
Abstract
Background
Monogenea van Beneden, 1858 is a group of parasitic flatworms, commonly found infecting bony fish. Several genera, such as Cichlidogyrus Paperna, 1960, are reported to include potential pathogenic species that can negatively impact aquaculture fish stocks. They can switch from introduced to native fish and vice versa. In Africa (and all over the world), fish species belonging to Cichlidae are often kept in aquaculture and represent a major source of food. Thus, research on the biodiversity and occurrence of monogenean species on these fish is of importance for aquaculture and conservation. The present study is a survey of the diversity of species of Cichlidogyrus in the south of the Democratic Republic of the Congo (DRC) on three cichlid species: Orthochromis sp. ‘Lomami’, Serranochromis cf. macrocephalus, and Tilapia sparrmanii Smith, 1840.
Methods
Specimens of Cichlidogyrus were isolated from the gills and mounted on glass slides with Hoyer’s medium. The genital and haptoral hard parts were measured and drawn using interference contrast.
Results
In total, six species of Cichlidogyrus were found, all new to science: C. bulbophallus n. sp. and C. pseudozambezensis n. sp. on S. cf. macrocephalus, C. flagellum n. sp. and C. lobus n. sp. on T. sparrmanii, C. ranula n. sp. on S. cf. macrocephalus and Orthochromis sp. ‘Lomami’, and C. maeander n. sp. found on Orthochromis sp. ‘Lomami’ and T. sparrmanii. The first four species are considered to be strict specialists, C. ranula n. sp. an intermediate generalist and C. maeander n. sp. a generalist. These parasite species show morphological similarities to species found in the Lower Guinea and Zambezi ichthyofaunal provinces, which might be explained by past river capture events between river systems of the Congo Province and both these regions.
Conclusions
Serranochromis cf. macrocephalus and Orthochromis sp. ‘Lomami’ can harbour respectively three and two species of Cichlidogyrus, all described in this study. Tilapia sparrmanii can harbour seven species, of which three are described in the present study. These results highlight the species diversity of this parasite genus in the Congo Basin.
Similar content being viewed by others


Background
Since the second half of the 20th century, the number of described monogenean species increased substantially, as interest in fish parasites increased in general because of intensified aquaculture and stocking [1, 2]. At the moment, more than 5500 species and 750 genera have been described [3, 4]. Within the Monogenea, the Gyrodactylidae van Beneden & Hesse, 1863 and the Dactylogyridae Bychowsky, 1933 are the most species-rich families, each consisting of more than 500 to thousands of species described [2, 3].
Several genera of the Monogenea, such as Cichlidogyrus Paperna, 1960, Dactylogyrus Diesing, 1850 and Gyrodactylus von Nordmann, 1832, are reported to include potential fish pathogens, especially in aquaculture stocks [3,4,5]. There is a possibility that they switch from introduced to native fish, where they can cause high mortality (see [5,6,7] and references herein). Additionally, spillback phenomena from native to introduced fish have been observed [8]. Thus, research on the biodiversity and occurrence of monogenean species is of importance for aquaculture and conservation, especially in a time of intense aquaculture and worldwide translocation of their hosts.
In Africa (and all over the world), fish species belonging to the Cichlidae are often kept in aquaculture and represent a major source of food [9]. Among the non-ostariophysan groups, the Cichlidae is the most species-rich family, with about 1700 described species belonging to 250 genera. More than 1100 of these species occur in the inland waters of Africa [10,11,12]. At present, African cichlid fishes are known to harbour six genera of Monogenea: two of them are mesoparasites living in the host’s body cavity and four are ectoparasites found on the gills of their host [13]. The gill parasites include three genera belonging to the Dactylogyridae (Cichlidogyrus [14], Onchobdella Paperna, 1968 [15] and Scutogyrus Pariselle & Euzet, 1995 [16]) and one genus belonging to the Gyrodactylidae (Gyrodactylus) [17]. Among these, Cichlidogyrus is the most species-rich, including 125 described species [18,19,20]. Species of this genus naturally occur on cichlids from Africa and the Middle East [21, 22].
The present study is a survey of the diversity of species of Cichlidogyrus on cichlid species in the south of the Democratic Republic of the Congo (DRC). The diversity of this parasite genus is investigated for the fish hosts Orthochromis sp. ‘Lomami’ and Serranochromis cf. macrocephalus from the upper course of the Lomami River, and Tilapia sparrmanii Smith, 1840 from the Ngulungu River, an affluent of the upper course of the Lomami River (DRC). These three fish species belong to the so-called haplotilapiine lineage [23,24,25]. Tilapia sparrmanii belongs to the tribe Tilapiini which, together with the tribe Steatocranini, form the sister taxon to the group comprising the East African radiations. The latter contains Orthochromis Greenwood and Serranochromis Regan. Orthochromis is a polyphyletic genus consisting of haplochromine cichlid species inhabiting exclusively riverine habitats [23]. One monophyletic taxon occurs in the Malagarasi River system (suggested by Salzburger et al. [23] to be named Schwetzochromis), the other taxon in the Congo River system. The taxon occurring in the Congo River system falls in a cluster consisting of two main monophyletic clades: one clade including species of Orthochromis, the other including Serranochromis [23].
The Lomami River is a left bank affluent of the Middle Congo, running through the Upper Congo ecoregion [26]. For most of its course it runs parallel to the Upper Congo (Lualaba), joining the Middle Congo downstream from Kisangani, near Yangambi. The species richness of fish in the Congo Basin is largest in the main courses of the major rivers [27]. However, a relatively low number of fish species are reported from the Lomami River, which is most likely an indication of limited inventorying in this river [27]. By consequence, the knowledge on the diversity of monogenean fish parasites in this river is also non-existent, despite the aforementioned conservational and economic importance of their cichlid hosts [2]. Therefore, additional sampling in this river is needed to better understand the true diversity of fish and their parasites [27].
To date, only few studies have reported gill parasites of Serranochromis macrocephalus (Boulenger), T. sparrmanii or species of Orthochromis. Five species of Cichlidogyrus have been reported from S. macrocephalus: C. dossoui Douëllou, 1993 [28]; C. halli (Price & Kirk, 1967) [28]; C. quaestio Douëllou, 1993 [28]; C. sclerosus Paperna & Thurston, 1969 [28]; and C. zambezensis Douëllou, 1993 [28, 29]. Four species of Cichlidogyrus has been reported from T. sparrmanii (C. dossoui [30]; C. papernastrema Price, Peebles & Bamford, 1969 [29, 31]; C. quaestio [30]; and C. tiberianus Paperna, 1960 [30]) and only one species of Cichlidogyrus has been reported from a representative of Orthochromis (C. consobrini Jorissen, Pariselle & Vanhove 2018 [29]). However, this is likely an underestimation of the true species richness of this genus on these hosts. Indeed, taxonomic reports of Cichlidogyrus often indicate that the diversity on a single host can reach high levels, e.g. 25 species of Cichlidogyrus have been reported from the host Coptodon guineensis (Günther) [18].
Methods
Collection, sample preparation and conservation
Fish were bought from local fishermen from the Upper Lomami Basin in the Haut-Lomami province (DRC), during a field expedition in May 2017. Specimens of Serranochromis cf. macrocephalus and Orthochromis sp. ‘Lomami’ were caught in the main stream (8°33′36″S, 24°36′36″E) on 29–30 May 2017. Specimens of Tilapia sparrmanii were captured from the Ngulungu River (8°44′09″S, 24°43′58″E), an affluent of the Lomami River, on 22 May 2017 (Fig. 1). The specimens of Orthochromis sp. ‘Lomami’ were caught using dip nets in the rapids of the river. Specimens of Serranochromis cf. macrocephalus were caught using seine nets. Specimens of Tilapia sparrmanii were caught using a combination of dip nets and fish pots. Fish were fixed in the field with formaldehyde (10%). Gills of the right gill chamber were dissected in the laboratory and screened exhaustively for monogenean parasites using an entomological needle under an Optica 4.0.0 stereomicroscope. Parasites were mounted on glass slides under a coverslip, directly in a drop of Hoyer’s medium. Coverslips were sealed with nailpolish.

Map of the DRC situated on the African continent (left) and sampling region (right). Rivers in grey with in black the main streams (left) and Lomami Basin (right). Sampling locations are indicated as red dots (shapefiles downloaded from the Digital Chart of the World database using DIVA-GIS, maps created using QGis 2.18.14 software)
Fish were deposited in the ichthyology collection at the Royal Museum of Central Africa in Tervuren (Belgium) and stored in 70% ethanol. Specimens of Serranochromis cf. macrocephalus are stored under the collection numbers RMCA_Vert_2017.018.P.0019–0021. Specimens of Tilapia sparrmanii are stored under the collection numbers RMCA_Vert_2017.018.P.0022–0029. Specimens of Orthochromis sp. ‘Lomami’ are stored under the collection numbers RMCA_Vert_2017.018.P.0001 and RMCA_Vert_2017.018.P.0004–0005. Mounted parasite specimens were deposited in the invertebrate collection of the Royal Museum of Central Africa, Tervuren, Belgium (RMCA); the collection of the research group Zoology: Biodiversity and Toxicology at Hasselt University, Diepenbeek, Belgium (HU); the Finnish Museum of Natural History, Helsinki, Finland (MZH); and the Iziko South African Museum, Cape Town, Republic of South Africa (SAMC) (see “Type-material” for details on repositories and accession numbers).
Microscopy and illustrations
As dactylogyrid monogeneans are often differentiated based on their hard parts [21] and because soft internal organs were no longer visible in the whole-mounted specimens after fixation, the species descriptions focus on the details of the sclerotised parts, i.e. haptor, male copulatory complex (MCC) and vagina (when sclerotised). These hard parts were studied on the whole-mounted specimens with a Nikon Eclipse 80i compound microscope using interference contrast. Measurements were based on those of Pariselle & Euzet [32]. Additional measurements were taken for the uncinuli and the MCC (Table 1, Fig. 2). In total, 42 different metrical features were measured. For each species, the average, the range and number of measured specimens are provided. Measurements and drawings were made with the aid of a drawing tube at a magnification of 1000× (objective ×100 immersion, ocular ×10). Drawings were made freehand and edited in Adobe Illustrator CS5.1. Drawings are from multiple specimens in case not all structures were clearly visible in one single specimen. Micrographs were also taken with the Nikon Eclipse 80i and processed using NIS-Elements. Filaments associated with anchors and uncinuli are not presented as they are not diagnostically informative.

Measurements used in the description of the new species of Cichlidogyrus (measurements on the MCC and vagina are based on those of C. bulbophallus n. sp.). Abbreviations: A, anchors (a, anchor total length; b, anchor blade length; c, anchor shaft length; d, anchor guard length; e, anchor point length); U, uncinuli (U, total axial length; Us, axial length of secondary shaft; Uw, maximum width of first uncinuli); VB, ventral transverse bar (w, ventral bar maximum width; x, axial length of one branch); DB, dorsal transverse bar (h, axial length of the dorsal bar auricle; w, dorsal bar maximum width; y, distance between auricles; x, dorsal bar total length); MCC, male copulatory complex (Stl, axial length of penis stylet; Stw, maximum width of the penis stylet; Apl, axial length of accessory piece; Ap, linear length of the accessory piece; Apw, maximum width of the accessory piece; He, partial length of the heel; Hel, total length of the heel; Hew, width of the heel); VG, vagina (Vgl, total axial length; Vgw, maximum width), BODY, body
In literature, several terms are used to indicate specific structures of the posterior adhesive organ of representatives of the Dactylogyridae: the posterior adhesive organ is called opisthhaptor or simply haptor, the large hooks are called anchors or hamuli, small marginal hooks are indicated by the term hooklets or uncinuli, and the crosspieces connecting the large hooks are called the V-shaped/ventral and compound/dorsal supporting/connective/transverse bars [3, 14, 16, 28, 33]. Throughout this paper, we follow the terminology proposed by Fannes et al. [34]. Uncinuli are numbered as in Pariselle & Euzet [32]. We refer to the penis or copulatory tube as penis stylet as it essentially consists of a sclerotised hard structure [33]. Species were identified to genus level following Paperna [14] and the identification key of Pariselle & Euzet [21]. Different species of Cichlidogyrus were first assigned to one of the four morphological groups according to the relative size of their haptoral sclerites [21]: one group having long uncinuli I and long uncinuli III to VII, a second group consisting of species with short uncinuli I and long uncinuli III to VII, a third group consisting of species with short uncinuli I and short uncinuli III to VII, and finally a fourth group including species with long uncinuli I and short uncinuli III to VII. Uncinuli were called long or short based on their standardised length i.e. the division of their total length by the total length of the second uncinuli, which retain their larval size [21].
For the identification of new species, the phylogenetic species concept was adopted which states that species are reproductively isolated groups of natural populations that originate through a speciation event and end with the next speciation or vanish through extinction [35]. Practically, this means that a group of specimens is defined as a new species when they consistently differ from another group of specimens in at least one attribute [36]. Note that the authors of the new species are different from the authors of this article (International Commission on Zoological Nomenclature 2015 [37]).
Museum specimens from the invertebrate collection of the Royal Museum of Central Africa, Tervuren, Belgium, were used for morphological comparison with the new species (voucher specimens of C. zambezensis: MT.37714, MT.38165–MT.38171, MT.37984–MT.37986, MT.37988–MT.37992; paratypes of C. reversati Pariselle & Euzet 2003: MT.37400–MT.37402; and a paratype of C. legendrei Pariselle & Euzet 2003: MT.37400).
To comply with the regulations set out in article 8.5 of the amended 2012 version of the International Code of Zoological Nomenclature (ICZN) [38], details of the new species have been submitted to ZooBank. The Life Science Identifier (LSID) of the article is urn:lsid:zoobank.org:pub:845B6B53-7ED0-4100-8051-CC8A47B3481B. For each new species, the Life Science Identifier (LSID) is reported in the taxonomic summary.
Results
Three specimens of S. cf. macrocephalus, three of Orthochromis sp. ‘Lomami’, and eight of T. sparrmanii were sampled. A total of 171 monogeneans were found on these cichlid species: 132 specimens on S. cf. macrocephalus, 30 specimens on T. sparrmanii and 9 specimens on Orthochromis sp. ‘Lomami’. These all belong to six species of Cichlidogyrus, which are all new to science.
Species descriptions are presented below. Measurements of the hard parts are provided in Table 2.
Family Dactylogyridae Bychowski, 1933
Genus Cichlidogyrus Paperna, 1960
Cichlidogyrus bulbophallus Geraerts & Muterezi Bukinga n. sp.
Type-host:Serranochromis cf. macrocephalus (Perciformes: Cichlidae).
Type-locality: Lomami River (8°33′36″S, 24°36′36″E), Democratic Republic of the Congo.
Type-material: The holotype (RMCA 39079) and 4 paratypes (RMCA 39071, 39073) are deposited in the invertebrate collection of the Royal Museum of Central Africa, Tervuren, Belgium (RMCA); 44 paratypes (HU nos 622, 629, 631–635, 637–640, 645, 650, 654, 655, 659, 660, 662, 663, 666, 670, 678, 679, 681–684, 690–704, 706, 708) are deposited in the collection of the research group Zoology: Biodiversity and Toxicology at Hasselt University, Diepenbeek, Belgium (HU); 8 paratypes (MZH KN.13824-KN.13827, KN.13829, KN.13830, KN.13834, KN.13835) are deposited in the Finnish Museum of Natural History, Helsinki, Finland (MZH); and 9 paratypes (SAMC A091369-A091371) are deposited in the Iziko South African Museum, Cape Town, Republic of South Africa (SAMC).
Site in host: Gills.
Prevalence and intensity: In 3 out of 3 hosts studied. All three specimens of S. cf. macrocephalus harboured specimens of C. bulbophallus n. sp. with an infection intensity of 3, 22 and 41, respectively.
ZooBank registration: The Life Science Identifier (LSID) for Cichlidogyrus bulbophallus Geraerts & Muterezi Bukinga n. sp. is urn:lsid:zoobank.org:act:6CEC158A-33DB-40B8-80B7-9907EABA291A.
Etymology: The species epithet is a combination of the Latin words bulbus (m) (= bulb) and phallus (m) (= penis). The name refers to the bulb-shaped swollen part of the penis stylet.
Description
[Based on 66 specimens; metrical data in Table 2; see Fig. 3.] Anchors 2 pairs. Ventral anchors large, with guard approximately 2 times as long as shaft. Dorsal anchors of about same total length as ventral anchors; guard and shaft pronounced, asymmetrical; guard approximately 3 times as long as shaft. Blades of both ventral and dorsal anchors arched. Ventral transverse bar V-shaped, with 2 branches with wing-shaped attachments along distal half. Dorsal transverse bar made up of thick midsection, tapering towards its extremities, and 2 pronounced auricles inserted at its dorsal face. Uncinuli 7 pairs; uncinuli I long; uncinuli III to VII long.
MCC consisting of long penis stylet, accessory piece and heel. Proximal (18–33 µm, n = 61) and distal (46–78 µm, n = 66) ends of stylet tubiform, middle part (46–65 µm, n = 66) forming prominent bulb-shaped enlargement; stylet tube with approximately same diameter at proximal and distal ends. Base of penis stylet lacking distinct swollen bulb, attached to pronounced leaf-like heel. Accessory piece shorter than stylet, with a cap at distal end, connected to base of stylet proximally. Vagina coiled, long and wide, with two turns.

Sclerotised structures of Cichlidogyrus bulbophallus n. sp. Drawings based on five specimens: the holotype RMCA 39079 for the MCC and vagina; paratype HU no 633 for the VA and DB; paratype HU no 694 for the VB; paratype HU no 634 for the DA; and paratype HU no 635 for the uncinuli. Abbreviations: I-VII, uncinuli; VA, ventral anchor; VB, ventral transverse bar; DA, dorsal anchor; DB, dorsal transverse bar; MCC, male copulatory complex with penis stylet in white, and accessory piece and heel in grey; VG, vagina. Scale-bar: 20 µm
Differential diagnosis
The specimens display all diagnostic features of Cichlidogyrus: (i) two pairs of anchors (one dorsal and one ventral), two transverse bars (ventral bar V-shaped, dorsal bar with two auricles); (ii) 14 uncinuli; (iii) a male copulatory complex consisting of a penis stylet and usually an accessory piece; and (iv) a vagina which may or may not be sclerotised [14, 21]. Cichlidogyrus bulbophallus n. sp. shows an unusual haptor configuration: it has long uncinuli I and long uncinuli III to VII. Such combination has up to now only been described for five species: C. arthracanthus Paperna, 1960 [14], C. inconsultans Birgi & Lambert, 1987 [39, 40], C. centesimus Vanhove, Volckaert & Pariselle, 2011 [41], C. calycinus Kusters, Jorissen, Pariselle & Vanhove, 2018 (in some specimens) [20] and C. habluetzeli Rahmouni, Vanhove & Šimková, 2018 [19]. Based on the identification key, Cichlidogyrus bulbophallus n. sp. is similar to C. inconsultans (reported from Polycentropsis abbreviata Boulenger in Cameroon) [39]), C. calycinus (reported from Hemichromis elongatus (Guichenot) in the DRC) and C. habluetzeli (reported from Cardiopharynx schoutedeni Poll and Cyphotilapia frontosa (Boulenger) in the DRC) in having a sclerotised vagina (the vagina of C. arthracanthus and C. centesimus is not sclerotised). Cichlidogyrus bulbophallus n. sp. can be distinguished from these species, and any other species of Cichlidogyrus, based on the typical shape of its penis stylet, starting and ending as a tube of approximately the same diameter and with a distinct and greatly developed swollen middle portion. Also, the morphology of the vagina, which is long and wide, making two turns, is unique within the genus. Contrary to that of other species of Cichlidogyrus, the penis stylet of C. bulbophallus n. sp. does not show a distinctly swollen bulb at the base. This combination of characters has not yet been observed within Cichlidogyrus. Therefore, C. bulbophallus n. sp. can easily be recognised as a new species.
Cichlidogyrus pseudozambezensis Geraerts & Muterezi Bukinga n. sp.
Type-host:Serranochromis cf. macrocephalus (Perciformes: Cichlidae).
Type-locality: Lomami River (8°33′36″S, 24°36′36″E), Democratic Republic of the Congo.
Type-material: The holotype (RMCA 39078) and 7 paratypes (RMCA 39072, 39074, 39078) are deposited in the invertebrate collection of the Royal Museum of Central Africa, Tervuren, Belgium (RMCA); 43 paratypes (HU nos 624–628, 630, 636, 642–644, 646–649, 651–653, 656–658, 661, 664, 667–669, 671–677, 680, 685–689, 705, 707, 709–712) are deposited in the collection of the research group Zoology: Biodiversity and Toxicology at Hasselt University, Diepenbeek, Belgium (HU); 6 paratypes (MZH KN.13828, KN.13831-KN.13833, KN.13836, KN.13837) are deposited in the Finnish Museum of Natural History, Helsinki, Finland (MZH); and 5 paratypes (SAMC A091372) are deposited in the Iziko South African Museum, Cape Town, Republic of South Africa (SAMC).
Site in host: Gills.
Prevalence and intensity: In 3 out of 3 hosts studied. All three specimens of S. cf. macrocephalus harboured specimens of C. pseudozambezensis n. sp. with an infection intensity of 2, 2 and 59, respectively.
ZooBank registration: The Life Science Identifier (LSID) for Cichlidogyrus pseudozambezensis Geraerts & Muterezi Bukinga n. sp. is urn:lsid:zoobank.org:act:586511EB-98A5-4125-B867-0C741E432653.
Etymology: The prefix pseudo- (Ancient Greek for ‘false, fake’) of the species epithet is used to emphasise the resemblance of the MCC to that of C. zambezensis.
Description
[Based on 63 specimens: metrical data in Table 2; see Fig. 4.] Anchors 2 pairs. Ventral anchors with guard slightly longer and more robust than shaft. Dorsal anchors of about same total length as ventral anchors; base more asymmetrical than that of ventral anchors; guard and shaft pronounced; guard approximately 2 times as long as shaft. Ventral transverse bar V-shaped with 2 branches with wing-shaped projections along distal half. Dorsal transverse bar made up of thick midsection, narrowing towards its extremities, and 2 pronounced auricles inserted at its dorsal face. Uncinuli 7 pairs; uncinuli I long, uncinuli IV to VII long, uncinuli III intermediate.
MCC consisting of penis stylet, accessory piece and heel. Penis stylet drop-shaped with pointed distal end and enlarged proximal basal bulb; midsection broadened. Base of penis stylet surrounded by irregular-shaped heel. Wall of penis stylet swollen at point where basal bulb attaches to heel. Accessory piece large and attached to base of stylet. Vagina C-shaped tube narrowing distally.

Sclerotised structures of Cichlidogyrus pseudozambezensis n. sp. Drawings are based on five specimens: the holotype RMCA 39078 for the vagina; paratype RMCA 39078 for the MCC; paratype HU no 627 for the uncinuli VA and DA; paratype HU no 651 for the DB; and paratype HU no 653 for the VB. Abbreviations: I-VII, uncinuli; VA, ventral anchor; VB, ventral transverse bar; DA, dorsal anchor; DB, dorsal transverse bar; MCC, male copulatory complex with penis stylet in white, and accessory piece and heel in grey; VG, vagina. Scale-bar: 20 µm
Differential diagnosis
The specimens display all diagnostic features of Cichlidogyrus (see “Differential diagnosis” for C. bulbophallus n. sp.). Cichlidogyrus pseudozambezensis n. sp. resembles C. arthracanthus, C. inconsultans, C. centesimus, C. calycinus and C. habluetzeli in having long uncinuli I and long uncinuli IV to VII [21, 22]. However, the MCC of C. pseudozambezensis n. sp. is morphologically similar to that of C. zambezensis (found on species of Serranochromis in Zimbabwe and the DRC, on Oreochromis mortimeri (Trewavas) in Zimbabwe [28], and on the Sargochromis mellandi (Boulenger) species complex in the DRC [29, 30]), a species that is positioned in a morphology-based group characterised by short uncinuli I and short uncinuli III to VII [21, 22]. Cichlidogyrus pseudozambezensis n. sp. also differs from C. zambezensis in the shape of the vagina, which is described as triangular, funnel-shaped or hat-like in C. zambezensis [28, 29]. Additionally, the accessory piece in C. zambezensis has a pointed distal end, in contrast to that in C. pseudozambezensis n. sp., which distally ends in a more rounded tip.
Cichlidogyrus flagellum Geraerts & Muterezi Bukinga n. sp.
Type-host:Tilapia sparrmanii Smith (Perciformes: Cichlidae).
Type-locality: Ngulungu River (8°44′09″S, 24°43′58″E), Democratic Republic of the Congo.
Type-material The holotype (RMCA 39075) is deposited in the invertebrate collection of the Royal Museum of Central Africa, Tervuren, Belgium (RMCA); 4 paratypes (H:U nos 615, 618, 619, 721) are deposited in the collection of the research group Zoology: Biodiversity and Toxicology at Hasselt University, Diepenbeek, Belgium (HU); 2 paratypes (MZH KN.13821, KN.13838) are deposited in the Finnish Museum of Natural History, Helsinki, Finland (MZH); and 1 paratype (SAMC A091368) is deposited in the Iziko South African Museum, Cape Town, Republic of South Africa (SAMC).
Site in host: Gills.
Prevalence and intensity: In 7 out of 8 hosts studied. Six specimens of T. sparrmanii harboured specimens of C. flagellum n. sp. with an infection intensity of 1, one with an infection intensity of 2.
ZooBank registration: The Life Science Identifier (LSID) for Cichlidogyrus flagellum Geraerts & Muterezi Bukinga n. sp. is urn:lsid:zoobank.org:act:8C8DF873-A2DA-4819-8DE1-05C955E1F625.
Etymology: The species epithet refers to the whip-like appearance of the penis stylet; flagellum (Lat., n) = whip.
Description
[Based on 8 specimens; metrical data in Table 2; see Fig. 5.] Anchors 2 pairs. Ventral anchors with guard approximately 2 times as long as shaft. Dorsal anchors somewhat shorter and less robust than ventral ones; guards, like those of ventral anchors, approximately 2 times as long as shafts. Ventral transverse bar V-shaped, with 2 branches with ridge along their length. Due to compression during preparation of microscope slides, ventral transverse bar not appearing V-shaped, but folded into other shapes. In paratype KN.13838 e.g., ventral transverse bar looking W-shaped (Fig. 5). Dorsal transverse bar more or less same width over its entire length; auricles short and inserted at dorsal face of bar. Uncinuli 7 pairs; uncinuli I short with short secondary shafts; uncinuli III and VII short; uncinuli IV, V, and VI long.
MCC consisting of penis stylet, accessory piece and heel. Penis stylet beginning as enlarged bulb, attached to small egg-shaped heel; penis stylet distinctly swollen at proximal third, proceeding distally as slender elongated whip-like tube, forming loop at distal end in all specimens. Accessory piece fish-hook-shaped, forming loop at beginning and joining penis stylet distally; large hook present at the end; accessory base proximally connected to base of penis stylet. Vagina sclerotised, folded and proximally broadened.

a Sclerotised structures of Cichlidogyrus flagellum n. sp. Drawings are based on four specimens: the holotype RMCA 39075 for uncinuli I, II, IV-VII and the vagina; paratype HU no 615 for uncinuli III; paratype MZH KN.13838 for the VB and MCC; and paratype HU no 721 for the VA, DA and DB. b, c Micrographs of the VB taken from paratype KN.13838 at two different focal depths. In (b), the two branches of the VB are indicated by an arrow. In (c), the middle part of the VB is indicated by an arrow. The VB looks W-shaped in the drawing (a) and micrographs (b, c) due to folding of this structure during the preparation of the microscope slide mount. Abbreviations: I-VII, uncinuli; VA, ventral anchor; VB, ventral transverse bar; DA, dorsal anchor; DB, dorsal transverse bar; MCC, male copulatory complex with penis stylet in white, and accessory piece and heel in grey; VG, vagina. Scale-bars: 20 µm
Differential diagnosis
The specimens display all diagnostic features of Cichlidogyrus (see “Differential diagnosis” for C. bulbophallus n. sp.). Cichlidogyrus flagellum n. sp. resembles C. douellouae Pariselle, Bilong Bilong & Euzet, 2003 (found on species of Sarotherodon Rüppell in West and West Central Africa [42]) in having short uncinuli I, long uncinuli IV to VI, a large vagina, a penis stylet length of more than 65 μm and an accessory piece ending in a hook. Clear differences between these two species can be found in the morphology of the reproductive organs. The vagina of C. flagellum n. sp. is longer (48–67 vs 12–17 µm in C. douellouae) and more curved than that of C. douellouae. The proximal third of the penis stylet of C. flagellum n. sp. is distinctly swollen and the penis stylet shows a loop near its distal end, in contrast to the penis stylet of C. douellouae, which does not have such swollen portion and loop. The accessory piece of C. flagellum n. sp. is fish-hook-shaped, while that of C. douellouae is S-shaped with a large and perpendicular diverticulum at its proximal third.
Cichlidogyrus maeander Geraerts & Muterezi Bukinga n. sp.
Type-host:Tilapia sparrmanii (Perciformes: Cichlidae).
Other host:Orthochromis sp. ‘Lomami’ (Perciformes: Cichlidae).
Type-locality: Ngulungu River (8°44′9″S, 24°43′58″E), the Democratic Republic of the Congo.
Other locality: Lomami River (8°33′36″S, 24°36′36″E) on Orthochromis sp. ‘Lomami’, the Democratic Republic of the Congo.
Type-material: The holotype (RMCA 39068) and 3 paratypes (RMCA 39068, 39077) are deposited in the Royal Museum of Central Africa, Tervuren, Belgium (RMCA); 7 paratypes (HU nos 614, 616, 620, 719, 722–724) are deposited in the collection of the research group Zoology: Biodiversity and Toxicology at Hasselt University, Diepenbeek, Belgium (HU); 3 paratypes (MZH KN.13822, KN.13840, KN.13841) are deposited in the Finnish Museum of Natural History, Helsinki, Finland (MZH); and 1 paratype (SAMC A091373) is deposited in the Iziko South African Museum, Cape Town, Republic of South Africa (SAMC).
Site in host: Gills.
Prevalence and intensity: In 6 out of 8 specimens of T. sparrmanii studied. Three specimens of T. sparrmanii were infected with an infection intensity of 3, two with an infection intensity of 2, and one with an infection intensity of 1. In 1 out of 3 specimens of Orthochromis sp. ‘Lomami’ studied. One specimen of Orthochromis sp. ‘Lomami’ was infected with an infection intensity of 1.
ZooBank registration: The Life Science Identifier (LSID) for Cichlidogyrus maeander Geraerts & Muterezi Bukinga n. sp. is urn:lsid:zoobank.org:act:23713587-C596-46F3-983B-0B58F69E0E64.
Etymology: The species epithet is deduced from Latin (maeander (m) = a road with a winding course) and refers to the sinusoid turning of the accessory piece around the penis stylet.
Description
[Based on 15 specimens; metrical data in Table 2; see Fig. 6.] Anchors 2 pairs. Ventral anchors with guard approximately 2 times as long as shaft. Dorsal anchors longer than ventral ones with longer shaft and guard; base consisting of pronounced pointed guard and blunt shaft, again with guard being approximately 2 times as long as shaft. Ventral transverse bar long and V-shaped with wing-like attachments along distal third of branches. Dorsal transverse bar arched with thick midsection, narrowing towards its extremities, with two pronounced auricles at its dorsal face. Uncinuli 7 pairs; uncinuli I long; uncinuli IV to VII long; uncinuli III of intermediate length.
MCC consisting of penis stylet, accessory piece and heel. Penis stylet short, forming enlarged bulb at base; base attached to pronounced heel; penis stylet distally curved, with pointed end. Accessory piece sinusoid and pointed at distal end; accessory piece narrowed proximally and attached to basal bulb of penis stylet. Vagina not observed.

Sclerotised structures of Cichlidogyrus maeander n. sp. Drawings are based on three specimens: the holotype RMCA 39068 for the DA and MCC; paratype HU no 723 for the uncinuli, DB and VA; and paratype HU no 614 for the VB. Abbreviations: I-VII, uncinuli; VA, ventral anchor; VB, ventral transverse bar; DA, dorsal anchor; DB, dorsal transverse bar; MCC, male copulatory complex with penis stylet in white, and accessory piece and heel in grey. Scale-bar: 20 µm
Differential diagnosis
The specimens display all diagnostic features of Cichlidogyrus (see “Differential diagnosis” for C. bulbophallus n. sp.). Cichlidogyrus maeander n. sp. resembles C. arthracanthus and C. centesimus; all three of them have long uncinuli I and long uncinuli IV to VII, and lack a sclerotised vagina [14, 39, 40]. Cichlidogyrus maeander n. sp. can, however, easily be distinguished from both based on the shape of the MCC. The morphology of the MCC resembles more that of C. reversati (found on Pelmatolapia cabrae (Boulenger) in the mouth of the Bas Kouilou River (DRC) [32]). However, the distal end of the penis stylet of C. maeander n. sp. is pointed, while it is wide-mouthed in C. reversati, and the accessory piece of C. maeander is more pointed than that of C. reversati. Additionally, C. maeander n. sp. differs from C. reversati in the length of uncinuli III-VII (C. reversati having short uncinuli III-VII) [21]. Also, uncinuli I are more robust compared to those of C. reversati.
Cichlidogyrus lobus Geraerts & Muterezi Bukinga n. sp.
Type-host:Tilapia sparrmanii (Perciformes: Cichlidae).
Type-locality: Ngulungu River (8°44′9″S, 24°43′58″E), the Democratic Republic of the Congo.
Type-material: The holotype (RMCA 39069) and 1 paratype (RMCA 39076) are deposited in the invertebrate collection of the Royal Museum of Central Africa, Tervuren, Belgium (RMCA); 1 paratype (HU no 617) is deposited in the collection of the research group Zoology: Biodiversity and Toxicology at Hasselt University, Diepenbeek, Belgium (HU); 1 paratype (MZH KN.13839) is deposited in the Finnish Museum of Natural History, Helsinki, Finland (MZH); and 1 paratype (SAMC A091373) is deposited in the Iziko South African Museum, Cape Town, Republic of South Africa (SAMC).
Site in host: Gills.
Prevalence and intensity: In 4 out of 8 hosts studied. One specimen of T. sparrmanii was infected with an infection intensity of 2 and three specimens were infected with an infection intensity of 1.
ZooBank registration: The Life Science Identifier (LSID) for Cichlidogyrus lobus Geraerts & Muterezi Bukinga n. sp. is urn:lsid:zoobank.org:act:E6AB8EA6-1ED8-4972-8EBA-5FD5FAC371F7.
Etymology: The species epithet refers to the lobed accessory piece; lobus (Lat.; m) = lobe.
Description
[Based on 5 specimens; metrical data in Table 2; see Fig. 7.] Anchors 2 pairs. Ventral anchors with robust base; guard approximately 2 times as long as shaft. Dorsal anchors approximately as long as ventral anchors, but thinner; base more symmetrical than that of ventral anchor; guard slightly longer than shaft; point slender and less curved compared to that of ventral one. Ventral transverse bar V-shaped with two robust branches with rough ridge over their entire length. Dorsal transverse bar made up of thick midsection, tapering towards its extremities, and 2 pronounced auricles inserted at its dorsal face. Uncinuli 7 pairs; uncinuli I short; uncinuli IV short; uncinuli III, V, VI, and VII long.
MCC consisting of penis stylet, accessory piece and heel. Penis stylet hook-shaped and pointed at distal end; base enlarged to ovoid basal bulb and attached to large heel. Penis stylet seeming to have swollen portion distally from basal bulb in holotype (Fig. 7), being merely an artefact due to folding of basal bulb during preparation of microscope slide mount. Accessory piece complex, consisting of several lobes; accessory piece proximally attached to basal bulb. Vagina short, thick-walled, and slightly arched in some specimens.

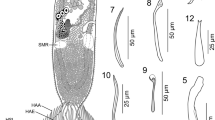
a Sclerotised structures of Cichlidogyrus lobus n. sp. Drawings are based on four specimens: the holotype RMCA 39069 for the MCC; paratype HU no 617 for the uncinuli and vagina; paratype SAMC A091373 for the VB and DB; and paratype MZH KN.13839 for the VA and DA. b–e Micrographs of the MCC of two specimens of C. lobus n. sp. b, c Micrographs taken from the holotype at two different focal depths. d, e Micrographs taken from paratype KN.13839 at two different focal depths. The arrows point at the basal bulb of the penis stylet. The penis stylet of the holotype seems to have a swollen portion distally from the basal bulb. This seemingly swollen portion is merely an artefact due to the folding of the basal bulb during preparation of the microscope slide mount. The basal bulb of paratype KN.13839 is not folded and shows that the penis stylet does not possess a swollen portion. Abbreviations: I-VII, uncinuli; VA, ventral anchor; VB, ventral transverse bar; DA, dorsal anchor; DB, dorsal transverse bar; MCC, male copulatory complex with penis stylet in white, and accessory piece and heel in grey; VG, vagina. Scale-bars: 20 µm
Differential diagnosis
The specimens display all diagnostic features of Cichlidogyrus (see “Differential diagnosis” for C. bulbophallus n. sp.). Cichlidogyrus lobus n. sp. resembles C. legendrei (found on Pelmatolapia cabrae in Lake Cayo (DRC) [32]). They can be distinguished from each other based on several details in the morphology of the haptor and MCC. Uncinuli IV of C. lobus n. sp. are short, while those of C. legendrei are long. The ventral transverse bar of C. legendrei has a slender appearance with two distinct wing-shaped appendages at the distal end of the branches. In contrast, the ventral transverse bar of C. lobus n. sp. is more robust with a rough ridge along the length of the branches; the wing-shaped appendages are absent. The penis stylet of C. lobus n. sp. is hook-shaped, in contrast to the sinuous penis stylet of C. legendrei. The pronounced hook at the end of the accessory piece of C. legendrei is not present in C. lobus n. sp.
Cichlidogyrus ranula Geraerts & Muterezi Bukinga n. sp.
Type-host:Orthochromis sp. ‘Lomami’ (Perciformes: Cichlidae).
Other host:Serranochromis cf. macrocephalus (Perciformes: Cichlidae).
Type-locality: Lomami River (8°33′36″S, 24°36′36″E), the Democratic Republic of the Congo.
Type-material: The holotype (RMCA 39070) and 1 paratype (RMCA 39070) are deposited in the invertebrate collection of the Royal Museum of Central Africa, Tervuren, Belgium (RMCA); and 1 paratype (HU no 641) is deposited in the collection of the research group Zoology: Biodiversity and Toxicology at Hasselt University, Diepenbeek, Belgium (HU).
Site in host: Gills.
Prevalence and intensity: In 1 out of 3 specimens of Orthochromis sp. ‘Lomami’ studied; this specimen was infected with an infection intensity of 2. In 1 out of 3 specimens of S. cf. macrocephalus studied; this specimen was infected with an infection intensity of 1.
ZooBank registration: The Life Science Identifier (LSID) for Cichlidogyrus ranula Geraerts & Muterezi Bukinga n. sp. is urn:lsid:zoobank.org:act:19CF0F3F-9971-4983-A637-87CBB303E0DE.
Etymology: The species epithet refers to the tadpole-shaped penis stylet; ranula (Lat., f) = tadpole.
Description
[Based on 3 specimens; metrical data in Table 2; see Fig. 8.] Anchors 2 pairs. Ventral anchors with robust asymmetric base; long guard approximately 2 times as long as shaft. Dorsal anchors slightly shorter than ventral ones; guard also approximately 2 times as long as shaft. Ventral transverse bar V-shaped with two long robust branches with wing-shaped attachments along distal third of each branch. Dorsal transverse bar made up of typically thick midsection, tapering towards its extremities, and two pronounced auricles at its dorsal face. Uncinuli 7 pairs; uncinuli I short with short secondary shafts; uncinuli III to VII long.
MCC consisting of penis stylet, accessory piece and heel. Penis stylet tadpole-shaped having swollen midsection and tapered, curved distal end; base enlarged to ovoid basal bulb and attached to long heel. Accessory piece extending from heel and having sinusoid course, ending distally in small hook. Vagina tube-shaped and thick-walled.

Sclerotised structures of Cichlidogyrus ranula n. sp. Drawings are based on three specimens: the holotype RMCA 39070 for the VA and MCC; paratype RMCA 39070 for the VB, DB, DA, uncinuli I and vagina; and paratype HU no 641 for uncinuli II-VII. Abbreviations: I-VII, uncinuli; VA, ventral anchor; VB, ventral transverse bar; DA, dorsal anchor; DB, dorsal transverse bar; MCC, male copulatory complex with penis stylet in white, and accessory piece and heel in grey; VG, vagina. Scale-bar: 20 µm
Differential diagnosis
The specimens display all diagnostic features of Cichlidogyrus (see “Differential diagnosis” for C. bulbophallus n. sp.). C. ranula n. sp. falls in the morphological group of species of Cichlidogyrus with short uncinuli I and long uncinuli III to VII. This species resembles C. legendrei and C. lobus n. sp. in having a short, rather straight, tubular vagina with a smooth wall. Clear differences with these two species are visible in the shape of the penis stylet and accessory piece: C. legendrei and C. lobus n. sp. have a penis stylet which lacks a broadened midsection (vs a broadened midsection in C. ranula n. sp.) and have a complex accessory piece (vs a simple accessory piece with a small hook at the end in C. ranula n. sp.). The MCC of C. ranula n. sp. shows some resemblance to that of C. papernastrema (found on Tilapia sparrmanii in South Africa and the DRC [29, 31]), which also has a penis stylet with a broadened midsection, tapering and curving towards the end, and an accessory piece ending in a hook. However, C. papernastrema lacks a sclerotised vagina and falls in a group of species showing a different haptoral construction, with long uncinuli I and short unicinuli III to VII.
Remarks
Juveniles were identified by having a relatively large haptor compared to their body size, and no or not fully sclerotised copulatory organs. In total, twelve specimens of Cichlidogyrus were considered to be juveniles. In eight of these juveniles the copulatory organs were not yet developed: five of them occurring on Orthochromis sp. ‘Lomami’, the other three on Tilapia sparrmanii. The other four juveniles already possessed an MCC, though not yet fully sclerotised. These juveniles were not considered in the species descriptions and the calculation of the infection intensities.
Discussion
Monogenean species richness on the cichlid hosts
This present morphological survey revealed the presence of six new species of Cichlidogyrus: C. bulbophallus n. sp. and C. pseudozambezensis n. sp. found on Serranochromis cf. macrocephalus; C. flagellum n. sp. and C. lobus n. sp. on Tilapia sparrmanii; C. ranula n. sp. occurring on Serranochromis cf. macrocephalus and Orthochromis sp. ‘Lomami’; and C. maeander n. sp. found on Orthochromis sp. ‘Lomami’ and Tilapia sparrmanii.
Four species of Cichlidogyrus have previously been described from T. sparrmanii: C. dossoui [30]; C. papernastrema [29, 31]; C. quaestio [30]; and C. tiberianus [30]. Cichlidogyrus papernastrema, of which the MCC shows some resemblance with that of C. ranula n. sp., was described from the Zambezian Lowveld ecoregion (Natal, South Africa) [26, 31] and the Bangweulu-Mweru ecoregion (DRC) [26, 29, 31]. In the latter region, this species was also found on Coptodon rendalli (Boulenger) and Oreochromis mweruensis Trewavas.
Five species of Cichlidogyrus have been reported from Serranochromis macrocephalus: C. dossoui [28]; C. halli [28]; C. quaestio [28]; C. sclerosus [28]; and C. zambezensis [28, 29]. Cichlidogyrus zambezensis, of which the MCC shows some resemblance with that of C. pseudozambezensis n. sp., was originally described from the host S. macrocephalus from the Kariba Lake in Zimbabwe [28], where this species was also found to infect Oreochromis mortimeri [28]. In the Bangwuelu-Mweru ecoregion, Cichlidogyrus zambezensis was also found on Serranochromis robustus jallae (Günther), S. stappersi Trewavas, 1964, S. angusticeps (Boulenger), S. thumbergi (Castelnau) and the Sargochromis mellandi species complex [29, 30].
The only species of Cichlidogyrus described from Orthochromis spp. is C. consobrini on Orthochromis katumbii Schedel, Vreven, Manda, Abwe, Manda & Schliewen, 2018 from the Kiswishi River (DRC), where it was also found to infect the Sargochromis mellandi species complex [29, 43].
To sum up, after this study Serranochromis cf. macrocephalus (assuming it is a new fish species) and Orthochromis sp. ‘Lomami’ are known to harbour respectively three and two species of Cichlidogyrus, all described in this study. Tilapia sparrmanii is known to harbour seven species, of which three are described in the present study. The discovery of new species of Cichlidogyrus is not surprising, given the high species richness of this genus and the limited amount of studies reporting gill parasites from these hosts [21, 43].
Morphological comparison with species from the Lower Guinea and Zambezi ichthyofaunal provinces
Cichlidogyrus bulbophallus n. sp., C. pseudozambezensis n. sp., C. flagellum n. sp. and C. ranula n. sp. all have a penis stylet with a swollen portion. Other congeners with such a distinct swollen portion are C. amphoratus Pariselle & Euzet, 1996 [44], C. giostrai Pariselle, Bilong Bilong & Euzet, 2003 [42], C. n**ei Pariselle, Bilong Bilong & Euzet, 2003 [42], C. ornatus Pariselle & Euzet, 1996 [44], C. papernastrema [29, 31], C. sanjeani Pariselle & Euzet, 1997 [45] and C. zambezensis [28, 29]. These species are mainly reported from ichthyofaunal provinces in West Africa and West Central Africa (Upper Guinea province and Lower Guinea province) and Central Africa (Congo province and Zambezi province), all occurring on cichlids from the haplotilapiine lineage (Table 3) [12, 23, 24].
Some species described in this study also showed morphological similarities to species reported from the ichthyofaunal province of Lower Guinea (C. legendrei and C. reversati) and the Upper Congo-Zambezi province (C. zambezensis and C. papernastrema) (see above and Table 3).
The Lomami River belongs to the Congo ichthyofaunal province. The Lower Guinea and the Congo ichthyofaunal province share part of their fish fauna, possibly as a consequence of the historic capture of some Lower Congo rivers by rivers in the Lower Guinea province [12]. Similarly, the Congo province and the Zambezi province show ichthyofaunal similarities, probably due to the past connection between the Upper Zambezi and the Kasai Basin and between the Upper Zambezi and the Upper Congo [12, 27, 46]. The morphological similarities of the MCC of species of Cichlidogyrus from the Lomami River Basin to those of the Lower Guinea and Upper Congo-Zambezi could be explained by convergent evolution, or alternatively be explained by these past river connections, which would have allowed the colonisation of Lower Guinean and Zambezian fish fauna into the Congo province. It would be worthwhile to use genetic data to investigate whether these morphological similarities are due to convergent evolution, typical to the monogenean fauna of certain ichthyofaunal provinces or host species, or whether this occurrence pattern is rather an artefact due to a sampling bias towards these regions and cichlid lineage.
Host specificity
Adopting the delimitation of host specificity used in Mendlová & Šimková [47] and considering the limited number of hosts and parasites studied, C. bulbophallus n. sp., C. pseudozambezensis n. sp., C. flagellum n. sp. and C. lobus n. sp. appear to be strict specialists, infecting only one host species. Cichlidogyrus ranula n. sp. seems to be an intermediate generalist, infecting non-congeneric cichlid species from the same tribe (i.e. Haplochromini). Finally, C. maeander n. sp. appears to be a generalist, infecting non-congeneric cichlid species of at least two different tribes (i.e. Tilapiini and Haplochromini) [24, 25].
Comparable to what was reported by Jorissen et al. [29] for the Bangwuelu-Mweru ecoregion (Upper Congo Basin), the species of Cichlidogyrus found in the Lomami River (Middle Congo Basin) also range from strict specialists to generalists. The generalist C. maeander n. sp. infects both the substrate brooding T. sparrmanii as well as the mouthbrooding Orthochromis sp. ‘Lomami’. This is in line with the hypothesis that decreasing host specificity is associated with infecting fish hosts with each exhibiting a different form of parental care [47] and contradicts Pouyaud et al. [48], who stated that species of Cichlidogyrus are not able to infect hosts with different forms of parental care.
The parasites already described from T. sparrmanii, S. macrocephalus and O. katumbii are all intermediate generalists (C. consobrini and C. halli) or generalists (C. dossoui, C. papernastrema, C. quaestio, C. sclerosus and C. zambezensis) [29, 43]. This may suggest that the host range of the species of Cichlidogyrus that were described in this study as strict specialists may expand with the parasitological examination of more fish species. Therefore, additional sampling of more potential hosts at different locations is needed to confirm or reject the host specificity of the described parasite species because the host range of a parasite species may differ between different regions or considering different scales [29, 47, 49, 50]. A more thorough parasitological screening of the Lomami River and adjacent regions is also needed to answer the question whether the distribution pattern of these species of Cichlidogyrus is determined by the geography of the basin or rather mirrors the host biogeography [30].
Perspectives
Species were initially identified following Paperna [14] and the identification key of Pariselle & Euzet [21]. The key of Pariselle & Euzet starts by splitting up the species of Cichlidogyrus into four haptoral groups (overview: Vignon et al. [22]). This split is based on the relative length of the uncinuli, considering pair III to VII of approximately the same relative length. This division into morphological groups was confirmed by Vignon et al. [22] and Pouyaud et al. [48], using molecular phylogenetic analyses based on 18S rDNA and ITS1 sequences. They found that the morphological groups are partially congruent with the monophyletic groups obtained with the molecular phylogeny. In this study, however, some species of Cichlidogyrus (C. pseudozambezensis n. sp., C. flagellum n. sp., C. maeander n. sp. and C. lobus n. sp.) were found to have uncinuli pairs III to VII of different relative length. Rahmouni et al. [51] also found several species of Cichlidogyrus from Lake Tanganyika which do not fit into the previously reported classification and suggested the existence of more than four haptoral groups. Because many new species are being described, some with a different haptoral anatomy than previously known, revision of the current classification is greatly needed.
Nowadays, a genetic approach is an integral part of taxonomic studies. Genetic analyses can be used to distinguish cryptic species and to reveal the existence of phenotypic plasticity within a species and the convergent evolution of the shape and dimensions of the hard parts within Cichlidogyrus [48, 52, 53]. Regrettably, material for such molecular analyses was not available for the current study due to fixation with formaldehyde (see Methods).
Conclusions
Sampling of fish in the Lomami River (DRC) revealed several species of Monogenea on the fish species Serranochromis cf. macrocephalus, Orthochromis sp. ‘Lomami’ and Tilapia sparrmanii. A total of six new species of Cichlidogyrus were found, ranging from strict specialists to generalists. The number of species of Cichlidogyrus possibly infesting Serranochromis cf. macrocephalus, T. sparrmanii and Orthochromis sp. ‘Lomami’ is raised substantially by the present study. Serranochromis cf. macrocephalus and Orthochromis sp. ‘Lomami’ are now known to harbour respectively three and two species of Cichlidogyrus, all described in this study. Tilapia sparrmanii are now known to harbour seven species, of which three are described in the present study. These results highlight the species diversity of Cichlidogyrus in the Congo Basin. The species described in this study show morphological similarities to species reported both from haplotilapiine fish from the Lower Guinea province and the Upper Congo-Zambezi province. Future research and additional sampling are needed to investigate whether these morphological similarities are due to convergent evolution, typical for the monogenean fauna of certain ichthyofaunal provinces or host species, or whether this occurrence pattern is rather an artefact due to a sampling bias towards these regions and cichlid lineage.
Availability of data and materials
The data supporting the findings of this article are included within the article. The type-material of the new species described in this study is deposited in the invertebrate collection of the Royal Museum of Central Africa, Tervuren, Belgium; the collection of the research group Zoology: Biodiversity and Toxicology at Hasselt University, Diepenbeek, Belgium; the Finnish Museum of Natural History, Helsinki, Finland; and the Iziko South African Museum, Cape Town, Republic of South Africa (see “Type-material” for details on repositories and accession numbers). Tissue samples of the fish hosts are available in the ichthyology collection of the Royal Museum of Central Africa, Tervuren, Belgium.
Abbreviations
- MCC:
-
male copulatory complex
References
Gibson DI, Timofeeva TA, Gerasev PI. A catalogue of the nominal species of the monogenean genus Dactylogyrus Diesing, 1850 and their host genera. Syst Parasitol. 1996;35:3–48.
Cribb TH, Chisholm LA, Bray RA. Diversity in the Monogenea and Digenea: does lifestyle matter? Int J Parasitol. 2002;32:321–8.
Pugachev ON, Gerasev PI, Gussev AV, Ergens R, Khotenowsky I. Guide to Monogenoidea of freshwater fish of Palaearctic and Amur regions. 1st ed. Milan, Italy: Ledizioni; 2010.
Paladini G, Longshaw M, Gustinelli A, Shinn AP. Parasitic diseases in aquaculture: their biology, diagnosis and control. In: Austin B, Newaj-Fyzul A, editors. Diagnosis and Control of Diseases of Fish and Shellfish. Hoboken: Wiley; 2017. p. 37–107.
Bakke TA, Harris PD, Cable J. Host specificity dynamics: observations on gyrodactylid monogeneans. Int J Parasitol. 2002;32:281–308.
Bauer ON. Spread of parasites and diseases of aquatic organisms by acclimatization: a short review. J Fish Biol. 1991;39:679–86.
Šimková A, Řehulková E, Rasoloariniaina JR, Jorissen MWP, Scholz T, Faltýnková A, et al. Transmission of parasites from introduced tilapias: a new threat to endemic Malagasy ichthyofauna. Biol Invasions. 2019;21:803–19.
Jiménez-García MI, Vidal-Martínez VM, López-Jiménez S. Monogeneans in introduced and native cichlids in Mexico: evidence for transfer. J Parasitol. 2001;87:907–9.
FAO. The state of world fisheries and aquaculture 2014. Rome: Food and Agriculture Organization of the United Nations; 2014.
Kullander SO. A phylogeny and classification of the South American Cichlidae (Teleostei: Perciformes). In: Malabarba LR, Reis RE, Vari RP, Lucena CAS, editors. Phylogeny and classification of Neotropical fishes. Brasil: Edipucrs; 1998. p. 461–98.
Froese R, Pauly D, editors. FishBase. World Wide Web electronic publication. http://www.fishbase.org/search.php. Version 06/2018. Accessed 1 Jan 2018.
Snoeks J, Harrison IJ, Stiassny MLJ. The status and distribution of freshwater fishes. In: Darwall WRT, Smith KG, Allen DJ, Holland RA, Harrison IJ, Brooks EGE, editors. The diversity of life in African freshwaters: underwater, under threat. Gland, Switzerland and Cambridge, UK: IUCN; 2011. p. 42–73.
Pariselle A, Boeger WA, Snoeks J, Bilong Bilong CF, Morand S, Vanhove MPM. The monogenean parasite fauna of cichlids: a potential tool for host biogeography. Int J Evol Biol. 2011;2011:471480.
Paperna I. Studies on monogenetic trematodes in Israel. 2. Monogenetic trematodes of cichlids. Bamidgeh. 1960;12:20–33.
Paperna I. Onchobdella n. gen. New genus of monogenetic trematodes (Dactylogyridae, Bychowski, 1933 from cichlid fish from West Africa. Proc Helminthol Soc Wash. 1968;1968(35):200–6.
Pariselle A, Euzet L. Scutogyrus gen. n. (Monogenea, Ancyrocephalidae) for Cichlidogyrus longicornis minus Dossou, 1982, C. l. longicornis, and C. l. gravivaginus Paperna and Thurston, 1969 with description of three new species parasitic on African cichlids. J Helminthol Soc Wash. 1995;62:157–73.
Christison KW, Shinn AP, van As JG. Gyrodactylus thlapi n. sp. (Monogenea) from Pseudocrenilabrus philander philander (Weber) (Cichlidae) in the Okavango Delta, Botwana. Syst Parasitol. 2005;60:165–73.
Scholz T, Vanhove MPM, Smit N, Jayasundera Z, Gelnar M. A guide to the parasites of African freshwater fishes. Brussels: Royal Belgian Institute of Natural Sciences; 2018.
Rahmouni C, Vanhove MPM, Šimková A. Seven new species of Cichlidogyrus Paperna, 1960 (Monogenea: Dactylogyridae) parasitizing the gills of Congolese cichlids from northern Lake Tanganyika. PeerJ. 2018;6:e5604.
Jorissen MWP, Pariselle A, Huyse T, Vreven EJ, Snoeks J, Decru E, et al. Six new dactylogyrid species (Platyhelminthes, Monogenea) from the gills of cichlids (Teleostei, Cichliformes) from the Lower Congo Basin. Parasite. 2018;25:64.
Pariselle A, Euzet L. Systematic revision of dactylogyridean parasites (Monogenea) from cichlid fishes in Africa, the Levant and Madagascar. Zoosystema. 2009;31:849–98.
Vignon M, Pariselle A, Vanhove MPM. Modularity in attachment organs of African Cichlidogyrus (Platyhelminthes: Monogenea: Ancyrocephalidae) reflects phylogeny rather than host specificity or geographic distribution. Biol J Linn Soc Lond. 2011;102:694–706.
Salzburger W, Mack T, Verheyen E, Meyer A. Out of Tanganyika: genesis, explosive speciation, key-innovations and phylogeography of the haplochromine cichlid fishes. BMC Evol Biol. 2005;5:17.
Dunz AR, Schliewen UK. Molecular phylogeny and revised classification of the haplotilapiine cichlid fishes formerly referred to as “Tilapia”. Mol Phylogenet Evol. 2013;68:64–80.
Meyer BS, Matschiner M, Salzburger W. A tribal level phylogeny of Lake Tanganyika cichlid fishes based on a genomic multi-marker approach. Mol Phylogenet Evol. 2015;83:56–71.
Abell R, Thieme ML, Revenga C, Bryer M, Kottelat M, Bogutskaya N, et al. Freshwater ecoregions of the world: a new map of biogeographic units for freshwater biodiversity conservation. Bioscience. 2008;58:403–14.
Stiassny MLJ, Brummett RE, Harrison IJ, Monsembula R, Mamonekene V. The status and distribution of freshwater fishes in central Africa. In: Brooks EGE, Allen DJ, Darwall WRT, editors. The status and distribution of freshwater biodiversity in central Africa. Gland, Switzerland: IUCN; 2011. p. 27–46.
Douëllou L. Monogeneans of the genus Cichlidogyrus Paperna, 1960 (Dactylogyridae: Ancyrocephalinae) from cichlid fishes of Lake Kariba (Zimbabwe) with descriptions of five new species. Syst Parasitol. 1993;25:159–86.
Jorissen MWP, Pariselle A, Huyse T, Vreven EJ, Snoeks J, Volckaert FAM, et al. Diversity and host specificity of monogenean gill parasites (Platyhelminthes) of cichlid fishes in the Bangweulu-Mweru ecoregion. J Helminthol. 2018;92:417–37.
Vanhove MPM, Van Steenberge M, Dessein S, Volckaert FAM, Snoeks J, Huyse T, et al. Biogeographical implications of Zambezian Cichlidogyrus species (Platyhelminthes: Monogenea: Ancyrocephalidae) parasitizing Congolian cichlids. Zootaxa. 2013;3608:398–400.
Price CE, Peebles HE, Bamford T. The monogenean parasites of African fishes. IV. Two new species from South African hosts. Rev Zool Bot Afr. 1969;79:117–24.
Pariselle A, Euzet L. Four new species of Cichlidogyrus (Monogenea: Ancyrocephalidae), gill parasites of Tilapia cabrae (Teleostei: Cichlidae), with discussion on relative length of haptoral sclerites. Folia Parasitol. 2003;50:195–201.
Hyman LH. The invertebrates: Platyhelminthes and Rhynchocoela, the acoelomate Bilateria, vol. 2. New York: McGraw-Hill; 1951.
Fannes W, Vanhove MPM, Huyse T. Redescription of Cichlidogyrus tiberianus Paperna, 1960 and C. dossoui Douëllou, 1993 (Monogenea: Ancyrocephalidae), with special reference to the male copulatory organ. Syst Parasitol. 2017;94:133–44.
Wägele J. Foundations of phylogenetic systematics. 2nd ed. München: Verlag Dr. Friedrich Pfeil; 2005.
Davis JI, Nixon KC. Populations, genetic variation, and the delimitation of phylogenetic species. Syst Biol. 1992;41:421–35.
International Commission on Zoological Nomenclature. Article 50: Authors of names and nomenclatural acts. In: Ride WDL, Cogger HG, Dupuis C, Kraus O, Minelli A, Thompson FC, et al., editors. International code of zoological nomenclature. 4th ed. London: The International Trust for Zoological Nomenclature; 1999. p. 2015.
ICZN. International Commission on Zoological Nomenclature: Amendment of articles 8, 9, 10, 21 and 78 of the International Code of Zoological Nomenclature to expand and refine methods of publication. Bull Zool Nomencl. 2012;69:161–9.
Birgi E, Lambert A. Présence chez un Nandidae (Téléostéen), Polycentropsis abbreviata Boulenger, 1901, du genre Cichlidogyrus (Monogenea, Monopisthocotylea, Ancyrocephalidae) - Description de Cichlidogyrus nandidae n. sp. et de Cichlidogyrus euzeti n. sp. Ann Parasitol Hum Comp. 1986;61:521–8.
Birgi E, Lambert A. A propos de Cichlidogyrus euzeti Birgi et Lambert, 1986 décrit au Cameroun chez un Nandidae, Polycentropsis abbreviata. Ann Parasitol Hum Comp. 1987;62:103.
Vanhove MPM, Volckaert FAM, Pariselle A. Ancyrocephalidae (Monogenea) of Lake Tanganyika: I: Four new species of Cichlidogyrus from Ophthalmotilapia ventralis (Teleostei: Cichlidae), the first record of this parasite family in the basin. Zoologia. 2011;28:253–63.
Pariselle A, Bilong Bilong CF, Euzet L. Four new species of Cichlidogyrus Paperna, 1960 (Monogenea, Ancyrocephalidae), all gill parasites from African mouthbreeder tilapias of the genera Sarotherodon and Oreochromis (Pisces, Cichlidae), with a redescription of C. thurstonae Ergens, 1981. Syst Parasitol. 2003;56:201–10.
Schedel FDB, Vreven E, Manda BK, Abwe E, Manda AC, Schliewen UK. Description of five new rheophilic Orthochromis species (Teleostei: Cichlidae) from the Upper Congo drainage in Zambia and the Democratic Republic of the Congo. Zootaxa. 2018;4461:301–49.
Pariselle A, Euzet L. Cichlidogyrus Paperna, 1960 (Monogenea, Ancyrocephalidae): gill parasites from West African Cichlidae of the subgenus Coptodon Regan, 1920 (Pisces), with descriptions of six new species. Syst Parasitol. 1996;34:109–24.
Pariselle A, Euzet L. New species of Cichlidogyrus Paperna, 1960 (Monogenea, Ancyrocephalidae) from the gills of Sarotherodon occidentalis (Daget) (Osteichthyes, Cichlidae) in Guinea and Sierra Leone (West Africa). Syst Parasitol. 1997;38:221–30.
Van Steenberge M, Vreven E, Snoeks J. The fishes of the Upper Luapula area (Congo basin): a fauna of mixed origin. Ichthyol Explor Freshwaters. 2014;24:329–45.
Mendlová M, Šimková A. Evolution of host specificity in monogeneans parasitizing African cichlid fish. Parasit Vectors. 2014;7:69.
Pouyaud L, Desmarais E, Deveney M, Pariselle A. Phylogenetic relationships among monogenean gill parasites (Dactylogyridea, Ancyrocephalidae) infesting tilapiine hosts (Cichlidae): systematic and evolutionary implications. Mol Phylogenet Evol. 2006;38:241–9.
Poulin R, Krasnov BR, Mouillot D. Host specificity in phylogenetic and geographic space. Trends Parasitol. 2011;27:355–61.
Krasnov BR, Poulin R. Ecological properties of a parasite: species-specific stability and geographical variation. In: Morand S, Krasnov BR, editors. The biogeography of host-parasite interactions. New York: Oxford University Press; 2010. p. 99–114.
Rahmouni C, Vanhove MPM, Šimková A. Underexplored diversity of gill monogeneans in cichlids from Lake Tanganyika: eight new species of Cichlidogyrus Paperna, 1960 (Monogenea: Dactylogyridae) from the northern basin of the lake, with remarks on the vagina and the heel of the male copulatory organ. Parasit Vectors. 2017;10:591.
Kmentová N, Gelnar M, Mendlová M, Van Steenberge M, Koblmüller S, Vanhove MPM. Reduced host-specificity in a parasite infecting non-littoral Lake Tanganyika cichlids evidenced by intraspecific morphological and genetic diversity. Sci Rep. 2016;6:39605.
Messu Mandeng FDM, Bilong Bilong CF, Pariselle A, Vanhove MPM, Bitja Nyom AR, Agnèse J-F. A phylogeny of Cichlidogyrus spp. (Monogenea, Dactylogyridae) clarifies a host-switch between fish families and reveals an adaptive component to attachment organ morphology of this parasite genus. Parasit Vectors. 2015;8:582.
Acknowledgements
We appreciate the help of the people involved in the field work and sampling procedure. Furthermore, we would like to thank Maarten Van Steenberge (IRSNB, Belgium), Ulrich Schliewen (SNSB, Germany), Fredi Schedel (SNSB, Germany) and Bauchet Katemo Manda (Royal Museum of Central Africa and KU Leuven, Belgium; BEZHU, DRC) for their help in the identification of the fish species. Finally, we would like to thank Ria Vanderspikken (Hasselt University) and Natascha Steffanie (Hasselt University) for their technical support in the laboratory.
Funding
The research leading to the results presented in this publication was partly carried out with infrastructure funded by EMBRC Belgium - FWO project GOH3817N. This study was also supported by the Belgian Federal Science Policy Office (BRAIN-be Pioneer Project BR/132/PI/TILAPIA), Czech Science Foundation Project No. P505/12/G112 (ECIP) and the “MbiSa Congo” project (2013–2018), a framework agreement project of the RMCA with the Belgian Development Cooperation. MG is supported by the Bijzonder Onderzoeksfonds (BOF-Project 7NI02) of Hasselt University for the analysis, interpretation of data and the writing of the manuscript.
Author information
Authors and Affiliations
Contributions
TA, MPMV and TH designed and supervised this study. ACM contributed to sampling, the collection and identification of fish, and provided scientific background information on the cichlids from the Lomami Basin. EV helped in the identification of the fish specimens. FMB collected and prepared the gill parasites, and did a first microscopical examination. MG performed the morphological characterisation and described the species. TA, MPMV and MG analysed the data and wrote the paper. AP helped with the interpretation and discussion of the results, and provided scientific background in the field of monogenean research. TA and MPMV critically revised the draft. All authors critically read and edited the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Fish were acquired from local fishermen and not taken from the wild especially for this study. In the absence of relevant animal welfare regulations in the Democratic Republic of the Congo, guidelines in force in Europe were applied.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Geraerts, M., Muterezi Bukinga, F., Vanhove, M.P.M. et al. Six new species of Cichlidogyrus Paperna, 1960 (Platyhelminthes: Monogenea) from the gills of cichlids (Teleostei: Cichliformes) from the Lomami River Basin (DRC: Middle Congo). Parasites Vectors 13, 187 (2020). https://doi.org/10.1186/s13071-020-3927-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-020-3927-4