

Key Points
-
The elucidation of the HIV life cycle from the mid-1980s onwards revealed targets for therapeutic intervention, ultimately leading to the development of several classes of antiretroviral drugs (ARVs), which are now usually combined in specific multidrug regimens.
-
However, such therapy eventually fails in the vast majority of patients and is typically associated with the emergence of drug resistance. Agents that target different steps in the HIV life cycle could address this problem.
-
HIV-mediated fusion of the virus to the target host cell is a multi-step process involving a series of conformational changes in the envelope glycoproteins of HIV, gp120 and gp41.
-
Enfuvirtide, a 36-amino-acid peptide derived from gp41, was found to have antiviral properties. It was subsequently shown that enfuvirtide binds to a region of gp41 and disrupts the conformational changes associated with virus–host-cell fusion, thereby blocking virus entry and inhibiting viral replication.
-
The major unmet need is for antiretroviral drugs with activity against HIV strains resistant to existing ARV drugs. The clinical development programme for enfuvirtide thus focused on patients that had been exposed to and/or had documented resistance to each of the three existing classes of ARVs.
-
In the two pivotal Phase III trials, enfuvirtide plus an individually optimized background regimen (consisting of 3–5 ARVs) demonstrated superior virological and immunological activity to an individually optimized background regimen alone after 24 weeks. Accelerated approval was granted to enfuvirtide in the United States and Europe on the basis of these results.
-
Data collected after 48 weeks from the Phase III trials confirmed the durability of enfuvirtide added to an optimized background regimen.
Abstract
Highly active antiretroviral therapy (HAART) based on combinations of drugs that target key enzymes in the life-cycle of human immunodeficiency virus (HIV) has considerably reduced morbidity and mortality from HIV infection since its introduction in the mid-1990s. However, the growing problem of the emergence of HIV strains that are resistant not only to individual drugs, but to whole drug classes, means that agents with new mechanisms of action are needed. Here, we describe the discovery and development of enfuvirtide (Fuzeon), the first drug to inhibit the entry of HIV-1 into host cells.
Similar content being viewed by others


Main
The history of combination antiretroviral therapy, also known as highly active antiretroviral therapy (HAART), for HIV infection has been characterized by an aggressive scientific and pharmaceutical response to the HIV epidemic. The elucidation of the HIV life cycle (Fig. 1) from the mid-1980s onwards revealed targets for therapeutic intervention, ultimately leading to the development of several classes of antiretroviral (ARV) drugs. Today, ARVs are usually combined in specific multidrug regimens, consisting of combinations of nucleoside (and nucleotide) reverse transcriptase inhibitors (NRTIs), non-nucleoside reverse transcriptase inhibitors (nNRTIs) and/or protease inhibitors (PIs), which are tailored to the patient and the virus.

Human immunodeficiency virus (HIV) adsorbs to the surface of host CD4 lymphocytes. The HIV envelope glycoproteins undergo a series of interactions with the CD4 receptor and one or more co-receptors (usually CCR5 or CXCR4). The culmination of these interactions is the fusing of the HIV outer membrane and the host-cell membrane, leading to virus–host-cell fusion. The viral capsid core then disassembles and viral nucleic acids enter the cytosol in association with virion proteins. A reverse-transcription complex containing the HIV reverse transcriptase catalyses complementary DNA synthesis, and the resulting complex containing viral cDNA is transported to the host-cell nucleus. Here, HIV integrase catalyses integration of viral cDNA into host-cell DNA. Expression of the viral DNA produces a precursor polypeptide that is proteolytically processed by HIV protease to give mature structural and functional viral proteins. These proteins assemble with HIV RNA at the cell membrane, from which they bud to release new virions. The targets against which approved anti-HIV agents act are indicated. nNRTI, non-nucleoside reverse transcriptase inhibitor; NRTI, nucleoside/ tide reverse transcriptase inhibitor. Adapted from Ref. 88.
HAART therapy changed the prognosis for patients with HIV infection from that of high morbidity and rapid mortality to, for many at least, a chronic, manageable, but still complicated disease1,2,3. Considering the low fidelity of the HIV reverse transcriptase enzyme combined with the high replication rate of the virus, it is not surprising that even triple-class HAART therapy eventually fails in the vast majority of patients and is typically associated with the emergence of drug resistance. Patients therefore need to be treated with sequential HAART regimens progressively, using up the diminishing pool of active drugs that are left available.
Mechanistic and in vitro genetic studies of viral isolates have signalled considerable cross-resistance within structural ARV classes, such that resistance to one member of a class translates to resistance against other members of the class. By 2000, a high proportion of patients in the United States and Europe had experienced a lengthy exposure to ARV agents and harboured virus resistant to one or more drugs4. More worryingly, it is not unusual for patients to be infected with virus already resistant to several ARVs at the time of diagnosis5. With chronic ARV use, more patients also experienced a broadening array of drug toxicities and rates of adequate compliance inevitably declined. Management of these advanced patients is a considerable concern for the treating physician6,7,8,9.
There is therefore a substantial unmet medical need for new ARVs with activity against HIV strains resistant to existing ARVs. Enfuvirtide is such an agent, developed through academic and pharmaceutical partnerships, and it is the first drug in an entirely new ARV class, the fusion inhibitors. Enfuvirtide was born in the laboratories of Duke University, nurtured by Trimeris, Inc. and developed by the collaboration of Trimeris and Roche. As the first approved ARV agent to inhibit extracellular fusion of HIV-1 to host cells, enfuvirtide faced several unique challenges, which are discussed here.
Investigation of new drug targets
HIV fusion. Among the first responses in 1983 to the identification of HIV as the aetiological agent of acquired immune deficiency syndrome (AIDS) was the search for a vaccine. HIV membrane proteins were logical targets for a vaccine, resulting in the rapid identification of these viral proteins and the investigation of their function in HIV-mediated membrane fusion10,11. The HIV envelope glycoprotein (env) is a type I integral membrane protein that is translated as a polyprotein (gp160)12,13,14. Proteolytic cleavage of gp160 yields the functional glycoproteins gp120 and gp41, which are inserted into the HIV membrane. In the viral membrane, gp120 and gp41 associate as oligomeric trimers that are non-covalently bound.
HIV-mediated fusion of the virus to the target host cell is a multi-step cascade involving a series of conformational changes in gp120 and gp41 (Fig. 2). The first event is the binding of gp120 to the CD4 receptor on the surface of LYMPHOCYTES expressing CD4. This causes unmasking of a second gp120-binding site for target chemokine co-receptors CXCR4 and CCR5 and binding of gp120 to either or both co-receptors. Although CXCR4 and CCR5 are the major co-receptors, other co-receptors can interact with gp120 in some settings11. The completion of co-receptor binding leads to the fusion-active conformation of the viral transmembrane fusion protein gp41 (Ref. 10). The ectodomain of gp41 contains two heptad repeat regions: HR1 (proximal to the N terminus) and HR2 (proximal to the C terminus). The hydrophobic fusion peptide region inserts into the host cell membrane, whereas the HR1 regions of gp41 form a trimeric coiled coil structure. HR2 regions then fold back within the hydrophobic grooves of the HR1 coiled coil, forming a hairpin structure containing a thermodynamically stable six-helix bundle that draws the viral and cellular membranes together for fusion. The exposure of gp41 and conformational changes leading to fusion are not instantaneous processes10. A transient pre-hairpin intermediate stage can last for several minutes, causing exposure of the N terminus of gp41 (Ref. 15).

It is thought that HIV enters the host cell through several separate but cooperative steps — typically abbreviated to attachment (b), co-receptor binding (c) and fusion (d–f) — which are shown here. HIV predominantly infects T cells carrying the CD4 antigen (a) through an initial association of the viral envelope glycoprotein gp120 with the CD4 receptor on the host cell (b). After this initial attachment, a conformational change is believed to occur in gp120 that allows its further association with host-cell chemokine co-receptors CCR5 and CXCR4 (c). Subsequently, a conformational change in the second viral envelope glycoprotein gp41 allows it to insert the hydrophobic N terminus into the host-cell membrane (d). The HR2 domain of gp41 then folds back on itself and associates with the HR1 domain (e); this process (known as gp41 zip**) leads to fusion of the viral and host-cell membranes and infection of the cell (f). However, in the presence of a fusion inhibitor, such as enfuvirtide (shown in yellow), an association between the fusion inhibitor and gp41 (g) prevents the successful completion of gp41 zip** (h), thereby blocking infection (i). Reproduced with permission from F. Hoffmann-La Roche and Trimeris Inc., 2001; copyright owned by Trimeris Inc. An animation of the HIV fusion process and its inhibition by enfuvirtide can be found on http://www.Roche-HIV.com.
HIV fusion targets derived from HR1 and HR2. Initially, synthetic peptides derived from gp41 were not targeted as inhibitors of HIV-1 fusion but were investigated as part of epitope-map** experiments aimed at evaluating strategies in vaccine development. Serendipitously, it was observed that when these were incubated with human T cells, an antiviral effect was seen. An understanding of the fusion process, and how envelope glycoproteins interact, led to the appreciation of how these peptides inhibited fusion and thereby were able to interrupt the HIV life cycle (Fig. 2)16.
The first synthetic peptide mimetic (DP-107) was a 38-amino-acid molecule corresponding to gp160 residues 558–595, the N-terminal region of gp41 (Ref. 17). DP-107 possessed a highly ordered, stable helical structure. In in vitro experiments, DP-107 inhibited HIV replication at concentrations approaching 6 μg per ml. Amino-acid substitutions and chemical modifications within DP-107 destabilize secondary structure and significantly reduce its antiviral activity18.
Subsequent studies examined the C terminus of gp41, because it too possessed sequences that were predicted to form an α-helical structure19,20. The 36-amino-acid peptide DP-178, later to be designated enfuvirtide (Fuzeon, formerly T-20), corresponded to amino-acid residues 643–678 of HIV-1LAI gp160 (Fig. 3). DP-178 blocked HIV-1LAI-induced SYNCYTIUM formation with an IC90 of 1.5 ng per ml (100 times more potent than DP-107). DP-178 contained the epitope for the human monoclonal antibody 2F5, implying that this region was accessible during the conformational changes leading to fusion16.

Structure of enfuvirtide.
A recombinant gp41 peptide was constructed that contained both regions, but it had little antiviral activity. However, a mutation within the N-terminal (DP-107) region restored activity. The mutation probably caused loss of helical constraint of the N-terminal region, thereby liberating the C-terminal (DP-178) region. Indeed, alteration or truncation of C-terminal sequences also eliminated activity21. Subsequent studies showed that the pre-hairpin intermediate is the target for enfuvirtide binding22.
Enfuvirtide selectively inhibits HIV-1; neither HIV-2 nor simian immunodeficiency virus (SIV) are susceptible. On the basis of its distinct extracellular mechanism of action, enfuvirtide was expected to be active against HIV-1 strains with resistance to any of the three other classes of ARVs and to multiple-drug-resistant virus, and this expectation was confirmed in both subsequent in vitro and early clinical studies23,24,25.
Early clinical studies of enfuvirtide
At the start of the enfuvirtide clinical programme, there was some scepticism as to the therapeutic potential of a peptide inhibitor of HIV fusion. Although preclinical data demonstrated substantial viral suppression19,20, enfuvirtide represented the first ARV to act extracellularly to inhibit viral entry and so it was essential to establish definitive proof-of-principle in vivo. The appropriate choice and management of patients was also crucial. The unmet medical need was in very treatment-experienced patients who had already been exposed to the existing ARVs and who had failed therapy because of the emergence of virus resistant to the existing therapies. As enfuvirtide was active against virus with multiple resistance to other ARVs, it had great potential if it could be used in heavily pre-treated patients to meet this unmet medical need. This patient population had rarely been characterized sufficiently for large clinical trials and drug registration, so outcomes would therefore be unpredictable.
A simple proof-of-principle result came reassuringly quickly. In a 14-day trial (TRI-001) to establish dose–response, enfuvirtide was administered as monotherapy by intravenous infusion in the dose range 3–100 mg, twice daily, to 16 patients. At the highest dose of 100 mg, median viral suppression was −1.96 log10 copies per ml HIV-1 RNA and dose–response was linear within the entire range23.
Further Phase II studies established the longer-term safety profile of enfuvirtide-containing regimens of 48–96 weeks26,27,28,29, and also investigated dose–response over 48 weeks of treatment27,28.
Formulation and pharmacokinetics. Human trials with enfuvirtide established predictable pharmacokinetic indices24,28,30,31. These Phase II trials also demonstrated that enfuvirtide could be administered by subcutaneous injection24 and that exposure to enfuvirtide was independent of the site of injection30. Figure 4 shows the predicted and mean observed plasma concentration–time profiles after subcutaneous injection across three doses31; the inset plot shows the predicted versus observed data. Calculated from model-independent analyses, Table 1 provides selected pharmacokinetic parameters after subcutaneous and intravenous administration.

Predicted (solid line) and mean observed (symbols) plasma concentration–time profiles after administration of a single subcutaneous bolus dose of 45 mg (squares), 90 mg (circles) and 180 mg (triangles) enfuvirtide. Inset: plot of predicted versus observed data (180 mg). Adapted from Zhang et al31.
Pharmacokinetic studies also provided confidence that sufficient plasma concentrations of enfuvirtide would be available to inhibit HIV-1 during a 12-hour dosing period. Extrapolated from Fig. 4, plasma concentrations of enfuvirtide surpass the upper range of viral sensitivity for at least 12 hours. Taking into account area under the curve (AUC)12h, plasma half-life and other pharmacokinetic measures, enfuvirtide clearly possessed the pharmacokinetic properties and potency necessary for further study in HIV-infected patients.
Drug–drug interactions. Being a peptide, enfuvirtide is expected to undergo catabolism to its constituent amino acids, followed by retention and recycling of these amino acids in the body pool rather than metabolism by cytochrome P450 (CYP450). Therefore, there is a low potential for enfuvirtide to affect or be affected by other drugs. To confirm this, several studies have evaluated the drug–drug interaction potential of enfuvirtide.
The results of the so-called Pittsburgh cocktail study (T20-502) showed that the administration of enfuvirtide (90 mg twice daily) to HIV-infected patients was not associated with clinically relevant effects on the pharmacokinetics of several drugs selected because of their known route of metabolism (caffeine for CYP1A2, chlorzoxone for CYP2E1, dapsone for CYP3A4, debrisoquine for CYP2D6 and mephenytoin for CYP2C19)32.
A further three studies have investigated the effect of rifampicin, ritonavir (Norvir; Abbott) and ritonavir-boosted saquinavir (Invirase/Fortovase; Roche) on the pharmacokinetics of enfuvirtide33. Rifampicin was selected because it is a well-characterized and potent enzyme inducer and is used in the treatment of tuberculosis in HIV-infected patients. Ritonavir was selected because multiple studies have shown that this HIV protease inhibitor is the most potent CYP450 inhibitor clinically studied, and ritonavir-boosted saquinavir was selected as a commonly administered boosted protease-inhibitor regimen. All three studies showed that enfuvirtide was not affected to a clinically meaningful extent by any of the other drugs administered33.
Manufacturing of enfuvirtide. Relatively small quantities of other medical peptides, such as calcitonin, are successfully manufactured synthetically and the early proof-of-concept studies with enfuvirtide also used synthetically produced drug. This method of manufacturing was considered to have the greatest likelihood of success for rapidly scaling up to a commercial scale, although significant challenges were anticipated. These challenges are obvious when one considers that enfuvirtide is a 36-amino-acid peptide with a complex structure and a molecular weight of 4,492 Da. Due to its large size, the manufacture of enfuvirtide is very complex, involving 106 steps in the chemical pathway. In comparison, a typical manufacturing process for small-molecule drugs usually involves between eight and twelve steps. It is the most complex synthetic peptide manufactured at such a massive scale (the current manufacturing capacity is >3 metric tonnes per year).
Enfuvirtide, which is supplied as a 100-mg lyophilized, carbonate-buffered powder that must be reconstituted with 1.1 ml of sterile water at the time of administration, is given twice daily by a 1.0 ml subcutaneous injection. The approved dose is 90 mg twice daily, although a dose higher than 180 mg per day has not been studied clinically. However, the approved dose gave a strong pharmacodynamic profile with evidence that the 180 mg per day dose was on the plateau portion of the exposure–response curve34. Furthermore, higher doses would entail a higher injection burden for the patient that might be expected to impact the acceptance of therapy.
Efficacy of enfuvirtide
The pivotal Phase III TORO (T-20 versus Optimized Regimen Only) trials were successful in several respects. With ∼1,000 treatment-experienced patients enrolled, they offered definitive proof of the therapeutic value of enfuvirtide. Because of their inclusiveness, the TORO studies enrolled treatment-experienced patients with broad demographic, disease and virological characteristics (Table 2).
TORO 1 was conducted in the United States, Mexico, Brazil and Canada35 and TORO 2 was conducted in Europe and Australia36. The trial designs were almost identical, differing only in the minimum length of previous experience for ARVs from each class (six months for TORO 1 versus three months for TORO 2), and the number of previous PIs that patients must have received (two for TORO 1 and one for TORO 2). To enter the TORO studies, patients must have been exposed to, and/or have documented resistance to, each of the three existing classes of ARVs (NRTIs, nNRTIs and PIs). Figure 5 schematically describes the TORO study design.

The novel TORO study design accommodated treatment-experienced patients by analysing virus at screen genotype (GT) and phenotype (PT) and selecting an optimized treatment background (OB) that patients began at baseline (BL). After week eight, patients who failed OB therapy could switch to revised OB regimens and receive enfuvirtide. The initial randomization was 2 (enfuvirtide + OB): 1 (OB alone). First planned analysis was at week 24 and the second was at week 48. *Criteria for virological failure based on two consecutive values: <0.5 log10 decrease from BL starting at week six and eight; <1.0 log10 decrease from BL starting at week fourteen and sixteen; and ≥ 2 log response and and >1 log rebound at any time.
HIV-1 from each patient underwent genotypic and phenotypic analysis during screening. Investigators and patients were instructed to choose an individualized 'Optimized Background' (OB) ARV regimen of three to five drugs on the basis of the viral drug resistance testing as well as the patient's previous history and tolerability. Once investigators and patients had committed to this OB regimen, they were randomized (at a 2:1 ratio) to enfuvirtide plus OB (enfuvirtide group) or to the OB regimen alone (control group). If patients experienced virological failure during the TORO trials while receiving the OB regimen alone, they could revise their OB regimen on the basis of repeat viral resistance testing and add enfuvirtide (referred to as 'switch').
Through 24 weeks, the primary efficacy endpoint was change from baseline in HIV-1 RNA — a departure from most previous Phase III ARV trials evaluating HAART therapies in ARV-naive patients that used a categorical endpoint of proportion of patients below limit of assay detection of HIV-1 RNA. Such categorical endpoints were secondary endpoints in the TORO analyses, because of the uncertainty about how many experienced patients would reach these levels and how well the OB regimen might perform. (If the categorical response on the control arm was very high or very low, there was a risk that a true treatment difference could be missed.)
In both TORO trials, therapy with enfuvirtide together with OB for 24 weeks resulted in a further reduction in viral load from baseline, approaching 1.0 log10 copies per ml35,36. Immunological response was also greater in the enfuvirtide group, with an additional benefit of +44 cells per mm3 in TORO 1 and +28 cells per mm3 in TORO 2 (Refs 35,36). This was a particularly impressive finding given the advanced stage of disease in the patient group enrolled.
Combining the data from both TORO studies was pre-planned. As the studies were of almost identical design, if the outcomes of the two studies were similar, then the combined database would provide a more accurate estimate of the treatment effect and also provide additional power for subgroup analyses. In this merged data set37, enfuvirtide plus OB again demonstrated superior virological and immunological activity to OB alone in a 24-week period.
Accelerated approval was granted to enfuvirtide in the United States and Europe on the basis of these Phase III 24-week results. Regulatory authorities and the medical community both demand long-term data for conventional approval and for reassurance of the durability of response. Data collected after 48 weeks from TORO 1 and TORO 2 were combined and analysed by INTENT-TO-TREAT (ITT) methods. Differing from the 24-week analyses, the categorical endpoints were the primary endpoints for the 48-week analysis of durability of response. These analyses confirmed the durability of enfuvirtide added to an OB regimen38 (Table 3), and showed that a high proportion of patients in each treatment group who responded at week 24 (HIV-1 RNA <400 copies per ml) also responded at week 48 (80% in the enfuvirtide group and 70% in the control group39).
Subgroup analyses. The principal 48-week results extended to demographic and baseline subgroups. Enfuvirtide worked equally well in men and women, irrespective of age or race40. Patients that were healthier at the onset of treatment (higher CD4 count, lower viral load) derived the greatest benefit from enfuvirtide; however, patients with advanced disease (viral load >100,000 copies per ml and CD4 cell count <100 cells per mm3) also demonstrated significant virological and immunological response. These findings confirmed earlier analyses based on the TORO 1 and TORO 2 24-week database, in which patients receiving enfuvirtide plus OB showed greater virological and immunological improvement across all subgroups compared with patients receiving OB alone41,42.
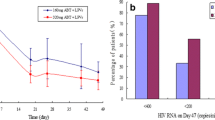
The TORO patient population had broad treatment experience with a median prior exposure to 12 ARVs and seven years of therapy. Viral suppression was significantly greater with enfuvirtide plus OB treatment than with OB treatment alone across all subgroups when classified according to the number of active ARVs in the background regimen, including those patients with no active ARVs in their background regimen40. Response in both treatment groups improved with increasing numbers of active drugs in the background regimen (Fig. 6), and response to enfuvirtide plus OB in patients with one active ARV in their background regimen was similar to, or greater than, response to OB alone in patients with two, three, four or five active ARVs in the background regimen. Not surprisingly, patients who had been previously exposed to fewer ARVs also responded better to enfuvirtide plus OB, presumably an indication of less advanced disease and a reflection of the increased options that these patients had to include fully active ARVs in their background regimen.

Percent responders with HIV-1 RNA <400 copies per ml, week 48 (intent-to-treat, discontinuation or virological failure = failure). GSS is the actual number of antiretrovirals that the baseline virus is sensitive to as indicated by standard primary mutations that each virus possesses. ENF, enfuvirtide; OB, optimized treatment background.
A simplified model investigating prognostic factors that might predict response to ARV therapy including enfuvirtide has identified four factors that are associated with greater response to enfuvirtide: first, CD4 count >100 cells per mm3; second, HIV-1 RNA <100,000 copies per ml; third, experience of ≤10 prior ARVs; and last, combining enfuvirtide with at least two active ARVs43. For patients that have all four positive prognostic factors, the chance of achieving a viral load <400 copies per ml at week 24 when receiving enfuvirtide plus OB is 80%, compared with 50% for patients receiving OB alone43. For the treating physician, this provides valuable insight and suggests that although enfuvirtide will benefit all patients, earlier use in the treatment algorithm, combined with other fully active ARVs, can provide the greatest benefit.
Virological failure. Median time to protocol-defined failure during 48 weeks of therapy was three times as long in the enfuvirtide group (32 weeks) as in the control group (11 weeks, P <0.0001)38. Patients who were characterized as failing therapy were more likely to be on the OB arm (71% versus 46%). Patients that failed over the course of the trials tended to have fewer active drugs in their background regimen, lower baseline CD4 cell counts and had been exposed to more ARVs before enrolling in the TORO trials44.
Resistance to enfuvirtide
Although enfuvirtide is active in patients whose virus is resistant to the other classes of ARVs, as is the case with all other antiretrovirals, continued exposure, especially if the virus is not completely suppressed, can lead to emergence of viral resistance to enfuvirtide.
In vitro studies. The N-terminal region of gp41 was evaluated with laboratory and clinical viral isolates with reduced susceptibility to enfuvirtide. Rimsky et al. identified susceptibility loci by SERIAL PASSAGE of viral strains HIV-1IIIB and NL4-3 in increasing concentrations of enfuvirtide45. All escape mutants mapped to the HR1, N-terminus of gp41 and in particular to a contiguous sequence within the heptad-repeat motif. This sequence is a deep cavity within the HR1 trimer and spanned positions 32–41 within the HR1 region of gp41, with positions 36 (G36D) and 38 (V38M) being most sensitive46,47.
Resistance to enfuvirtide in clinical studies. An early clinical trial exposed patients to short courses of enfuvirtide monotherapy to evaluate dose–response relationships23. Viral isolates were sequenced by amplifying the HR1 and HR2 regions of gp41 (Ref. 48). The lowest (10 mg twice daily) and highest (100 mg twice daily) doses of enfuvirtide did not select for resistance, but virus isolated during exposure to, and rebound from, intermediate doses showed amino-acid substitutions within the HR1 region. Amino-acid substitutions occurred at gp41 positions 32 (Q to R/H), 36 (G to S), 38 (V to A) and 39 (Q to R). Subsequent analysis of sequences derived from clinical isolates continued to highlight the crucial core region of residues 36–45 within HR1 (Refs 49,50).
The evolution of resistance to enfuvirtide at amino-acid positions 36–45 was analysed from virus isolated before and during treatment51. At baseline, virus from 177 enfuvirtide-naive patients was sequenced with 80% showing the consensus sequence at positions 36–45, and 15% of the viral isolates showing an N42S polymorphism. The remaining 5% had substitutions other than N42S. At protocol-defined virological failure, the virus from 78% (31 out of 40) of patients had substitutions associated with resistance at positions 36–45. Investigation of the replicative capacity of virus with enfuvirtide mutations confirmed that enfuvirtide-resistance mutations in HR1 identified in patient isolates obtained after treatment with enfuvirtide conferred a significant growth disadvantage in the absence of drug as compared with wild type52,53.
Viral preference for chemokine co-receptor varies with the phase of HIV infection; during acute infection, CCR5-tropic HIV variants predominate, whereas CXCR4-tropic variants emerge during later stages of disease. In vitro evidence from laboratory virus and from a small number of clinical isolates indicated that sensitivity to enfuvirtide might be influenced by chemokine co-receptor TROPISM54,55,56. Enfuvirtide clinical trials provided the opportunity to examine a larger number of clinical isolates that expressed tropism for dual or individual co-receptors. Using several cell-based assays, Greenberg et al. evaluated co-receptor phenotype and syncytium formation of 151 clinical trial isolates and 8 isolates from the National Institute of Allergy and Infectious Disease Repository. CCR5-, CXCR4- and dual-tropic virus exhibited similar sensitivities57. Whitcomb et al. examined 612 baseline clinical-trial isolates using a recombinant assay to target cells that expressed CD4 and either CCR5 or CXCR4 co-receptors58. CCR5 isolates comprised 62% of the population, 34% of isolates were dual-tropic and only 4% of baseline isolates used CXCR4. The geometric mean IC50 was 44 ng per ml for CXCR4 and 25 ng per ml for CCR5 isolates. Although the IC50 for CXCR4 isolates was slightly higher than for the other two types, mean virological response (change from baseline) to enfuvirtide at 24 weeks (ITT, last observation carried forward (LOCF)) was generally similar among patients who harboured CCR5 (−1.55 log10 copies per ml), dual tropic (−1.64 log10 copies per ml) or CXCR4 (−1.34 log10 copies per ml) viruses at baseline (P = 0.4)59. These data would therefore indicate that viral tropism has no clinically relevant effect on treatment response to an enfuvirtide-containing regimen.
The clinical safety of enfuvirtide
As enfuvirtide entered the clinic, several safety questions were foremost: what would be the overall safety profile in treatment-experienced patients on various combinations of ARVs?; what would be the consequences of chronic, twice-daily injections?; as a foreign peptide, would enfuvirtide cause local inflammatory or more serious immunological reactions?; and last, would the human immune system eventually reject enfuvirtide? Analysis of clinical data from Phase II and the Phase III TORO trials addressed these questions.
Overall surveillance during the TORO trials included 663 patients randomized to enfuvirtide plus OB and 334 patients randomized to OB alone. The combined safety record of the TORO trials through 24 weeks has been described37. During the 48 weeks, 26.5% of patients in the enfuvirtide group and 36.6% in the control group discontinued therapy. Discontinuation due to adverse events occurred in 8.9% of patients in the enfuvirtide group and 3.6% of patients in the control group. Injection-site reactions (ISRs) were tabulated separately and accounted for a discontinuation rate of 4.4%.
During the 48-week period, the exposure to enfuvirtide plus OB, switch, combined enfuvirtide [(enfuvirtide plus OB) plus switch] and OB was 557, 120, 677 and 162 patient years, respectively. The most commonly reported events were diarrhoea (37.1 per 100 patient-years on combined enfuvirtide versus 73.4 on OB), nausea (26.2 versus 50.0) and fatigue (25.0 versus 37.6)60. Other common constitutional signs/symptoms reported at a lower frequency on combined enfuvirtide than on OB were headache (15.8 per 100 patient years on combined enfuvirtide versus 24.1 on OB), insomnia (16.6 versus 19.7) and vomiting (15.8 versus 26.5). Among the events reported more commonly on the combined enfuvirtide arms were peripheral neuropathy (16.3 per 100 patient years on combined enfuvirtide versus 13.6 on OB), weight decrease (12.1 versus 10.5), sinusitis (9.3 versus 6.2), appetite decrease (8.3 versus 4.9) and lymphadenopathy (7.1 versus 1.2). An imbalance in the rate of pneumonia, primarily bacterial, was also observed at 48 weeks. In the combined enfuvirtide group, 6.7 patients per 100 patient years showed pneumonia, in contrast to 0.6 patients per 100 patient years in the control group60. Of note, the rate observed in the enfuvirtide arm was within ranges reported for this patient population in the literature. A review of data from the Adult Spectrum of Disease study (Seattle) indicated that in the post-HAART-era pneumonia rates have declined, but are still in the range seen on the enfuvirtide arm in the TORO studies and that the risk of pneumonia is closely linked with CD4 count.
Clinical laboratory findings identified only increased eosinophila (>0.7 × 109 cells per l) as imbalanced between treatment groups, occurring in 11.1 per 100 patient years in combined enfuvirtide patients and 3.7 per 100 patient years in OB patients. However, when using the higher (>1.4 × 109 per l) threshold for marked eosinophilia, similar rates per 100 patient years were seen for both treatment groups. Eosinophila was not associated with hypersensitivity in either treatment group. Overall, Grade 4 treatment-emergent laboratory abnormalities were seen in 15.5 per 100 patient years of combined enfuvirtide patients and in 25.9 per 100 patient years of patients on OB regimen alone.
Throughout 48 weeks, almost all patients (98.3%) receiving enfuvirtide cumulatively reported ISRs. At any given visit, 46–53% of patients reported mild tenderness, 15–20% moderate pain without limitation of usual activities and 0.9–3.3% severe pain requiring analgesics or limiting usual activities. The most frequent signs and symptoms of a local ISR were pain/discomfort (96%), erythema (91%), induration (90%) and nodules and cysts (80%)60. Importantly, there was no evidence of an increase in severity over time for any of the signs and symptoms of a local ISR and few patients experienced infection at the injection site.
Several histopathology and histochemistry studies have been conducted to help elucidate possible mechanisms of ISRs. Ball and Kinchelow61 reported on excisional biopsy findings from seven patients with ISRs in enfuvirtide clinical trials. Although responses varied, inflammatory infiltrates were found that were consistent with a localized reaction. Conspicuous eosinophils and histocytes were present, but lymphocytes and plasma cells were rare. Multinucleated forms occurred that in two patients were similar to a granulomatous response. There was no relationship between inflammatory response and clinical findings.
Questions of antibody neutralization of enfuvirtide and induction of inflammatory responses or other adverse events have been addressed40,62. Serum samples for assaying of anti-gp41 antibody that crossreacted to enfuvirtide were taken during the TORO trials. Across both treatment arms, approximately 75% of patients were positive for anti-gp41 antibody and 23% of patients had non-quantifiable levels of antibody at baseline; less than 2% of patients were antibody-negative at baseline. At 24 weeks, 65% of enfuvirtide patients and 13% of OB patients had decreases of 30% or more in antibody levels.
The presence of anti-gp41 antibody did not seem to influence the activity of enfuvirtide. Mean change from baseline in HIV-1 RNA at 24 weeks was identical among enfuvirtide groups with negative, non-quantifiable and positive antibody levels (1.6 log10 copies per ml) and was in a narrow range (0.6–0.8 log10 copies per ml) in the OB group. Rates of adverse events, hypersensitivity reactions and Grade 3 or Grade 4 laboratory toxicities were comparable, regardless of baseline or on-treatment antibody status or magnitude of antibody level.
Quality of life
The TORO studies included a Subcutaneous Injection Survey (SIS) to evaluate patient attitude to self-injection63,64. Patients were surveyed at weeks 8, 24 and 48. The majority of patients assessed all items relating to ease of injection as 'very easy' or 'easy' and assessed their daily functioning as 'not at all' or 'a little' limited by enfuvirtide self-injection. The results of the SIS indicated that most patients with appropriate education and training were able to learn to integrate enfuvirtide dosing into their daily routines. Patients were also evaluated using the Medical Outcomes Study (MOS)-HIV questionnaire65. This is a well-established and comprehensive measure of Heath Related Quality of Life (HRQoL). Patients were evaluated at 24 and 48 weeks. The results of the MOS-HIV showed that for most parameters evaluated there was a slight improvement in HRQoL during the 48 weeks of therapy in both patients receiving enfuvirtide plus OB and those receiving OB alone65.
The future of entry inhibitors
Enfuvirtide ushered in a new class of ARVs that can suppress multidrug-resistant virus by virtue of a distinct mechanism of action. Entry inhibitors also have the potential for marked synergism with, and lack of cross-resistance to, NRTIs, nNRTIs and PIs66. Each step in the entry process is a potential target, and there are a number of compounds in development directed against each step that show promise, some of which are now in clinical trials. The initial step in HIV entry is the recognition and binding of viral gp120 to host CD4 receptors, an interaction that is the target of several small molecules67, monoclonal antibodies68 and recombinant molecules that link CD4 receptors to IgG2 constant regions69,70,71,72,73. The CD4-IgG2 tetramer seems to be highly synergistic in vitro with enfuvirtide73. A single-dose, dose-escalation study of the intravenously administered monoclonal antibody TNX-355 demonstrated antiviral activity at all doses68.
CCR5 and CXCR4 co-receptor inhibitors have also been intensely investigated. CCR5 inhibitors include RANTES antagonists and other small molecules, some of which have already demonstrated early clinical activity and also in vitro synergism with enfuvirtide. The TAK series has good in vitro activity74,75 and synergism with enfuvirtide has been reported76,77. A monoclonal antibody (PRO-140; Progenics) inhibits viral entry without influencing CCR5 function in vitro, and antiviral activity was seen in a therapeutic mouse model78. The RANTES antagonist SCH-C (Schering-Plough) performed well in initial dose-range trials56. SCH-C is synergistic with enfuvirtide79, and along with enfuvirtide is active in multiple cell types, including peripheral blood mononuclear cells, dendritic cells and macrophages80. To date, escape mutants do not seem to switch to a CXCR4 phenotype81,82. The development of inhibitors of CXCR4 binding has been somewhat less successful and there is concern about the consequences of blocking a receptor that is widely present in different cell types66. Results from a clinical study with the small molecule AMD-3100 (AnorMed) did not show consistent antiviral activity83.
Similarly to enfuvirtide, T-1249 is a peptide fusion inhibitor that interacts with the gp41 HR1 region of HIV-1. Its sequence overlaps with, but is not identical to, enfuvirtide and contains short sequences derived from SIV and HIV-2. Owing to these differences, T-1249 is active against laboratory and clinical viral isolates that are resistant to enfuvirtide84. Although not presently in clinical development, the results of a dose-ranging trial in 115 patients during a 14-day period demonstrated dose-dependent viral suppression with a median HIV-1 RNA suppression of −1.96 log10 copies per ml at the highest dose (200 mg per day) during monotherapy. No dose-limiting toxicity occurred85. Furthermore, T-1249 activity was not affected by ARV resistance or prior treatment history86,87.
Discussion
Enfuvirtide is the first ARV fusion inhibitor approved for the treatment of HIV-1-infected patients. It is by far the most complex ARV ever manufactured at such a large scale, which provided both production and economic challenges.
The target population for enfuvirtide is treatment-experienced patients with limited remaining therapeutic options. This population is steadily rising as increasing numbers of patients progress through the treatment algorithm. The most gratifying aspect of the development of enfuvirtide is the hope it has given to these individuals, who are some of the most difficult to treat HIV-infected patients.
The subgroup analyses uncovered clues to the optimal use of enfuvirtide and the management of treatment-experienced patients. Regardless of baseline status, all patient groups benefited from enfuvirtide therapy. Multiple- and logistic-regression analyses identified four key prognostic factors for virological and immunological response: lower baseline viral load, higher baseline CD4, more active ARVs in the OB and a fewer number of prior ARVs. Although recognition of such patterns might also be thought of as common-sense logic in treating HIV patients, it takes on additional gravity when demonstrated for heavily treatment-experienced patients and provides confidence when devising treatment strategies.
The TORO studies succeeded with the utilization of state-of-the-art patient management, incorporating both physician and nursing support. An often overlooked finding is that patients receiving the background therapy alone demonstrated an uncharacteristically good clinical response. This probably reflects the use of antiviral-resistance testing before study treatment. Most patients could construct an OB regimen with at least partial activity. The fact that the addition of enfuvirtide to this OB regimen significantly enhanced the response in this patient population highlights the additional benefits of adding an ARV agent from a new class.
Obviously at the start of the enfuvirtide clinical programme the impact of chronic daily injections was a major concern; injection burden, the routine of assembling supplies and preparing and administering injections could have affected the utility of enfuvirtide therapy. The experience of study patients for as long as four years indicates that injection burden or fatigue was not insurmountable and did not unduly affect activities of daily living.
The synergistic effects of investigational ARVs, such as CCR5 inhibitors with enfuvirtide, are now being investigated. Long-term data from these studies will be important in determining future treatment strategies and, once again, prolonging and enhancing the life of patients infected with HIV.
References
King, J. T. Jr et al. Long-term HIV/AIDS survival estimation in the highly active antiretroviral therapy era. Med. Decis. Making 23, 9–20 (2003).
Kitchen, C. M. et al. Initial virological and immunologic response to highly active antiretroviral therapy predicts long-term clinical outcome. Clin. Infect. Dis. 33, 466–472 (2001).
Valenti, W. M. HAART is cost-effective and improves outcomes. AIDS Read. 11, 260–262 (2001).
Richman, D. D. et al. The prevalence of antiretroviral drug resistance in the US. 41st Intersci. Conf. Antimicrob. Agents Chemother. A LB–17 (2001).
Little, S. E. et al. Antiretroviral-drug resistance among patients recently infected with HIV. N. Engl. J. Med. 347, 385–394 (2002). A retrospective analysis showing that the proportion of new HIV infections that involve drug-resistant virus is increasing in North America.
Hammer, S. M. HIV drug resistance: implications for management. Top. HIV Med. 10, 10–15 (2002).
Kuritzkes, D. R. et al. Current management challenges in HIV: antiretroviral resistance. AIDS Read. 13, 133–142 (2003).
Montaner, J. S. & Mellors, J. W. Antiretroviral therapy for previously treated patients. N. Engl. J. Med. 345, 452–455 (2001).
Richman, D. D. HIV chemotherapy. Nature 410, 995–1001 (2001).
Eckert, D. M. & Kim, P. S. Mechanisms of viral membrane fusion and its inhibition. Annu. Rev. Biochem. 70, 777–810 (2001).
LaBranche, C. C. et al. HIV fusion and its inhibition. Antiviral Res. 50, 95–115 (2001).
Einfeld, D. & Hunter, E. Oligomeric structure of a prototype retrovirus glycoprotein. Proc. Natl Acad. Sci. USA 85, 8688–8692 (1988).
Pinter, A. et al. Oligomeric structure of gp41, the transmembrane protein of human immunodeficiency virus type 1. J. Virol. 63, 2674–2679 (1989).
Wyatt, R. & Sodroski, J. The HIV-1 envelope glycoproteins: fusogens, antigens, and immunogens. Science 280, 1884–1888 (1998).
Munoz-Barroso, I. et al. Dilation of the human immunodeficiency virus-1 envelope glycoprotein fusion pore revealed by the inhibitory action of a synthetic peptide from gp41. J. Cell. Biol. 140, 315–323 (1998).
Matthews, T. J. et al. Structural rearrangements in the transmembrane glycoprotein after receptor binding. Immunol. Rev. 140, 93–104 (1994).
Wild, C. et al. A synthetic peptide inhibitor of human immunodeficiency virus replication: correlation between solution structure and viral inhibition. Proc. Natl Acad. Sci. USA 89, 10537–10541 (1992).
Wild, C. et al. Propensity for a leucine zipper-like domain of human immunodeficiency virus type 1 gp41 to form oligomers correlates with a role in virus-induced fusion rather than assembly of the glycoprotein complex. Proc. Natl Acad. Sci. USA 91, 12676–12680 (1994).
Wild, C., Greenwell, T. & Matthews T. A synthetic peptide from HIV-1 gp41 is a potent inhibitor of virus-mediated cell-cell fusion. AIDS Res. Hum. Retroviruses 9, 1051–1053 (1993). First report showing that the synthetic peptide DP-178, later to be called enfuvirtide, was a potent inhibitor of virus infection and virus-mediated cell-to-cell fusion.
Wild, C. T. et al. Peptides corresponding to a predictive α-helical domain of human immunodeficiency virus type 1 gp41 are potent inhibitors of virus infection. Proc. Natl Acad. Sci. USA 91, 9770–9774 (1994).
Chen, C. H. et al. A molecular clasp in the human immunodeficiency virus (HIV) type 1 TM protein determines the anti-HIV activity of gp41 derivatives: implication for viral fusion. J. Virol. 69, 3771–3777 (1995).
Kliger, Y. et al. Mode of action of an antiviral peptide from HIV-1. J. Biol. Chem. 276, 1391–1397 (2001).
Kilby, J. M. et al. Potent suppression of HIV-1 replication in humans by T-20, a peptide inhibitor of gp41-mediated virus entry. Nature Med. 4, 1302–1307 (1998). This study (TRI-001) provided proof-of-concept that viral entry could be successfully blocked in vivo.
Kilby, J. M. et al. The safety, plasma pharmacokinetics, and antiviral activity of subcutaneous enfuvirtide (T-20), a peptide inhibitor of gp41-mediated virus fusion, in HIV-infected adults. AIDS Res. Hum. Retroviruses 18, 685–693 (2002).
Sista, P. et al. Subgroup analysis of baseline (BL) susceptibility and early virological response to enfuvirtide in the combined TORO studies. Antivir. Ther. 8, S60 (2003).
Lalezari, J. P. et al. A phase II clinical study of the long-term safety and antiviral activity of enfuvirtide-based antiretroviral therapy. AIDS 17, 691–698 (2003).
Lalezari, J. P. et al. A controlled Phase II trial assessing three doses of enfuvirtide (T-20) in combination with abacavir, amprenavir, ritonavir and efavirenz in non-nucleoside reverse transcriptase inhibitor-naive HIV-infected adults. Antivir. Ther. 8, 279–287 (2003).
Wheeler, D. A. et al. Safety, tolerability and plasma pharmacokinetics of high-strength formulations of enfuvirtide (T-20) in treatment-experienced HIV-1-infected patients. J. Clin. Virol. (in the press).
Drobnes, C. et al. Tolerability of enfuvirtide (T-20) during chronic therapy in Phase II trials. 42nd Intersci. Conf. Antimicrob. Agents Chemother. A H-171 (2002).
Lalezari, J. P. et al. Influence of subcutaneous injection site on the steady state pharmacokinetics of enfuvirtide (T-20) in HIV-1-infected patients. J. Clin. Virol. 28, 217–222 (2003).
Zhang, X. et al. Pharmacokinetics of plasma enfuvirtide after subcutaneous administration to patients with human immunodeficiency virus: inverse Gaussian density absorption and 2-compartment disposition. Clin. Pharmacol. Ther. 72, 10–19 (2002).
Zhang, X. et al. Assessment of metabolic inhibition potential of enfuvirtide using a 5-drug cocktail in HIV-1 infected patients. Am. Soc. Clin. Pharmacol. Ther. A PIII–73 (2003).
Boyd, M. et al. Enfuvirtide — investigations on the drug interaction potential in HIV-infected patients. 10th Conf. Retrovir. Opportunistic Infect. A541 (2003).
Zhang, X. et al. Enfuvirtide pharmacokinetic-pharmacodynamic (PK-PD) relationship. 9th Eur. AIDS Conf. A4.1/4 (2003).
Lalezari, J. P. et al. Enfuvirtide, an HIV-1 fusion inhibitor, for drug-resistant HIV infection in North and South America. N. Engl. J. Med. 348, 2175–2185 (2003). The 24-week efficacy and safety results from the pivotal Phase III study set in North America and Brazil, TORO 1.
Lazzarin, A. et al. Efficacy of enfuvirtide in patients infected with drug-resistant HIV-1 in Europe and Australia. N. Engl. J. Med. 348, 2186–2195 (2003). The 24-week efficacy and safety results from the pivotal Phase III study set in Europe and Australia, TORO 2.
Delfraissy, J. F. et al. Summary of pooled efficacy and safety analyses of enfuvirtide (ENF) treatment for 24 weeks in TORO 1 and TORO 2 Phase III trials in highly antiretroviral (ARV) treatment-experienced patients. 10th Conf. Retrovir. Opportunistic Infect. A568 (2003).
Katlama, C. et al. Enfuvirtide TORO studies: 48 week results confirm 24 week findings. 2nd Int. AIDS Soc. Conf. HIV Pathog. Treat. A LB2 (2003). Presentation of the integrated TORO 1 and TORO 2 efficacy and safety results at 48 weeks, demonstrating that enfuvirtide therapy is durable and tolerable.
Trottier, B. et al. Durability of response of enfuvirtide through 48 weeks in the TORO trials. 43rd Intersci. Conf. Antimicrob. Agents Chemother. A H-835 (2003).
Walmsley, S. et al. Efficacy of enfuvirtide in subgroups of patients through 48 weeks of therapy in the TORO trials. 9th Eur. AIDS Conf. A7.3/15 (2003). Presentation reporting that enfuvirtide was effective across all subgroups based on demographics, baseline characteristics as well as number of active antiretrovirals in the background regimen, as evaluated at 48 weeks.
Lalezari, J. et al. Enfuvirtide (T-20) in combination with an optimized background (OB) regimen vs. OB alone: week 24 response among categories of treatment experience and baseline (BL) HIV antiretroviral (ARV) resistance. 42nd Intersci. Conf. Antimicrob. Agents Chemother. A3232 (2002).
Lange, J. et al. Enfuvirtide (T-20) in combination with an optimized background (OB) regimen vs. OB alone: week 24 response among categories of treatment experience and baseline (BL) demographics, treatment experience and HIV anti-retroviral (ARV) resistance. 6th Int. Congress Drug Ther. HIV Infect. A PL14.3 (2002).
Montaner, J. et al. Analysis of virological response to enfuvirtide in TORO: implications for patient management. 2nd Intern AIDS Soc. Conf. HIV Pathog. Treat. A116 (2003).
Battegay, M. et al. Analysis of virological failure through 24 weeks of treatment in the TORO pivotal studies. 2nd Int. AIDS Soc. Conf. HIV Pathog. Treat. A574 (2003).
Rimsky, L. T., Shugars, D. C. & Matthews, T. J. Determinants of human immunodeficiency virus type 1 resistance to gp41-derived inhibitory peptides. J. Virol. 72, 986–993 (1998). These experiments demonstrated that the mechanism of action of enfuvirtide was binding to the HR1 region of gp41 and inhibition of the HR1-HR2 complex formation within the gp41 protein.
Chan, D. C. et al. Core structure of gp41 from the HIV envelope glycoprotein. Cell 89, 263–273 (1997).
Weissenhorn, W. et al. Atomic structure of the ectodomain from HIV-1 gp41. Nature 387, 426–430 (1997).
Wei, X. et al. HIV-1 selection in response to inhibition of virus fusion and entry. 6th Conf. Retrovir. Opportunistic Infect. A611 (1999).
Mink, M. et al. Impact of HIV-1 gp41 amino acid substitutions (positions 36-45) on susceptibility of T-20 (enfuvirtide) in vitro: analysis of primary virus isolates recovered from patients during chronic enfuvirtide treatment and site-directed mutants in NL4-3. Antivir. Ther. 7, S24 (2002).
Mink, M. A. et al. Correlation of gp41 binding and antiviral potencies of the T-20 fusion inhibitor using clinical trial isolate-derived sequences. 8th Conf. Retrovir. Opportunistic Infect. A474 (2001).
Sista, P. et al. Characterization of baseline and treatment-emergent resistance to T-20 (enfuvirtide) observed in Phase II trials: substitutions in gp41 amino acids 36-45 and enfuvirtide susceptibility of virus isolates. Antivir. Ther. 7, S16 (2002).
Lu, J. & Kuritzkes, D. R. Impaired fitness of human immunodeficiency virus type 1 site-directed mutants resistant to T-20. 5th Int. Workshop HIV Drug Resist. Treat. Strateg. A23 (2001).
Lu, J. et al. Fitness of HIV-1 clinical isolates resistant to T-20 (enfuvirtide). Antivir. Ther. 7, S74 (2002).
Derdeyn, C. A. et al. Sensitivity of human immunodeficiency virus type 1 to fusion inhibitors targeted to the gp41 first heptad repeat involves distinct regions of gp41 and is consistently modulated by gp120 interactions with the coreceptor. J. Virol. 75, 8605–8614 (2001).
Derdeyn, C. A. et al. Sensitivity of HIV-1 to the fusion inhibitors T-20 and T-649 is modulated by coreceptor specificity and involves distinct regions of gp41. 8th Conf. Retrovir. Opportunistic Infect. A475 (2001).
Reynes, J. et al. SCH C: safety and antiviral effects of a CCR5 receptor antagonist in HIV-1 infected subjects. 9th Conf. Retrovir. Opportunistic Infect. A1 (2002).
Greenberg, M. L. et al. Virus sensitivity to T-20 and T-1249 is independent of coreceptor usage. 8th Conf. Retrovir. Opportunistic Infect. A473 (2001).
Whitcomb, J. M. et al. Analysis of baseline enfuvirtide (T20) susceptibility and co-receptor tropism in two Phase III study populations. 10th Conf. Retrovir. Opportunistic Infect. A557 (2003).
Greenberg, M. et al. Baseline and on-treatment susceptibility to enfuvirtide seen in TORO 1 and TORO 2 through 24 weeks. 10th Conf. Retrovir. Opportunistic Infect. A141 (2003).
Eron, J. et al. Safety of enfuvirtide (ENF) through 48 weeks of therapy in the TORO trials. 43rd Intersci. Conf. Antimicrob. Agents Chemother. A H-836 (2003).
Ball, R. A. & Kinchelow, T. Injection site reactions with the HIV-1 fusion inhibitor enfuvirtide. J. Am. Acad. Dermatol. 49, 826–831 (2003).
Walmsley, S. et al. Enfuvirtide (T-20) cross-reactive gp41 antibody has no effect on the efficacy or safety of enfuvirtide in the TORO 1 and TORO 2 Phase III trials. J. Infect. Dis. 188, 1827–1833 (2003).
Cohen, C. et al. Patient acceptance of self-injected enfuvirtide at 8 and 24 weeks. HIV Clin. Trials 4, 347–357 (2003).
Cohen, C. et al. Patient acceptance with self-injection of enfuvirtide (ENF) for HIV over 48 weeks of treatment. 9th Eur. AIDS Conf. A7.1/1 (2003).
Clumeck, N. et al. Impact of enfuvirtide on health-related quality of life at 48 weeks. 9th Eur. AIDS Conf. A 7.3/19 (2003).
Kilby, J. M. & Eron, J. J. Novel therapies based on mechanisms of HIV-1 cell entry. N. Engl. J. Med. 348, 2228–2238 (2003).
Lin, P. F. et al. Identification and characterization of a novel inhibitor of HIV-1 entry - II: mechanism of action. 9th Conf. Retrovir. Opportunistic Infect. A10 (2002).
Kuritzkes, D. R. et al. Safety and preliminary anti-HIV activity of an anti-CD4 mAb (TNX-355; formerly Hu5A8) in HIV-infected patients. 10th Conf. Retrovir. Opportunistic Infect. A13 (2003).
Franti, M. et al. PRO 542 (CD4-IgG2) has a profound impact on HIV-1 replication in the Hu-PBL-SCID mouse model. 9th Conf. Retrovir. Opportunistic Infect. A410-T (2002).
Jacobson, J. M. et al. Single-dose safety, pharmacology, and antiviral activity of the human immunodeficiency virus (HIV) type 1 entry inhibitor PRO 542 in HIV-infected adults. J. Infect. Dis. 182, 326–329 (2000).
Nagashima, K. et al. Mechanisms of synergy between HIV-1 attachment, coreceptor and fusion inhibitors. 8th Conf. Retrovir. Opportunistic Infect. A310 (2001).
Olson, W. C. et al. Potent, synergistic inhibition of HIV-1 by combinations of the viral entry inhibitors PRO-542 and T-20. 40th Intersci. Conf. Antimicrob. Agents Chemother. A549 (2000).
Trkola, A. et al. Cross-clade neutralization of primary isolates of human immunodeficiency virus type 1 by human monoclonal antibodies and tetrameric CD4-IgG. J. Virol. 69, 6609–6617 (1995).
Dragic, T. et al. A binding pocket for a small molecule inhibitor of HIV-1 entry within the transmembrane helices of CCR5. Proc. Natl Acad. Sci. USA 97, 5639–5644 (2000).
Takashima, K. et al. Inhibitory effects of small-molecule CCR5 antagonists on human immunodeficiency virus type 1 envelope-mediated membrane fusion and viral replication. Antimicrob. Agents Chemother. 45, 3538–3543 (2001).
Tremblay, C. L. et al. In vitro synergy observed between the fusion inhibitor T-20 and a CCR5 inhibitor TAK-779. 40th Intersci. Conf. Antimicrob. Agents Chemother. A1164 (2000).
Tremblay, C. L. et al. TAK-220, a novel small molecular inhibitor of CCR5 has favorable anti-HIV interactions with other antiretrovirals in vitro. 10th Conf. Retrovir. Opportunistic Infect. A562 (2003).
Franti, M. et al. The CCR5 co-receptor inhibitor PRO 140 effectively controls established HIV-1 infections in vivo. 9th Conf. Retrovir. Opportunistic Infect. A403-T (2002).
Tremblay, C. L. et al. Anti-human immunodeficiency virus interactions of SCH-C (SCH 351125), a CCR5 antagonist, with other antiretroviral agents in vitro. Antimicrob. Agents Chemother. 46, 1336–1339 (2002).
Ketas, T. J. et al. Entry inhibitors SCH-C, RANTES, and T-20 block HIV type 1 replication in multiple cell types. AIDS Res. Hum. Retroviruses 19, 177–186 (2003).
Riley, J. et al. Genotypic and phenotypic analysis of in vitro generated HIV-1 escape isolates to the CCR5 antagonist SCH-C. 9th Conf. Retrovir. Opportunistic Infect. A397-T (2002).
Stoddart, C. et al. Evaluation of in vivo HIV-1 escape from SCH-C (SCH 351125). 10th Conf. Retrovir. Opportunistic Infect. A614 (2003).
Tremblay, C. et al. Strong in vitro synergy observed between the fusion inhibitor T-20 and a CXCR4 blocker AMD-3100. 7th Conf. Retrovir. Opportunistic Infect. A500 (2000)
Greenberg, M. L. et al. Enfuvirtide (T-20) and T-1249 resistance: observations from Phase II clinical trials of enfuvirtide in combination with oral antiretrovirals (ARVs) and a Phase I/II dose-ranging monotherapy trial of T-1249. 11th Int. HIV Drug Resist. Workshop. A128 (2002).
Gulick, R. et al. Complete analysis of T1249-101: safety, pharmacokinetics, and antiviral activity of T-1249, a peptide inhibitor of HIV membrane fusion. 42nd Intersci. Conf. Antimicrob. Agents Chemother. A850 (2002).
Miralles, G. D. et al. Baseline genotype and prior antiretroviral history do not affect virological response to T-1249. 5th Int. Workshop HIV Drug Resist. Treat. Strateg. A3 (2001).
Miralles, G. D. et al. Genotypic resistance to protease and reverse transcriptase inhibitors and antiretroviral history do not affect virologic response to T-1249. 41st Intersci. Conf. Antimicrob. Agents Chemother. A669 (2001).
De Clercq, E. Strategies in the design of antiviral drugs. Nature Rev. Drug Disc. 1, 13–25 (2002).
Acknowledgements
The authors would like to thank the hundreds of people involved in the discovery and development of enfuvirtide; the Roche and Trimeris Research and Development Teams, the Study Investigators and the many patients who participated in the clinical trials.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
M.S. and J.C. are employees of Roche Laboratories, a division of F. Hoffmann-La Roche Ltd. T.M., M.G., R.M. and D.B. are employees of Trimeris, Inc. Trimeris and Roche are collaborative partners in develo** enfuvirtide (Fuzeon).
Related links
Related links
DATABASES
LocusLink
FURTHER INFORMATION
Encyclopedia of Life Sciences
AIDS: understanding HIV transmission
Glossary
- LYMPHOCYTES
-
White blood cells of lymphoid origin that function as part of the immune system.
- SYNCYTIUM
-
A multi-nucleated mass caused by the fusion of several individual cells; often virally induced.
- INTENT TO TREAT
-
A conservative form of clinical trial analysis in which the numerator is the number of patients in a given arm achieving an end-point and the denominator is the total number of patients assigned to that arm, with discontinuations and/or missing data classed as failures.
- SERIAL PASSAGE
-
Innoculation of a chain of in vitro cell cultures with samples of an infectious agent drawn from the previously infected culture.
- TROPISM
-
The affinity of a virus for a particular biological feature that affects its infectivity or growth.
Rights and permissions
About this article
Cite this article
Matthews, T., Salgo, M., Greenberg, M. et al. Enfuvirtide: the first therapy to inhibit the entry of HIV-1 into host CD4 lymphocytes. Nat Rev Drug Discov 3, 215–225 (2004). https://doi.org/10.1038/nrd1331
Issue Date:
DOI: https://doi.org/10.1038/nrd1331
- Springer Nature Limited
This article is cited by
-
Porphyrins with combinations of 4-carboxyphenyl and 4-hydroxyphenyl substituents in meso-positions as anti-HIV-1 agents
Scientific Reports (2024)
-
A stapled lipopeptide platform for preventing and treating highly pathogenic viruses of pandemic potential
Nature Communications (2024)
-
Inactivation of cell-free HIV-1 by designing potent peptides based on mutations in the CD4 binding site
Medical & Biological Engineering & Computing (2024)
-
Strategies to improve the physicochemical properties of peptide-based drugs
Pharmaceutical Research (2023)
-
Development of a HPLC fluorometric method for the quantification of enfuvirtide following in vitro releasing studies on thermosensitive in situ forming gel
Drug Delivery and Translational Research (2023)