LR leachates increased during the litter decay process as highly leachable structures in HR litter were depleted. In contrast, chemistry of leachates from LR litter changed little during the litter decay process. The oxygenated phenolic compounds from HR litter were driving the lability of HR leachates and the changes in relative abundance of molecules during DOM incubation. This appeared to result in the creation of stable aliphatic secondary microbial compounds. In LR leachates, lability was driven by labile aliphatic compounds, while more resistant phenolic compounds were associated with recalcitrance. These results show how DOM dynamics follow different paths depending on litter reactivity, which has important implications for soil biogeochemistry and C sequestration.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Boreal forests cover nearly 15% of the earth’s land surface (Melillo et al. 1993; Gower et al. 2001) and contain as much as 370–1700 Pg of carbon (C), stored to ~ 95% below ground (Bradshaw and Warkentin 2015). Of the total net primary production (NPP) in boreal forests, 20–60% contributes to litterfall (Chen et al. 2017) and, thus, to subsequent decomposition on the forest floor, where 7–40% of litter C loss leaches as dissolved organic matter (DOM) (Soong et al. 2015). Through infiltrating rainfall and snowmelt water, this DOM is conveyed through soils where it plays a key role in controlling soil formation and C sequestration (Kalbitz et al. 2000, 2005; Cotrufo et al. 2015). Thus, DOM is an important link between NPP and C storage in boreal regions.
Both labile and recalcitrant fractions of DOM can lead to the formation of persistent soil organic matter (SOM) (Grandy and Neff 2008; Prescott 2010; Cotrufo et al. 2015). Labile DOM is an important microbial C source contributing to the production of secondary microbial products (Kalbitz et al. 2000; Bowen et al. 2009), which have been shown to form the most stable and persistent fraction of SOM (Marschner and Kalbitz 2003; Miltner et al. 2012). The portion of DOM that is more recalcitrant, on the other hand, consists partly of aromatic molecules with high affinity for stabilization through sorption (Kalbitz et al. 2005; Kleber et al. 2007). Although some labile compounds such as amino acids can also be prone to sorption (Kleber et al. 2007), the decrease in decay is smaller for sorbed labile DOM than recalcitrant DOM (Kalbitz et al. 2005). Thus, while the quantity of the litter to soil DOM flux is imperative to the global C cycle, the chemical composition of the DOM is key to understanding the mechanisms controlling the fate and preservation of soil DOM.
Litter chemistry has been proposed to be a key regulator of both initial rates of the mass loss and, specifically, of the loss of C through DOM leaching (Berg and Staaf 1980; Melillo et al. 1982, 1984; Aber et al. 1990; Cornelissen 1996; Pérez-Harguindeguy et al. 2000; Preston and Trofymow 2000; Soong et al. 2015; Campbell et al. 2016). Litter mass loss and leaching tend to go hand in hand, as early phase decay is faster in litter with a higher soluble content (Osono and Takeda 2005). Over time, soluble compounds like polyphenols, certain carbohydrates and non-lignified cellulose are exhausted, leading to decreasing rates of mass loss (Berg et al. 1982; Melillo et al. 1989; Lorenz et al. 2000, 2004). In addition, litter containing highly labile DOM compounds is subject to higher lignin degradation rates as a result of priming, with phenolic compounds released as byproducts (Klotzbücher et al. 2011b). Thus litter with rapid initial mass loss undergo greater changes in chemical composition and leach more DOM compared with litter that is initially more resistant to decay (Soong et al. 2015). There are good reasons to expect a corresponding difference in the composition of leachates between rapid and slow decaying litter species, but little is known about the chemical composition of DOM from different litter species and the changes throughout litter decay.
The characterization of litter-leached DOM has often not been detailed enough to provide a molecular understanding of the leaching dynamics and involved processes (Brock et al. 2020). For example, while it is well-documented that there are large variations in microbial bioavailability of DOM (BDOM) between DOM leached from different litter sources (Cleveland et al. 2004; Silveira 2011; Hensgens et al. 2020) and at different litter decay stages (Don and Kalbitz 2005; Hensgens et al. 2020), it is unknown if corresponding differences exist on a molecular composition level. This leaves the fundamental question of how BDOM relates to litter molecular DOM composition unanswered. At the same time, while soil science is largely moving away from chemical humification preservation models (Schmidt et al. 2011; Lehmann and Kleber 2015), knowledge about the chemical composition of litter derived DOM and the changes occurring during litter decay may prove instrumental in the development and testing of alternative soil C preservation models (Kaiser and Kalbitz 2012).
A variety of different techniques exist to measure the chemistry of DOM and are reviewed in (Minor et al. 2014). Fourier-transform ion cyclotron resonance mass spectrometry (FT-ICR MS) provides ultrahigh accuracy mass measurements and is widely used for DOM characterization (Nebbioso and Piccolo 2013). However, it requires ionization of the DOM molecules, and when combined with direct infusion measurements is prone to ionization artifacts like ion suppression and matrix effects (Nebbioso and Piccolo 2013). In addition, solid phase extraction (SPE), which is required for direct infusion FT-ICR MS does not recover all DOM and is known to selective enrichment of non-polar compounds from freshwater DOM (Raeke et al. 2016). Lastly, quantification of DOM compounds is not possible because there are currently no standards available for DOM. While other techniques, such as fluorescence spectroscopy, Fourier-transform infrared (FTIR) or nuclear magnetic resonance (NMR), can be used for quantification, they lack the (molecular) resolution to advance the field on the molecular scale. In this study, we used ultra-high resolution FT-ICR MS measurements on litter DOM leachates from eight different sources in a field litter decomposition experiment, with the goal to explore the dynamics in DOM chemistry as related to litter decomposition and to BDOM. The DOM was extracted from fresh, 10-, 48- and 180-day decomposed litter (Fig. 1). FT-ICR MS measurements were done before and after half-year DOM incubations, during which DOM concentrations were measured periodically and its reactivity and BDOM were calculated. We tested the idea that similar to how litter reactivity is known to govern the initial changes in litter chemistry, it also controls the corresponding initial changes in leachate chemical composition, with consequences for DOM bioavailability. In order to explore the effects of litter reactivity on DOM chemistry we conceptually divided the litter species into two separate groups based on litter mass loss and the solubility of litter C: ‘high reactive’ (HR) and ‘low reactive’ (LR) litter (Fig. 1). Early litter mass loss has been shown to be comparatively low in spruce and pine litter as compared to alder or birch litter (Berg and Ekbohm 1991). In addition, as early litter decomposition is inversely correlated with structural strength (Pérez-Harguindeguy et al. 2000), we assigned birch, alder and V. myrtillus, as HR litter and the more tough structured spruce, pine and wood litter as LR litter.

Conceptualizing high reactive (HR) and low reactive (LR) litter species based on litter mass loss rates and C solubility. Simplified data from (Hensgens et al. 2020). Dashed lines show mass loss and DOM leaching potential (mg C g−1) at 10, 48 and 180 days of litter decay for HR (yellow) and LR (blue) species. In reality, LR species could show either a slight decrease or a slight increase in leachable material depending on its source litter (Don and Kalbitz 2005; Hensgens et al. 2020)
The objective of this project was to compare HR (i.e. birch, alder and Vaccinium) and LR (i.e. spruce, pine and wood) litter species with regard to changes in DOM chemistry and bioavailability of leachates throughout the early phase of litter decay. More specific, we hypothesized that as a result of highly soluble material in HR litter at the start of litter decay (1.1) the DOM reactivity is higher in HR than LR litter species and (1.2) it decreased relatively more rapid for HR litter species during litter decay, as highly soluble and reactive material in parent litter is exhausted over time. Furthermore, we predict that (2.1) as litter decomposition progresses, leachate chemistry will show greater changes in HR as opposed to LR species, while (2.2) a higher degree of similarity is seen in leachate chemistry towards the later stages of litter decomposition. Finally, if the above hypotheses can be supported, we expect that fresh HR litter leachates contain unique chemical features that are linked to their higher overall reactivity. Based on our findings we discuss how our approach and results provide useful knowledge for a wide range of subjects from soil C respiration, SOM buildup and soil DOM leaching into the aquatic network.
Material and methods
Sample collection
Fresh litter samples were collected on September 25 and 28, 2015, in the Krycklan catchment in northern Sweden (Laudon et al. 2013) from the most abundant litter species: Picea abies (spruce), Pinus sylvestris (pine), Betula spp. (birch), Alnus spp. and Vaccinium myrtillus. For spruce, pine and birch species small branches were collected as wood samples. Litter samples were dried, weighed and put into the field for in situ litter decomposition. Litter samples were retrieved after 0-, 10-, 48- and 180-day field decomposition.
DOM extraction and sampling
Following collection, pure water extractions were carried out using a 48-h shaking (140 RPM) protocol at room temperature (Hensgens et al. 2020). After filtration (0.7 µm; Whatman GF/F) and dilution to 5.9 mg C l−1, samples were taken for FT-ICR MS analyses, acidified to pH ~ 2 using ultrapure HCl and immediately frozen (− 24 °C). In order to keep factors constant besides inherent C reactivity, and in line with the finding that soil heterotrophic communities are primarily carbon limited (Soong et al. 2020), the samples for microbial incubations were diluted to a final concentration of 6 mg C l−1, enriched with inorganic nutrients to achieve a C:N:P ratio above the Redfield ratio (25:5:1) and added the same inoculum (1% v/v of unfiltered Krycklan stream water). Incubations were done in a dark constant room at 20 °C in 400 ml HDPE bottles. After the end of the incubation (180 days), samples for FT-ICR MS analyses were taken, acidified and stored. All equipment had been acid washed and samples were kept in acid washed HDPE bottles. A more detailed description of the setup can be found in a companion paper (Hensgens et al. 2020), where DOM leaching is discussed in relation to litter production of the understory versus canopy species.
DOM decay dynamics
Dark incubations were initiated within two days from DOM extraction and DOM carbon concentrations measured after 0, 28 and 182 days of incubation. For fresh litter additional measurements were made after 14, 45 and 91 days. Parallel to this, 7-day dark incubations were started in 5 ml duplicates glass vials for each DOM solution using SDR (SensorDish Readers, PreSense) nonintrusive oxygen sensors for respiration monitoring every 2 h in a high temperature precision 20 °C climate chamber (Soares et al. 2018). The DOM was measured as total organic carbon (TOC) on 30–40 ml samples acidified using 50 µl of ultrapure (20%) HCl and stored at 5 °C before analysis performed on an OI analytical Aurora TOC analyzer.
Changes in dissolved oxygen concentrations (O2) were transformed to DOMt values according to Eq. 1, where Cmass is the molar mass of C and O2mass is the molar mass of dioxygen.
This assumes a respiratory quotient (RQ) of 1, close to RQs measured in soils (Müller et al. 2004; Dilly and Zyakun 2008). Short-term data was trimmed to 1 value per day and merged with long-term incubation measurements.
The reactivity continuum model (Eq. 2 was fitted on DOM concentrations measured throughout the incubation converted to relative DOM concentrations DOMt/DOM0 (Koehler 2012). Fitting was done using a general linear mixed effects (glme) model, with litter decay state as fixed and species as random factor.
FT-ICR MS methods
Sample preparation
Leachate samples (30–360 ml) were extracted via solid-phase extraction using an automated sample preparation system (FreeStyle, LC Tech, Obertaufkirchen, Germany) on 50 mg styrene-divinyl-polymer type sorbens (Bond Elut PPL, Agilent Technologies, Santa Clara, CA, United States) to desalt the sample for subsequent DI-ESI–MS according to Raeke et al. (2016). The SPE-DOM was eluted with 1 ml methanol (Biosolve, Valkenswaard, Netherlands), diluted to 20 ppm and mixed 1:1 (v/v) with ultrapure water immediately prior FT-ICR MS analysis. Carbon based extraction efficiency was approximately (38.9 ± 10.4)% (n = 62). SRFA measured in triplicate was used to check instrument variability and solvent and extraction blanks were prepared.
FT-ICR MS measurement
An FT-ICR mass spectrometer equipped with a dynamically harmonized analyzer cell (solariX XR, Bruker Daltonics Inc., Billerica, MA, USA) and a 12 T refrigerated actively shielded superconducting magnet (Bruker Biospin, Wissembourg, France) instrument was used in ESI negative ionization mode (capillary voltage: 5 kV). Extracts were analyzed in random order with an autosampler (infusion rate: 10 μl min−1). For each spectrum, 256 scans were co-added in the mass range 150—3000 m/z with 15 ms ion accumulation time and 4 MW time domain (resolution@400 m/z ca. 470,000). Mass spectra were internally re-calibrated with a list of peaks (247–609.2 m/z, n > 52) commonly present in terrestrial DOM and the mass accuracy after linear calibration was better than 0.11 ppm (n = 62). Peaks were considered if the signal-to-noise (S/N) ratio was greater than four. Raw spectra were processed with Compass DataAnalysis 4.1 (Bruker Daltonics Inc., Billerica, MA, USA).
FT-ICR MS data evaluation
Molecular formulas were assigned to peaks in the range 0–750 m/z allowing for elemental compositions C1–60 H0–122 N0–3 O0–40 S0–2 with an error range of ± 0.5 ppm according to Lechtenfeld and others 2014. Briefly, the following rules were applied: 0.3 ≤ H/C ≤ 2.5, 0 ≤ O/C ≤ 1, 0 ≤ N/C ≤ 1.5, 0 ≤ DBE ≤ 25 (double bound equivalent, DBE = 1 + 1/2 (2C–H + N) (Koch et al. 2014), − 10 ≤ DBE-O ≤ 10 (Herzsprung et al. 2014), and element probability rules proposed by Kind and Fiehn (2007). Isotopologue formulas (13C, 34S) were used for quality control but removed from the final data set as they represent duplicate chemical information. Relative abundance (RA) values of assigned peaks were calculated based on the sum of all assigned peaks in each mass spectrum.
The aromaticity index (AI) was calculated as [1 + c − o − s − 0.5 * (h + n)] / [c − o − s − n] (Koch and Dittmar 2006, 2016). In this article we refer to molecular formulas as compounds although each formula may comprise different isomers. Compounds classes were assigned according to Table 1. The RA values of formulas in each group were summed (%RA) and the difference of %RA values calculated between two samples (Δ%RA = [%RA(Day 10, 48, or 180) − %RA(Day 0)] / %RA(Day 0) for further evaluation.
Statistics
Statistics were performed using the R core program and specific statistical packages where stated (3.5.1. 2018). The Welch’s two sample one-sided t-test was used to test whether the DOM reactivity leached from HR litter was higher than that of LR litter in Fig. 3. Tests were done for each litter decomposition time. The two-sided Welch’s t-test was used to test which components changed significantly in RA during litter decomposition (Fig. 4). A non-dimetric dimensional scaling (NMDS) was performed using the vegan package on the normalized RA of individual formulas (Fig. 5). Linear regression modeling was done using the standard R functions on non-transformed (Fig. 4) and log–log transformed (Fig. 5a:b) data. Partial least squares (PLS) analysis was done using the pls and spls packages on HR and LR leachates separately with BDOM as predictor variable. First, only formulas occurring in 50% of the dataset were selected. For both models the 3 component model was selected. Variable selection was done using the sparse PLS method, a method designed to deal with multicollinearity often seen in large genetic datasets (Chun and Keleş 2010). For the final models, the O-scores method was used together with cross-validation. The directional effect of individual molecular formulas on BDOM of the leachates was extracted from the model and simplified to negative or positive for Fig. 8 and Table 3.
Results
Litter mass loss and DOM leaching
The cumulative mass loss of HR litter reached 4.1 ± 1.2%, 11.9 ± 4.1% and 20.6 ± 3.6% after 10, 48 and 180 days respectively. The mass loss of LR litter was relatively much lower at 0.8 ± 2.1%, 1.6 ± 2.1% and 6.2 ± 0.5% after 10, 48 and 180 days, respectively. Daily mass loss rates decreased from 0.4% day−1 during the first 10 days to 0.07% day−1 between day 48 and 180 for HR species. For LR species a decrease was seen from 0.08% day−1 during the first 10 days to 0.03% day−1 between day 48 and 180. DOM leaching for HR litter decreased from 81.7 ± 42 mg g−1 to 7.4 ± 3.9 mg g−1 for fresh and 180 day decomposed litter respectively. For LR litter DOM leaching was consistently low at an average of 4.9 ± 4.1 mg g−1 for fresh and 3.4 ± 2.5 mg g−1 for 180 decomposed litter. Ranges given depict the standard deviation.
DOM decay dynamics and BDOM
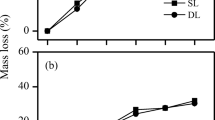
The DOM decay followed the reactivity continuum model (Eq. 2), showing high initial decay rates that decrease over time. The overall DOM reactivity was high across all fresh litter sources and decreased over time as litter was decomposed, with the exception of small initial increases after 10 days in some species (Fig. 2). Overall, only two out of three wood species had lower reactive DOM than non-wood litter species (Fig. 2). However, at the start of the DOM incubation the initial decay rate (k) of fresh and 10-day decomposed HR litter was significantly higher than that of LR litter samples (Fig. 3a). This difference disappeared as litter decay progressed. A similar, though less significant pattern was found for the BDOM after 180 days of incubation (Fig. 3b).

Fraction remaining DOM over incubation time as modelled by the reactivity continuum (Koehler et al. 2012) model. HR litter species are in yellow, LR in blue. Different lines denote different litter field decay times (0 to 180 days)

The decay coefficient k (a) and the total bioavailability of DOM (b) for HR and LR litter leachates related to litter decay. Significant differences between HR and LR leachates are denoted (*p < 0.05, •p < 0.1)
DOM chemistry
The FT-ICR MS analysis resulted in on average 2613 to 4270 assigned molecular formulas per timepoint (Table 2). Less formulas were assigned for HR than LR litter samples. The number of assigned formulas increased after DOM incubation, although only slightly for LR (Table 2). In HR litter the assigned formulas increased during litter decay, but for LR litter this was not seen. The double bond equivalent (DBE) of HR litter decreased during litter decay and the amount of nitrogen (N) containing formulas increased. In addition, after incubation the number of N containing formulas was larger (Table 2), while the average mass of formulas was slightly lower before incubation.
As an example, spruce wood (LR species) and Vaccinium litter (HR species) are plotted on van Krevelen plots (Fig. 4). Only few compounds increase in intensity during litter decay for Vaccinium, while most compounds decrease in intensity. For spruce wood an increase and decrease can be seen across the plot, although the increasing formulas are spread along a wider OC gradient. Differences in C:H:N:O:S are shown in supplementary Figs. 2 and 3.

Molecular formulas detected for the spruce wood (a, b) and Vaccinium (c, d) displayed by their molecular H/C versus O/C ratios and the change in normalized peak intensities (color coded ΔRI). Formulas with decreasing normalized intensity during litter decay (0—> 180 days) have ΔRI values < 0.45 (a, c), while formulas with increasing intensity have ΔRI values > 0.55 (b, d). Formulas with 0.45 < ΔRI < 0.55 are not shown
The relative abundance (RA) of compounds in leachate DOM samples was dominated by phenolic and aliphatic compounds (Fig. 5). Unsaturated and polyphenolic compounds were seen at lower RA followed by condensed aromatics. For HR litter, decomposition of the source material lead to a relative increase of aliphatic (p < 0.05, R2 = 0.33) and a decrease of high OC phenolic (p < 0.05, R2 = 0.36) RA (Fig. 5a). In a similar manner, no significant change was found in the composition of LR leachates. While for fresh (0 and 10 day decomposed) litter a significant difference was found between HR and LR aliphatic (p < 0.01), phenolic high OC (p < 0.01) and phenolic low OC (p < 0.05), in aged (48 and 180 day decomposed) litter only phenolic high OC (p < 0.05) compounds showed a significant difference.

Leachate DOM composition of HR (a) and LR (b) litter species at day 0 to day 180 of litter decomposition. Changes in relative abundance (RA) are shown as relative difference from Day 0 in color scheme with a max change of 100%
These results were consistent with a nonmetric dimensional scaling model (stress = 0.042 k = 2), which showed greater distance—and thus greater change in chemistry—between sequential HR litter decomposition samples than for the LR counterparts (Fig. 6). Leachates from more decomposed HR litter were in general in closer vicinity to LR litter samples, showing greater resemblance between HR and LR leachates as litter decomposed. Changes in position were correlated with changes in DOM leaching (p < 0.01) and litter mass loss (p < 0.05), showing that greater changes in chemistry occurred for litter species undergoing more rapid changes in litter structure (Fig. 6b, c). Further, it shows a relatively clear separation between LR litter with low leaching, mass loss and changes in chemistry and HR litter with high leaching, mass loss and changes in chemistry.

NMDS based on the relative abundancy of individual molecular formulas showing (a) the changes occurring as litter decomposes for HR and LR litter species and (b, c) the changes in chemistry as related to changes in (b) leaching and (c) mass loss during litter decomposition. Delta MDS is the total change in position of the data on both MDS axes
Changes in DOM chemistry through incubation
The RA of unsaturated and phenolic high OC compounds decreased for fresh HR litter while the RA of unsaturated and phenolic low OC and aliphatic compounds increased (Fig. 7a). For aged HR litter phenolic RA decreased less or only slightly increased, while aliphatic RA decreased (Fig. 7a). The RA of LR litter peaks was relatively stable for phenolic compounds and decreased for aliphatic compounds (Fig. 7b). Results for polyphenolic compounds were variable, while condensed aromatic compounds in general increased slightly in RA for both LR and HR species (Fig. 7).

DOM composition after incubation. Composition is shown for a HR and b LR litter at day 0 to 180 of litter decomposition. Changes in relative abundance (RA) are shown as a difference with pre-incubation DOM in color scheme with a max change of 100%. Grey bars show the pre-incubation RAs
The relationship between DOM chemical change and BDOM
For both HR and LR litter, a 3-component PLS model was selected using 86.93 and 94.59% of the variance in the FT-ICR MS data to explain 95.65 and 95.92% of the variance in BDOM respectively. Compounds which increased in RA with increased BDOM (Fig. 8a) were associated with either recalcitrance or microbial products formed during the decomposition process. Recalcitrance in this study is defined as molecular formulas that show a relative increased intensity over the incubation period. Lability is defined as molecular formulas that show a relative decreased intensity over the incubation period. Compounds which decreased in RA with increased BDOM (Fig. 8b) were associated with a higher relative lability. While only a marginal difference between HR and LR could be seen for recalcitrance associated molecules (Fig. 8a), a comparative clear difference was seen for molecules associated with lability, which showed relatively more oxygen compared to hydrogen and carbon (Fig. 8b). Both for HR and LR samples, molecules associated with recalcitrance were in general limited to low O and comparatively lower DBE molecules, while molecules associated with lability showed high O content with a large DBE range (Fig. 8c, d).

Results of the PLS models for high reactive (HR) and low reactive (LR) litter. a and b show molecular formulas indexed by their oxygen to carbon (OC) and hydrogen to carbon (HC) ratio. Shown are molecular formulas that a increased and b decreased in relative abundance (RA) with higher BDOM. E and F show the molecular formulas as related to the double bond equivalent (DBE) and number of O atoms for LR (a) and HR (f) litter species. The formulas (e, f) or compound class (c, d, g, h) that increased with higher BDOM are shown in brown (e, f, g, c), those that decreased in turquoise (e, f, d, h)
Recalcitrance was associated with aliphatic and unsaturated phenolic low OC compounds for HR litter (Fig. 8c). The HR PLS model was driven by compounds increasing in intensity over incubation rather than decreasing (Fig. 8c, d, Table 3). Lability was associated primarily with phenolic compounds and a lesser degree with aliphatic compounds. The LR PLS model was more equally driven by increasing and decreasing compounds than the HR model (Fig. 8g, h, Table 3). Here, low OC phenolic compounds were associated with recalcitrance and aliphatic and high OC phenolic compounds with lability.
Discussion
In line with our hypotheses, our results showed a clear difference in DOM lability between leachates from HR (i.e. birch, alder, Vaccinium) and LR (i.e. spruce, pine, wood) species. Specifically, we found a relatively higher DOM lability of HR litter that decreased as litter decay progressed, thereby eliminating the HR and LR difference (Fig. 3). Changes in DOM chemistry were positively related to changes in mass loss and DOM leaching (Fig. 6) and more profound in birch, alder and Vaccinium than spruce, pine and wood species over the same litter decay period. Towards the later stages of litter decay the chemistry of LR and HR leachates showed a higher degree of similarity (Fig. 5, 6). Over the DOM incubation period we saw clear differences in bioavailable compounds between fresh HR and LR samples (Fig. 7, 8). Furthermore, there was a high abundance of high OC phenolic compounds associated with the high reactivity seen in fresh HR litter. These compounds seem to be derivatives of increased lignin degradation in HR source litter. These findings show a diversity in leachate chemistry and dynamics from different litter sources that so far has been understudied and stress the importance of highly reactive litter in soil biochemical processes.
Litter leached DOM chemistry and litter decay
Phenolic high OC compounds were highest in fresh HR leachates and subsequently decreased as HR litter decay progressed. Oxidized phenolic compounds are a product of lignin degradation (Klotzbücher et al. 2011a) as lignin undergoes gradual oxidative transformations during the degradation process (Kögel-Knabner 2002). A decreasing yield of lignin-derived phenols is a sign of strong lignin degradation, that has been linked to a high supply of available C (Klotzbücher et al. 2011b). Alternatively, phenolic compounds can also be found in certain tannins (Lorenz et al. 2007) and a rapid decrease of tannins has been seen in spruce litter (Lorenz et al. 2000). However, it has been pointed out that the decrease in tannins found in the latter study might be an artifact due to the formation of non-extractable complexes or structural changes that prevent their detection as litter decay progresses (Kögel-Knabner 2002). In addition, tannins can have strong antimicrobial effects (Kögel-Knabner 2002). However, the highly oxidized phenolic compounds extracted in our fresh HR litter samples decreased significantly during DOM incubation. Thus, the oxidized phenolic compounds are likely lignin derivatives produced during early litter lignin degradation in HR litter as a result of the high C leaching rates.
While the RA of aliphatic and phenolic low OC compounds were found to increase during HR litter decay, this is arguably a secondary result of the decreasing RA of phenolic formulas with high OC and consequently increase of the other RAs. Aliphatic compounds have been shown to accumulate during litter decay (Hempfling et al. 1987; Zech et al. 1987; Bonanomi et al. 2015; Cepáková and Frouz 2015), indicating a lower relative leachability than other compounds. As litter decay progresses high leachable substrates are depleted first (Lorenz et al. 2004), resulting in a proportional increase in the RA of low leachable compounds such as aliphatics. This occurs in the context of quantitatively smaller litter DOM leachates, i.e. the total amount of DOM leached is smaller. As a result, in absolute terms, aliphatic compounds are still leached at higher quantities in fresh HR than decayed litter. For LR litter no such relationships were found. This indicates that no additional lignin degradation takes place due to high C availability as LR litter shows low C leaching rates throughout the decay experiment and because no preferential leaching of certain compounds takes place. It should be noted that these results show DOM dynamics during initial litter decay only. Although lignin has a chemical structure that makes it more resistant to decay, ultimately it is broken down and oxidized (Grandy and Neff 2008), possibly resulting in the leaching of lignin derivatives in later phases of the litter decay process.
Increased chemical similarity through microbial decomposition
Microbial processing of DOM leads to an increased chemical in soils (Strid et al. 2016; Thieme et al. 2019), rivers (Mosher et al. 2015) and oceans (Lechtenfeld et al. 2015; Mentges et al. 2017). Our results show that this process starts already at the primary source—aboveground with litter microbial processing increasing the similarity of leachates between litter types. Similarly, litter chemistry converges as litter decay furthers (Preston et al. 2009b, a). This highlights the uniform direction of carbon decay processes and indicates that an increase in molecular homogeneity in soils, inland waters and oceans is not necessarily the result of increased mixing of different chemically unique sources. Instead, it is rather an effect of the accumulation of chemically similar decay products and the disappearance of unique molecular formulas as decomposition furthers.
However, while differences between species decreased, the number of molecules found in litter DOM extracts increased with HR litter decay. Thus, while microbial processing of litter leads to increased similarity in leachates between sources, at the same time it results in an interspecific increase in chemo-diversity. The latter might be explained by the presence of a few high RA peaks in fresh HR litter, which could prevent the detection of less abundant compounds that as a result fall under the detection limit. Still, this suggests a less diverse composition, similar to how high abundancies of one species can prevent the detection of rare species and results in lower diversity scores (Chao and Shen 2003). It should be pointed out that while the SPE carbon recovery rate was within bounds of previous research (20–60% (Raeke et al. 2016), it varied between samples and was often higher at more decayed litter than fresh leachates (Suppl. Fig. 1). A lower SPE carbon recovery might indicate that a larger fraction or different types of molecules were not accessible to FT-ICR MS.
The chemistry of labile vs. recalcitrant litter leachates
A higher BDOM was associated with a greater reduction of highly oxidized phenolic compounds in HR litter, while for LR litter it was associated with the reduction of aliphatics (Fig. 7, 8; Table 3). In soils an increase in aliphatics and a decrease in phenolics can be seen in DOM with increasing depth, although the reason for this is thought to be related to sorption mechanics. Phenolic compounds are sorbed to the soil matrix, while aliphatics remain in soluble form (Kaiser et al. 2004; Traversa et al. 2008). Our results suggest that part of the decrease in phenolic compounds seen in soils could also be due to the decomposition of the highly oxidized labile DOM leached from HR litter species. However, in situ detection of these compounds, even in the litter layer of soils, is highly situational due to the rapid decomposition of fresh HR litter leachates (Fig. 2).
A higher BDOM was associated with a greater increase of aliphatics and low oxidized phenolics for HR and LR litter leachates respectively that remained persistent over the incubation period (Fig. 8; Table 3). The chemistry of the DOM after incubation might also reflect microbial products, as overall there were high DOM decay rates (Fig. 2) and large shifts in chemical composition. This is especially true for the fresh and 10-day decomposed HR litter leachates. As such the increase in aliphatics, as related to HR litter BDOM, might be related to the production of secondary microbial compounds, as substantial amounts of aliphatics can be found in bacteria (Kögel-Knabner 2002). In comparison, the increase of low oxidized phenolic compounds in LR litter is more likely accumulation of undecomposed litter leachates as phenolic compounds are associated more with plant than bacterial or fungal tissues (Kögel-Knabner 2002).
Importance for soil biogeochemical processing
The flux of DOM from litter to soils determines to a large extent terrestrial ecosystem C sequestration (Kalbitz et al. 2000). The bioavailability of DOM suggests that HR litter leachates are more easily processed by microbes. On a short timescale this might lead to a faster return of CO2 through higher mineralization of the DOM. However, long term C sequestration is linked to microbial processing of C and as the DOM of HR species has a higher bioavailability this might lead to increased persistent SOM formation through the so called DOM-microbial pathway (Cotrufo et al. 2013, 2015). Microbially processed C has been shown to be a very stable component of SOM (Marschner and Kalbitz 2003; Miltner et al. 2012). In addition, aliphatic components have been shown to accumulate in soils and can persist for millennia (Lorenz et al. 2007). Our results indicate the production of stable aliphatic compounds as fresh HR litter leachates are degraded. In contrast, supplying subsoils with fresh plant derived C can stimulate the mineralization of aged C (Fontaine et al. 2007). However, as DOM moves through the soil it is degraded, transformed and sorbed along the way (Shen et al. 2015), meaning that the labile C of fresh litter leachates is less likely to arrive at subsoil depths unaltered.
Since birch, alder or Vaccinium litter initially produce far more DOM than spruce, pine or wood (i.e. LR) litter, the lability and chemistry of these leachates might have an overall larger effect on soil biogeochemistry than the abundant coniferous litter species. In our boreal research area more than 90% of potentially fresh litter leached DOM can be attributed to birch, alder and Vaccinium, while producing less than 50% of the total litterfall (Hensgens et al. 2020). This difference is bound to seasonality as for these species litter C leaching decreases sharply with increased decomposition, while for coniferous and wood litter C leaching stays more or less equal during the first 180 days. Moreover, previously we showed that approximately 70% of fresh litter leached DOM comes from the understory species V. myrtillus (Hensgens et al. 2020). The chemistry of V. myrtillus is most differentiating from the rest, showing the highest amount of oxygenized phenolic compounds released of all species. As such, the understory species V. myrtillus might have a large effect on soil biogeochemistry where it is abundant and possibly lead to higher soil C sequestration rates.
Conclusion
Our results show that following litterfall, litter decay and leaching dynamics follow different paths depending on litter species and more importantly litter reactivity. The reactive birch, alder and Vaccinium (HR) litter produces quantitatively more leachable and labile DOM. As microbial processing of litter decomposition progress, the degree of similarity in chemistry and bioavailability of the leachates increase. In addition, DOM of different litter species also increase in chemical similarity as it is decomposed. This reiterates the importance of microbial processing, not mixing of different terrestrial DOM sources, in sha** the increasingly homogenous chemistry of DOM found in soils, surface waters and oceans. The high lability of DOM across litter sources in combination with the uni-directional changes in chemistry offer an insight in microbial processing and explains why the distinct chemistry of litter sources have little effect on long-term evolving soil or terrestrially exported DOM chemistry.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
5.1. RDCT (2018) A Language and Environment for Statistical Computing. R Found. Stat. Comput. 2:https://www.R-project.org
Aber JD, Melillo JM, McClaugherty CA (1990) Predicting long-term patterns of mass loss, nitrogen dynamics, and soil organic matter formation from initial fine litter chemistry in temperate forest ecosystems. Can J Bot 68:2201–2208. https://doi.org/10.1139/b90-287
Berg B, Ekbohm G (1991) Litter mass-loss rates and decomposition patterns in some needle and leaf litter types. Long-term decomposition in a Scots pine forest. VII Can J Bot 69:1449–1456. https://doi.org/10.1139/b91-187
Berg B, Hannus K, Popoff T, Theander O (1982) Changes in organic chemical components of needle litter during decomposition. Long-term decomposition in a Scots pine forest: I. (Pinus sylvestris). Can J Bot 60:1310–1319. https://doi.org/10.1139/b82-167
Berg B, Staaf H (1980) Decomposition rate and chemical changes of Scots pine needle litter, II. Influence of chemicel composition. Struct Funct North Conifer For—an Ecosyst study
Bonanomi G, Senatore M, Migliozzi A et al (2015) Decomposition of submerged plant litter in a Mediterranean reservoir: A microcosm study. Aquat Bot 120:169–177. https://doi.org/10.1016/j.aquabot.2014.05.006
Bowen SR, Gregorich EG, Hopkins DW (2009) Biochemical properties and biodegradation of dissolved organic matter from soils. Biol Fertil Soils 45:733–742. https://doi.org/10.1007/s00374-009-0387-6
Bradshaw CJA, Warkentin IG (2015) Global estimates of boreal forest carbon stocks and flux. Glob Planet Change 128:24–30. https://doi.org/10.1016/j.gloplacha.2015.02.004
Brock O, Helmus R, Kalbitz K, Jansen B (2020) Non-target screening of leaf litter-derived dissolved organic matter using liquid chromatography coupled to high-resolution mass spectrometry (LC-QTOF-MS). Eur J Soil Sci 71:420–432. https://doi.org/10.1111/ejss.12894
Campbell EE, Parton WJ, Soong JL et al (2016) Using litter chemistry controls on microbial processes to partition litter carbon fluxes with the Litter Decomposition and Leaching (LIDEL) model. Soil Biol Biochem 100:160–174. https://doi.org/10.1016/j.soilbio.2016.06.007
Cepáková S, Frouz J (2015) Changes in chemical composition of litter during decomposition: a review of published 13C NMR spectra. J Soil Sci Plant Nutr 15:805–815. https://doi.org/10.4067/S0718-95162015005000055
Chao A, Shen TJ (2003) S Index of diversity when there are unseen species in sample. Environ Ecol Stat 10:429–443
Chen HYH, Brant AN, Seedre M et al (2017) The contribution of litterfall to net primary production during secondary succession in the Boreal Forest. Ecosystems 20:830–844. https://doi.org/10.1007/s10021-016-0063-2
Chun H, Keleş S (2010) Sparse partial least squares regression for simultaneous dimension reduction and variable selection. J R Stat Soc Ser B Stat Methodol 72:3–25. https://doi.org/10.1111/j.1467-9868.2009.00723.x
Cleveland CC, Neff JC, Townsend AR, Hood E (2004) Composition, dynamics, and fate of leached dissolved organic matter in terrestrial ecosystems: results from a decomposition experiment. Ecosystems 7:275–285. https://doi.org/10.1007/s10021-003-0236-7
Cornelissen JHC (1996) An experimental comparison of leaf decomposition rates in a wide range of temperate plant species and types. J Ecol 84:573. https://doi.org/10.2307/2261479
Cotrufo MF, Soong JL, Horton AJ et al (2015) Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat Geosci 8:776–779. https://doi.org/10.1038/ngeo2520
Cotrufo MF, Wallenstein MD, Boot CM et al (2013) The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Glob Chang Biol 19:988–995. https://doi.org/10.1111/gcb.12113
Dilly O, Zyakun A (2008) Priming effect and respiratory quotient in a forest soil amended with glucose. Geomicrobiol J 25:425–431. https://doi.org/10.1080/01490450802403099
Don A, Kalbitz K (2005) Amounts and degradability of dissolved organic carbon from foliar litter at different decomposition stages. Soil Biol Biochem 37:2171–2179. https://doi.org/10.1016/j.soilbio.2005.03.019
Fontaine S, Barot S, Barré P et al (2007) Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 450:277–280. https://doi.org/10.1038/nature06275
Gower ST, Krankina O, Olson RJ et al (2001) Net primary production and carbon allocation patterns of boreal forest ecosystems. Ecol Appl 11:1395–1411. https://doi.org/10.1890/1051-0761(2001)011[1395:NPPACA]2.0.CO;2
Grandy AS, Neff JC (2008) Molecular C dynamics downstream: the biochemical decomposition sequence and its impact on soil organic matter structure and function. Sci Total Environ 404:297–307. https://doi.org/10.1016/j.scitotenv.2007.11.013
Hempfling R, Ziegler F, Zech W, Schulten HR (1987) Litter decomposition and humification in acidic forest soils studied by chemical degradation, IR and NMR spectroscopy and pyrolysis field ionization mass spectrometry. Z Pflanzenernähr Bodenkd 150:179–186. https://doi.org/10.1002/jpln.19871500311
Hensgens G, Laudon H, Peichl M et al (2020) The role of the understory in litter DOC and nutrient leaching in boreal forests. Biogeochemistry 149:87–103. https://doi.org/10.1007/s10533-020-00668-5
Herzsprung P, Hertkorn N, Von Tumpling W et al (2014) Understanding molecular formula assignment of Fourier transform ion cyclotron resonance mass spectrometry data of natural organic matter from a chemical point of view. Anal Bioanal Chem 406:7977–7987. https://doi.org/10.1007/s00216-014-8249-y
Kaiser K, Guggenberger G, Haumaier L (2004) Changes in dissolved lignin-derived phenols, neutral sugars, uronic acids, and amino sugars with depth in forested Haplic Arenosols and Rendzic Leptosols. Biogeochemistry 70:135–151. https://doi.org/10.1023/B:BIOG.0000049340.77963.18
Kaiser K, Kalbitz K (2012) Cycling downwards—dissolved organic matter in soils. Soil Biol Biochem 52:29–32. https://doi.org/10.1016/j.soilbio.2012.04.002
Kalbitz K, Schwesig D, Rethemeyer J, Matzner E (2005) Stabilization of dissolved organic matter by sorption to the mineral soil. Soil Biol Biochem 37:1319–1331. https://doi.org/10.1016/j.soilbio.2004.11.028
Kalbitz K, Solinger S, Park J-H et al (2000) Controls on the dynamics of dissolved organic matter in soils: a review. Soil Sci 165:277–304
Kind T, Fiehn O (2007) Seven Golden Rules for heuristic filtering of molecular formulas obtained by accurate mass spectrometry. BMC Bioinf 8:1–20. https://doi.org/10.1186/1471-2105-8-105
Kleber M, Sollins P, Sutton R (2007) A conceptual model of organo-mineral interactions in soils: self-assembly of organic molecular fragments into zonal structures on mineral surfaces. Biogeochemistry 85:9–24. https://doi.org/10.1007/s10533-007-9103-5
Klotzbücher T, Filley TR, Kaiser K, Kalbitz K (2011a) A study of lignin degradation in leaf and needle litter using 13C-labelled tetramethylammonium hydroxide (TMAH) thermochemolysis: Comparison with CuO oxidation and van Soest methods. Org Geochem 42:1271–1278. https://doi.org/10.1016/j.orggeochem.2011.07.007
Klotzbücher T, Kaiser K, Guggenberger G et al (2011b) A new conceptual model for the fate of lignin in decomposing plant litter. Ecology 92:1052–1062. https://doi.org/10.1890/10-1307.1
Koch BP, Dittmar T (2006) From mass to structure: an aromaticity index for high-resolution mass data of natural organic matter. Rapid Commun Mass Spectrom 20:926–932. https://doi.org/10.1002/rcm.2386
Koch BP, Dittmar T (2016) Erratum: From mass to structure: An aromaticity index for high-resolution mass data of natural organic matter (Rapid Communications in Mass Spectrometry (2006) 20 (926–932) DOI: https://doi.org/10.1002/rcm.2386). Rapid Commun Mass Spectrom 30:250. https://doi.org/https://doi.org/10.1002/rcm.7433
Koch BP, Kattner G, Witt M, Passow U (2014) Molecular insights into the microbial formation of marine dissolved organic matter: Recalcitrant or labile? Biogeosciences 11:4173–4190. https://doi.org/10.5194/bg-11-4173-2014
Koehler B, Von Wachenfeldt E, Kothawala D, Tranvik LJ (2012) Reactivity continuum of dissolved organic carbon decomposition in lake water. J Geophys Res Biogeosciences 117:1–14. https://doi.org/10.1029/2011JG001793
Kögel-Knabner I (2002) The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol Biochem 34:139–162. https://doi.org/10.1016/S0038-0717(01)00158-4
Laudon H, Taberman I, Ågren A et al (2013) The Krycklan Catchment study—a flagship infrastructure for hydrology, biogeochemistry, and climate research in the boreal landscape. Water Resour Res 49:7154–7158. https://doi.org/10.1002/wrcr.20520
Lechtenfeld OJ, Hertkorn N, Shen Y et al (2015) Marine sequestration of carbon in bacterial metabolites. Nat Commun 6:1–8. https://doi.org/10.1038/ncomms7711
Lechtenfeld OJ, Kattner G, Flerus R et al (2014) Molecular transformation and degradation of refractory dissolved organic matter in the Atlantic and Southern Ocean. Geochim Cosmochim Acta 126:321–337. https://doi.org/10.1016/j.gca.2013.11.009
Lehmann J, Kleber M (2015) The contentious nature of soil organic matter. Nature 528:60–68. https://doi.org/10.1038/nature16069
Lorenz K, Lal R, Preston CM, Nierop KGJ (2007) Strengthening the soil organic carbon pool by increasing contributions from recalcitrant aliphatic bio(macro)molecules. Geoderma 142:1–10. https://doi.org/10.1016/j.geoderma.2007.07.013
Lorenz K, Preston CM, Krumrei S, Feger KH (2004) Decomposition of needle/leaf litter from Scots pine, black cherry, common oak and European beech at a conurbation forest site. Eur J For Res 123:177–188. https://doi.org/10.1007/s10342-004-0025-7
Lorenz K, Preston CM, Raspe S et al (2000) Litter decomposition and humus characteristics in Canadian and German spruce ecosystems: information from tannin analysis and. Soil Biol Biochem 32:1–14
Marschner B, Kalbitz K (2003) Controls of bioavailability and biodegradability of dissolved organic matter in soils. Geoderma 113:211–235. https://doi.org/10.1016/S0016-7061(02)00362-2
Melillo JM, Aber JD, Linkins AE et al (1989) Carbon and nitrogen dynamics along the decay continuum: plant litter to soil organic matter. Plant Soil 115:189–198. https://doi.org/10.1007/BF02202587
Melillo JM, Aber JD, Muratore JF (1982) Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 63:621–626. https://doi.org/10.2307/1936780
Melillo JM, McGuire AD, Kicklighter DW et al (1993) Global climate change and terrestrial net primary production. Nature 363:234–240. https://doi.org/10.1038/363234a0
Melillo JM, Naiman RJ, Aber JD, Linkins AE (1984) Factors controlling mass loss and nitrogen dynamics of plant litter decaying in northern streams. Bull Mar Sci 35:341–356
Mentges A, Feenders C, Seibt M et al (2017) Functional molecular diversity of marine dissolved organic matter is reduced during degradation. Front Mar Sci 4:1–10. https://doi.org/10.3389/fmars.2017.00194
Miltner A, Bombach P, Schmidt-Brücken B, Kästner M (2012) SOM genesis: microbial biomass as a significant source. Biogeochemistry 111:41–55. https://doi.org/10.1007/s10533-011-9658-z
Minor EC, Swenson MM, Mattson BM, Oyler AR (2014) Structural characterization of dissolved organic matter: a review of current techniques for isolation and analysis. Environ Sci Process Impacts 16:2064–2079. https://doi.org/10.1039/c4em00062e
Mosher JJ, Kaplan LA, Podgorski DC et al (2015) Longitudinal shifts in dissolved organic matter chemogeography and chemodiversity within headwater streams: a river continuum reprise. Biogeochemistry 124:371–385. https://doi.org/10.1007/s10533-015-0103-6
Müller C, Abbasi MK, Kammann C et al (2004) Soil respiratory quotient determined via barometric process separation combined with nitrogen-15 labeling. Soil Sci Soc Am J 68:1610–1615. https://doi.org/10.2136/sssaj2004.1610
Nebbioso A, Piccolo A (2013) Molecular characterization of dissolved organic matter (DOM): a critical review. Anal Bioanal Chem 405:109–124. https://doi.org/10.1007/s00216-012-6363-2
Osono T, Takeda H (2005) Decomposition of organic chemical components in relation to nitrogen dynamics in leaf litter of 14 tree species in a cool temperate forest. Ecol Res 20:41–49. https://doi.org/10.1007/s11284-004-0002-0
Pérez-Harguindeguy N, Díaz S, Cornelissen JHC et al (2000) Chemistry and toughness predict leaf litter decomposition rates over a wide spectrum of functional types and taxa in central Argentina. Plant Soil 218:21–30. https://doi.org/10.1023/A:1014981715532
Prescott CE (2010) Litter decomposition: what controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 101:133–149. https://doi.org/10.1007/s10533-010-9439-0
Preston CM, Nault JR, Trofymow JA (2009a) Chemical changes during 6 years of decomposition of 11 litters in some Canadian forest sites. Part 2. 13C abundance, solid-state 13C NMR spectroscopy and the meaning of “lignin.” Ecosystems 12:1078–1102. https://doi.org/10.1007/s10021-009-9267-z
Preston CM, Nault JR, Trofymow JA, Smyth C (2009b) Chemical changes during 6 years of decomposition of 11 litters in some Canadian forest sites. Part 1. elemental composition, tannins, phenolics, and proximate fractions. Ecosystems 12:1053–1077. https://doi.org/10.1007/s10021-009-9266-0
Preston CM, Trofymow JA (2000) Variability in litter quality and its relationship to litter decay in Canadian forests. Can J Bot 78:1269–1287. https://doi.org/10.1139/cjb-78-10-1269
Raeke J, Lechtenfeld OJ, Wagner M et al (2016) Selectivity of solid phase extraction of freshwater dissolved organic matter and its effect on ultrahigh resolution mass spectra. Environ Sci Process Impacts 18:918–927. https://doi.org/10.1039/c6em00200e
Schmidt MWI, Torn MS, Abiven S et al (2011) Persistence of soil organic matter as an ecosystem property. Nature 478:49–56. https://doi.org/10.1038/nature10386
Shen Y, Chapelle FH, Strom EW, Benner R (2015) Origins and bioavailability of dissolved organic matter in groundwater. Biogeochemistry 122:61–78. https://doi.org/10.1007/s10533-014-0029-4
Silveira ML (2011) Litter decomposition and soluble carbon, nitrogen, and phosphorus release in a forest ecosystem. Open J Soil Sci 01:86–96. https://doi.org/10.4236/ojss.2011.13012
Soares ARA, Kritzberg ES, Custelcean I, Berggren M (2018) Bacterioplankton responses to increased organic carbon and nutrient loading in a boreal estuary—separate and interactive effects on growth and respiration. Microb Ecol 76:144–155. https://doi.org/10.1007/s00248-017-1115-7
Soong JL, Fuchslueger L, Marañon-Jimenez S et al (2020) Microbial carbon limitation: the need for integrating microorganisms into our understanding of ecosystem carbon cycling. Glob Chang Biol 26:1953–1961. https://doi.org/10.1111/gcb.14962
Soong JL, Parton WJ, Calderon F et al (2015) A new conceptual model on the fate and controls of fresh and pyrolized plant litter decomposition. Biogeochemistry 124:27–44. https://doi.org/10.1007/s10533-015-0079-2
Strid A, Lee BS, Lajtha K (2016) Homogenization of detrital leachate in an old-growth coniferous forest, OR: DOC fluorescence signatures in soils undergoing long-term litter manipulations. Plant Soil 408:133–148. https://doi.org/10.1007/s11104-016-2914-1
Thieme L, Graeber D, Hofmann D et al (2019) Dissolved organic matter characteristics of deciduous and coniferous forests with variable management: different at the source, aligned in the soil. Biogeosciences 16:1411–1432. https://doi.org/10.5194/bg-16-1411-2019
Traversa A, D’Orazio V, Senesi N (2008) Properties of dissolved organic matter in forest soils: Influence of different plant covering. For Ecol Manage 256:2018–2028. https://doi.org/10.1016/j.foreco.2008.07.038
Zech W, Johansson MB, Haumaier L, Malcolm RL (1987) CPMAS 13C NMR and IR spectra of spruce and pine litter and of the Klason lignin fraction at different stages of decomposition. Z Pflanzenernähr Bodenkd 150:262–265. https://doi.org/10.1002/jpln.19871500413
Acknowledgements
Open access funding provided by Lund University. This research was supported by core funding from Formas (#239-2014-698) and the Royal Physiography Society Lund. The authors appreciate the support of ProVIS Center for Chemical Microscopy by the Helmholtz Centre for Environmental Research – UFZ and the funds for ProVIS establishment provided by EFRE (Europäischer Fonds für regionale Entwicklung) and the federal state of Saxony (Freistaat Sachsen). The support from Krycklan via SITES is also appreciated.
Funding
This research was supported by core funding from Formas (#239-2014-698), the Royal Physiography Society Lund, Åforsk as well as the funding for the ProVIS Centre for Chemical Microscopy, which was established with the European Regional Development Funds (EFRE—Europe funds Saxony) at the Helmholtz Centre for Environmental Research and was supported by the Helmholtz Association. Additional support from Krycklan via SITES.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Responsible Editor: James Sickman
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hensgens, G., Lechtenfeld, O.J., Guillemette, F. et al. Impacts of litter decay on organic leachate composition and reactivity. Biogeochemistry 154, 99–117 (2021). https://doi.org/10.1007/s10533-021-00799-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-021-00799-3