
Abstract
Background
Pre-implantation development is a crucial step in successful implantation and pregnancy in mammals. It has been studied in depth, but mostly in laboratory animal models. Less is known about the regulatory mechanism involved in the pre-implantation development in humans and about the comparative aspects.
Methods
Here, we employed the microarray datasets from the public database library of GEO and applied comparative analysis of genome wide temporal gene expression data based on statistical analysis and functional annotation for both mouse and human, demonstrating the discordance between the regulatory mechanisms of both mouse and human pre-implantation development.
Results
There were differences between mouse and human pre-implantation development both in the global gene expression pattern and in the expression changes of individual genes at each stage, including different major transient waves of transcription profiles and some stage-specific genes and significantly related pathways. There also appeared to be different functional changes from one stage to another between mouse and human.
Conclusions
The analysis presented here lead to interesting and complementary conclusions that the regulatory mechanism of human pre-implantation development is not completely the same as the mouse. Not as the fact that 1-cell to 2-cell stage is important for mouse pre-implantation development, the 4-cell stage and 8-cell stage are both essential for human. Unlike in mouse, of which most of pathways found were related to energy, RNA and protein metabolism, the identified pathways in human were mostly disease-related and associated with human pre-implantation embryonic development. All of these suggest that a further comparative analysis should be required for applying the result of mouse expression data to human research or therapy, particularly in pre-implantation developments. Our study provides several potential targets of genes and pathways for studying the regulatory mechanism of human pre-implantation development using mouse model.
Similar content being viewed by others


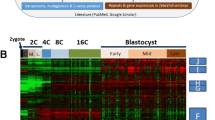
Background
Pre-implantation development is a mammalian-specific occurrence, which encompasses the period from fertilization to implantation and involves a number of important events [1]. Understanding pre-implantation development is important, both for basic reproductive biology and for practical applications including regenerative medicine and livestock production. However, due to the scarcity of the materials, both in size (about 0.1 mm diameter) and in quantity (only a few to tens of oocytes from each ovulation) are limited in related research, which has hampered the molecular analysis of human pre-implantation embryos. Thus using the mouse model system has formed the current perfect paradigm about gene expression during pre-implantation development. Recently, more and more global gene expression profiles during mouse pre-implantation development have been examined, two principal transient waves of de novo transcription have been identified [2, 3]. Additionally, several important transcripts have been reported to have the core roles at each developmental stage. For example, H1foo and Msy2 have been reported as the oocyte-specific transcripts, which are not re-expressed later in development, destruction of these maternal mRNAs restricts the length of time that these genes can function [4, 5]; Recent studies have shown that JNK and p38 are involved in cavity formation during pre-implantation development [6, 7]. As the unclear confidence of the previously identified genes, their roles in the regulation of mouse pre-implantation development must be further discussed and the consistence with human must also be considered.
In our study, we employed time course expression datasets of early mouse and human embryo both from the same series. Through our comparative analysis of genome wide temporal gene expression data based on significance analysis and functional annotation, we found that there were many differences in the expression patterns of pre-implantation development between mouse and human, for both the regulatory waves and the identified genes.
Methods
Data collection
We searched GEO [8] for the gene expression profiling studies related to pre-implantation development. Finally, we chose the data set GSE18290 contributed by ** of the human and murine orphan receptors ERRalpha (ESRRA) and ERRbeta (ESRRB) and identification of a novel human ERRalpha-related pseudogene. Genomics. 1997, 45: 320-326. 10.1006/geno.1997.4939." href="/article/10.1186/1477-7827-8-41#ref-CR21" id="ref-link-section-d75648876e930">21]. The second group appears to represent genes that showed a dramatic decrease throughout the stages, abundant in one cell stage but degraded during pre-implantation development, which is cluster 3 (1883 probe sets). The third group is exactly opposite to the second one, it appears to represent genes that showed a dramatic increase throughout the developmental stages, including cluster 5 (690 probe sets). The last group includes cluster 4 (1721 probe sets) and cluster 6 (690 probe sets), the genes here were firstly activated from one cell stage to 8-cell stage, peaked at 8-cell stage but degraded at later stages. In fact, the last group contains Mapk1 and Mapk3, respectively in cluster 4 and cluster 6, which are both well-known genes having central roles in diverse cellular functions [26]. In our study, the profiles of XIST expression in both mouse and human pre-implantation development were known. As a result, the expression of **st was significantly regulated at mouse 2-cell stage and blastocyst, but almost significantly regulated through human pre-implantation development (from 2-cell to blastocyst). On the other hand, in the k-means clustering of significant genes, the mouse **st gene was in Cluster 5 but the human XIST gene was in Cluster 3. The details were shown in Additional file 1 and 2, colored in red. Thus, all of these demonstrate that not as in the mouse, the pattern of human XIST expression is not consistent with a role for early expression in the choice of paternal X inactivation but participates in regulating the whole human early embryo development.
Significantly related GO terms and pathways
We then sought to determine the functions associated with the identified significant genes between each adjacent stage, analysis of over-represented annotations and pathways were performed by using DAVID [14, 15]. If p value was less than 0.01, it was considered as significant GO annotation or pathway. The significant GO terms of over-represented genes were shown in Table 2 and 3 respectively for mouse or human. The significant pathways of over-represented genes were shown in Table 4 and 5 respectively for mouse or human.
With comparison to mouse, due to the less different identified genes from one-cell stage to two-cell stage and from two-cell stage to 4-cell stage for human, there were less GO terms during these periods. The GO terms for human focused on such as "cell division", "cellular metabolic process", "primary metabolic process" and "RNA processing". These were all obtained for mouse, except for the GO term "death". Just like to be delayed, most of previous GO terms for mouse were appearing from 4-cell to 8-cell stage for human, such as "macromolecule metabolic process", "macromolecule localization", "establishment of protein localization" and "establishment of RNA localization". These findings support the idea that mid-preimplantation gene activation (MGA) drives the overt morphological changes in the subsequent stages, compaction and bifurcation into two-cell lineages. With the preparation of basic cellular machinery, dramatic biological processes were happening during Human Phase 3 (morula and blastocyst), which is related to the event of implantation.
Briefly, through our pathway analysis, most of the identified pathways were involved in after human 4-cell stage, but the number of identified pathways shared in mouse pre-implantation development. Moreover, many of the features and signaling pathways that are required during human pre-implantation development are also active during tumourigenesis [27]. According to our analysis based on human data, p53 signaling pathway, the pathways of Eenteropathogenic E. coli (EPEC) infection and Enterohemorrhagic E. coli (EHEC) infection were identified separately in 4-cell and 8-cell stages. These pathways were also thought to be related to the formation of cancer and the regulation of human reproduction [28–31]. But these above pathways were not identified from microarray analysis in mouse 4-cell and 8-cell stages. On the contrary, the pathway of oxidative phosphorylation was mostly identified through mouse pre-implantation development, which hardly appeared in human data. The oxidative phosphorylation pathway was reported as one of the obligatory energy metabolism pathways in most species throughout pre-implantation development [32]. And the pathway of Citrate cycle (TCA cycle) was identified after mouse 8-cell stage, which was not identified in human data. It is well know that the mitochondrial TCA cycle is the major source of reducing equivalents in the cytosol so that any change in mitochondrial function in the embryo will be reflected in changes in the intracellular redox state. In the mouse, the metabolic substrates used by the oocyte and early embryo each have a different impact on the intracellular redox state [33]. Surprisingly, the common identified pathway from 4-cell to 8-cell stage between human and mouse was only the Ubiquitin mediated proteolysis pathway, which was reported as an important role in eukaryotic cellular processes [34]. Thus it is advisable to choose the Ubiquitin mediated proteolysis pathway as the candidate pathway for further study on reproductive biology and regenerative medicine using mouse model. Interestingly, some of the known metabolic pathways e.g. biosynthesis of steroids pathway and fatty acid metabolism pathway were shown significant in both human and mouse blastocysts. All of these suggest that unlike in mouse, of which most of pathways found were related to energy, RNA and protein metabolism, the identified pathways in human were mostly disease-related and associated with human pre-implantation embryonic development. By contrast, there are some common metabolic pathways participate in regulating the mammal early embryonic development.
Furthermore, in view of the fact that many researchers working in mice interpret their results in relation to human reproduction, mice and human reproduction differ distinctly in many aspects; e.g. the reproduction in mice is characterized by a very short oestrus cycle and thus, displays a distinctly different endocrine dynamic pattern if compared with humans or domestic animals [35–38]. On the other hand, due to the difference in the inner environment of embryo and the external environment of uterus, there are species differences in the implantation process between mice and human [39–41]. The same applies for the regulatory mechanisms of early embryonic development that can be not simply transferable to the human species.
In sum, pre-implantation development involves a number of biologically significant events, such as compaction and blastocyst formation, which represent morphologically dynamic changes, especially for mouse and human [27]. Although the models of mouse have been well examined, molecular mechanisms regulating the early embryo development of human have been scarcely reported and the credibility of using the mouse model to explain the regulatory mechanism of human pre-implantation development remains unclear. Briefly, in this study, we have shown differences between mouse and human pre-implantation developments both in the global gene expression pattern and the expression changes of individual genes at each stage, including different major transient waves of transcription profiles and some stage-specific genes and pathways. Undeniably, Studies in mice have provided insights into the molecular basis of human pre-implantation development because of their shared features. In both species, early embryo development leads to a complex regulatory mechanism. However, the nature of human embryonic signals that influence uterine functions is more difficult than rodents', especially at the later stages of pre-implantation development (morula and blastocyst). Thus, it limits the availability of adequate amounts of tissues for mouse analysis. Moreover, the quality and quantity of samples chosen in microarray experiments also play essential roles in effecting the accuracy of mouse model, especially the conditions for in vitro maturation and fertilization are crucial for the proper early embryo development and may result in developmental aberrations.
Conclusions
By comparison between mouse and human expression profiles, we have noted that the regulatory mechanism of human pre-implantation development is different from the mouse, and even more complex. Through our analysis, we have found differences between mouse and human transcription profiles both in the global expression pattern of genes and in expression of individual genes within the gene clusters identified and in the significantly related pathways. Not as the fact that 1-cell to 2-cell stage is important for mouse pre-implantation development, the 4-cell stage and 8-cell stage are both essential for human. Unlike in mouse, of which most of pathways found were related to energy, RNA and protein metabolism, the identified pathways in human were mostly disease-related and associated with human pre-implantation embryonic development. Different expression patterns and significantly related biological processes during each stage between mouse and human suggest that a further comparative analysis should be required for applying the result of mouse expression data to human research or therapy, particularly in pre-implantation developments. Our study also provides several potential targets of genes and pathways for studying the regulatory mechanism of human pre-implantation development using mouse model, such as the gene of **st and the Ubiquitin mediated proteolysis pathway.
References
Edwards RG: Aspects of the molecular regulation of early mammalian development. Reprod Biomed Online. 2003, 6: 97-113. 10.1016/S1472-6483(10)62061-5.
Hamatani T, Carter MG, Sharov AA, Ko MS: Dynamics of global gene expression changes during mouse preimplantation development. Dev Cell. 2004, 6: 117-131. 10.1016/S1534-5807(03)00373-3.
Wang QT, Piotrowska K, Ciemerych MA, Milenkovic L, Scott MP, Davis RW, Zernicka-Goetz M: A genome-wide study of gene activity reveals developmental signaling pathways in the preimplantation mouse embryo. Dev Cell. 2004, 6: 133-144. 10.1016/S1534-5807(03)00404-0.
Tanaka M, Hennebold JD, Macfarlane J, Adashi EY: A mammalian oocyte-specific linker histone gene H1oo: homology with the genes for the oocyte-specific cleavage stage histone (cs-H1) of sea urchin and the B4/H1 M histone of the frog. Development. 2001, 128: 655-664.
Yu J, Hecht NB, Schultz RM: Expression of MSY2 in mouse oocytes and preimplantation embryos. Biol Reprod. 2001, 65: 1260-1270. 10.1095/biolreprod65.4.1260.
Maekawa M, Yamamoto T, Tanoue T, Yuasa Y, Chisaka O, Nishida E: Requirement of the MAP kinase signaling pathways for mouse preimplantation development. Development. 2005, 132: 1773-1783. 10.1242/dev.01729.
Natale DR, Paliga AJ, Beier F, D'Souza SJ, Watson AJ: p38 MAPK signaling during murine preimplantation development. Dev Biol. 2004, 268: 76-88. 10.1016/j.ydbio.2003.12.011.
Gene Expression Omnibus. [http://www.ncbi.nlm.nih.gov/geo]
**e D, Chen CC, Ptaszek LM, **ao S, Cao X, Lewin HA, Cowan C, Zhong S: Rewirable gene regulatory networks in the preimplantation embryonic development of three mammalian species. Genome Res. 2010
Gautier L, Cope L, Bolstad BM, Irizarry RA: affy--analysis of Affymetrix GeneChip data at the probe level. Bioinformatics. 2004, 20: 307-315. 10.1093/bioinformatics/btg405.
Irizarry RA, Hobbs B, Collin F, Beazer-Barclay YD, Antonellis KJ, Scherf U, Speed TP: Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics. 2003, 4: 249-264. 10.1093/biostatistics/4.2.249.
Park T, Yi SG, Lee S, Lee SY, Yoo DH, Ahn JI, Lee YS: Statistical tests for identifying differentially expressed genes in time-course microarray experiments. Bioinformatics. 2003, 19: 694-703. 10.1093/bioinformatics/btg068.
Team RDC: R: A language and environment for statistical computing. 2009, R Foundation for Statistical Computing, Vienna, Austria
Dennis G, Sherman BT, Hosack DA, Yang J, Gao W, Lane HC, Lempicki RA: DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4: P3-10.1186/gb-2003-4-5-p3.
Huang da W, Sherman BT, Lempicki RA: Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009, 4: 44-57. 10.1038/nprot.2008.211.
Braude P, Bolton V, Moore S: Human gene expression first occurs between the four- and eight-cell stages of preimplantation development. Nature. 1988, 332: 459-461. 10.1038/332459a0.
Jeong HJ, Kim HJ, Lee SH, Kwack K, Ahn SY, Choi YJ, Kim HG, Lee KW, Lee CN, Cha KY: Gene expression profiling of the pre-implantation mouse embryo by microarray analysis: comparison of the two-cell stage and two-cell block. Theriogenology. 2006, 66: 785-796. 10.1016/j.theriogenology.2006.01.059.
Adjaye J, Herwig R, Brink TC, Herrmann D, Greber B, Sudheer S, Groth D, Carnwath JW, Lehrach H, Niemann H: Conserved molecular portraits of bovine and human blastocysts as a consequence of the transition from maternal to embryonic control of gene expression. Physiol Genomics. 2007, 31: 315-327. 10.1152/physiolgenomics.00041.2007.
Saeed AISV, White J, Li J, Liang W, Bhagabati N, Braisted J, Klapa M, Currier T, Thiagarajan M, Sturn A, Snuffin M, Rezantsev A, Popov D, Ryltsov A, Kostukovich E, Borisovsky I, Liu Z, Vinsavich A, Trush V, Quackenbush J: TM4: a free, open-source system for microarray data management and analysis. Biotechniques. 2003, 34: 374-378.
Larsen LH, Rose CS, Sparso T, Overgaard J, Torekov SS, Grarup N, Jensen DP, Albrechtsen A, Andersen G, Ek J, Glumer C, Borch-Johnsen K, Jorgensen T, Hansen T, Pedersen O: Genetic analysis of the estrogen-related receptor alpha and studies of association with obesity and type 2 diabetes. Int J Obes (Lond). 2007, 31: 365-370. 10.1038/sj.ijo.0803408.
Sladek R, Beatty B, Squire J, Copeland NG, Gilbert DJ, Jenkins NA, Giguere V: Chromosomal map** of the human and murine orphan receptors ERRalpha (ESRRA) and ERRbeta (ESRRB) and identification of a novel human ERRalpha-related pseudogene. Genomics. 1997, 45: 320-326. 10.1006/geno.1997.4939.
Graf K, ** XP, Yang D, Fleck E, Hsueh WA, Law RE: Mitogen-activated protein kinase activation is involved in platelet-derived growth factor-directed migration by vascular smooth muscle cells. Hypertension. 1997, 29: 334-339.
Nohara A, Ohmichi M, Koike K, Masumoto N, Kobayashi M, Akahane M, Ikegami H, Hirota K, Miyake A, Murata Y: The role of mitogen-activated protein kinase in oxytocin-induced contraction of uterine smooth muscle in pregnant rat. Biochem Biophys Res Commun. 1996, 229: 938-944. 10.1006/bbrc.1996.1905.
Pearson G, Robinson F, Beers Gibson T, Xu BE, Karandikar M, Berman K, Cobb MH: Mitogen-activated protein (MAP) kinase pathways: regulation and physiological functions. Endocr Rev. 2001, 22: 153-183. 10.1210/er.22.2.153.
Zernicka-Goetz M, Verlhac MH, Geraud G, Kubiak JZ: Protein phosphatases control MAP kinase activation and microtubule organization during rat oocyte maturation. Eur J Cell Biol. 1997, 72: 30-38.
Daniels R, Zuccotti M, Kinis T, Serhal P, Monk M: XIST expression in human oocytes and preimplantation embryos. Am J Hum Genet. 1997, 61: 33-39. 10.1086/513892.
Wang H, Dey SK: Roadmap to embryo implantation: clues from mouse models. Nat Rev Genet. 2006, 7: 185-199. 10.1038/nrg1808.
Crane JK, Naeher TM, Choudhari SS, Giroux EM: Two pathways for ATP release from host cells in enteropathogenic Escherichia coli infection. Am J Physiol Gastrointest Liver Physiol. 2005, 289: G407-G417. 10.1152/ajpgi.00137.2005.
Allende-Vega N, Saville MK: Targeting the ubiquitin-proteasome system to activate wild-type p53 for cancer therapy. Semin Cancer Biol. 2010, 20 (1): 29-39. 10.1016/j.semcancer.2009.10.004. Epub 2009 Nov 6
Hu W, Feng Z, Levine AJ: The regulation of human reproduction by p53 and its pathway. Cell Cycle. 2009, 8: 3621-3622. 10.4161/cc.8.22.9938.
Maddocks OD, Short AJ, Donnenberg MS, Bader S, Harrison DJ: Attaching and effacing Escherichia coli downregulate DNA mismatch repair protein in vitro and are associated with colorectal adenocarcinomas in humans. PLoS One. 2009, 4: e5517-10.1371/journal.pone.0005517.
Harvey AJ, Kind KL, Thompson JG: REDOX regulation of early embryo development. Reproduction. 2002, 123: 479-486. 10.1530/rep.0.1230479.
Dumollard R, Carroll J, Duchen MR, Campbell K, Swann K: Mitochondrial function and redox state in mammalian embryos. Semin Cell Dev Biol. 2009, 20: 346-353. 10.1016/j.semcdb.2008.12.013.
Sudheer S, Adjaye J: Functional genomics of human pre-implantation development. Brief Funct Genomic Proteomic. 2007, 6: 120-132. 10.1093/bfgp/elm012.
Coutifaris C, Omigbodun A, Coukos G: Integrins, endometrial maturation, & human embryo implantation. Semin Reprod Endocrinol. 1998, 16: 219-229. 10.1055/s-2007-1016280.
Bernard DJ, Woodruff TK: Inhibin binding protein in rats: alternative transcripts and regulation in the pituitary across the estrous cycle. Mol Endocrinol. 2001, 15: 654-667. 10.1210/me.15.4.654.
Lee GS, Jeung EB: Uterine TRPV6 expression during the estrous cycle and pregnancy in a mouse model. Am J Physiol Endocrinol Metab. 2007, 293: E132-E138. 10.1152/ajpendo.00666.2006.
Kaminska K, Wasielak M, Bogacka I, Blitek M, Bogacki M: Quantitative expression of lysophosphatidic acid receptor 3 gene in porcine endometrium during the periimplantation period and estrous cycle. Prostaglandins Other Lipid Mediat. 2008, 85: 26-32. 10.1016/j.prostaglandins.2007.10.001.
Allegra A, Marino A, Coffaro F, Lama A, Rizza G, Scaglione P, Sammartano F, Santoro A, Volpes A: Is there a uniform basal endometrial gene expression profile during the implantation window in women who became pregnant in a subsequent ICSI cycle?. Hum Reprod. 2009, 24: 2549-2557. 10.1093/humrep/dep222.
Arimoto-Ishida E, Sakata M, Sawada K, Nakayama M, Nishimoto F, Mabuchi S, Takeda T, Yamamoto T, Isobe A, Okamoto Y, Lengyel E, Suehara N, Morishige K, Kimura T: Up-regulation of alpha5-integrin by E-cadherin loss in hypoxia and its key role in the migration of extravillous trophoblast cells during early implantation. Endocrinology. 2009, 150: 4306-4315. 10.1210/en.2008-1662.
Najwa AR, Sengupta J, Ghosh D: A systems biology approach towards understanding the process of blastocyst implantation. Indian J Physiol Pharmacol. 2009, 53: 197-208.
Acknowledgements
**e D and Sheng Z are acknowledged for providing biological materials for this study. Our work is funded by National Natural Science Foundation of China (No.30871782), the Ministry of Science and Technology of the People's Republic of China (No.2006AA10Z1E3), National Programs for High Technology Research and Development of China (863 program)(No.2008AA101009), and the National Key Technology R&D Program (grant No. 2008BADB2B11).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
KH designed the study, collected the datasets from databases and analyzed the data, then wrote the manuscript. QSW is acknowledged for excellent technical assistance with text data mining and statistical algorithm. HBZ designed the bioinformatics study and reviewed the manuscript. YCP designed the study and reviewed the manuscript. All authors read and approved the final manuscript.
Electronic supplementary material
12958_2010_666_MOESM1_ESM.XLSX
Additional file 1:The list of mouse significantly regulated genes in each stage comparison. The data provided represent the list of mouse significantly regulated genes in each stage comparison, including from 1-cell to 2-cell (1v2); from 2-cell to 4-cell (2v4); from 4-cell to 8-cell (4v8); from 8-cell to Morula (8 vM); from Morula to Blastocyst (MvB), and containing the information of probe set IDs, Fold change, regulation type (down or up), gene symbols and gene titles. (XLSX 728 KB)
12958_2010_666_MOESM2_ESM.XLSX
Additional file 2:The list of human significantly regulated genes in each stage comparison. The data provided represent the list of human significantly regulated genes in each stage comparison, including from 1-cell to 2-cell (1v2); from 2-cell to 4-cell (2v4); from 4-cell to 8-cell (4v8); from 8-cell to Morula (8 vM); from Morula to Blastocyst (MvB), and containing the information of probe set IDs, Fold change, regulation type (down or up), gene symbols and gene titles. (XLSX 814 KB)
12958_2010_666_MOESM3_ESM.XLS
Additional file 3:significant genes of K means clustering for mouse. The data provided represent the list of significant genes in 6 clusters for mouse, containing probe set IDs, gene symbols and gene titles. (XLS 1 MB)
12958_2010_666_MOESM4_ESM.XLS
Additional file 4:significant genes of K means clustering for human. The data provided represent the list of significant genes in 6 clusters for human, containing probe set IDs, gene symbols and gene titles. (XLS 2 MB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
He, K., Zhao, H., Wang, Q. et al. A comparative genome analysis of gene expression reveals different regulatory mechanisms between mouse and human embryo pre-implantation development. Reprod Biol Endocrinol 8, 41 (2010). https://doi.org/10.1186/1477-7827-8-41
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1477-7827-8-41