
Abstract
Background
We have previously identified 1,25-dihydroxyvitamin D3 [1,25(OH)2D3], the bioactive form of vitamin D3, as a potent regulator of energy-utilization and nutrient-sensing pathways in prostate cancer cells. In the current study, we investigated the effects of 1,25(OH)2D3 on breast cancer (BCa) cell metabolism using cell lines representing distinct molecular subtypes, luminal (MCF-7 and T-47D), and triple-negative BCa (MDA-MB-231, MDA-MB-468, and HCC-1143).
Methods
1,25(OH)2D3’s effect on BCa cell metabolism was evaluated by employing a combination of real-time measurements of glycolysis/oxygen consumption rates using a biosensor chip system, GC/MS-based metabolomics, gene expression analysis, and assessment of overall energy levels. The influence of treatment on energy-related signaling molecules was investigated by immunoblotting.
Results
We show that 1,25(OH)2D3 significantly induces the expression and activity of the pentose phosphate pathway enzyme glucose-6-phosphate dehydrogenase (G6PD) in all BCa cell lines, however differentially influences glycolytic and respiratory rates in the same cells. Although 1,25(OH)2D3 treatment was found to induce seemingly anti-oxidant responses in MCF-7 cells, such as increased intracellular serine levels, and reduce the expression of its putative target gene thioredoxin-interacting protein (TXNIP), intracellular reactive oxygen species levels were found to be elevated. Serine accumulation in 1,25(OH)2D3-treated cells was not found to hamper the efficacy of chemotherapeutics, including 5-fluorouracil. Detailed analyses of the nature of TXNIP’s regulation by 1,25(OH)2D3 included genetic and pharmacological inhibition of signaling molecules and metabolic enzymes including AMP-activated protein kinase and G6PD, as well as by studying the ITCH (E3 ubiquitin ligase)-TXNIP interaction. While these investigations demonstrated minimal involvement of such pathways in the observed non-canonical regulation of TXNIP, inhibition of estrogen receptor (ER) signaling by tamoxifen mirrored the reduction of TXNIP levels by 1,25(OH)2D3, demonstrating that the latter’s negative regulation of ER expression is a potential mechanism of TXNIP modulation.
Conclusions
Altogether, we propose that regulation of energy metabolism contributes to 1,25(OH)2D3’s anti-cancer effects and that combining 1,25(OH)2D3 with drugs targeting metabolic networks in tumor cells may lead to synergistic effects.
Similar content being viewed by others

Background
Breast cancer (BCa) is the main cause of cancer death in females in both developed and develo** countries [1]. Several risk factors have been implicated in the pathogenesis of the disease, including lifestyle-related factors, for example smoking, obesity, and physical inactivity, as well as genetic factors, like mutations in breast cancer susceptibility genes (BRCA 1 and 2) [1, 2]. In addition to these risk factors, mounting evidence has demonstrated that vitamin D deficiency contributes to both BCa incidence and survival [3, 4].
Vitamin D is a seco-steroid that is produced in humans upon exposure of the skin to ultraviolet-B radiation [5]. The resulting hormonal precursor molecule is further metabolized to yield its hormonally active form—1,25-dihydroxyvitamin D (1,25(OH)2D3, also known as calcitriol)—through a hydroxylation step in the liver, followed by a second activating hydroxylation in the kidneys [5]. 1,25(OH)2D3 subsequently binds to its nuclear vitamin D receptor (VDR), present in diverse cell types, and regulates the expression of hundreds of genes known to influence proliferation, differentiation, and angiogenesis [5]. Both in vitro and animal studies have illustrated that 1,25(OH)2D3 and its analogues are promising chemotherapeutics against BCa [3, 6], with reports showing induction of cell cycle arrest and apoptosis, as well as regulation of estrogen signaling with treatment [3, 6]. Additionally, studies have shown that VDR expression inversely correlates with breast cancer mortality and that CYP24A1, the vitamin D-catabolizing enzyme, is a putative oncogene [6].
BCa is classified into a number of molecular subtypes, namely (i) basal-like, also known as triple-negative BCa (TNBC) due to the absence of estrogen receptor (ER), progesterone receptor, and HER2; (ii) luminal A/B, characterized by the presence of ER; and finally, (iii) HER2 positive BCa, with amplified ERBB2 gene expression [7, 8]. Clinically, these subtypes exhibit different responsiveness to chemotherapy and thus are administered different therapy regimens [9]. For example, luminal BCa is typically treated with chemotherapeutics that interfere with ER signaling, such as anti-estrogens and aromatase inhibitors [10]. Additionally, although controversial, different BCa subtypes exhibit distinct aberrations in their metabolic profiles [10, 11]. For instance, ER+ BCa has been shown to exhibit more “classical” Warburg metabolism, illustrated by an increase in glucose consumption and lactate production, whereas ER− BCa is known to rely more on glutamine metabolism and subsequent TCA replenishment/anaplerosis [10]. Moreover, metabolic adaptation has been reported as a potential resistance mechanism, which BCa cells adopt in response to hormone therapy [12]. Therefore, identification of drugs that target these altered metabolic networks may prove beneficial in the treatment of BCa as monotherapies, as well as enhance the efficacy/reduce resistance associated with currently established chemotherapeutics.
Recent work has shown that 1,25(OH)2D3 modulates glucose, glutamine, and fatty acid metabolism in several experimental models including breast and prostate cancer cells [13,14,15,16,17], which prompted us to thoroughly evaluate the ability of this molecule to regulate metabolic networks in BCa cell lines representing different molecular subtypes. The effect of 1,25(OH)2D3 on energy metabolism of luminal (MCF-7 and T-47D) and TNBC (MDA-MB-231, MDA-MB-468, and HCC-1143) cells was evaluated using real-time measurements of glycolytic/respiratory rates, GC/MS-based quantification of TCA cycle intermediates and diverse amino acids, mRNA expression analysis of metabolism-related genes, and finally overall energy charge. 1,25(OH)2D3 was found to induce both similar and different metabolic effects in these cell lines, such as induction in glucose-6-phosphate dehydrogenase (G6PD) expression and activity in all cell lines, and disparate regulation of glycolytic and respiratory capacities. In MCF-7 cells, seemingly pro-survival metabolic perturbations induced by treatment, such as accumulation of intracellular serine, were not found to antagonize the anti-tumor efficacy of chemotherapeutics including 5-fluorouracil (5-FU). Furthermore, 1,25(OH)2D3 was found to negatively regulate TXNIP expression in MCF-7 cells, possibly through reduction of estrogen receptor (ER) expression.
Methods
Cell culture
The human BCa cell lines MCF-7, T-47D, MDA-MB-231, MDA-MB-468, and HCC-1143 were cultured in Dulbecco’s Modified Eagle Medium (DMEM) (Gibco, Germany) containing 10% FCS (v/v) (Gibco, Germany) and 1% penicillin/streptomycin (v/v) (Gibco, Germany) and kept in a standard cell culture incubator (37 °C; 5% CO2). T-47D, MDA-MB-468, and HCC-1143 were kind gifts from Dr. Stefan Wiemann (DKFZ, Heidelberg). 1,25(OH)2D3 (Cayman Chemical-Biomol GmbH, Germany) treatments were performed for the various indicated time points in standard medium, at a final concentration of 100 nM. Additional drugs used in the study were added to either fresh or conditioned medium for different time points (as described in the “Results” section) and included 10 mM 2-deoxyglucose (Fluka-Sigma-Aldrich, Germany), 5 μM MG-132 (Sigma-Aldrich, Germany), 20 μM leupeptin (Sigma-Aldrich, Germany), 100 nM calcipotriol (Cayman Chemical-Biomol GmbH, Germany), and 20 μM BAPTA-AM (Cayman Chemical-Biomol GmbH, Germany), as well as increasing concentrations of dehydroepiandrosterone (DHEA) (Cayman Chemical-Biomol GmbH, Germany), Na Oxamate (Cayman Chemical-Biomol GmbH, Germany), AZD-3965 (Cayman Chemical-Biomol GmbH, Germany), 5-fluorouracil (5-FU) (Fluka-Sigma-Aldrich, Germany), and CBR-5884 (Cayman Chemical-Biomol GmbH, Germany).
RNA isolation, cDNA synthesis, and RT-qPCR
BCa cells were treated with 1,25(OH)2D3 (100 nM) for 72 h, after which total RNA was isolated using QIAzol lysis reagent (Qiagen, Germany), following the manufacturer’s instructions. RNA concentration and purity were determined using NanoDrop 2000 UV-Vis Spectrophotometer (Thermo Scientific, Germany). cDNA was synthesized from 500 ng of total RNA/sample using ProtoScript® II first strand cDNA synthesis kit (New England BioLabs, Germany). The thermal cycle qTower (Analytik Jena AG) was used to assess mRNA expression of various genes in real time. Primers used in the study are listed in Additional file 1: Table S1. The LightCycler® 480 SYBR Green I Master (Roche, Germany) reaction mix was used. The ∆∆Ct method was used to calculate the relative expression of the investigated genes in response to treatment, with vinculin as the housekee** gene.
RNA interference
Anti-AMPKα1 and anti-G6PD siRNAs were synthesized by Riboxx (Riboxx GmbH, Radebeul, Germany) (Additional file 2: Table S2). MCF-7 cells were transfected with negative control, anti-AMPKα1, or anti-G6PD oligonucleotides as described in an earlier study [16]. Briefly, 50 pmol of the respective siRNA was diluted in 100 μL Opti-MEM Reduced Serum Medium (Gibco, Germany)/well in a 24-well plate and complexed with the 1.5 μL transfection reagent Lipofectamine® 3000 (Thermo Fischer, Germany)/well. Sixty thousand cells suspended in 500 μL of DMEM (10% FCS, without antibiotics) were added to each well and mixed gently. The plate was kept overnight in a standard incubator, and the treatment was performed on the following day as indicated.
On-line measurement of extracellular acidification, oxygen consumption, and impedance using BIONAS 2500
Changes in cellular glycolytic, respiratory, and impedance rates were monitored in response to 1,25(OH)2D3 (100 nM) treatment using the Bionas 2500 biosensor chip system (Bionas, Rostock, Germany) as previously described [16, 18]. Briefly, 6 sensor chips (SC1000) were seeded with BCa cells at a density of 200,000 cells/chip in 450 μL of full medium and kept in a standard cell culture incubator overnight. The 6 chips were then transferred to the sensor chip system, where either 3 were continuously fed running medium (RM) containing either DMSO or 1,25(OH)2D3. The RM used was prepared from DMEM powder (Pan-Biotech GmbH, Germany) containing neither glucose, sodium pyruvate, L-glutamine, NaHCO3, nor phenol red. The medium was then supplemented with 1 g/L glucose, 2 mM glutamine, 1 mM HEPES, and 10 mg/L phenol red. Additionally, 0.1% (v/v) FCS and 1% (v/v) penicillin/streptomycin were added. Data were analyzed using the software provided by the manufacturer.
Glucose uptake and ATP measurements
With regard to glucose uptake, culture medium was replaced with medium containing the 50 μM of the fluorescently labeled glucose analogue, 2-[N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl) amino]-2-deoxy-D-glucose (2-NBDG) (Cayman Chemical-Biomol GmbH, Germany), at the end of the indicated treatment periods. Cells were harvested 2 h later, and FACS analysis was performed to determine the amount of intracellular 2-NBDG.
Intracellular ATP levels were assessed using the ATPlite™ 1 step (Perkin Elmer, Germany), following the manufacturer’s instructions. Cells were seeded at a density of 5000 cells/well in a black 96-well plate with a transparent bottom and placed inside a standard cell culture incubator overnight. The cells were then treated for 24 h with 100 μL of full medium containing either DMSO or 1,25(OH)2D3 (100 nM), after which 100 μL of the substrate solution was added to the wells, and luminescence was measured kinetically using the Tecan Ultra plate reader (Tecan, Germany). The signal obtained from each well was then normalized to the corresponding protein content.
Analysis of glucose-6-phosphate dehydrogenase (G6PD) enzymatic activity
G6PD enzymatic activity was determined as previously described [19, Full size image
To further investigate the influence of treatment on glucose metabolism, we studied the regulation of TCA cycle intermediates by 1,25(OH)2D3 using GC/MS in representative cell lines of each molecular subtype, MCF-7 (luminal) and MDA-MB-231 (TNBC) cells. Interestingly, the levels of the investigated intermediates were differentially impacted by 1,25(OH)2D3, where treatment was found to reduce citrate levels and the levels of downstream metabolites in MCF-7 cells (Fig. 1b). In MDA-MB-231 cells on the contrary, citrate levels were found to be increased by 1,25(OH)2D3 (Fig. 1b). Additionally, we investigated the levels of select essential and non-essential amino acids in response to 1,25(OH)2D3 in both cell lines. While levels of all investigated amino acids were altered in both cell lines by 1,25(OH)2D3 treatment to varying degrees, a profound increase in serine levels was observed in MCF-7 cells (Fig. 1b). Additionally, glycine levels were found to be reduced and increased in response to 1,25(OH)2D3 in MCF-7 and MDA-MB-231 cells, respectively (Fig. 1b).
In view of the effects 1,25(OH)2D3 exerted on the investigated metabolic parameters in BCa cells, we postulated that such effects may at least in part be explained by 1,25(OH)2D3-mediated changes in the expression of metabolism-related genes. We thus performed a comprehensive mRNA screen of genes encoding players involved in diverse metabolic processes, such as glucose, glutamine, and fatty acid metabolism, in MCF-7 and MDA-MB-231 cells. Furthermore, given the different p53 backgrounds exhibited by the two investigated cell lines, and since p53 is known to be a powerful regulator of cell metabolism and associated processes (e.g., autophagy) [24, 25], we included in our screen a number of p53 target genes, as well as genes encoding autophagy regulators. While several investigated genes were only mildly regulated by 1,25(OH)2D3 treatment in both MCF-7 and MDA-MB-231 cells, a clear induction in the mRNA levels of the G6PD gene, encoding glucose-6-phosphate dehydrogenase, was observed in both cell lines (Fig. 1c). Similarly, a significant induction in G6PD protein expression was observed with 1,25(OH)2D3 treatment in both cell lines (Fig. 1d, e). G6PD catalyzes the first rate-limiting step in the PPP, which provides cells with reducing equivalents for both reductive biosynthesis and protection against oxidative stress [26]. On the other hand, G6PD expression has been shown to be elevated in cancers and its activity induced by oncogenic signaling [26, 27]. With regard to vitamin D, the promoter of the G6PD gene has been shown to harbor vitamin D response elements [28], and 1,25(OH)2D3 has been repeatedly shown to induce either the expression and/or activity of this enzyme in different experimental models, including BCa [29, 30]. We then aimed to investigate if the upregulation of G6PD expression with 1,25(OH)2D3 treatment is observed in the other BCa cell lines. Indeed, G6PD mRNA and protein levels were markedly induced with 1,25(OH)2D3 in all the investigated BCa cell lines (Fig. 1f, g). Importantly, G6PD enzymatic activity in all cell lines was found to be significantly increased in response to treatment (Fig. 1h).
We also sought to investigate changes in cellular ATP levels in response to 1,25(OH)2D3 treatment in all BCa cell lines. In MCF-7 cells, 1,25(OH)2D3 was found to induce fluctuations in cellular ATP levels over the investigated time course, with an overall significant decrease with treatment after 72 h (Fig. 1i). In MDA-MB-231 cells, no clear differences in ATP levels were observed with treatment after 24 h; however, a profound induction was observed after 48 and 72 h (Fig. 1i).
Seemingly pro-survival, 1,25(OH)2D3-induced metabolic changes do not hamper the efficacy of different anti-cancer strategies
Results of the metabolic characterization described in the previous section indicate that certain effects induced by 1,25(OH)2D3 might enhance cancer cell proliferation and survival. For example, in MCF-7 cells, treatment was found to increase intracellular serine levels and G6PD expression and activity. G6PD, as previously mentioned, is a putative oncogene, whose expression significantly correlates with reduced overall survival in ER+ and total BCa patients (Fig. 2a). Similarly, serine is a recognized onco-metabolite that is known to contribute to both the folate and methionine cycles, thus enhancing nucleotide biosynthesis, NADPH production, and S-adenosylmethionine generation, possibly inducing epigenetic changes [31].

Seemingly pro-survival metabolic effects of 1,25(OH)2D3 do not hamper the efficacy of metabolism-targeting therapeutic regimens. a Kaplan-Meier plots demonstrating the significant inverse correlation between G6PD mRNA expression and overall survival in ER+ (left) and total (right) BCa patients. b Knocking-down G6PD in MCF-7 cells significantly reduces cell survival, independent of 1,25(OH)2D3 (100 nM; 72 h). Statistical comparison between DMSO- and 1,25(OH)2D3-treated cells was made using a two-tailed Student’s t test. P values less than or equal to 0.05, 0.01 and 0.001 are depicted by *, **, and ***, respectively. Error bars ± SD; n = 3. Cellular survival was assessed using SRB assay, and % survival is calculated by normalizing the absorbance value obtained with the different experimental conditions to DMSO-treated cells transfected with negative control (NC) siRNA. 1,25(OH)2D3 significantly enhances the anti-tumor effects of DHEA assessed by SRB assay. MCF-7 cells were treated for 72 h with increasing concentrations of DHEA in the presence of either DMSO or 1,25(OH)2D3, and % survival is calculated by normalizing the absorbance value obtained with the different experimental conditions to DMSO-treated cells. c Schematic overview of cellular serine uptake, synthesis, and metabolism. 3PS 3-phosphoserine. d 72 h treatment of MCF-7 cells with 1,25(OH)2D3 differentially influences the mRNA expression of enzymes involved in serine synthesis but upregulates the expression of those involved in serine metabolism. Relative expression was calculated using the ∆∆Ct method, with vinculin as the housekee** gene. e 1,25(OH)2D3 significantly influences cellular survival in response to amino acid deprivation. MCF-7 cells were cultivated in medium lacking serine, glycine, or both, in the presence of either DMSO or 1,25(OH)2D3 for 72 h, after which SRB assay was performed. 1,25(OH)2D3 was found to mildly however significantly dampen the reduction in survival of MCF-7 cells in response to serine deprivation. 1,25(OH)2D3 was also found to enhance the efficacy of CBR-5884 and 5-FU, inhibitors of PHGDH and TYMS, respectively, assessed by SRB assay
G6PD inhibition has been previously shown to limit cancer cell survival [32]. To investigate if 1,25(OH)2D3’s induction of G6PD expression and activity would hamper such anti-tumor strategies, G6PD was genetically and pharmacologically inhibited in MCF-7 cells using siRNA and DHEA, respectively, in the presence and absence of 1,25(OH)2D3, and cellular survival was assessed by SRB assay. While 1,25(OH)2D3 treatment did not dampen the significant reduction in cell survival in response to G6PD knockdown, the inhibitory (anti-cancer) effect of DHEA was found to be enhanced in the presence of 1,25(OH)2D3 (Fig. 2b).
We then aimed at investigating the underlying mechanism of serine accumulation in response to 1,25(OH)2D3 in MCF-7 cells. Serine is a non-essential amino acid that cells acquire either by de novo synthesis, using the glycolytic intermediate 3-phosphoglycerate (3PG) as a starting molecule, or by uptake from the medium (Fig. 2c). We therefore investigated changes in the mRNA expression of players involved in serine synthesis, metabolism, and uptake, in response to 1,25(OH)2D3 treatment of MCF-7 cells. While treatment was found to differentially regulate the mRNA expression of enzymes in the serine synthesis pathway (SSP), for example non-significant alteration of PHGDH (phosphoglycerate dehydrogenase), induction of PSAT1 (phosphoserine aminotransferase 1), and reduction of PSPH (phosphoserine phosphatase), enzymes involved in serine metabolism, namely serine hydroxymethyltransferase 1 (SHMT1) and 2 (SHMT2), as well as thymidylate synthase (TYMS), were found to be significantly induced with 1,25(OH)2D3 (Fig. 2d). SHMT1 and SHMT2 are responsible for the interconversion of serine and glycine, and, concurrently, tetrahydrofolate (THF) and 5,10-methylene-THF [31]. The latter could be used in the synthesis of thymidylate, in a reaction catalyzed by TYMS, which is the intracellular target of the classical chemotherapeutic 5-fluorouracil (5-FU) [33].
To investigate the impact of serine accumulation in 1,25(OH)2D3-treated MCF-7 cells on the efficacy of 5-FU, cells were treated with increasing concentrations of 5-FU, alone and in combination with 1,25(OH)2D3, and cellular survival was evaluated. The addition of 1,25(OH)2D3 was found to significantly improve 5-FU’s anti-cancer effects (Fig. 2e). We then aimed to elucidate whether serine accumulation was the result of increased de novo synthesis or uptake. We therefore treated MCF-7 cells with the small molecule PHGDH inhibitor CBR-5884 in the presence and absence of 1,25(OH)2D3. PHGDH catalyzes the first reaction of the SSP, namely the conversion of 3PG to 3-phosphopyruvate (3PHP) [31]. CBR-5884 has been shown to be toxic to cancer cells that depend on de novo serine synthesis, but not to cancer cells reliant on extracellular sources for serine [34]. Mullarky et al. [34] demonstrated that MCF-7 cells are dependent on serine in the medium and were therefore insensitive to CBR-5884. However, upon cultivation in serine-depleted medium, MCF-7 cells were rendered susceptible to the drug’s toxic effects [34]. We thus postulated that an increased SSP activity in response to 1,25(OH)2D3 would render MCF-7 sensitive to CBR-5884’s anti-tumor effects on the one hand, as well as dampen the growth inhibiting effects of serine deprivation on the other. Indeed, treating MCF-7 cells with a combination of 1,25(OH)2D3 and increasing concentrations of CBR-5884 led to substantial inhibition of proliferation compared to cells treated with CBR-5884 alone (Fig. 2e). Additionally, 1,25(OH)2D3 was found to mildly however significantly reduce the sensitivity of MCF-7 cells to serine deprivation. Altogether, these results indicate that 1,25(OH)2D3 may enhance the ability of cells to synthesize serine. It remains unclear, however, if the proposed increase in serine synthesis utilizes the glycolytic metabolite 3PG or glycine as a starting intermediate. In support of the latter possibility is the reduction in glycine levels and the induction in SHMT1 and 2 mRNA expression in 1,25(OH)2D3-treated MCF-7 cells.
1,25(OH)2D3 differentially regulates the AMPK-TXNIP signaling axis in BCa cells
In view of the clear metabolic rewiring induced by 1,25(OH)2D3 in BCa cells, we hypothesized that key signaling molecules involved in regulating energy levels and glucose homeostasis might be affected by treatment. We thus investigated changes in TXNIP expression and AMPK signaling activation in BCa cells in response to 1,25(OH)2D3 treatment. TXNIP, originally identified by Chen and DeLuca in HL-60 cells as the vitamin D upregulated protein 1 (VDUP1) [35], regulates both redox balance and glucose uptake [36]. With regard to redox regulation, TXNIP binds to reduced thioredoxin and prevents its anti-oxidant actions [36]. More recent studies however have shown that TXNIP is a pivotal member of glucose-sensing mechanisms, involving glycolytic intermediates, glucose transporters, and the heterodimers MondoA-MLX [37, 38]. Briefly, an increase in glucose uptake and subsequently glycolytic intermediates triggers the nuclear translocation of the aforementioned heterodimer, which binds to carbohydrate response elements on the TXNIP gene, thereby inducing its expression [37]. TXNIP in turn reduces glucose uptake by reducing the expression and inducing the internalization of glucose transporter 1 [38]. AMPK on the other hand is a heterotrimeric complex that acts as an intracellular energy gauge through sensing changes in the AMP:ATP ratio [39]. In response to energetic stress, AMPK inhibits energy-consuming processes such as fatty acid synthesis and activates energy-producing processes, including glucose uptake and mitochondrial respiration [39]. Recently, AMPK has been shown to induce glucose uptake through triggering the degradation of TXNIP [38].
BCa cells were therefore treated with 1,25(OH)2D3 for 72 h, after which TXNIP levels and AMPK signaling activity were investigated using immunoblotting. Treatment was found to activate AMPK signaling, as observed by the increased phosphorylation of acetyl coA carboxylase (ACC; serine 79)—AMPK’s substrate—in MCF-7 and MDA-MB-231 cells (Fig. 3a), which correlates well with the alterations in ATP levels observed in these two cell lines with 1,25(OH)2D3 treatment (Fig. 1i). Furthermore, in cell lines expressing detectable levels of TXNIP, namely MCF-7, T-47D, and MDA-MB-231 cells, canonical regulation of TXNIP expression by 1,25(OH)2D3 was not observed (Fig. 3a). In fact, treatment was found to reduce TXNIP levels in MCF-7 cells (Fig. 3a), an effect that was statistically significant after both 24 and 72 h of treatment (Fig. 3b, c). This is in line with our previous studies demonstrating reduction of TXNIP levels by 1,25(OH)2D3 in prostate cancer cells [16], as well as the cell line-specific regulation of the protein by treatment [40].

Regulation of AMPK signaling and TXNIP expression in BCa cells by 1,25(OH)2D3. a The AMPK-TXNIP signaling axis was found to be differentially regulated by a 72-h treatment with 1,25(OH)2D3 (100 nM). AMPK signaling was found to be induced and TXNIP expression reduced in MCF-7 cells in response to treatment. Western blot and densitometric analysis of TXNIP expression in MCF-7 cells treated for 24 (b) and 72 h (c) with 1,25(OH)2D3. Statistical significance between DMSO- and 1,25(OH)2D3-treated cells is calculated using a two-tailed Student’s t test, where p values less than or equal to 0.05, 0.01, and 0.001 are depicted in the figures by *, **, and ***, respectively. Error bars ± SD; n = 3. d–g 72 h treatment with 1,25(OH)2D3 significantly induces intracellular ROS levels. Microscopic analysis of cellular superoxide using DHE staining (d) and associated quantification (e). Hoechst dye was used to stain nuclei. Mitochondrial superoxide was stained using MitoSOX Red, and mitochondria were stained using MitoTracker Green (f). Microscopic images were taken every 15 min for 5 h, and fluorescence intensity of MitoSOX Red was quantified (g)
Since TXNIP engages in both redox balance and glucose homeostasis [37, 41], we hypothesized that the reduction in TXNIP level in MCF-7 cells by treatment may also influence such critical cellular processes. We therefore evaluated intracellular ROS levels in DMSO- and 1,25(OH)2D3-treated MCF-7 cells using DHE and MitoSOX Red staining. Although 1,25(OH)2D3 was found to significantly induce G6PD expression and activity (Fig. 1d, h) as well as intracellular serine levels in MCF-7 cells (Fig. 1c), which together with the reduction in TXNIP expression serve as a strong basis for anti-oxidant activity, we observed significant ROS accumulation in response to treatment (Fig. 3d–g; Additional files 3 and 4: Movies S1 and S2). This is in line with the results of Koren et al. [42], who illustrated that 1,25(OH)2D3 may act as a pro-oxidant in MCF-7 cells by increasing the ratio of oxidized to reduced glutathione. Moreover, we observed a significant reduction in glucose uptake after 24, but not 72 h of treatment (Additional file 5: Figure S1), indicating that the reduction in intracellular glycolytic intermediates in response to 1,25(OH)2D3 may have led to the decrease in TXNIP expression.
Non-canonical regulation of TXNIP by 1,25(OH)2D3 is independent of metabolic changes
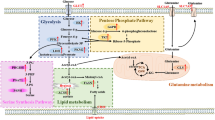
As previously mentioned, the TXNIP gene is a direct target of the glucose-sensing transcriptional heterodimer MondoA-MLX and is induced in response to an increase in upstream glycolytic intermediates, namely glucose-6-phosphate. We postulated that distinct metabolic changes induced by 1,25(OH)2D3 in MCF-7 cells, including increased G6PD expression and activity, induction of the SSP, and enhanced glycolytic rate, may contribute to the reduction in TXNIP expression by depleting the levels of glycolytic intermediates capable of inducing TXNIP levels. We thus postulated that genetic and/or pharmacological inhibition of such metabolic pathways in 1,25(OH)2D3-treated cells would rescue TXNIP expression (Fig. 4a).

TXNIP regulation by 1,25(OH)2D3 is independent of metabolic reprogramming. a Schematic overview of treatment-induced metabolic changes that may reduce levels of glycolytic intermediates capable of inducing the nuclear translocation of MondoA/MLX and hence TXNIP expression. Genetic and/or pharmacological strategies for inhibiting key metabolic players are depicted. b Time-dependent increase in G6PD mRNA expression in MCF-7 cells with 1,25(OH)2D3 (100 nM) is associated with an attenuation of the initial increase in TXNIP mRNA levels with treatment. Upper and lower ends of floating bars represent maximum and minimum values, respectively, whereas lines in the middle represent the means. The ∆∆Ct method was used to calculate the relative expression with vinculin as the housekee** gene. The data are obtained from two biological replicates. c Similar to the trend observed on the mRNA level, the induction in G6PD protein level in response to 1,25(OH)2D3 first occurs after 12 h, which coincides with the first reduction in TXNIP protein expression. d Co-treatment with DHEA, but not knocking-down G6PD levels, elevates TXNIP mRNA levels in 1,25(OH)2D3-treated MCF-7 cells. Cells transfected with either anti-G6PD siRNA or a NC were treated with 1,25(OH)2D3 for 72 h, whereas increasing concentrations of DHEA were added to the conditioned medium of MCF-7 cells treated for 48 h with either DMSO or 1,25(OH)2D3, for an additional 24 h. e G6PD inhibition using siRNA or DHEA reduces TXNIP protein expression independent of 1,25(OH)2D3. f–h Western blot analysis of TXNIP expression in MCF-7 cells treated with a combination of either DMSO or 1,25(OH)2D3 and various metabolic inhibitors. Cells were initially treated with DMSO or 1,25(OH)2D3 for 48 h, after which increasing concentrations of the different inhibitors were added to the conditioned medium for an additional 24 h. All inhibitors were found to reduce TXNIP protein levels in 1,25(OH)2D3-treated cells. In the absence of 1,25(OH)2D3, AZD 3965 and CBR-5884 were found to mildly induce TXNIP levels
Since G6P is a major glycolytic metabolite implicated in inducing TXNIP expression [43], we first investigated the impact of G6PD induction by 1,25(OH)2D3 on TXNIP regulation. Time-course studies of TXNIP and G6PD mRNA and protein regulation by 1,25(OH)2D3 illustrated that the initial induction in TXNIP mRNA levels by treatment (1–3 h) was found to be largely attenuated at 6 h, coinciding with the induction in G6PD mRNA levels by 1,25(OH)2D3 (Fig. 4b). Similarly, on the protein level, the reduction and induction of TXNIP and G6PD levels, respectively, in response to treatment, first occurs after 12 h (Fig. 4c), indicating that shunting of G6P into the PPP by G6PD may be responsible for TXNIP reduction. We then knocked-down G6PD levels in MCF-7 cells using siRNA and investigated TXNIP mRNA and protein regulation by 1,25(OH)2D3. Additionally, cells were treated with either DMSO or 1,25(OH)2D3 for 48 h, and DHEA—the non-competitive inhibitor of G6PD [44]—was added to the conditioned medium for an additional 24 h, after which TXNIP levels were also investigated. Independent of 1,25(OH)2D3, knocking-down G6PD levels were not found to influence TXNIP mRNA levels, whereas different concentrations of DHEA were found to induce TXNIP mRNA levels (Fig. 4d). Furthermore, TXNIP protein expression was found to be largely depleted, and reduced in a dose-dependent manner, in cells with knocked-down G6PD levels, and in DHEA-treated cells, respectively, independent of 1,25(OH)2D3 (Fig. 4e). We therefore concluded that the induction in G6PD expression and activity by treatment does not contribute to the reduction in TXNIP expression.
To assess the contribution of other 1,25(OH)2D3-induced metabolic changes to TXNIP regulation, MCF-7 cells were treated with increasing concentrations of Na oxamate and AZD 3965, inhibitors of lactate dehydrogenase and monocarboxylate transporter (MCT) 1, respectively, as well as CBR-5884, alone and in combination with 1,25(OH)2D3. While AZD 3965 and CBR-5884 led to slight increases in TXNIP protein levels, all inhibitors were found to reduce TXNIP expression in the presence of 1,25(OH)2D3 (Fig. 4f–h), indicating that inhibition of one metabolic pathway in 1,25(OH)2D3-treated cells does not lead to an accumulation of glycolytic intermediates since such metabolites might be shunted into other activated pathways.
1,25(OH)2D3 reduces TXNIP expression through inducing its proteasomal degradation
Since 1,25(OH)2D3 treatment had no profound impact on TXNIP mRNA levels at time points in which protein levels were found to be clearly reduced by treatment (12, 24, and 72 h), we speculated that 1,25(OH)2D3 reduces TXNIP levels through inducing its proteasomal degradation. Numerous studies have demonstrated that 1,25(OH)2D3 influences protein stability through a number of distinct mechanisms, namely regulating the expression of players in the ubiquitin proteasomal pathway, proteases, and protease inhibitors [45]. Moreover, TXNIP degradation through the ubiquitin proteasomal pathway has been demonstrated by studies implicating the E3 ubiquitin ligase ITCH as well as AMPK signaling in TXNIP stability [38, 46].
To investigate the possible degradation of TXNIP by 1,25(OH)2D3, the proteasomal and lysosomal inhibitors MG-132 and leupeptin, respectively, were added to the conditioned medium of DMSO- and 1,25(OH)2D3-treated MCF-7 cells 6 h prior to the end of the treatment period (72 h). A similar treatment plan was employed for 2-deoxyglucose, which was used as a positive control of TXNIP regulation, since this molecule is phosphorylated by hexokinase to yield 2-deoxyglucose-6-phosphate, which accumulates intracellularly, thus inducing the nuclear translocation of MondoA/Mlx leading to an increase in TXNIP expression [37]. MG-132 and 2-deoxyglucose, but not leupeptin, were found to rescue TXNIP levels in 1,25(OH)2D3-treated cells (Fig. 5a).

1,25(OH)2D3 induces proteasomal degradation of TXNIP in MCF-7 cells. a The reduction in TXNIP protein levels by 1,25(OH)2D3 (100 nM) in MCF-7 cells is rescued by MG-132 (5 μM) or 2-deoxyglucose (10 mM), but not leupeptin (20 μM). The various molecules were added to the conditioned medium of DMSO- and 1,25(OH)2D3-treated (66 h) MCF-7 cells, for an additional 6 h. b, c ITCH mRNA and protein expression is not markedly influenced by 1,25(OH)2D3. Relative expression was calculated using the ∆∆Ct method with vinculin as the housekee** gene. Error bars ± SD; n > 3. d Overall protein ubiquitination in MCF-7 cells was not changed by 1,25(OH)2D3 treatment. e Co-immunoprecipitation studies illustrate that the TXNIP-ITCH interaction is not altered by 1,25(OH)2D3 treatment of MCF-7 cells. f Negative regulation of TXNIP protein expression by 1,25(OH)2D3 is observed in MCF-7 cells with knocked-down AMPKα1 levels. g The non-calcemic 1,25(OH)2D3 analogue, calcipotriol (100 nM; 72 h) induces similar effects on TXNIP expression as 1,25(OH)2D3. The cell permeable Ca2+ chelator BAPTA-AM (20 μM) does not hamper 1,25(OH)2D3’s effects on TXNIP expression. BAPTA-AM was added to the conditioned medium of DMSO- and 1,25(OH)2D3-treated MCF-7 cells, 2 h prior to the end of the treatment period (72 h)
In view of this, we proposed that 1,25(OH)2D3 may influence TXNIP proteasomal degradation by either inducing ITCH expression or by enhancing its interaction with TXNIP. Assessment of changes in ITCH mRNA and protein levels in MCF-7 cells in response to 1,25(OH)2D3 did not reveal an induction in ITCH expression by treatment (Fig. 5b, c). Similarly, overall protein ubiquitination in MCF-7 cells was unchanged by 1,25(OH)2D3 treatment (Fig. 5d). Furthermore, 1,25(OH)2D3 was not found to markedly alter the levels of ITCH co-precipitating with TXNIP (Fig. 5e). We then hypothesized that AMPK, which was found to be activated in response to 1,25(OH)2D3 in MCF-7 cells (Fig. 3a), may be implicated in TXNIP degradation by 1,25(OH)2D3. We therefore knocked-down AMPK⍺1 levels in MCF-7 cells and investigated TXNIP protein expression in response to a 24- and 72-h treatment with 1,25(OH)2D3. Interestingly, the reduction in TXNIP expression by 1,25(OH)2D3 was observed in cells with knocked-down AMPK⍺1 levels (Fig. 5f).
Additionally, we postulated that TXNIP degradation by 1,25(OH)2D3 may be Ca2+-dependent, for example, by increasing intracellular Ca2+ levels leading to AMPK activation or by activating Ca2+-dependent proteases, namely calpains. To this end, we employed calcipotriol, a hypo-calcemic analog of 1,25(OH)2D3, as well as the cell permeable Ca2+ chelator BAPTA-AM. Similar to the parental molecule, calcipotriol treatment led to a reduction in TXNIP levels in MCF-7 cells (Fig. 5g). Moreover, addition of BAPTA-AM to the conditioned medium of 1,25(OH)2D3-treated MCF-7 cells did not rescue TXNIP levels (Fig. 5g). These data indicate the lack of involvement of Ca2+ signaling in mediating 1,25(OH)2D3’s effects on TXNIP expression.
Negative regulation of TXNIP protein levels by 1,25(OH)2D3 is possibly ER dependent
Previous reports have highlighted the ability of 1,25(OH)2D3 to decrease the expression of ER⍺ in BCa cells [47, 48]. Since estradiol stimulates glucose uptake and metabolism in BCa cells [49, 50], we investigated the possible involvement of ER in 1,25(OH)2D3’s regulation of TXNIP. Firstly, we sought to confirm the reduction in ER⍺ mRNA and protein expression in MCF-7 cells by 1,25(OH)2D3. Indeed, treatment was found to reduce ER⍺ expression in MCF-7 cells (Fig. 6a–c). We then co-treated cells with ER signaling regulators, namely the selective ER modulator tamoxifen, and estradiol, in the presence of either DMSO or 1,25(OH)2D3. Tamoxifen, independent of 1,25(OH)2D3, was found to reduce TXNIP protein levels in a time-dependent manner (Fig. 6d). Estradiol on the other hand did not markedly influence TXNIP protein levels in either DMSO- or 1,25(OH)2D3-treated cells (Fig. 6e). On the mRNA level, both ER agonist and antagonist did not significantly alter TXNIP expression, independent of 1,25(OH)2D3 (Fig. 6f).

Reduction of TXNIP expression in MCF-7 cells by 1,25(OH)2D3 is possibly ER-dependent. a–c 1,25(OH)2D3 (100 nM) treatment significantly reduces ER⍺ mRNA (72 h) and protein expression in MCF-7 cells. Relative expression was calculated using the ∆∆Ct method, with vinculin as the housekee** gene. Statistical significance between DMSO- and 1,25(OH)2D3-treated cells is calculated using a two-tailed Student’s t test, where p values less than or equal to 0.05, 0.01, and 0.001 are depicted in the figures by *, **, and ***, respectively. Error bars ± SD; n = 3. Tamoxifen (10 μM) treatment (d), but not estradiol (100 nM) (e), reduces TXNIP protein expression in a time-dependent manner. f Non-significant regulation of TXNIP mRNA levels is observed in MCF-7 cells treated for 24 h with either tamoxifen or estradiol, alone and in combination with 1,25(OH)2D3. g Glucose uptake in MCF-7 cells is significantly reduced by various concentrations of tamoxifen (24 h), an effect that is significantly ablated in the presence of 1,25(OH)2D3. Estradiol reduces glucose uptake in a concentration-dependent manner, with 1,25(OH)2D3 only influencing regulation of glucose uptake in response to 10 nM treatment with estradiol
Given TXNIP’s role in glucose homeostasis, and since GLUT1 expression has been shown to be induced by estradiol treatment [51], we aimed to study the influence of co-treating MCF-7 cells with ER modulators and either DMSO or 1,25(OH)2D3 on glucose uptake. While different concentrations of tamoxifen markedly reduced glucose uptake, 1,25(OH)2D3 co-treatment was found to significantly attenuate this effect (Fig. 6g). Estradiol treatment on the other hand led to a concentration-dependent reduction in glucose uptake, an effect that was not largely impacted by 1,25(OH)2D3 co-treatment (Fig. 6g).
