
Abstract
Background
Gexia-Zhuyu Tang (GZT), a traditional Chinese medicine formula, is used to treat a variety of diseases. However, its roles in gastric cancer (GC) remain unclear.
Objective
The aim of this study was to explore the roles and underlying molecular mechanisms of modified GZT in GC.
Methods
The effects of modified GZT on GC were investigated by constructing mouse xenograft models with MFC cell line. The fecal samples from low-dose, high-dose, and without modified GZT treatment groups were collected for the 16S rRNA gene sequencing and fecal microbiota transplantation (FMT). Histopathological alterations of mice were evaluated using the hematoxylin–eosin (HE). Immunohistochemical (IHC) analysis with Ki67 and GSDMD was performed to measure tissue cell proliferation and pyroptosis, respectively. Proteins associated with pyroptosis, invasion, and metastasis were detected by Western blotting. Enzyme-linked immunosorbent assay (ELISA) was used to assess inflammation-related factors levels.
Results
Modified GZT inhibited GC tumor growth and reduced metastasis and invasion-related proteins expression levels, including CD147, VEGF, and MMP-9. Furthermore, it notably promoted caspase-1-dependent pyroptosis, as evidenced by a dose-dependent increase in TNF-α, IL-1β, IL-18, and LDH levels, along with elevated protein expression of NLRP3, ASC, and caspase-1. Additionally, modified GZT increased species abundance and diversity of the intestinal flora. FMT assay identified that modified GZT inhibited GC tumor progression through regulation of intestinal flora.
Conclusions
Modified GZT treatment may promote pyroptosis by modulating gut microbiota in GC. This study identifies a new potential approach for the GC clinical treatment.
Similar content being viewed by others


Background
Gastric cancer (GC) is a prevalent malignancy, ranking fifth in terms of new cases in 2020 and fourth in terms of mortality, causing approximately 800,000 deaths [1, 2]. Even with various treatment options like surgery, chemotherapy, radiotherapy, drugs, and immunotherapy, the prognosis for patients with GC remains unsatisfactory [3]. Therefore, the identification of alternative and effective treatments for GC are of paramount importance.
Having been practiced for over 5000 years, Traditional Chinese Medicine (TCM) has gained recognition as a credible and promising alternative therapy for cancer. Gexia-Zhuyu Tang (GZT) is a TCM Formula (TCMF) that shows promise in treating chronic diseases [4]. It is composed of 12 Chinese herbal medicines, including Peach kernel, Safflower, Angelica sinensis, Chuanxiong, Red peony, Peony bark, YanhuSuo, Wulingzhi, Wuyao, Fructus Aurantii, ** between the M and H groups, accounting for 6.33%, while the H group had 15888 unique OTUs, accounting for 53.77% (Fig. 5A). At the phylum level, the phyla Firmicutes, Bacteroidetes, and Proteobacteria were relatively abundant (Fig. 5B). With high-dose modified GZT treatment, the abundance of Bacteroidetes, Proteobacteria, and Tenericutes increased, and the abundance of Firmicutes decreased. At the genus level, the relative abundance of Bacteroides, Lactobacillus, and Oscillospira ranked in the top three. In the H group, the abundance of Bacteroides decreased compared to the M group, while the abundance of Coprococcus and Psychrobacter increased (Fig. 5B). The heatmap was generated for the top 50 genera based on the mean abundance. The high-dose modified GZT treatment reduced the abundance of Desulfovibrio, Helicobacter, and Facklamia, while increased the abundance of Alistipes, Sutterella, Paraprevotella, and Psychrobacter (Fig. 5C).

The modified GZT treatment alters the community of intestinal flora in mice with GC. A Venn diagram showed the overlap** OTUs in different groups. B Average relative abundance of gut microbiota at the phylum and genus level, respectivly, in different regions of the Venn diagram. C Differences in gut microbiota in the Model, Lower dose, and Higher dose group at the genus level were demonstrated by heatmaps
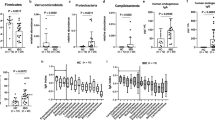
By analyzing the iconic species, notable differences in ASV/out were found between the M group and the H group (Additional file 1: Figure S3A). The abundance of Desulfovibrionaceae and Desulfovibrio was significantly higher in the M group compared to the H group (Additional file 1: Figure S3B). The analysis of marker species showed significant differences in the abundance of species between the M group and the H group (Additional file 1: Figure S3C). The taxonomic hierarchy, from phylum to genus, in each group of samples was shown in Additional file 1: Figure S1D. At the order, family, and genus levels, the marker species in the modified GZT high-dose treatment group were Turicibacterales, Turicibacteraceae, and Turicibacter.
The PCA method was used to study the degree of dispersion of the differential species. The results demonstrated a distinct trend of separation between the M and H groups, with the ratio of the difference in species abundance composition to the total difference for all samples at 43.3% (Additional file 1: Figure S4A). At the same time, the results of OPLS-DA indicated that the abundance composition of the differential species between the M and H groups exhibited a separation trend with only a few overlap** individual parts and a high overall separation, indicating a better classification effect (Additional file 1: Figure S4B).
Modified GZT inhibits the GC development by regulating intestinal flora
The Paraprevotella, Psychrobacter, Trematoda, Alistipes, Coprococcus, Oscillospira, and Sutterella are not the predominant bacterial community in GC. Therefore, the study opted for a more comprehensive analysis by using the entire fecal microbiota for the FMT experiment, which provided insights into how GZT treatment affected the gut microbiota and its potential impact on gastric cancer in mice. Fecal microorganisms from donor mice were extracted and transferred to recipient mice to evaluate the impact of FMT on GC progression in mice (Fig. 6A). The results indicated that a noteworthy decrease was observed in both the tumor volume and weight in the high-dose of the modified GZT (FMT) group compared to the Model (FMT) group (Fig. 6B–D). Moreover, there was a significant reduction in immune cell infiltration and inflammatory cells in the Higher Dose (FMT) group compared to the Model (FMT) group (Fig. 6E). IHC assay results showed a significant increase in GSDMD expression in the Higher Dose (FMT) group compared with the Model (FMT) group (Fig. 6F). Furthermore, modified GZT (FMT) decreased the number of Ki67-positive cells (Fig. 6G). The expression levels of metastasis and invasion-associated proteins CD147, VEGF, and MMP-9 were significantly lower in the Higher Dose (FMT) group than in the Model (FMT) group (Fig. 6H).

The modified GZT treatment inhibits tumor growth in vivo by regulating intestinal flora. A Schematic diagram of fecal gavage experiment in BALB/c mice. B Photographs of tumors in mice. C Tumor volume growth curve. D Tumor weight change curve. E Histopathological changes observed by HE staining (200 × , scale bar: 100 μm); F The expression level of GSDMD was assessed by IHC (magnification: 200 × , scale bar: 50 μm). G Ki67 immunohistochemical staining was used to detect cell proliferation (200 × , scale bar: 100 μm). H Western blot was used to detect the protein expression levels of CD147, VEGF and MMP-9 in the Model (FMT) and the Higher Dose (FMT) group. *P < 0.05; **P < 0.01 vs. Model group
Modified GZT promotes pyroptosis by regulating gut microbiota
Next, we used FMT to detect whether gut microbiota altered by modified GZT had therapeutic benefits for GC. Fecal microbiota from Model or higher dose modified GZT-treated mice were transplanted into GC mice recipients. ELISA analysis showed that the levels of TNF-α, IL-1β, IL-18, and LDH levels were significantly lower in the Model (FMT) group compared to the Control group. In contrast, the levels of TNF-α, IL-1β, IL-18, and LDH levels in the Higher Dose (FMT) group were significantly higher than that in the Model (FMT) group (Fig. 7A, B). Furthermore, the expression of NLRP3, ASC, and caspase-1 proteins were increased in the Higher Dose (FMT) group than that in the Model (FMT) group (Fig. 7C).

The modified GZT inhibits pyroptosis by regulating gut microbiota. A–B ELISA assay was used to detect the expression levels of TNF-α, IL-1β, IL-18 and LDH. C Western blot was used to detect the protein expression of NLRP3, ASC, and caspase-1. *P < 0.05; **P < 0.01 vs. Control group; ##P < 0.01 vs. Model group
Discussion
GC is a malignancy that originates from the inner lining of the stomach with a high morbidity and mortality rate [36]. H. pylori infection, diet, family history, and alcohol consumption are all high risk factors for GC [37], of which H. pylori infection is responsible for approximately 70% of all GC cases worldwide [38]. Advances in molecular biology and sequencing technologies have enabled researchers to study the gut microbiome in greater depth and breadth in GC, not just H. Pylori. 16S rRNA sequencing has proven to be a powerful tool for exploring the diversity and composition of microbial communities. In the gut microbiome, 16S rRNA sequencing has been employed to study the effects of diet, drug therapy, and disease on microbial diversity and composition [39]. In the current study, we demonstrated that modified GZT inhibited the growth, proliferation, metastasis, invasion, and promoted pyroptosis of GC. Through 16S rRNA sequencing analysis, we detected alterations in the gut microbiota following modified GZT treatment. This study demonstrates that modified GZT effectively suppresses GC progression by modulating the gut microbiota.
First, we analyzed the metabolites of modified GZT by HPLC–MS/MS. The results showed that the metabolites of modified GZT mainly included dihydroxybenzoic acid, 4-methylcatechol, ailanthone, and 18β-glycyrrhetinic acid. In GC 3,4-dihydroxybenzoic acid promotes cell apoptosis through JNK/p38 MAPK signaling activation and exerts anti-tumor effects [40]. The 18β-glycyrrhetinic acid exhibits properties with potential benefits, including antitumor, anti-inflammatory, and antiviral effects [41,42,43]. Furthermore, 18β-glycyrrhetinic acid impedes the development and advancement of GC by downregulating COX-2 expression and suppressing Wnt-1 expression through miR-149–3 upregulation [44]. It inhibits the migration and invasion of GC cells through the ROS/PKC-α/ERK pathway [45]. 4-methylcatechol induces apoptosis via the intrinsic mitochondrial pathway and demonstrates cytotoxic effects on metastatic malignant melanoma cells [46]. Ailanthone possesses anti-inflammatory, anti-malarial, and anti-tumor effects. It can inhibit the progression of multiple tumors, including acute myeloid leukemia, lung, breast, melanoma, and gastric cancer [47]. These findings all suggest that modified GZT may have an important role in inhibiting tumor development.
Then we explored the effects of modified GZT on GC. The results revealed that modified GZT significantly inhibited tumor growth in a dose-dependent manner. The Ki-67 marker is a recognized parameter for measuring proliferation in malignant tumors [48]. IHC results showed that modified GZT reduced the number of Ki67-positive cells, inhibiting the proliferation of GC tumor tissue cells. CD147, which is highly expressed on the extracellular surface of numerous types of tumor tissue, potentially promotes tumor metastasis through its regulation of cellular substrate and adhesion mechanisms [49]. The role of CD147 in promoting tumor invasiveness has been recently confirmed in various human malignancies, including GC [33]. Angiogenesis plays a crucial role in the metastasis of solid tumors, with VEGF standing out as the most potent mediator of angiogenesis [50]. MMP-9, a member of the matrix metalloproteinase family, is involved in GC metastasis and invasion [51]. In the present study, we found that both low and high doses of modified GZT suppressed the expression of CD147, VEGF, and MMP-9, implying that modified GZT inhibited the metastasis and invasion of GC tumors. In addition, IHC staining demonstrated that modified GZT significantly increased the expression level of GSDMD. GSDMD is a widely investigated pore-forming protein in pyroptosis, and it is a substrate for activated caspase-1/4/5/11 [52]. Further investigation showed that modified GZT greatly promoted the protein expression levels of NLRP3, ASC and caspase-1, and the concentrations of TNF-α, IL-1β, IL-10 and LDH. Subsequent FMT showed that modified GZT inhibited the development of GC by modulating the gut microbiota.
Composition and species abundance of intestinal flora are significantly altered in GC. Studies have shown that as the progression from gastritis to intestinal metaplasia and GC occurs, there is a gradual decline in both diversity and abundance of gut microbiota [23, 53]. In GC, the abundance of Fusobacterium, Leptotrichia, Veillonella, Campylobacter and Haemophilus is notably elevated [54]. In comparison to patients with intestinal metaplasia and gastritis, those with GC exhibit a significantly higher abundance of Lactobacillus, Clostridium, and Lachnospiraceae [55]. The fecal microbiota promotes inflammation, affects cell proliferation and immune regulation, and disrupts DNA integrity, ultimately contributing to carcinogenesis [56]. The most common Helicobacter species is H. Pylori. The mechanism through which H. pylori infection induces GC mainly involves chronic gastritis, releasing gastrin and histamine, damaging DNA, and activating the proliferation of gastric epithelial cells through multiple pathways such as the induction of the PI3K/Akt pathway [57]. The increased abundance of Lachnospiracea in GC may be related to inflammatory regulation [58]. In our study, we found that after high-dose modified GZT treatment, the abundances of many gut flora species and the community composition were significantly changed. At the genus level, the abundance of several genera including Bacteroides, Desulfovibrio, Helicobacter, and Facklamia decreased, while the abundance of Paraprevotella, Psychrobacter, Trematoda, Alistipes, Coprococcus, Oscillospira, and Sutterella increased.
Numerous studies have shown that TCMF can regulate intestinal flora and affect the development of intestinal cancer. A previous study has found that the intestinal flora diversity, microbial composition, and abundance in patients with precancerous lesions of GC are altered after treatment with Weifuchun capsules [59]. Quxie Capsules elevate the abundance of Actinobacteria and Lachnospiraceae, while decrease the abundance of Bacteroides, Bacteroidetes, Escherichia Shigella, and Gammaproteobacteria, which may help protect against colorectal cancer tumors and enhance immunity [60]. Curcumin, a compound generated by the roots of the Curcuma longa plant, may have anti-carcinogenic qualities via preserving the diversity of gut bacteria [61]. Antibiotics are extensively employed in the realm of Western medicine for the management of a broad spectrum of ailments [62]. Nevertheless, their usage is concomitant with an alteration in the composition and a reduction in the diversity of the human microbiota [63, 64]. Investigation has revealed a progressive increase in the risk of GC associated with the cumulative number of penicillin courses administered [65]. Antibiotic administration not only influences the resistome of the subject to whom it is given, but also the whole population owing to selection for resistance to its function [66]. Prior research has demonstrated that the use of antibiotics in conjunction with prebiotics, probiotics, and synbiotics assists the gut microbiota in resisting H. pylori infections and lowers the proportion of drug-resistant bacteria [67]. Therefore, we hypothesize that modified GZT combined with antibiotic therapy plays a role in preserving gut microbiota homeostasis and decreasing drug resistance.
Inflammasomes in the gastrointestinal tract are associated with intestinal flora homeostasis and infection [68]. Studies have shown that TCM can reduce H. pylori-induced gastritis by decreasing the expression of IL-8, TNF-α, IL-6, iNOS, and IFN-γ [69]. These findings suggest that there is an important correlation between TCM and intestinal flora, inflammation, and pyroptosis. In this study, we found that high dose modified GZT facilitated the expression proinflammatory factors TNF-α, IL-1β, IL-18, and caspase-1-dependent pyroptosis by modulating intestinal flora. Caspase-1 activates primarily the pro-inflammatory factors IL-1β and IL-18 [70]. Moreover, studies have shown that some pathogen-associated molecular patterns, carried by bacteria, are delivered to the host cytoplasm where they activate inflammasomes [68]. Thus, we speculate that modified GZT promotes caspase-1-dependent pyroptosis by inhibiting harmful bacteria, which in turn enhances the release of proinflammatory factors.
However, there are also limitations in this investigation. Firstly, we relied on previous studies for GZT ingredients by HPLC analysis and did not conduct HPLC analysis of modified GZT in this study due to scientific limitations. Secondly, for the assessment of pyroptosis, we determined the expression levels of GSDMD, inflammatory factors, and pyroptosis-related proteins using IHC assay. However, morphological analysis was not performed through electron microscopy due to scientific limitations. Additionally, in the evaluation of modified GZT effect on the metastasis and invasion of GC, we solely assessed the protein expression levels of CD147, VEGF, and MMP-9, which will be further validated by in vitro experiments in our subsequent exploration.
Conclusion
In this study, we found that the modified GZT inhibited the growth, proliferation, metastasis, and invasion of GC by regulating the intestinal flora. Furthermore, modified GZT also promoted pyroptosis of GC. The current investigation revealed a novel therapeutic strategy and theoretical foundation for utilizing TCMF in the treatment of GC.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- TCMF:
-
Traditional Chinese medicine formula
- GC:
-
Gastric cancer
- GZT:
-
Gexia-Zhuyu Tang
- FMT:
-
Fecal microbiota transplantation
- HE:
-
Hematoxylin–eosin
- IHC:
-
Immunohistochemical
- ELISA:
-
Enzyme-linked immunosorbent assay
- HPLC:
-
High-performance liquid chromatography
- AQPs:
-
Aquaporins
- TNF-α:
-
Tumor necrosis factor-alpha
- IL-1β:
-
Interleukin-1 beta
- IL-18:
-
Interleukin-18
- LDH:
-
Lactate dehydrogenase
- OTUs:
-
Operational taxonomic units
- RDP:
-
Ribosomal database project
- PCoA:
-
Principal coordinates analysis
- PCA:
-
Principal component analysis
- OPLS-DA:
-
Orthogonal partial least squares discriminant analysis
- SD:
-
Means ± standard deviation
- ANOVA:
-
Analysis of variance
References
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 Countries. CA Cancer J Clin. 2021;71(3):209–49. https://doi.org/10.3322/caac.21660.
Smyth EC, Nilsson M, Grabsch HI, van Grieken NC, Lordick F. Gastric cancer. Lancet. 2020;396(10251):635–48. https://doi.org/10.1016/S0140-6736(20)31288-5.
Alsina M, Arrazubi V, Diez M, Tabernero J. Current developments in gastric cancer: from molecular profiling to treatment strategy. Nat Rev Gastroenterol Hepatol. 2023;20(3):155–70. https://doi.org/10.1038/s41575-022-00703-w.
Chen J, Chen H, Cheng J, Lin H, Tung Y, Lin C, Chen C. A Chinese herbal medicine, Gexia-Zhuyu Tang (GZT), prevents dimethylnitrosamine-induced liver fibrosis through inhibition of hepatic stellate cells proliferation. J Ethnopharmacol. 2012;142(3):811–8. https://doi.org/10.1016/j.jep.2012.06.005.
Hibasami H, Iwase H, Yoshioka K, Takahashi H. Glycyrrhizin induces apoptosis in human stomach cancer KATO III and human promyelotic leukemia HL-60 cells. Int J Mol Med. 2005;16(2):233–6.
Khan U, Karmakar BC, Basak P, Paul S, Gope A, Sarkar D, Mukhopadhyay AK, Dutta S, Bhattacharya S. Glycyrrhizin, an inhibitor of HMGB1 induces autolysosomal degradation function and inhibits Helicobacter pylori infection. Mol Med. 2023;29(1):51. https://doi.org/10.1186/s10020-023-00641-6.
Wang XZ, **a L, Zhang XY, Chen Q, Li X, Mou Y, Wang T, Zhang YN. The multifaceted mechanisms of Paeoniflorin in the treatment of tumors: State-of-the-Art. Biomed Pharmacother. 2022;149:112800. https://doi.org/10.1016/j.biopha.2022.112800.
Deng Z, Zhang S, Ge S, Kong F, Cao S, Pan Z. Gexia-Zhuyu decoction attenuates carbon tetrachloride-induced liver fibrosis in mice partly via liver angiogenesis mediated by myeloid cells. Med Sci Monit. 2019;25:2835–44. https://doi.org/10.12659/MSM.913481.
Liao KF, Chiu TL, Huang SY, Hsieh TF, Chang SF, Ruan JW, Chen SP, Pang CY, Chiu SC. Anti-cancer effects of radix angelica sinensis (Danggui) and N-Butylidenephthalide on gastric cancer: implications for REDD1 activation and mTOR inhibition. Cell Physiol Biochem. 2018;48(6):2231–46. https://doi.org/10.1159/000492641.
Yi B, Liu D, He M, Li Q, Liu T, Shao J. Role of the ROS/AMPK signaling pathway in tetramethylpyrazine-induced apoptosis in gastric cancer cells. Oncol Lett. 2013;6(2):583–9. https://doi.org/10.3892/ol.2013.1403.
Wang D, Wang S, Liu W, Li M, Zheng Q, Li D. Hydroxysafflor yellow B induces apoptosis via mitochondrial pathway in human gastric cancer cells. J Pharm Pharmacol. 2022. https://doi.org/10.1093/jpp/rgac044.
Huang Y, Yan S, Su Z, **a L, **e J, Zhang F, Du Z, Hou X, Deng J, Hao E. Aquaporins: a new target for traditional Chinese medicine in the treatment of digestive system diseases. Front Pharmacol. 2022;13:1069310. https://doi.org/10.3389/fphar.2022.1069310.
Lee T, Lee D, Lee S, Ko Y, Sung Kang K, Chung S, Kim K. Sesquiterpenes from Curcuma zedoaria rhizomes and their cytotoxicity against human gastric cancer AGS cells. Bioorg Chem. 2019;87:117–22. https://doi.org/10.1016/j.bioorg.2019.03.015.
Zhang R, Ma C, Wei Y, Wang X, Jia J, Li J, Li K, Cao G, Yang P. Isolation, purification, structural characteristics, pharmacological activities, and combined action of Hedyotis diffusa polysaccharides: A review. Int J Biol Macromol. 2021;183:119–31. https://doi.org/10.1016/j.ijbiomac.2021.04.139.
Wang C, **n P, Wang Y, Zhou X, Wei D, Deng C, Sun S. Iridoids and sfingolipids from Hedyotis diffusa. Fitoterapia. 2018;124:152–9. https://doi.org/10.1016/j.fitote.2017.11.004.
Shim JH, Gim H, Lee S, Kim BJ. Inductions of caspase-, MAPK- and ROS-dependent apoptosis and chemotherapeutic effects caused by an ethanol extract of Scutellaria barbata D. Don in human gastric adenocarcinom cells. J Pharmacopuncture. 2016;19(2):129–36. https://doi.org/10.3831/KPI.2016.19.014.
Bik EM, Eckburg PB, Gill SR, Nelson KE, Purdom EA, Francois F, Perez-Perez G, Blaser MJ, Relman DA. Molecular analysis of the bacterial microbiota in the human stomach. Proc Natl Acad Sci U S A. 2006;103(3):732–7. https://doi.org/10.1073/pnas.0506655103.
Deng Y, Tang D, Hou P, Shen W, Li H, Wang T, Liu R. Dysbiosis of gut microbiota in patients with esophageal cancer. Microb Pathog. 2021;150:104709. https://doi.org/10.1016/j.micpath.2020.104709.
Bai X, Wei H, Liu W, Coker OO, Gou H, Liu C, Zhao L, Li C, Zhou Y, Wang G, Kang W, Ng EK, Yu J. Cigarette smoke promotes colorectal cancer through modulation of gut microbiota and related metabolites. Gut. 2022;71(12):2439–50. https://doi.org/10.1136/gutjnl-2021-325021.
Stewart OA, Wu F, Chen Y. The role of gastric microbiota in gastric cancer. Gut Microbes. 2020;11(5):1220–30. https://doi.org/10.1080/19490976.2020.1762520.
Eslick GD. Helicobacter pylori infection causes gastric cancer? A review of the epidemiological, meta-analytic, and experimental evidence. World J Gastroenterol. 2006;12(19):2991–9. https://doi.org/10.3748/wjg.v12.i19.2991.
Peng X, Zhou L, Gong Y, Song Z, He L, Lin S, Zhang J. Non-pylori Helicobacters (NHPHs) induce shifts in gastric microbiota in helicobacter pylori-infected patients. Front Microbiol. 2017;8:1038. https://doi.org/10.3389/fmicb.2017.01038.
Ferreira RM, Pereira-Marques J, Pinto-Ribeiro I, Costa JL, Carneiro F, Machado JC, Figueiredo C. Gastric microbial community profiling reveals a dysbiotic cancer-associated microbiota. Gut. 2018;67(2):226–36. https://doi.org/10.1136/gutjnl-2017-314205.
Lu Y, Liu H, Yang K, Mao Y, Meng L, Yang L, Ouyang G, Liu W. A comprehensive update: gastrointestinal microflora, gastric cancer and gastric premalignant condition, and intervention by traditional Chinese medicine. J Zhejiang Univ Sci B. 2022;23(1):1–18. https://doi.org/10.1631/jzus.B2100182.
Bao Z, Wu G, Du J, Ye Y, Zheng Y, Wang Y, Ji R. The comparative efficacy and safety of 9 traditional Chinese medicines combined with standard quadruple therapy for Helicobacter pylori-associated gastritis: a systematic review and network meta-analysis. Ann Transl Med. 2022;10(24):1349. https://doi.org/10.21037/atm-22-5421.
Hao X, Zhou P, Yang Z, Yang T, Wang Y. The therapeutic effect of Huazhuojiedu decoction on precancerous lesions in a gastric cancer model via the regulation of lnc 517368. J Ethnopharmacol. 2022;283:114635. https://doi.org/10.1016/j.jep.2021.114635.
Yu P, Zhang X, Liu N, Tang L, Peng C, Chen X. Pyroptosis: mechanisms and diseases. Signal Transduct Target Ther. 2021;6(1):128. https://doi.org/10.1038/s41392-021-00507-5.
Sborgi L, Ruhl S, Mulvihill E, Pipercevic J, Heilig R, Stahlberg H, Farady CJ, Muller DJ, Broz P, Hiller S. GSDMD membrane pore formation constitutes the mechanism of pyroptotic cell death. EMBO J. 2016;35(16):1766–78. https://doi.org/10.15252/embj.201694696.
Joosten LA, Netea MG, Dinarello CA. Interleukin-1beta in innate inflammation, autophagy and immunity. Semin Immunol. 2013;25(6):416–24. https://doi.org/10.1016/j.smim.2013.10.018.
Zhang X, Li C, Chen D, He X, Zhao Y, Bao L, Wang Q, Zhou J, **e Y. H. pylori CagA activates the NLRP3 inflammasome to promote gastric cancer cell migration and invasion. Inflamm Res. 2022;71(1):141–55. https://doi.org/10.1007/s00011-021-01522-6.
Li L, Bao B, Chai X, Chen X, Su X, Feng S, Zhu X. The anti-inflammatory effect of Callicarpa nudiflora extract on H. Pylori-infected GES-1 Cells through the inhibition of ROS/NLRP3/Caspase-1/IL-1beta signaling axis. Can J Infect Dis Med Microbiol. 2022;2022:5469236. https://doi.org/10.1155/2022/5469236.
Zhao Z, Li Q, Ashraf U, Yang M, Zhu W, Gu J, Chen Z, Gu C, Si Y, Cao S, Ye J. Zika virus causes placental pyroptosis and associated adverse fetal outcomes by activating GSDME. eLife. 2022. https://doi.org/10.7554/eLife.73792.
Bae WJ, Kim S, Ahn JM, Han JH, Lee D. Estrogen-responsive cancer-associated fibroblasts promote invasive property of gastric cancer in a paracrine manner via CD147 production. FASEB J. 2022;36(11):e22597. https://doi.org/10.1096/fj.202200164RR.
Ma F, Zhang B, Ji S, Hu H, Kong Y, Hua Y, Luo S. Hypoxic macrophage-derived VEGF promotes proliferation and invasion of gastric cancer cells. Dig Dis Sci. 2019;64(11):3154–63. https://doi.org/10.1007/s10620-019-05656-w.
Krishnaveni D, Bhayal AC, Sri Manjari K, Vidyasagar A, Uma Devi M, Ramanna M, Jyothy A, Nallari P, Venkateshwari A. MMP 9 gene promoter polymorphism in gastric cancer. Indian J Clin Biochem. 2012;27(3):259–64. https://doi.org/10.1007/s12291-012-0210-2.
Petryszyn P, Chapelle N, Matysiak-Budnik T. Gastric cancer: where are we heading? Dig Dis. 2020;38(4):280–5. https://doi.org/10.1159/000506509.
Zhang R, Li H, Li N, Shi JF, Li J, Chen HD, Yu YW, Qin C, Ren JS, Chen WQ, He J. Risk factors for gastric cancer: a large-scale, population-based case-control study. Chin Med J (Engl). 2021;134(16):1952–8. https://doi.org/10.1097/CM9.0000000000001652.
Jain U, Saxena K, Chauhan N. Helicobacter pylori induced reactive oxygen species: a new and develo** platform for detection. Helicobacter. 2021;26(3):e12796. https://doi.org/10.1111/hel.12796.
Gao B, Chi L, Zhu Y, Shi X, Tu P, Li B, Yin J, Gao N, Shen W, Schnabl B. An introduction to next generation sequencing bioinformatic analysis in gut microbiome studies. Biomolecules. 2021. https://doi.org/10.3390/biom11040530.
Lin HH, Chen JH, Huang CC, Wang CJ. Apoptotic effect of 3,4-dihydroxybenzoic acid on human gastric carcinoma cells involving JNK/p38 MAPK signaling activation. Int J Cancer. 2007;120(11):2306–16. https://doi.org/10.1002/ijc.22571.
Pan B, Zheng L, Shi Y, Dong Z, Feng T, Yang J, Wei Y, Zhou Y. βSynthesis and antiviral and antitumor activities of Novel 18-glycyrrhetinic acid derivatives. Int J Mol Sci. 2023. https://doi.org/10.3390/ijms241915012.
Sun J, Liu H, Lv C, Qin J, Wu Y. Modification, antitumor activity, and targeted PPARγ study of 18β-glycyrrhetinic acid, an important active ingredient of licorice. J Agric Food Chem. 2019;67(34):9643–51. https://doi.org/10.1021/acs.jafc.9b03442.
Feng Y, Mei L, Wang M, Huang Q, Huang R. Anti-inflammatory and pro-apoptotic effects of 18beta-glycyrrhetinic acid in vitro and in vivo models of rheumatoid arthritis. Front Pharmacol. 2021;12:681525. https://doi.org/10.3389/fphar.2021.681525.
Cao D, Jia Z, You L, Wu Y, Hou Z, Suo Y, Zhang H, Wen S, Tsukamoto T, Oshima M, Jiang J, Cao X. 18β-glycyrrhetinic acid suppresses gastric cancer by activation of miR-149–3p-Wnt-1 signaling. Oncotarget. 2016;7(44):71960–73. https://doi.org/10.18632/oncotarget.12443.
Cai H, Chen X, Zhang J, Wang JJJonm,. 18β-glycyrrhetinic acid inhibits migration and invasion of human gastric cancer cells via the ROS/PKC-α/ERK pathway. J Nat Med. 2018;72(1):252–9. https://doi.org/10.1007/s11418-017-1145-y.
Payton F, Bose R, Alworth WL, Kumar AP, Ghosh R. 4-Methylcatechol-induced oxidative stress induces intrinsic apoptotic pathway in metastatic melanoma cells. Biochem Pharmacol. 2011;81(10):1211–8. https://doi.org/10.1016/j.bcp.2011.03.005.
Ding H, Yu X, Hang C, Gao K, Lao X, Jia Y, Yan Z. Ailanthone: a novel potential drug for treating human cancer. Oncol Lett. 2020;20(2):1489–503. https://doi.org/10.3892/ol.2020.11710.
Folescu R, Levai CM, Grigoras ML, Arghirescu TS, Talpos IC, Gindac CM, Zamfir CL, Poroch V, Anghel MD. Expression and significance of Ki-67 in lung cancer. Rom J Morphol Embryol. 2018;59(1):227–33.
Tang Y, Kesavan P, Nakada MT, Yan L. Tumor-stroma interaction: positive feedback regulation of extracellular matrix metalloproteinase inducer (EMMPRIN) expression and matrix metalloproteinase-dependent generation of soluble EMMPRIN. Mol Cancer Res. 2004;2(2):73–80.
Chen S, Zhang X, Peng J, Zhai E, He Y, Wu H, Chen C, Ma J, Wang Z, Cai S. VEGF promotes gastric cancer development by upregulating CRMP4. Oncotarget. 2016;7(13):17074–86. https://doi.org/10.18632/oncotarget.7717.
Zhao C, Yuan G, Jiang Y, Xu J, Ye L, Zhan W, Wang JJEcr,. Capn4 contributes to tumor invasion and metastasis in gastric cancer via activation of the Wnt/β-catenin/MMP9 signalling pathways. Exp Cell Res. 2020;395(2):112220. https://doi.org/10.1016/j.yexcr.2020.112220.
Yan H, Luo B, Wu X, Guan F, Yu X, Zhao L, Ke X, Wu J, Yuan J. Cisplatin induces pyroptosis via activation of MEG3/NLRP3/caspase-1/GSDMD pathway in triple-negative breast cancer. Int J Biol Sci. 2021;17(10):2606–21. https://doi.org/10.7150/ijbs.60292.
Coker OO, Dai Z, Nie Y, Zhao G, Cao L, Nakatsu G, Wu WK, Wong SH, Chen Z, Sung JJY, Yu J. Mucosal microbiome dysbiosis in gastric carcinogenesis. Gut. 2018;67(6):1024–32. https://doi.org/10.1136/gutjnl-2017-314281.
Castano-Rodriguez N, Goh KL, Fock KM, Mitchell HM, Kaakoush NO. Dysbiosis of the microbiome in gastric carcinogenesis. Sci Rep. 2017;7(1):15957. https://doi.org/10.1038/s41598-017-16289-2.
Sun L, Ma W, Gao W, **ng Y, Chen L, **a Z, Zhang Z, Dai Z. Propofol directly induces caspase-1-dependent macrophage pyroptosis through the NLRP3-ASC inflammasome. Cell Death Dis. 2019;10(8):542. https://doi.org/10.1038/s41419-019-1761-4.
Wroblewski LE, Peek RM Jr, Coburn LA. The role of the microbiome in gastrointestinal cancer. Gastroenterol Clin North Am. 2016;45(3):543–56. https://doi.org/10.1016/j.gtc.2016.04.010.
Xu W, Huang Y, Yang Z, Hu Y, Shu X, **e C, He C, Zhu Y, Lu NJO, therapy,. Helicobacter pylori promotes gastric epithelial cell survival through the PLK1/PI3K/Akt pathway. Onco Targets Ther. 2018;11:5703–13. https://doi.org/10.2147/ott.S164749.
Berry D, Reinisch W. Intestinal microbiota: a source of novel biomarkers in inflammatory bowel diseases? Best Pract Res Clin Gastroenterol. 2013;27(1):47–58. https://doi.org/10.1016/j.bpg.2013.03.005.
Bian Y, Chen X, Cao H, **e D, Zhu M, Yuan N, Lu L, Lu B, Wu C, Bahaji Azami NL, Wang Z, Wang H, Zhang Y, Li K, Ye G, Sun M. A correlational study of Weifuchun and its clinical effect on intestinal flora in precancerous lesions of gastric cancer. Chin Med. 2021;16(1):120. https://doi.org/10.1186/s13020-021-00529-9.
Sun L, Yan Y, Chen D, Yang Y. Quxie capsule modulating gut microbiome and its association with T cell regulation in patients with metastatic colorectal cancer: result from a randomized controlled clinical trial. Integr Cancer Ther. 2020;19:1534735420969820. https://doi.org/10.1177/1534735420969820.
McFadden RM, Larmonier CB, Shehab KW, Midura-Kiela M, Ramalingam R, Harrison CA, Besselsen DG, Chase JH, Caporaso JG, Jobin C, Ghishan FK, Kiela PR. The role of curcumin in modulating colonic microbiota during colitis and colon cancer prevention. Inflamm Bowel Dis. 2015;21(11):2483–94. https://doi.org/10.1097/MIB.0000000000000522.
Moser C, Lerche C, Thomsen K, Hartvig T, Schierbeck J, Jensen P, Ciofu O, Høiby N. Antibiotic therapy as personalized medicine—general considerations and complicating factors. APMIS. 2019;127(5):361–71. https://doi.org/10.1111/apm.12951.
Dethlefsen L, Huse S, Sogin M, Relman D. The pervasive effects of an antibiotic on the human gut microbiota, as revealed by deep 16S rRNA sequencing. Plos Biol. 2008;6(11):e280. https://doi.org/10.1371/journal.pbio.0060280.
Jakobsson H, Jernberg C, Andersson A, Sjölund-Karlsson M, Jansson J, Engstrand L. Short-term antibiotic treatment has differing long-term impacts on the human throat and gut microbiome. Plos ONE. 2010;5(3):e9836. https://doi.org/10.1371/journal.pone.0009836.
Boursi B, Mamtani R, Haynes K, Yang YX. Recurrent antibiotic exposure may promote cancer formation–Another step in understanding the role of the human microbiota? Eur J Cancer. 2015;51(17):2655–64. https://doi.org/10.1016/j.ejca.2015.08.015.
Blaser MJ. Antibiotic use and its consequences for the normal microbiome. Science. 2016;352(6285):544–5. https://doi.org/10.1126/science.aad9358.
Meng C, Bai C, Brown TD, Hood LE, Tian Q. Human gut microbiota and gastrointestinal cancer. Genomics Proteomics Bioinformatics. 2018;16(1):33–49. https://doi.org/10.1016/j.gpb.2017.06.002.
Man SM. Inflammasomes in the gastrointestinal tract: infection, cancer and gut microbiota homeostasis. Nat Rev Gastroenterol Hepatol. 2018;15(12):721–37. https://doi.org/10.1038/s41575-018-0054-1.
Song MY, Lee DY, Park SY, Seo SA, Hwang JS, Heo SH, Kim EH. Steamed ginger extract exerts anti-inflammatory effects in helicobacter pylori-infected gastric epithelial cells through inhibition of NF-kappaB. J Cancer Prev. 2021;26(4):289–97. https://doi.org/10.15430/JCP.2021.26.4.289.
Man SM, Kanneganti TD. Regulation of inflammasome activation. Immunol Rev. 2015;265(1):6–21. https://doi.org/10.1111/imr.12296.
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
TZ: conceptualization; investigation; methodology; project administration; formal analysis; validation; visualization; writing—review & editing; ZY: methodology; resources; visualization; writing—original draft; all authors have read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Compliance with the National Institutes of Health's guidelines for the care and use of laboratory animals, this study was approved by the Animal Ethics Committee of Southern Medical University (23410).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
Total Ion Chromatogram peak of the top 10 metabolites from negative ion model. Figure S2. Total Ion Chromatogram peak of the top 10 metabolites from positive ion model. Figure S3. Analysis of marker species of intestinal flora. A. MetagenomeSeq test results in the Model and the Higher Dose group. B. The log2 values of the top five ASV/OUT and ASV/OTU multiples that were significantly up- and down-regulated. Positive values represent upregulation in M group compared with H or L group. C. Histogram of distribution of LDA values for significantly different species. D. Taxonomic branching diagram showing taxonomic hierarchy relationships from phylum to genus in each taxon sample. Figure S4. PCA and OPLS-DA analysis of differential species. A. Two-dimensional sorting diagram of samples for PCA analysis. B. Sorting diagram of samples for OPLS-DA discriminant analysis.
Additional file 2: Table S1.
The gradient elution process of modified GZT. Table S2. The top 10 metabolites of modified GZT from negative ion model. Table S3. The top 10 metabolites of modified GZT from positive ion model.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhao, T., Yu, Z. Modified Gexia-Zhuyu Tang inhibits gastric cancer progression by restoring gut microbiota and regulating pyroptosis. Cancer Cell Int 24, 21 (2024). https://doi.org/10.1186/s12935-024-03215-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-024-03215-6