
Abstract
The influence of rainfall and hydrological variables on the abundance and diversity of the copepod community was investigated on a monthly basis over an annual cycle in the Taperaçu mangrove estuary. In general, the results show that there were no clear spatial or tidal patterns in any biological variables during the study period, which was related to the reduced horizontal gradient in abiotic parameters, determined mainly by the morphological and morphodynamic features of the estuary. Nevertheless, seasonal and monthly trends were recorded in both the hydrological data and the abundance of the dominant copepod species. In particular, Pseudodiaptomus marshi (6,004.6 ± 22,231.6 ind m−3; F = 5.0, p < 0.05) and Acartia tonsa (905.6 ± 2,400.9 ind m−3; F = 14.6, p < 0.001) predominated during the rainy season, whereas Acartia lilljeborgii (750.8 ± 808.3 ind m−3; U = 413.0, p < 0.01) was the most abundant species in the dry season. A distinct process of succession was observed in the relative abundance of these species, driven by the shift in the rainfall regime, which affected hydrological, in particular salinity, and consequently the abundance of copepod species. We suggest that this may be a general pattern governing the dynamics of copepod populations in the estuaries of the Brazilian Amazonian region.
Similar content being viewed by others

Introduction
Brazil has the second largest area of mangrove forest on Earth, distributed along a coast of approximately 6,800 km (Kjerfve et al. 1997). The country’s northernmost coastline is extremely irregular, with innumerable bays, muddy, and sand beaches, salt and freshwater swamps, islands, and estuaries, among other ecosystems (Souza-Filho et al. 2005) that shelter highly complex and productive ecosystems, in which the biological dynamics are influenced by combined effects of the local climate and the physical–chemical characteristics of the water (Giarrizzo and Saint-Paul 2008; Costa et al. 2011). The hydrological and hydrodynamic conditions of these environments are favorable for the development of the planktonic larvae of a wide range of marine fish and shellfish species, which use these habitats as nurseries by providing a rich food source (e.g., phytoplankton and detritus) and protection from predation, as reported to other coastal ecosystems worldwide (Dauvin and Desroy 2005; Nagelkerken et al. 2008).
The estuaries of the Amazonian coast are characterized by a relatively hot and humid equatorial climate which, combined with a semidiurnal macrotidal regime and vast areas of mangrove forest, creates a unique aquatic ecosystem, in terms of its physical–chemical and biological characteristics, especially in comparison with other tropical coastal environments in India (Dalal and Goswami 2001; Madhu et al. 2007), Africa (Champalbert et al. 2007), Australia (Duggan et al. 2008), Costa Rica (Brugnoli-Olivera and Morales-Ramírez 2008), and other regions of Brazil (Sankarankutty et al. 1995; Marcolin et al. 2010). In recent years, research into the region’s zooplankton (Krumme and Liang 2004; Costa et al. 2008) and especially its copepods Magalhães et al. (2006, 2009a, b, 2011; Leite et al. 2010) has increased considerably.
In temperate waters, the seasonal relationship of phyto- and mesozooplankton populations often shows what resembles coupled oscillations which might suggest a classical Lotka–Volterra predator–prey relationship (Kiørboe 1997), i.e., the phytoplankton populations are limited by predation, while the zooplankton (predator) populations are limited by the availability of feeding resources. This top-down versus bottom-up control has been observed in several tropical and subtropical estuaries around the world (Rios-Jara 1998; Ara 2002; Marques et al. 2007). However, ecological studies in the mangroves estuaries of the Amazonian coast (Atique 2009; Magalhães et al. 2011) suggest that the high productivity of these systems can be related to its detritus-based food web, in which the forest represents one of the main sources of carbon for aquatic consumers (Dobson and Frid 2009). Giarrizzo et al. (2011) confirmed the importance of the mangroves in this region as a source of carbon using stable isotopes, showing that this compound enters the food web indirectly as particulate organic matter (POM), which is assimilated by Calanoida copepods and transferred to higher trophic levels through the ingestion of these invertebrates by chaetognaths and small zooplanktivores pelagic fishes. Clearly, the role of the copepods as a direct and efficient path for the transfer of energy to higher level consumers is pivotal to the functioning of this system. Given this, understanding the biological attributes of copepod populations is essential for the reliable assessment of their contribution to the processes that structure the aquatic systems they inhabit, as well as the evaluation of the effects of anthropogenic impacts on these systems, to which these organisms are particularly sensitive (Park and Marshall 2000).
Despite the importance of copepods at the secondary trophic level, the scale and pattern of temporal variations in abundance and biodiversity are still poorly known in the specific context of tropical estuaries. Estimates of copepods abundance are fundamental to the determination of biomass and productivity, which are crucial for the understanding of energy flow and the trophodynamics of estuarine systems. In addition to all these considerations, the assessment of biodiversity on both a regional and global scale is an increasingly important issue in ecology, given the growing evidence of profound anthropogenic modifications to the biosphere (Palumbi 2001).
Given these considerations, the present study focused on a continuous 12-month analysis of the abundance and ecological indices (species diversity, evenness, and richness) of copepods in a tropical, tide-dominated estuary, the Taperaçu, in northern Brazil. These biological parameters provide an annual profile of the zooplankton community structure in an estuary with unique characteristics, such as the absence of fluvial discharge, a small catchment area, shallow waters, and strong tidal currents, which may be especially useful for comparative studies with similar systems in Brazil and in other parts of the world. A primary aim of this survey was to contribute to the understanding of the effects of regional coastal conditions (e.g., climatological and hydrological) on the temporal variation of copepod populations in tropical estuarine ecosystems.
Materials and methods
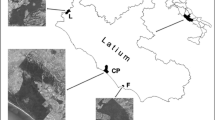
This study was carried out in the Taperaçu estuary (46°42′–46°45′W and 00°50′–00°57′S), located about 200 km southeast of the mouth of the Amazon River, in the municipality of Bragança, northern Brazil. It is classified as a shallow (mean depth = 4.2 m), permanently open estuary, with many sandbanks running down the middle, high turbidity (mean 378.1), and strong tidal currents of up to 2.04 m s−1 (Asp et al. 2012). Delimited by the Atlantic Ocean and the Maiaú Bay and Caeté estuary (Fig. 1), the Taperaçu has 21 km2 of surface water and a catchment area of approximately 40 km2, but no effective source of freshwater. A recent study conducted in this estuary indicates that the system is characterized by discreet vertical gradients in hydrological parameters (Asp et al. 2012). The main hydrodynamic feature is a semidiurnal tidal range of about 5 m, which may reach 6 m during the equinoctial spring tides (DHN 2008).

Study area: a South America; b Location of the Taperaçu estuary on the Amazonian coast of northern Brazil; c Positions of the sampling stations in the upper (1), middle (2), and lower (3) sectors of the Taperaçu estuary, with the black arrow indicating the position of Taici creek, which connects the Taperaçu and Caeté estuaries (modified from Mehlig 2001)
The local climate is humid equatorial, with a rainy season normally between January and July, and a dry (or less rainy) season between August and December (Moraes et al. 2005), although considerable variation may be observed in some years. This seasonal pattern is controlled primarily by shifts in the intertropical convergence zone (ITCZ), which provokes intense convective activity during the first half of the year. Mean annual rainfall is 2,401 mm (INMET 2012), with 75 % of the precipitation falling during the rainy season. Average monthly air temperatures are between 25.2 and 26.7 °C, with maximum daily amplitude between 20.4 and 32.8 °C (Schories and Mehlig 2000).
The Taperaçu estuary is surrounded by wetlands and extensive mangrove forests, with trees reaching up to 20 m in height, and a complex network of tidal creeks, which link the Taperaçu with the neighboring Maiaú Bay and Caeté estuary. Due to its biogeochemical, ecological, and socioeconomic importance, this ecosystem has been designated a conservation unit (RESEX Caeté-Taperaçu) by the Brazilian government.
Zooplankton samples were collected at monthly intervals in the Taperaçu estuary from January to December 2006 during spring tides at three stations located in the upper (1), middle (2), and lower (3) sectors of the estuary (Fig. 1). Each sample consisted of a 3-min subsurface horizontal tow of a conical plankton net (300 μm mesh) fitted with a mechanical flowmeter to estimate the volume of water filtered through the net. Two tows per station were conducted each month—one on the ebb tide and the other on the flood tide—resulting in a total of 72 samples. The tows were performed during the diurnal period, using a small powerboat running at an average speed of 1.5 knot. As soon as they were collected, the samples were preserved in a 4 % buffered formaldehyde seawater solution. Physical and chemical parameters of the water (salinity, temperature, pH, and dissolved oxygen (DO) concentrations) were measured in situ using a multiparameter analyzer. The salinity was measured using the Practical Salinity Scale. The transparency of the water was determined using a Secchi disk. Rainfall data for the sampling periods were obtained from the Tracuateua station of the National Meteorological Institute, which is located at 01°05′S, 47°10′W, about 20 km from the study area.
In the laboratory, the preserved zooplankton samples were rinsed to remove the formalin buffer and divided into aliquots between one and nine times in a Folsom splitter to provide standardized subsamples containing ~500 copepods, which were then identified (Björnberg 1981; Bradford-Grieve et al. 1999), classified taxonomically (WoRMS 2012), and counted in a gridded Petri dish under a stereomicroscope. The counts for each species were multiplied by the subsampling factor (1–9) to obtain the estimate of the total number of individuals in the full sample. The adult copepods were identified to species or the lowest possible taxonomic level. Copepodites were identified to the genus level. Adults and juveniles that could not be identified to the genus or species level were classified as unidentified copepod, together with copepod nauplii.
To analyze the monthly and seasonal patterns of variation in the biological attributes of the local copepod populations, the quantitative data obtained for each sample were used to calculate abundance and ecological indices (diversity, evenness, and species richness). Abundance was expressed as the number of individuals per m3 (ind m−3). Copepod diversity (H ′) and evenness (J ′) were calculated, respectively according to:
-
Shannon–Wiener index \(\left( {H^{{\prime }} } \right) = - \sum\limits_{i = 1}^{s} {\left( {pi\log_{2} pi} \right)}\), Shannon (1948);
-
Pielou index \(\left( {J^{{\prime }} } \right) = H^{{\prime }} /\log_{2} S\), Pielou (1977);
where pi = ni/N (proportion of the sample belonging to the ith species). S = the total number of species recorded in the sample (species richness) and N = the total number of individuals of all the species in a sample. The relative frequency of occurrence (FO) of a given species was determined by the proportion of samples in which the species was recorded.
The one-way analysis of variance (ANOVA) and the nonparametric Mann–Whitney U test were applied to compare variation in abiotic parameters, the abundance of the two principal groups of copepod taxa (total copepod fauna and the dominant species), and their ecological indices between sampling stations (S1, S2, and S3), tides (flood/ebb), months (January–December), and seasons (rainy/dry). When significant F values were recorded in the ANOVA, Tukey’s multiple comparisons test was used to discern significant differences between treatments (Zar 2010). All analyses were conducted in STATISTICA 5, with α = 0.05. In the present study, species contributing more than 10 % of the total number of copepods in a given month were classified as dominant.
A multivariate redundancy analysis (RDA) was carried out to determine the relationships between environmental variables and the abundance of copepod taxa, using the CANOCO 4.5 software package (ter Braak and Smilauer 1998). The results are presented as a biplot, in which species and environmental variables are plotted together. To assess the similarity among months, a hierarchical agglomerative cluster analysis and non-metric multidimensional scaling (NMDS) ordination were computed using the Bray–Curtis similarity index for log-transformed (x + 1) abundance data. A one-way analysis of similarity (ANOSIM) was subsequently used to test whether copepod community varied significantly between seasons. In addition, a similarity percentage (SIMPER) analysis was used to identify the species that contributed most to the similarities within and dissimilarities between the groups formed by the cluster and NMDS analyses. These analyses were run in the PRIMER 6 software package, following Clarke and Warwick (1994).
Results
Mean monthly rainfall in the dry season (42.7 ± 33.9 mm) was considerably lower than that recorded in the rainy season (313.9 ± 151.6 mm), with 88 % of the year’s total precipitation (1,883.3 mm) falling between February and July, which was thus defined as the rainy season (Fig. 2). During the rainy season months, significant reductions were recorded in salinity (U = 1.0, p < 0.01), pH (F = 30.6, p < 0.01), and water temperature (U = 232.5, p < 0.01), which appeared to be linked to the massive inflow of freshwater from rainfall runoff occurring during this period (Fig. 2). Mesohaline–marine regimes were predominant throughout the study period, with mean salinity ranging from 11.0 ± 0.6 in April to 38.7 ± 0.8 in November. The pH values were generally alkaline, ranging from 7.2 ± 0.1 in April to 7.8 ± 0.1 in October. While seasonal differences in mean water temperatures were highly significant (p < 0.01), temperatures were relatively constant throughout the year—which is typical of tropical environments—with an amplitude of only 2.3 °C.

Total monthly rainfall and Mean (±SD) variation in hydrological variables in the Taperaçu estuary, northern Brazil. The shaded portion of the graph indicates the rainy season (February–July)
Dissolved oxygen (DO) concentrations also varied little between seasons (p > 0.05), with water being slightly less oxygenated during the rainy season. The mean DO ranged from 3.0 ± 1.2 mg L−1 in July to 6.1 ± 1.3 mg L−1 in December. A similar trend was observed in the transparency of the water, with mean values varying from 13.5 ± 4.5 cm in October to 21.0 ± 7.3 cm in December. The reduced transparency recorded throughout the year was due to the high concentrations of suspended particulate material and nebulosity typical of this type of environment (Fig. 2).
Overall, no significant spatial patterns were identified in the biological data collected in the Taperaçu estuary, which allowed for the pooling of the data from the three sectors (mean ± standard deviation) for the analysis of monthly and seasonal patterns. In general, short-term variation, related to tidal cycles, was negligible.
The seasonal variation in the copepod populations tended to reflect that of the total abundance of zooplankton during the study period. The Copepoda was the dominant taxon, accounting for up to 57.9 % of total zooplankton abundance in the dry season and 90.6 % in the rainy season. The lowest copepod abundance was recorded in June (4.3 %) and January (28.1 %), when the abundance of brachyuran crab larvae increased considerably. A total of 22 copepods species, belonging to four orders (Calanoida, Cyclopoida, Harpacticoida, and Poecilostomatoida), were identified (Table 1). The Calanoida was the most abundant order and the richest in taxa, with seven families, nine genera, and 15 species, of which Pseudodiaptomus marshi Wright, 1936, Acartia tonsa Dana, 1849, Acartia lilljeborgii Giesbrecht, 1889, and Subeucalanus pileatus (Giesbrecht, 1888) were numerically dominant, contributing up to 99.6 % of total copepod abundance (in February).
While no clear seasonal pattern of abundance (p > 0.05) was recorded for the copepod community as a whole, marked seasonal variation was observed in some individual species (Fig. 3; Table 1). In particular, P. marshi presented marked seasonal fluctuations in abundance (F = 5.0, p < 0.05), increasing significantly from a mean of 1,418.7 ± 5,621.9 ind m−3 in the dry season to 6,004.6 ± 22,231.6 ind m−3 in the rainy season. This species accounted for 90.5 % of the total copepod abundance in February (20,909.69 ± 50,527.07 ind m−3) (Fig. 3). Spikes in the abundance of this species were also recorded in April, when it accounted for 63.8 % of total copepod abundance (9,260.0 ± 21,482.7 ind m−3) and August, when it contributed 75.9 % (7,644.6 ± 12,769.4 ind m−3). A similar seasonal trend was recorded in A. tonsa, with a mean abundance of 119.3 ± 246.9 ind m−3 in the dry season and 905.6 ± 2,400.9 ind m−3 in the rainy season (F = 14.6, p < 0.001). In this case, there was only a single peak in abundance, in April (4,604.182 ± 4,521.736 ind m−3; F = 5.9, p < 0.001), representing 31.7 % of total copepod abundance (Fig. 3).

Total monthly rainfall (histogram) and Mean (±SD) variation in the salinity and abundance (filled circle) of the principal copepod species in the Taperaçu estuary, northern Brazil. The results of the statistical tests refer to the differences between seasons
By contrast, the abundance of A. lilljeborgii was significantly higher (U = 413.0, p < 0.01) in the dry season (750.8 ± 808.3 ind m−3) in comparison with the rainy season (709.5 ± 1,286.3 ind m−3), although a significant peak was observed in the late rainy season month of July (2,227.9 ± 2,082.0 ind m−3; F = 4.5, p < 0.05), when it contributed 62.2 % of total copepod abundance (Fig. 3). Peaks in the abundance of this species were also recorded in February (1,646.4 ± 1,237.3 ind m−3; 7.1 %), August (1,445.5 ± 1,222.7 ind m−3; 14.4 %), September (1,208.6 ± 1,117.4 ind m−3; 62.8 %), and January (961.4 ± 307.8 ind m−3; 69.6 %). Of the four predominant species, S. pileatus was the least representative and was absent from the samples collected between May and July, when mesohaline–polyhaline conditions prevailed. While the mean abundance of S. pileatus was also higher in the dry season (24.3 ± 31.0 ind m−3), there was no significant seasonal variation in its abundance. Classified as a non-dominant species (<10 % of the total copepod abundance in a given month), Paracalanus quasimodo Bowman, 1971, had a high relative frequency of occurrence (61.1 %), with mean abundance varying significantly from 17.5 ± 64.1 ind m−3 in the dry season to 86.8 ± 219.5 ind m−3 in the rainy season (U = 434.0, p < 0.05; Table 1).
No evidence was found of any significant variation in the abundance of A. tonsa and A. lilljeborgii related to the tidal cycle or estuarine sector. This lack of any tidal or spatial pattern was also observed in S. pileatus, although the abundance of this species increased slightly toward the lower estuary during the ebb and flood tides of the rainy season (Fig. 4). However, significant spatial and tidal differences were recorded in P. marshi, which was most abundant at stations 1 (34,322.3 ± 47,972.6 ind m−3) and 2 (1,595.8 ± 1,114.9 ind m−3) during the flood tide in the rainy season, decreasing considerably toward the mouth of the estuary (31.2 ± 46.1 ind m−3), and during the ebb tide (Fig. 4; Table 2). A similar pattern was observed in this species during the dry season, although the different was not statistically significant.

Variation in the Mean (±SD) abundance of the four dominant copepod species in the Taperaçu estuary, northern Brazil, by season (rainy/dry), station (S1, S2, and S3), and tide (ebb/flood)
Copepod diversity (0.5 ± 0.2 to 1.2 ± 0.3 bits ind−1), evenness (0.2 ± 0.0 to 0.5 ± 0.3), and richness (4.2 ± 2.3 to 7.8 ± 2.3) were all relatively low overall (Fig. 5) and varied little (p > 0.05) between stations, tide cycles, months, or between seasons, indicating relative spatial and temporal homogeneity in comparison with other tropical and temperate estuaries worldwide (e.g., Islam et al. 2006; Rakhesh et al. 2013). The low species diversity recorded in the Taperaçu estuary was partly related to the large mesh size of the plankton net (300 μm) used for the collection of samples, which almost certainly underestimated species richness. This was also reflected in the overwhelming abundance of large species (e.g., P. marshi and A. lilljeborgii) in comparison with the smaller ones, such as Oithona oswaldocruzi Oliveira, 1945, and Euterpina acutifrons Dana, 1847 (Table 1).

Mean (±SD) ecological indices recorded over an annual cycle in the Taperaçu estuary, northern Brazil
The relationship between copepod abundance and environmental parameters is shown in the RDA ordination diagram (Fig. 6). Of the four predominant species, A. lilljeborgii and S. pileatus were positively correlated with salinity, which accounts for the higher values recorded in the dry season, when salinity was significantly higher. The opposite pattern was observed in P. marshi, A. tonsa, and P. quasimodo, which were influenced negatively by this parameter.

RDA analysis showing biplot of copepod taxa (large arrows heads) and environmental variables (small arrow heads). Environmental variables are as follows: Sal salinity, pH pH, DO dissolved oxygen, and Temp temperature. Copepods taxa are as follows: Al = Acartia lilljeborgii, At = Acartia tonsa, Pm = Pseudodiaptomus marshi, Pr = Pseudodiaptomus richardi, Sp = Subeucalanus pileatus, Pa = Pseudodiaptomus acutus, Psp = Pseudodiaptomus sp., Ca = copepodite of Acartia, Ea = Euterpina acutifrons, Ha = Harpacticoida, Pq = Paracalanus quasimodo, Oh = Oithona hebes, Oo = Oithona oswaldocruzi, Lf = Labidocera fluviatilis, Cv = Centropages velificatus, Pc = Parvocalanus crassirostris
The hierarchical cluster analysis illustrated clear temporal changes in the copepod community of the Taperaçu estuary (Fig. 7a). In particular, two distinct groups of months were delineated with 60 % similarity. Group I was characterized by the predominance of P. marshi and A. tonsa and included primarily rainy season months. In group II, by contrast, which contained mainly dry season months, there was a decline in the contribution of P. marshi and A. tonsa to total copepod abundance, and an increase in that of A. lilljeborgii (Fig. 7c). The clustering pattern was further confirmed by a NMDS two-dimensional plot (stress: 0.03) with no overlap between groups (Fig. 7b). There was 70.8 % similarity within the rainy season group (group I) and 73.2 % in the dry season group (group II). The results of the NMDS ordination analysis were supported by the ANOSIM (global R = 0.9, p < 0.01). Overall, P. marshi, A. lilljeborgii, A. tonsa, and S. pileatus accounted for over 85 % of the similarity within these groups and for 74 % of the dissimilarity between them. The SIMPER also indicated that P. marshi (62.1 %) and A. lilljeborgii (57.8 %) were the species that most contributed to the similarity within groups I and II, respectively, highlighting the importance of these species in the structural dynamics of the copepod community.

a Cluster and b NMDS analyses of the abundance of copepod species recorded during the study period in the Taperaçu estuary, northern Brazil. c Relative contributions of the dominant species to total copepod abundance. Dominance of oligomesohaline species (group I); dominance of marine species (group II). The main groups shown in the Cluster and NMDS (stress: 0.03) are separated by dashed lines at a similarity level of 60 %
Discussion
The horizontal gradients in hydrological parameters such as salinity, temperature, and turbidity constitute a crucial determinant of the distribution of copepods within estuarine environments (Lawrence et al. 2004; Chew and Chong 2011). The virtual absence of such gradients in the Taperaçu estuary is probably related to a number of morphological and morphodynamic factors, including the effective absence of fluvial discharge, the small catchment area (minimal, sporadic freshwater inflow), shallow depths, and strong tidal currents, which facilitate both horizontal and vertical mixing of the water column. Overall, the lack of any significant spatial variation in biological parameters presumably reflects the absence of any horizontal gradient in hydrological variables, especially salinity.
The Amazonian coast is characterized by pronounced annual fluctuations in rainfall, which, together with the associated variation in fluvial discharge, cause drastic seasonal changes in hydrological parameters. These fluctuations are related with the displacement of the ITCZ, which shifts to the southern hemisphere during the first half of the year, provoking an increase in precipitation levels, whereas in the second half of the year, it shifts back to the northern hemisphere, creating drier conditions (Marengo 1995). In the rainy season, the significant reduction in salinity, pH, and temperature were closely related to the increased in precipitation levels, which leads to a major increment in the discharge of local rivers (e.g., Caeté, Quatipuru), with a knock-on effect in the hydrological parameters, and consequently in the abundance of copepods. During the course of the dry season (<13 % of total annual precipitation), these abiotic variables increased progressively. By contrast, water transparency was relatively low overall due to the large amount of particulate material suspended in the water column, a typical characteristic of shallow Amazonian estuaries. In addition, the Taperaçu was different from other estuarine systems in northern (Guimarães et al. 2009) and northeastern Brazil (Cavalcanti et al. 2008), in the lack of any clear seasonal variation in dissolved oxygen concentrations.
Copepods were by far the most prominent zooplankton group, as found in other mangrove systems in different parts of the world, where they typically account for more than 80 % of total abundance (Rios-Jara 1998; Dias and Bonecker 2008). The copepod community in the Taperaçu estuary was characterized by the predominance of Pseudodiaptomus marshi, Acartia tonsa, Acartia lilljeborgii, and Subeucalanus pileatus. A similar composition has been recorded at the same site in previous studies (Costa et al. 2008; Magalhães et al. 2009a) and in other mangrove estuaries on the Brazilian coast (Almeida Prado-Por and Lansac-Tôha 1984; Sterza and Fernandes 2006). The contribution of other zooplankton groups, e.g., foraminifers, cnidarians, mollusks, amphipods, chaetognaths, brachyurans, appendicularians, and chordates, was highly variable, but together accounted for an average of 38.5 % of total zooplankton abundance. The contribution of the non-copepod zooplankton was greatest in January and June, due to the abundance of the larvae of the brachyuran crabs, which spawns during these months in this region (Diele 2000; Koch et al. 2005).
The variability in the abundance, biomass, and productivity of copepods in shallow tidal estuaries has been associated with the combined effects of regional climate patterns and local hydrological factors (Leandro et al. 2007; Muxagata et al. 2012), in addition to other multifactorial processes involving both biotic and abiotic parameters (Dalal and Goswami 2001; Azeiteiro et al. 2005). In the present study, the seasonal variation in the abundance of copepods appeared to be influenced primarily by the interaction of precipitation levels and salinity. In Taperaçu estuary, this seasonality was defined mainly by shifts in the relative contribution of the four predominant species (P. marshi, A. tonsa, A. lilljeborgii, and S. pileatus). Overall, P. marshi was the dominant taxon, especially in the rainy season months, when salinity decreased. During the dry season, by contrast, P. marshi declined sharply in abundance and was replaced by A. lilljeborgii. While P. marshi may be able to survive and even reproduce in hypersaline water (Medeiros et al. 2006), the results of redundancy analysis suggest a preference for the low to intermediate saline conditions of estuarine environments. P. marshi was more abundant in the Taperaçu estuary than in the neighboring Caeté estuary (Magalhães et al. 2010). These two estuaries are connected by the Taici creek (Fig. 1), through which water flows from the Caeté to the upper Taperaçu during the flood tide (Araújo and Asp 2013). This may have contributed to the recruitment of copepods, such as P. marshi and Pseudodiaptomus richardi (Dahl F., 1894), from the oligomesohaline Caeté estuary (Magalhães unpub. data). Considering the morphological, morphodynamic, and hydrological characteristics of the Taperaçu estuary, this process would account for the variation in the abundance of P. marshi during the rainy season (Fig. 4, Table 2), as well as the presence of organisms more typical of the inner Caeté estuary when marine conditions—as defined by the Venice classification system (Anonymous 1959)—prevailed (i.e., during the dry season months). These results suggest that P. marshi, together with other oligohaline copepods, does not establish resident populations throughout the year, or at least during the dry season. The presence of this species during the dry season months, when the system is clearly more marine in nature, was probably the result of immigration through the Taici creek. This would explain the peak in abundance of P. marshi recorded in August, which contradicts the decrease in precipitation and the increase in salinity recorded during this month.
In estuarine environments, two or more species of Acartia may coexist, with seasonal alternations in dominance (Lee and McAlice 1979; Sullivan and McManus 1986). In the case of the Taperaçu estuary, the increasing freshwater runoff in April, May, and June resulted in a sharp decline in salinity and consequently the progressive substitution of A. lilljeborgii by A. tonsa, due presumably to the relative tolerance of A. tonsa to lower salinity levels.
The planktonic copepod A. tonsa is a cosmopolitan species which occurs in the Atlantic, Indian, and Pacific oceans, and the Azov, Baltic, Black, and Mediterranean Seas, as well as the Caspian Sea, where it is a recent invader. The species is nevertheless distributed preferentially in estuarine and coastal areas (Paffenhöfer and Stearns 1988; Tester and Turner 1991) and is one of the most prominent in the estuaries of the Atlantic coast of South America, being present year-round, but exhibiting major seasonal fluctuations in population density (Gómez-Erache et al. 2000; Costa et al. 2009). Despite being considered a euryhaline and eurythermal species, capable of tolerating salinity levels ranging from 0 to 31.5 (Montú and Gloeden 1986), optimal salinity for A. tonsa is considered to be between 15 and 22 (Cervetto et al. 1999); although in the present study, the greatest abundance was recorded at a salinity of 11.
A number of field and laboratory studies have reported enhanced growth rates in A. tonsa at lower salinity levels (Peck and Holste 2006; Putland and Iverson 2007). The significant negative effects of salinity on the population dynamics of A. tonsa underpin its temporal distribution in the Taperaçu estuary, because the organism’s ability to osmoregulate affects its ecological tolerance. In the present study, the species presented a single annual peak of abundance in the rainy season month of April. This pattern differed from that observed in other Amazonian coastal systems (Leite et al. 2009; Magalhães et al. 2009b), where the species was abundant and predominant throughout much of the rainy season, when salinity was between seven and 19. The reduction in the abundance of A. tonsa during the dry season was determined by its ecological affinity with environments characterized by moderate salinity levels and suggests resting (diapause) eggs production, which is used as a survival strategy in response to seasonal and longitudinal fluctuations in conditions, as observed previously in temperate coastal and marine habitats (Castro-Longoria 2001; Katajisto 2006), where the eggs were maintaining in a resting state by environmental cues such as temperature and salinity. The benthic diapause phase could be one of the bases of the seasonal replacement of Acartia species at the present study site, in addition to the resumption of growth in the remaining adult population under favorable conditions.
Copepods of the genus Acartia inhabit many coastal and offshore environments, where they are usually among the most abundant zooplankton taxa (Greenwood 1981; Hubareva et al. 2008). This was also confirmed in the present study, given the predominance of A. lilljeborgii during most of the second half of the year. This marine–estuarine species is an indicator of coastal waters (Björnberg 1981) and is dominant in many Brazilian estuaries (Schwamborn et al. 2004; Dias et al. 2009). In the Taperaçu, the temporal fluctuations in the abundance of A. lilljeborgii can be explained by its preference for a given range of salinity. In the present study, values peaked at between 25 and 35, which may be the optimal range for egg production in this species. Ara (2001) recorded an increase in the production of eggs in A. lilljeborgii with increasing salinity and found that during periods of reduced abundance related to low salinity (<17), populations of this species were unable to recover rapidly through egg production. Temporal fluctuations in the abundance of this species are thus derived not only from its reproductive output, but also from the modification of this process by extrinsic biological factors and physical conditions, such as salinity. The affinity of this species for highly saline environments has been reported from a number of other tropical estuaries (Brugnoli-Olivera et al. 2004; Dias and Bonecker 2008), as a result of its greater tolerance to high salinities.
As mentioned above, the peak in the abundance of S. pileatus, a species typical of the warm coastal waters of the tropical and subtropical regions of the southwest Atlantic (Björnberg 1981), occurred primarily during the dry season, when marine conditions predominated, suggesting that the presence of this species in the studied area depends on recruitment from the adjacent coastal zone. This could account for the slight increase in its abundance toward the mouth of the estuary during the ebb and flood periods in the rainy season (Fig. 4). Dominant in most Brazilian coastal areas (Magalhães et al. 2009b; Eskinazi-Sant’Anna and Björnberg 2006), P. quasimodo is a marine–euryhaline species, occurring preferentially in waters of high salinity in tide-dominated areas (Lopes et al. 1998; Araújo et al. 2008). However, the opposite tendency was observed in the Taperaçu estuary, where the highest density of this species was recorded during the period of lowest salinity. The euryhaline character of P. quasimodo associated with the strong tidal currents may account for this pattern, as well as the high frequency of occurrence of the species (61.1 %) during the study period.
The lack of any significant variation in ecological indices in relation to estuary sector, tide cycle or climatic variables may be related to the unique configuration of the Taperaçu estuary, and the absence of any major and continuous inflow of freshwater, thus minimizing the horizontal gradients in these indices. Magalhães et al. (2011) also recorded a lack of circadian and tidal level variations in copepod diversity within the Taperaçu estuary, which was attributed to the shallow depths of the estuary combined with its strong tidal currents, which make the water column relatively homogeneous, and eliminate the effects of the vertical migration of the zooplankton on diversity patterns. This process may impede the mechanisms that retain resident copepods in the estuary (e.g., DVM and TVM—diel- and tide-induced vertical migrations), which would account for the reduced spatial–temporal variation recorded in the ecological indices during the course of the study period. The ecological indices recorded in the present study reflect the relatively low diversity typical of estuarine environments (Hopcroft et al. 1998). In tropical areas, however, this may be at least partly due to the presence of small-bodied species, which may be under sampled in nets with mesh sizes over 64 μm (Chisholm and Roff 1990). In this case, it would be necessary to collect further samples using nets of finer mesh in order to evaluate the full diversity of copepods in the study area. A negative effect of mesh size on zooplankton diversity has been recorded in other aquatic ecosystems (Favareto et al. 2009; Riccardi 2010).
Monthly and seasonal changes in the relative contributions of the four principal copepod species, in particular P. marshi and A. lilljeborgii, did not have a direct affect on the ecological indices, which were spatially and temporally homogeneous. However, the present study found a clear seasonal pattern of abundance in the copepod community, characterized by the ecological succession of the main species. The multivariate analyses confirmed the dominance of oligomesohaline species during the rainy season, in contrast with the predominance of estuarine–marine species during the dry season. These fluctuations may play a fundamental role in community structure, especially where salinity exceeds the levels tolerated by the different species and interferes with their osmoregulatory processes, leading to alterations in their reproductive potential and ultimately a decrease in the size of the population. In the present study, the absence of freshwater organisms was related to the lack of an effective river discharge.
The temporal variability of copepods in coastal waters is controlled mainly by salinity and temperature (Rodriguez et al. 1995; Mouny and Dauvin 2002), although bottom-up as well as top-down controls (i.e., food availability and predation pressure) also play an important role in this process (Hoffmeyer and Torres 2001; Islam et al. 2006). In the Taperaçu estuary, some of the dominant copepod species, such as A. lilljeborgii and A. tonsa, are classified as omnivores (Uye et al. 1987; Paffenhöfer 1991; Ara 2001) and present an ample range of feeding behavior, which likely reduces interspecific competition for resources, and may represent a secondary factor in the ecological succession of the different species. This is further supported by the relative abundance and diversity of autotrophic food sources (e.g., benthic and planktonic microalgae, mangrove macroalgae, and detritus) available in the local aquatic food web (Koch and Wolff 2002; Brenner and Krumme 2007; Matos et al. 2011), which may also contribute to a reduction in interspecific competition among the copepods. While predation may also have been an important mechanism of population control, data are only available for P. marshi and A. lilljeborgii in the neighboring Caeté estuary (Camargo and Isaac 2004). In this estuary, these two copepods were the main food items ingested by juvenile fishes of three species, Macrodon ancylodon (Bloch and Schneider, 1801), Stellifer rastrifer (Jordan, 1889), and Stellifer naso (Jordan, 1889). This scarcity of data on the role of predation as a control mechanism for copepod populations in Amazonian estuaries impedes the conclusive evaluation of the contribution of this factor to the structure of the zooplankton community of the Taperaçu estuary as a whole.
References
Almeida Prado Por MS, Lansac Tôha FA (1984) Distribution of brackish water Calanoida (Copepoda) along the coasts of Brazil. Hydrobiologia 113:147–150
Anonymous (1959) The Venice system for the classification of marine waters according to salinity. Arch Oceanogr Limnol 11:243–248
Ara K (2001) Daily egg production rate of the planktonic calanoid copepod Acartia lilljeborgi Giesbrecht in the Cananéia Lagoon estuarine system, São Paulo, Brazil. Hydrobiologia 445:205–215
Ara K (2002) Temporal variability and production of Temora turbinata (Copepoda: Calanoida) in the Cananéia Lagoon estuarine system, São Paulo, Brazil. Sci Mar 66:399–406
Araújo WP Jr, Asp NE (2013) Hydrodynamic connectivity between two macrotidal Amazonian estuaries. J Coast Res Spec Issue 65:1086–1091
Araújo HMP, Nascimento-Vieira DA, Neumann-Leitão S, Schwamborn R, Lucas APO, Alves JPH (2008) Zooplankton community dynamics in relation to the seasonal cycle and nutrient inputs in an urban tropical estuary in Brazil. Braz J Biol 68:751–762
Asp NE, Schettini CAF, Siegle E, Silva MS, Brito RNR (2012) The dynamics of a frictionally-dominated Amazonian estuary. Braz J Oceanogr 60:391–403
Atique P (2009) Microzooplâncton do estuário do rio Caeté (região amazônica, Bragança, Pará). MSc thesis, Universidade Federal do Pará, Brazil
Azeiteiro UM, Marques SC, Vieira LMR, Pastorinho MRD, Ré PAB, Pereira MJ, Morgado FMR (2005) Dynamics of the Acartia genus (Calanoida: Copepoda) in a temperate shallow estuary (the Mondego estuary) on the western coast of Portugal. Acta Adriat 46:7–20
Björnberg TKS (1981) Copepoda. In: Boltovskoy D (ed) Atlas del zooplancton del Atlántico Sudoccidental y métodos de trabajo con el zooplancton marino. Instituto Nacional de Investigación y Desarrollo Pesquero, Mar del Plata, pp 587–679
Bradford-Grieve JM, Markhaseva EL, Rocha CEF, Abiahy B (1999) Copepoda (volume 2). In: Boltovskoy D (ed) South Atlantic zooplankton. Backhuys Publishers, Leiden, pp 869–1098
Brenner M, Krumme U (2007) Tidal migration and patterns in feeding of the four-eyed fish Anableps anableps L. in a north Brazilian mangrove. J Fish Biol 70:406–427
Brugnoli-Olivera E, Morales-Ramírez A (2008) Trophic planktonic dynamics in a tropical estuary, Gulf of Nicoya, Pacific coast of Costa Rica during El Niño 1997 event. Rev Biol Mar Oceanogr 43:75–89
Brugnoli-Olivera E, Díaz-Ferguson E, Delfino-Machin M, Morales-Ramírez A, Arosemena AD (2004) Composition of the zooplankton community, with emphasis in copepods, in Punta Morales, Golfo de Nicoya, Costa Rica. Rev Biol Trop 52:897–902
Camargo M, Isaac V (2004) Food categories reconstruction and feeding consumption estimates for the Sciaenid Macrodon ancylodon (Bloch & Schneider), and the congeneric fishes Stellifer rastrifer (Jordan) and Stellifer naso (Jordan) (Pisces, Perciformes) in the Caeté Estuary, Northern Coast of Brazil. Rev Bras Zool 21:85–89
Castro-Longoria E (2001) Comparative observations on the external morphology of subitaneous and diapause eggs of Acartia species from Southampton water. Crustaceana 74:225–236
Cavalcanti EAH, Neumann-Leitão S, Vieira DAN (2008) Mesozooplankton of the estuarine system of Barra das Jangadas, Pernambuco, Brazil. Rev Bras Zool 25:436–444
Cervetto G, Gaudy R, Pagano M (1999) Influence of salinity on the distribution of Acartia tonsa (Copepoda, Calanoida). J Exp Mar Biol Ecol 239:33–45
Champalbert G, Pagano M, Sene P, Corbin D (2007) Relationships between meso and macro-zooplankton communities and hydrology in the Senegal River Estuary. Estuar Coast Shelf Sci 74:381–394
Chew L-L, Chong VC (2011) Copepod community structure and abundance in a tropical mangrove estuary, with comparisons to coastal waters. Hydrobiologia 666:127–143
Chisholm LA, Roff JC (1990) Abundances, growth rates, and production of tropical neritic copepods off Kingston, Jamaica. Mar Biol 106:79–89
Clarke KR, Warwick RM (1994) Changes in marine communities: an approach to statistical analysis and interpretation. Natural Environment Research Council, Plymouth
Costa RM, Leite NR, Pereira LCC (2009) Mesozooplankton of the Curuçá estuary (Amazon coast, Brazil). J Coast Res Spec Issue 56:400–404
da Costa KG, Pereira LCC, da Costa RM (2008) Short and long-term temporal variation of the zooplankton in a tropical estuary (Amazon region, Brazil). Bol Mus Para Emílio Goeldi Cienc Nat 3:127–141
da Costa RM, Atique P, da Costa KG, Pereira LCC (2011) Seasonal and spatial variation in hydrological parameters and microzooplankton communities in an Amazonian estuary. J Coast Res Spec Issue 64:1477–1481
Dalal SG, Goswami SC (2001) Temporal and ephemeral variations in copepod community in the estuaries of Mandovi and Zuari–west coast of India. J Plankton Res 23:19–26
Dauvin J-C, Desroy N (2005) The food web in the lower part of the Seine estuary: a synthesis of existing knowledge. Hydrobiologia 540:13–27
Dias CO, Bonecker SLC (2008) Inter-annual variability of planktonic copepods in a tropical bay in southeastern Brazil. Braz Arch Biol Techn 51:731–742
Dias CO, Araujo AV, Bonecker SLC (2009) Seasonal variability of planktonic copepods (Copepoda: Crustacea) in a tropical estuarine region in Brazil. Zoologia 26:705–715
Diele K (2000) Life history and population structure of the exploited mangrove crab Ucides cordatus cordatus (Linnaeus, 1763) (Decapoda: Brachyura) in the Caeté estuary, North Brazil. PhD thesis, University of Bremen, Germany
Diretoria de Hidrografia e Navegação (DHN) (2008). http://www.mar.mil.br/~dhn/tabuas. Accessed 25 January 2008
Dobson M, Frid C (2009) Ecology of aquatic systems. Oxford University Press, New York
Duggan S, McKinnon AD, Carleton JH (2008) Zooplankton in an Australian tropical estuary. Estuar Coast 31:455–467
Eskinazi-Sant’Anna EM, Björnberg TKS (2006) Seasonal dynamics of mesozooplankton in Brazilian coastal waters. Hydrobiologia 563:253–268
Favareto LR, Perbiche-Neves G, Serafim-Júnior M, Sartori LP (2009) Selectivity of plankton nets over planktonic Copepoda in two sub-tropical estuaries. Acta Limnol Bras 21:67–77
Giarrizzo T, Saint-Paul U (2008) Ontogenetic and seasonal shifts in the diet of the pemecou sea catfish Sciades herzbergii (Siluriformes: Ariidae), from a macrotidal mangrove creek in the Curuçá estuary, northern Brazil. Rev Biol Trop 56:861–873
Giarrizzo T, Schwamborn R, Saint-Paul U (2011) Utilization of carbon sources in a northern Brazilian mangrove ecosystem. Est Coast Shelf Sci 95:447–457
Gómez-Erache M, Norbis W, Bastreri D (2000) Wind effect as forcing factor controlling distribution and diversity of copepods in a shallow temperate estuary (Solis Grande, Uruguay). Sci Mar 64:87–95
Greenwood JG (1981) Occurrences of congeneric pairs of Acartia and Pseudodiaptomus species (Copepoda; Calanoida) in Moreton Bay, Queensland. Est Coast Shelf Sci 13:591–596
Guimarães DO, Pereira LCC, Monteiro M, Gorayeb A, da Costa RM (2009) Effects of urban development on the Cereja River and Caeté Estuary (Amazon coast, Brazil). J Coast Res Spec Issue 56:1219–1223
Hoffmeyer MS, Torres ER (2001) Morphometric variables and individual volume of Eurytemora americana and Acartia tonsa females (Copepoda, Calanoida) from the Bahía Blanca estuary, Argentina. Hydrobiologia 459:73–82
Hopcroft RR, Roff JC, Lombard D (1998) Production of tropical copepods in Kingston Harbour, Jamaica: the importance of small species. Mar Biol 130:593–604
Hubareva E, Svetlichny L, Kideys A, Isinibilir M (2008) Fate of the Black Sea Acartia clausi and Acartia tonsa (Copepoda) penetrating into the Marmara Sea through the Bosphorus. Est Coast Shelf Sci 76:131–140
Instituto Nacional de Meteorologia (INMET) (2012) The INMET register. http://www.inmet.gov.br. Accessed 28 May 2012
Katajisto T (2006) Benthic resting eggs in the life cycles of calanoid copepods in the northern Baltic Sea. PhD thesis, University of Helsinki, Finland
Kiørboe T (1997) Population regulation and role of mesozooplankton in sha** marine pelagic food webs. Hydrobiologia 363:13–27
Kjerfve B, Lacerda LD, Diop ES (1997) Mangrove ecosystem studies in Latin America and Africa. UNESCO, Paris
Koch V, Wolff M (2002) Energy budget and ecological role of mangrove epibenthos in the Caeté estuary, north Brazil. Mar Ecol Prog Ser 228:119–130
Koch V, Wolff M, Diele K (2005) Comparative population dynamics of four fiddler crabs (Ocypodidae, genus Uca) from a north Brazilian mangrove ecosystem. Mar Ecol Prog Ser 291:177–188
Krumme U, Liang T-H (2004) Tidal-induced changes in a copepod-dominated zooplankton community in a macrotidal mangrove channel in northern Brazil. Zool Stud 43:404–414
Lawrence D, Valiela I, Tomasky G (2004) Estuarine calanoid copepod abundance in relation to season, salinity, and land-derived nitrogen loading, Waquoit Bay, MA. Est Coast Shelf Sci 61:547–557
Leandro SM, Morgado F, Pereira F, Queiroga H (2007) Temporal changes of abundance, biomass and production of copepod community in a shallow temperate estuary (Ria de Aveiro, Portugal). Est Coast Shelf Sci 74:215–222
Lee WY, McAlice BJ (1979) Seasonal succession and breeding cycles of three species of Acartia (Copepoda: Calanoida) in a Maine estuary. Estuaries 2:228–235
Leite NR, Pereira LCC, da Costa RM (2009) Temporal distribution of the mesozooplankton in the Muriá creek, Pará, Brazil. Bol Mus Para Emílio Goeldi Cienc Nat 4:149–164
Leite NR, Pereira LCC, Abrunhosa F, Pires MAB, da Costa RM (2010) Occurrence of Cymbasoma longispinosum Bourne, 1890 (Copepoda: Monstrilloida) in the Curuçá river estuary (Amazon littoral). An Acad Bras Ciênc 82:577–583
Lopes RM, Vale R, Brandini FP (1998) Zooplankton composition, abundance and spatial distribution in the estuarine complex of Paranaguá during winter 1993 and summer 1994. Rev Bras Oceanogr 46:195–211
Madhu NV, Jyothibabu R, Balachandran KK, Honey UK, Martin GD, Vijay JG, Shiyas CA, Gupta GVM, Achuthankuty CT (2007) Monsoonal impact on planktonic standing stock and abundance in a tropical estuary (Cochin backwaters–India). Est Coast Shelf Sci 73:54–64
Magalhães A, Costa RM, Liang T-H, Pereira LCC, Ribeiro MJS (2006) Spatial and temporal distribution in density and biomass of two Pseudodiaptomus species (Copepoda: Calanoida) in the Caeté River Estuary (Amazon region–North of Brazil). Braz J Biol 66:421–430
Magalhães A, Bessa RSC, Pereira LCC, da Costa RM (2009a) Temporal variation in composition, occurrence and distribution of Copepoda (Crustacea) from Taperaçu estuary, Pará, Brazil. Bol Mus Para Emílio Goeldi Cienc Nat 4:133–148
Magalhães A, Leite NR, Silva JGS, Pereira LCC, da Costa RM (2009b) Seasonal variation in the copepod community structure from a tropical Amazon estuary, northern Brazil. An Acad Bras Ciênc 81:187–197
Magalhães A, Pereira LCC, Ribeiro MJS, Liang T-H, da Costa RM (2010) Populational dynamics of Pseudodiaptomus marshi (Crustacea: Copepoda) in the Caeté Estuary (Brazil). Trop Oceanogr 38:163–173
Magalhães A, Nobre DSB, Bessa RSC, Pereira LCC, da Costa RM (2011) Seasonal and short-term variations in the copepod community of a shallow Amazon estuary (Taperacu, northern Brazil). J Coast Res Spec Issue 64:1520–1524
Marcolin CR, Conceição BL, Nogueira MM, Júnior PM, Johnsson R (2010) Mesozooplankton and ichthyoplankton composition in two tropical estuaries of Bahia, Brazil. Check List 6:210–216
Marengo JA (1995) Interannual variability of deep convection over the tropical South American sector as deduced from ISCCP C2 data. Int J Climatol 15:995–1010
Marques SC, Pardal MA, Pereira MJ, Gonçalves F, Marques JC, Azeiteiro UM (2007) Zooplankton distribution and dynamics in a temperate shallow estuary. Hydrobiologia 587:213–223
Matos JB, Sodré DKL, da Costa KG, Pereira LCC, da Costa RM (2011) Spatial and temporal variation in the composition and biomass of phytoplankton in an Amazon estuary. J Coast Res Spec Issue 64:1525–1529
Islam MDS, Ueda H, Tanaka M (2006) Spatial and seasonal variations in copepod communities related to turbidity maximum along the Chikugo estuarine gradient in the upper Ariake Bay, Japan. Est Coast Shelf Sci 68:113–126
Medeiros GF, Medeiros LS, Henriques DMF, Carlos MTL, Faustino GVBS, Lopes RM (2006) Current distribution of the exotic copepod Pseudodiaptomus trihamatus Wright, 1937 along the northeastern coast of Brazil. Braz J Oceanogr 54:241–245
Mehlig U (2001) Aspects of tree primary production in an equatorial mangrove forest in Brazil. PhD thesis, University of Bremen, Germany
Montú M, Gloeden IM (1986) Atlas dos Cladocera e Copepoda (Crustacea) do estuário da Lagoa dos Patos (Rio Grande, Brasil). Nerítica 1:1–134
Moraes BC, Costa JMN, Costa ACL, Costa MH (2005) Variação espacial e temporal da precipitação no estado do Pará. Acta Amazon 35:207–214
Mouny P, Dauvin JC (2002) Environmental control of mesozooplankton community structure in the Seine estuary (English channel). Oceanol Acta 25:13–22
Muxagata E, Amaral WJA, Barbosa CN (2012) Acartia tonsa production in the Patos Lagoon estuary, Brazil. ICES J Mar Sci 69:475–482
Nagelkerken I, Blaber SJM, Bouillon S, Green P, Haywood M, Kirton LG, Meynecke J-O, Pawlik J, Penrose HM, Sasekumar A, Somerfield PJ (2008) The habitat function of mangroves for terrestrial and marine fauna: a review. Aquat Bot 89:155–185
Paffenhöfer G-A (1991) Some characteristics of abundant subtropical copepods in estuarine, shelf and oceanic waters. Bull Plankton Soc Japan Spec 201–216
Paffenhöfer G-A, Stearns DE (1988) Why is Acartia tonsa (Copepoda: Calanoida) restricted to nearshore environments? Mar Ecol Prog Ser 42:33–38
Palumbi SR (2001) Humans as the world’s greatest evolutionary force. Science 293:1786–1790
Park GS, Marshall HG (2000) Estuarine relationships between zooplankton community structure and trophic gradients. J Plankton Res 22:121–135
Peck MA, Holste L (2006) Effects of salinity, photoperiod and adult stocking density on egg production and egg hatching success in Acartia tonsa (Calanoida: Copepoda): optimizing intensive cultures. Aquaculture 255:341–350
Pielou EC (1977) Mathematical ecology. Wiley, New York
Putland JN, Iverson RL (2007) Ecology of Acartia tonsa in Apalachicola Bay, Florida, and implications of river water diversion. Mar Ecol Prog Ser 340:173–187
Rakhesh M, Raman AV, Ganesh T, Chandramohan P, Dehairs F (2013) Small copepods structuring mesozooplankton community dynamics in a tropical estuary-coastal system. Est Coast Shelf Sci 126:7–22
Riccardi N (2010) Selectivity of plankton nets over mesozooplankton taxa: implications for abundance, biomass and diversity estimation. J Limnol 69:287–296
Rios-Jara E (1998) Spatial and temporal variations in the zooplankton community of Phosphorescent Bay, Puerto Rico. Est Coast Shelf Sci 46:797–809
Rodriguez V, Guerrero F, Bautista B (1995) Egg production of individual copepods of Acartia grani Sars from coastal waters: seasonal and diel variability. J Plankton Res 17:2233–2250
Sankarankutty C, Medeiros GF, Oliveira JEL (1995) Diurnal and seasonal fluctuations of estuarine zooplankton of Potengi estuary, Natal, Northeast of Brazil. J Mar Biol Assoc India 37:201–211
Schories D, Mehlig U (2000) CO2 gas exchange of benthic microalgae during exposure to air: a technique for the rapid assessment of primary production. Wetl Ecol Manag 8:273–280
Schwamborn R, Bonecker SLC, Galvão IB, Silva TA, Neumann-Leitão S (2004) Mesozooplankton grazing under conditions of extreme eutrophication in Guanabara Bay, Brazil. J Plankton Res 26:983–992
Shannon CE (1948) A mathematical theory of communication. Bell Syst Technol J 27:379–423
Souza-Filho PWM, da Sales MEC, da Prost MTRC, Costa FR, de Souza LFMO (2005) Zona costeira amazônica: o cenário regional e os indicadores bibliométricos em C & T (volume 1). In: Souza-Filho PWM, da Cunha ERSP, da Sales MEC, Costa FR (eds) Bibliografia da zona costeira amazônica. Museu Paraense Emilio Goeldi/Universidade Federal do Pará/Petrobras, Belém, pp 9–20
Sterza JM, Fernandes LL (2006) Zooplankton community of the Vitória Bay estuarine system (southeastern Brazil). characterization during a three-year study. Braz J Oceanogr 54:95–105
Sullivan BK, McManus LT (1986) Factors controlling seasonal succession of the copepods Acartia hudsonica and A. tonsa in Narragansett Bay, Rhode Island: temperature and resting egg production. Mar Ecol Prog Ser 28:121–128
ter Braak CJF, Smilauer P (1998) CANOCO reference manual and user’s guide to Canoco for Windows: software for canonical community ordination (version 4). Ithaca, New York
Tester PA, Turner JT (1991) Why is Acartia tonsa restricted to estuarine habitats? Bull Plankton Soc Japan Spec Vol 603-611
Uye S-I, Kuwata H, Endo T (1987) Standing stocks and production rates of phytoplankton and planktonic copepods in the Inland Sea of Japan. J Oceanogr Soc Jpn 42:421–434
World Register of Marine Species (WoRMS) (2012). The WoRMS register. http://www.marinespecies.org. Accessed 8 Mar 2012
Zar JH (2010) Biostatistical analysis. Prentice Hall Inc, New Jersey
Acknowledgments
We thank the Universidade Federal do Pará, Instituto de Estudos Costeiros for providing logistical support during this survey. We are also indebted to Dr. Stephen Ferrari for his careful correction of the English. The author André Magalhães is grateful to Coordenação de Aperfeiçoamento de Pessoal de Nível Superior for the concession of PhD scholarship. The authors thank two anonymous referees for their useful remarks on the original version of the manuscript. This research was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (André Magalhães Grant Numbers 304061/2003–4 and 553062/2005–0, Luci Cajuerio C. Pereira Grant Numbers 308379/2011–0 and 200629/2014–0, and Rauquírio Marinho da Costa Grant Numbers 306061/2011–2 and 200622/2014–5).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.-D. Franke.
Rights and permissions
About this article

Cite this article
Magalhães, A., Pereira, L.C.C. & da Costa, R.M. Relationships between copepod community structure, rainfall regimes, and hydrological variables in a tropical mangrove estuary (Amazon coast, Brazil). Helgol Mar Res 69, 123–136 (2015). https://doi.org/10.1007/s10152-014-0421-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-014-0421-4