
Abstract
The immunoglobulin E (IgE)-associated locus on human chromosome 13q14 influencing asthma-related traits contains the genes PHF11 and SETDB2. SETDB2 is located in the same linkage disequilibrium region as PHF11 and polymorphisms within SETDB2 have been shown to associate with total serum IgE levels. In this report, we sequenced the 15 exons of SETDB2 and identified a single previously ungenotyped mutation (AT/G, rs386770867) in the 5′-untranslated region of the gene. The polymorphism was found to be significantly associated with serum IgE levels in our asthma cohort (P=0.0012). Electrophoretic mobility shift assays revealed that the transcription factor Ying Yang 1 binds to the AT allele, whereas SRY (Sex determining Region Y) binds to the G allele. Allele-specific transcription analysis (alleloty**) was performed in 35 individuals heterozygous for rs386770867 from a panel of 200 British families ascertained through probands with severe stage 3 asthma. The AT allele was found to be significantly overexpressed in these individuals (P=1.26 × 10−21). A dual-luciferase assay with the pGL3 luciferase reporter gene showed that the AT allele significantly affects transcriptional activities. Our results indicate that the IgE-associated AT/G polymorphism (rs386770867) regulates transcription of SETDB2.
Similar content being viewed by others


Introduction
Asthma is a complex disease underpinned by a combination of genetic and environmental factors. Positional cloning and genome-wide association studies have resulted in the identification of a number of loci that influence asthma and asthma-related traits such as total serum immunoglobulin E (IgE).1 An understanding of the function of the genes within these loci as well as their regulation will provide important insights into their roles in disease.
Previously, we have reported a highly significant genetic association between a locus on human chromosome 13 and total serum IgE.2 The locus contains the genes PHF11 and SETDB2. Both genes are in the same linkage disequilibrium (LD) block and are separated by a mere 4 kb. The association between single-nucleotide polymorphisms in PHF11 and asthma, or asthma-associated traits, has been investigated in many subsequent studies, with reports confirming the association in a number of ethnic populations.3, 4, 5 It is notable, however, that two studies have failed to replicate the original finding6, 7 potentially due to true differences in the effect of PHF11 variants between groups (e.g. due to variation in environmental exposures or through epistatic or gene–environment interactions) and variance in the patterns of LD between typed and true causative variants.
SETDB2 is transcribed in the same direction as PHF11 and cotranscripts extending from SETDB2 to PHF11 exist in many human tissues.2 Owing to the small intervening distance and the LD structure between these genes, it is not currently possible to delineate the relative roles of SETDB2 and PHF11 in asthma and its associated traits. SETDB2 has a methyl-CpG-binding domain and SET domain that modulates gene expression epigenetically through histone H3 lysine methylation. Abnormal histone methylation has been found in many diseases including asthma.8 In this report, we sequenced the 15 exons of SETDB2 to discover new polymorphisms that may underlie asthma-associated traits. We identified an AT/G mutation (rs386770867), comprising both an insertion and substitution at the same locus, positioned in the 5′-untranslated region (UTR) of SETDB2. We genotyped this variant in our family-based cohorts of asthma and then investigated its role in the regulation of SETDB2 using electrophoretic mobility shift assays (EMSAs), alleoty** and dual reporter gene assay analysis.
Results
SETDB2 sequencing and genoty**
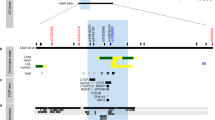
Sequencing of the 15 exons of SETDB2 in 10 diploid genomes (five unrelated individuals with atopic disease and five unrelated control individuals) together with a pool of DNA from 32 unrelated individuals resulted in the identification of three mutations. Two missense mutations located in exons 7 and 10 had been genotyped in our previous work (referenced as d8ex7 and d8ex10).2 The third variant was an AT/G mutation located in the 5′-UTR of exon 1 and subsequently was designated as rs386770867 in the NCBI dbSNP database (Figures 1a and b). Previously, we had identified and genotyped 12 SNPs in the SETDB2 region in an asthma cohort, finding association of IgE with three (rs7998427, P=0.02; b1_2, P=0.008; rs11619265, P=0.002).2 We therefore genotyped rs386770867 first in the Australian panel of families, and found that the mutant allele (AT) was significantly associated not only with total serum IgE levels (P=0.0012) but also to the phenotype of RASTI (Radio AllergoSorbent Testing Index; the index of the sum of serum IgE levels to the two allergens of house dust mite and grass pollen) (P=0.002), both of which are asthma-related phenotypes. The significant association with RASTI was also found in the UK panel (P=0.01), making this SNP the most strongly associated with asthma-related traits in SETDB2 (Table 1).

(a) The location of the AT/G mutation in the UTR of SETDB2. Position of the AT/G mutation is on chr13: 50 018 841–50 018 842 (Build Hg19), it is located at −492/−493 before the translation codon ATG of SETDB2. (b) Sequence showing the rs386770867 polymorphism.
Transcription factor binding analysis of rs386770867
The sequence spanning rs386770867 was examined using the transcription factor binding prediction programs TFSearch,9 TFScan10 and MatInspector.11 Three transcription factors (HS$IL6_06, HS$GG_12 [NF-E] and SRY) were predicted to bind the region independently of the mutation. One transcription factor, HS$GMCSF_04 (Ying Yang 1 (YY1)), was predicted to bind the AT allele only, whereas v-Myb was predicted to bind only the G allele (Table 2).
Transcription factor binding is modulated in vitro by rs386770867
Initial EMSAs using BEAS-2B (bronchial epithelial cell line), Calu-3 (airway epithelial cell line) and Daudi (male B lymphoblast) nuclear extracts identified three major complexes (Figure 2a). Complex 1 was present for both alleles for all the nuclear extracts. Complex 3 formed for the AT allele alone, and again was present for all three nuclear extracts. Complex 2, however, was only observed using the G allele probe and the Daudi nuclear extract. To confirm the specificity of complexes 2 and 3, a series of competition assays were performed using molar excesses of unlabelled probes for each allele, and an unrelated sequence. Competition assays using Calu-3 nuclear extract demonstrated that for both the AT and G probes binding to complex 1 was efficiently competed using unlabelled probes for either allele. The intensity of complex 3 was, however, only reduced in the presence of the unlabelled AT probe (Figure 2b). This was also seen for the Daudi nuclear extract (Figure 2c). Complex 2 was only efficiently competed in the presence of the unlabelled G probe (Figure 2c). The data demonstrated the specific binding of proteins for both AT and G alleles of rs386770867.

(a) EMSA results for rs386770867 using BEAS-2B, Calu-3 and Daudi nuclear extracts A total of 5 μg of nuclear extract was used per reaction. (b) Competition EMSA using Calu-3 nuclear extract. Excess of cold probe used was 10x, 50x and 100x with 5 μg of nuclear extract used per reaction. (c) Competition EMSA using 5 μg of Daudi nuclear extract and 10x, 50x and 100x excesses of cold probe. (d) Supershift assay using 5 μg of Daudi cell nuclear extract per reaction.
To identify the proteins responsible for the allele-specific complexes, supershift assays were performed using antibodies for the transcription factors implicated by the bioinformatic analyses of the region: SRY, YY1 and c-Myb. In addition, a reaction using an Oct-1 antibody, not implicated to bind either allele, was included as a negative control. Complex 3 was supershifted in the presence of anti-YY1, whereas complex 2 was abolished by anti-SRY (Figure 2d). Complex 1 was unaffected by any of the antibodies tested.
Alleloty**
To determine the effect of the allele-specific transcription factor binding on SETDB2, the relative expression of the two alleles in heterozygous individuals was investigated by alleloty**.12, 13 The 35 heterozygotes for rs386770867 were identified by genoty** the MRC-A/RNA cohort. For each of the heterozygous individuals, eight replicate matrix-assisted laser desorption/ionization genoty** reactions were performed using both genomic DNA and cDNA for each individual.
The AT:G ratio of peak areas for each replicate genomic DNA result was calculated for all samples. Replicate results outside the 1.5x interquartile range were excluded as outliers (N=9), and the mean of the remaining data calculated (mean=1.78; N=295). The AT:G peak area ratio in the cDNA of the 35 individuals was compared with the mean ratio from the genomic DNA using a paired-samples t-test. This revealed a modest, but highly significant preferential expression of the AT allele across the 35 individuals as a whole (cDNA mean=2.07; gDNA mean=1.78; difference of means=0.286; 95% confidence interval (CI) of the difference=0.232–0.340; fold change=1.16; P=1.26 × 10−21).
Limiting the analysis to include only those individuals with no outlying replicate cDNA results (N=29) confirmed the original finding (cDNA mean=2.10; gDNA mean=1.78; difference of means=0.314; 95% CI of the difference=0.269–0.358; fold change=1.18; P=5.66 × 10−32) (Table 3). Comparison of each patient separately to the genomic DNA ratio identified 25 individuals with preferential expression of the AT allele, and 1 for the G allele (P<0.05) (Table 3). Significant differences were also observed for both sexes when analysed separately, although preferential expression of the AT allele appears to be stronger in males (cDNA mean=2.17; gDNA mean=1.78; difference of means=0.381; 95% CI of the difference=0.306–0.457; fold change=1.21; P=5.07 × 10−18) compared with that in females (cDNA mean=1.96; gDNA mean=1.78; difference of means=0.171; 95% CI of the difference=0.100–0.242; fold change=1.10; P=6.35 × 10−6) (Table 3).
Dual-luciferase reporter assay results
Jurkat T cells were cultured for the dual-luciferase reporter assay. The Renilla pRL-SV40 plasmid was used as a control for the transfection efficiency. The luciferase expression was documented as average relative light units/pGL3 control. The AT allele plasmid showed significantly greater expression levels of luciferase compared with the pGL3 vector (P<0.01). The G-allele plasmid showed slightly lower expression of luciferase compared with the pGL3 vector, but this difference was not statistically significant in Jurkat T cells (Figure 3).

Dual-luciferase reporter assay results of AT and G alleles. The Renilla pRL-SV40 plasmid was used as a control for the transfection efficiency. The luciferase expression was documented as average relative light units/pGL3 control.
Discussion
Chromosome 13q was first shown to associate with asthma-related traits by whole-genome linkage analysis in 1996.14 Subsequently, the genes PHF11 and SETDB2 were implicated by positional cloning of the locus.2, 15 Our earlier genetic analyses had indicated that variation in total serum IgE levels was attributable, at least in part, to variation in the PHF11 gene on chromosome 13q14. The coordinate regulation of PHF11 and the nearby gene SETDB2 and the possible presence of more distant alleles influencing association suggested, however, that investigation of the function of the locus should include the two genes within this locus.
SETDB2 is a histone H3 lysine 9 methyltransferase that modulates gene expression epigenetically through histone H3 methylation.16 Methylation of histone H3 at lysine 9 has emerged as an important player in the formation of heterochromatin, chromatin condensation and transcriptional repression. Depletion of SETDB2 coincides with a loss of CENP proteins and delayed mitosis, suggesting that SETDB2 participates in chromosome condensation and segregation.16 Knockdown of Setdb2 in zebrafish results in a massive expansion of dorsal organizer markers floating head, goosecoid and chordin, as well as a significant increase of fibroblast growth factor 8.17 The family of fibroblast growth factors regulates a plethora of developmental processes, including brain patterning, branching morphogenesis and limb development. The recent discovery of the crucial roles of the endocrine-acting fibroblast growth factor 19 subfamily in bile acid, glucose and phosphate homeostasis has sparked renewed interest in the pharmacological potential of this family.18
In this report, we first systematically sequenced the 15 exons of SETDB2 and identified an AT/G mutation in the 5′-UTR (rs386770867). The AT allele of this mutation was most highly associated with IgE levels. EMSA analysis of rs386770867 revealed three main bandshifts, an AT allele-specific complex (band 3), a G-allele-specific complex (band 2) and a complex present for both alleles (band 1). The AT-specific and constant complexes were seen in all three nuclear extracts used (Calu-3, BEAS-2B and Daudi), whereas the G-allele-specific band was only seen in the presence of the Daudi nuclear extract. Supershift experiments indicated that the AT allele complex was due to the YY1 transcription factor binding the probe, whereas the G allele was due to SRY binding. The differences in binding patterns between cell lines may therefore indicate varying regulation of SETDB2 in different tissue types.
YY1 is a transcription factor, so called because of its ability to act both as an initiator, activator and repressor of transcription.19, 20 It is also known as a member of the GLI-Krüppel family of zinc-finger transcription factors.19 The protein is highly conserved and ubiquitously expressed. Examples of its role as a repressor are the P5 promoter of adeno-associated virus, c-fos and interleukin-5.21, 22, 23 It is believed that YY1 is a repressor by default, but under certain conditions its function can be altered to that of an activator.24
An association between YY1 and asthma has already been previously reported. Tumor growth factor-β is an asthma candidate gene, the product of which inhibits B and T cells, decreases IgE production and mast cell proliferation and induces eosinophil apoptosis. Subsequently, it has been shown to be associated with asthma. YY1 has also been shown to inhibit tumor growth factor-β-induced cell differentiation.25
Sry is the gene required for testis determination and differentiation in mammals.26 Recent work has also indicated additional roles for Sry, for instance, in the regulation of tyrosine hydroxylase gene transcription.27 SRY is sex-specific and so the possibility remains that this could be related to the sex-specific influence on asthma, which has previously been reported. For instance, Weiss et al.28 reported significant sex-specific differences in FEV1 (forced expiratory volume in 1 s), eosinophils and IgE levels, all of which are quantitative traits that are associated with asthma. In addition, two traits, FEV1/FVC (forced vital capacity) and eosinophil levels, were found to have sex-specific linkage to regions of the genome and had significant genotype–sex interactions.28 Interestingly, our alleloty** data did indicate a stronger preferential expression of the AT allele in males compared with females. This perhaps indicates a greater suppression of allele G expression because of SRY binding in males.
The results of the EMSAs indicate that rs386770867 changes the sequence from a YY1 binding site to that of SRY. Both transcription factors are able to activate or repress transcription based on the circumstances in which binding takes place. Alleoty** revealed that the AT allele had a modest, but highly significant, increased expression in heterozygous individuals relative to the G allele. Although the scale of the difference was subtle, this could be of biological significance in conditions enhancing IgE production in vivo. The dual-luciferase reporter assay showed that the AT allele significantly increases the transcription of luciferase. This therefore indicates that changing from YY1 to SRY binding as a result of the rs386770867 variant decreases expression of SETDB2, consistent with our findings in relation to allele expression in males and females.
Although this work provides additional clarity regarding the mechanisms underlying the chromosome 13q14 asthma locus, many unanswered questions remain. Recent investigation on PHF11 found that it not only regulates the T-cell function29, 30 but also is involved in immunoglobulin class switching on B cells.31 Therefore, our findings support the role of both genes at this locus influencing asthma-associated traits, although their relative influences are as yet unknown. Future work will need to focus on the functions of PHF11 and SETDB2 in asthma pathophysiology, in particular, how the two molecules work together during immune cell regulation. Additionally, using expression quantitative trait loci data will help to determine both the functional effects of rs386770867 and the functional role of SETDB2. The dissection of regulating elements in the locus will also increase our understanding of the factors that influence the expression of these genes.
Materials and methods
SETDB2 exon sequencing and genoty** of rs386770867
Sanger sequencing of 10 diploid genomes (five unrelated individuals with atopic disease and five unrelated control individuals) together with a pool of DNA from 32 unrelated individuals for all 15 exons of SETDB2 was performed. This gave us 99.9% probability of detecting alleles with a minimum frequency of 0.2 and 99% probability of detecting alleles with a minimum frequency of 0.1. Dilution experiments with known alleles indicated that we were able to detect allele frequencies >0.15 with the pool. An AT/G mutation in the 5′-UTR was identified, subsequently this mutation was designated as rs386770867. The mutation was genotyped by a PCR-based restriction fragment length polymorphism method (forward primer: 5′-CGACAGTTCCTCTAGCCG-3′; reverse primer: 5′-CTGAGACAGACAGGCTGTA-3′—generating a PCR product of 167/168 bp). The reverse primer contained a modified A to T change (bold) to generate a restriction enzyme site (5′-GTAC-3′) for Rsal, allowing discrimination between the AT and G alleles on a 2.5% agarose gel. The mutation was genotyped in family-based asthma cohorts, which have been described in detail previously.1 Briefly, the Australian panel consisted of 364 subjects in 80 nuclear families of European descent from the rural town of Busselton in Western Australia. The families contained a total of 203 offspring forming 172 sib-pairs. The mean age of the children was 12.6±1.3 (s.e.) years, their geometric mean IgE was 55.7±1.1 k Ul−1 and their mean skin test index was 4.0±0.41 mm.14 The UK panel consisted of 87 nuclear families of British descent recruited when a child of the family attended an asthma clinic in the Oxford region. The families contained 216 offspring (148 sibling pairs) and consisted of 4 large pedigrees and 66 nuclear families. For analysis, three-generation pedigrees were divided into independent two-generation pedigrees, to give a total of 88 nuclear families. The father was atopic in 31 parent pairs, the mother in 17, both parents in 21 and neither parent in 5.32 Ethical approvals were given by Australia or the United Kingdom Multicentre Research Ethics Committee. All subjects or their parents gave written informed consent to the study. The genotypes were checked by two experienced researchers. The quantitative trait loci were analysed by the QTL program as described previously.2
Transcription factor binding search programs
Putative transcription factor binding sites in the sequence spanning rs386770867 were identified using the programs TFSEARCH,9 TFSCAN10 and MatInspector (Genomatix Software GmbH, Munich, Germany).11 Programs were run using the default settings.
Cell culture and nuclear protein extraction
Daudi (male B lymphoblast), Calu-3 (airway epithelial), BEAS-2B (bronchial epithelial) and Jurkat (T lymphoblast) cell lines were obtained from the American Type Culture Collection (Teddington, UK). Airway epithelium and B and T lymphoblasts are of importance in asthma pathogenesis, making these cell lines relevant for EMSA investigations. In addition, a SETDB2–PHF11 gene complex has been shown to be expressed in lung tissue and lymphocytes.2 Jurkat and Daudi cells were cultured in RPMI 1640 media (Sigma, Gillingham, UK) supplemented to contain final concentrations of 10% fetal bovine serum (Sigma), 10 mmol l−1 HEPES (Sigma), 1 mmol l−1 sodium pyruvate (Sigma), 4.5 g l−1 glucose (Sigma), 2 mmol l−1 l-glutamine (Sigma), 20 U ml−1 penicillin and 0.1 mg ml−1 streptomycin (Sigma). Calu-3 cells were cultured in Dulbecco's modified Eagle's medium (Sigma), supplemented to contain final concentrations of 10% fetal bovine serum, 1 mmol l−1 sodium pyruvate, 2 mmol l−1 l-glutamine, 20 U ml−1 penicillin and 0.1 mg ml−1 streptomycin. BEAS-2B cells were cultured in keratinocyte-SFM (serum-free medium) (Gibco-BRL, Paisley, UK) media supplemented with 2 mmol l−1 l-glutamine, 5 ng ml−1 epidermal growth factor (Gibco-BRL), 50 mg ml−1 bovine pituitary extract (Gibco-BRL), 20 U ml−1 penicillin and 0.1 mg ml−1 streptomycin. All cell lines were cultured at 37 °C and 5% CO2. Nuclear extracts were prepared by using a modified Schreiber protocol33 and quantified using the Bradford assay.34
Electrophoretic mobility shift assays
Sense and antisense single-stranded oligonucleotides for both alleles of rs386770867 were designed and synthesized. Each consisted of the SNP allele and 15 bases of 5′ and 3′ flanking sequence. An AGCT tag was added to the 5′ end of each oligonucleotide to facilitate subsequent radiolabelling (rs386770867 in bold): AT allele: forward, GACTGAGTTTCCAACCTCCATTTCAGCCTGTCTGTC; reverse, GACTGACAGACAGGCTGAAATGGAGGTTGGAAACTC; G allele: forward, GACTGAGTTTCCAACCTCCGTTCAGCCTGTCTGTC; reverse, GACTGACAGACAGGCTGAACGGAGGTTGGAAACTC.
The oligonucleotides were annealed to form probes for both alleles and labelled with [α-32P]CTP (Amersham Biosciences, Little Chalfont, UK) using the Klenow fragment (New England Biolabs, Hitchin, UK).35 Typically, EMSA binding reactions contained 3–5 μg of nuclear extract, 2 μl radiolabelled probe (40 counts per second per μl), 1 × binding buffer (Promega, Hampshire, UK), 0.05–0.1 mg ml−1 poly(dIdC)·poly(dI−dC) (Sigma) and 4% glycerol (Sigma) in a total reaction volume of 15 μl. For competition assays, reactions included unlabelled competitor probe at a concentration of 10x, 50x or 100x that of the labelled probe. For supershift assays, 2 μg antibodies were added to the reactions before the first incubation step. The antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA) used for supershift assays were YY1, SRY and c-Myb. Reactions were incubated without the labelled probe for 10 min at room temperature. On addition of the labelled probe, reactions were incubated for a further 20 min at room temperature. Products were run on a 6% non-denaturing polyacrylamide gel at 4 °C and 100 V for ~2 h using 0.5x TBE (Tris-borate-EDTA) run buffer before visualization by exposure to Kodak X-Omat AR film (Sigma).
Alleloty** subjects
Allele-specific expression analysis was performed in a subset of the MRC-A population, MRC-A/RNA. The MRC-A panel consists of 195 siblings and their parents in 95 nuclear pedigrees in which we have previously carried out genome-wide association studies for global gene expression and asthma status.36, 37 Our sample contained 357 subjects (183 male) with a mean age in children of 12.2 years (ranging from 2 to 39) and adults of 42 years (ranging from 27 to 61 years). One hundred and thirteen children had doctor diagnosed asthma. Epstein–Barr virus-immortalized lymphoblastoid cell lines were created from blood taken from all the children in the population. The MRC-A/RNA cohort is a subset of the MRC-A population. It is made up of 94 children from the 49 sibling pairs selected from the 200 families on the basis of being the most highly discordant for IgE levels. The Epstein–Barr virus cell lines provided a source of both DNA and RNA from these individuals. Ethical approval was given by the United Kingdom Multicentre Research Ethics Committee. All subjects or their parents gave written informed consent to the study.
Alleloty**
Allele-specific expression analysis was performed using DNA and RNA from a subset of the MRC-A panel, MRC-A/RNA. Details of the alleloty** method have been described previously.36 Briefly, 10 μg of RNA was used to synthesize double-stranded cDNA using the One-cycle cDNA Synthesis Kit (Affymetrix, Santa Clara, CA, USA). Allele peak areas in the Sequenom hME Assay (Sequenom, San Diego, CA, USA) mass spectra are directly proportional to the amount of starting material in the assay. Consequently, the peak areas provide a method of comparing the relative amounts of two alleles for a heterozygote. To determine whether allele-specific expression is occurring, the ratio of the peak areas in the genomic DNA is compared with that in the cDNA to account for any natural bias in peak areas. If the two ratios differ significantly, preferential allele expression is indicated. Amplification was performed in 5 μl reaction volumes with either 5 ng template genomic DNA or cDNA equivalent to 5 ng total RNA. The PCR program was as detailed previously.38 Replicate PCRs (N=8) for both genomic DNA and cDNA were performed for each subject. PCR products were prepared for mass spectrometry following the standard Sequenom method.39
Statistical analyses of the allele ratios for alleloty** were performed using Excel 2010 (Microsoft). For paired-samples t-test, unless otherwise stated, the ratio of the peak areas from the mass spectrometry results of allele AT relative to allele G in the cDNA was calculated and compared with the mean ratio in the genomic DNA for all samples to minimize the effect of any potential variation in genomic DNA ratio.
Dual-luciferase reporter assay
Using the pLG3 promoter plasmid, a construct was made incorporating a 281/280 bp fragment containing rs386770867. PCR primers used were: forward, ATAAGCTAGCGAGCCAGGTGAAACCAAGGC; reverse, GCTCGCTCGAGCTTTGACAACCTCAGGTTCC (GCTAGC, Nhel site; CTCGAG, Xhol site). PCRs were carried out in individuals homozygous for either the AT or G allele. PCR products were then digested with Nhel and Xhol and the bands obtained purified and ligated into the pLG3 promoter plasmid. Clones were picked, cultured overnight for DNA extraction and then sequenced to identify the appropriate constructs.
Jurkat T cells were seeded into 12-well plates at 1x106 cells per ml and 1 μg plasmids were transfected with Lonza Amaxa Cell Line NucleofectorT Kit V (Lonza AG, Castleford, UK). Co-transfection with the Renilla pRL-SV40 plasmid (1 μg per transfect) provided a control for transfection efficiency in a dual-luciferase reporter assay system (Promega). The untransfected pGL3 promoter plasmid was used as the negative control. After 24 h transfection, a dual-luciferase assay was carried out and expression of luciferase was measured both according to the manufacturer's protocol. Briefly, the medium was removed from cultured cells and 100 μl 1x Passive Lysis Buffer was added to the wells; then, 25 μl Passive Lysis Buffer lysate was transferred to a new 96-well plate and Luciferase Assay Buffer II was added. Firefly luciferase activity was measured; then, 100 μl Stop & Glo reagent (dual-luciferase reporter assay kit, Promega) was added, and Renilla luciferase activity was measured. Three independent experiments were performed and results were analysed using the Student’s t-tests.
Change history
12 May 2016
This article has been corrected since publication and a corrigendum has also been published.
References
Zhang Y, Moffatt MF, Cookson WO . Genetic and genomic approaches to asthma: new insights for the origins. Curr Opin Pulm Med 2012; 18: 6–13.
Zhang Y, Leaves NI, Anderson GG, Ponting CP, Broxholme J, Holt R et al. Positional cloning of a quantitative trait locus on chromosome 13q14 that influences immunoglobulin E levels and asthma. Nat Genet 2003; 34: 181–186.
Gao J, Li W, Willis-Owen SA, Jiang L, Ma Y, Tian X et al. Polymorphisms of PHF11 and DPP10 are associated with asthma and related traits in a Chinese population. Respiration 2010; 79: 17–24.
Hersh CP, Raby BA, Soto-Quiros ME, Murphy AJ, Avila L, Lasky-Su J et al. Comprehensive testing of positionally cloned asthma genes in two populations. Am J Respir Crit Care Med 2007; 176: 849–857.
Jang N, Stewart G, Jones G . Polymorphisms within the PHF11 gene at chromosome 13q14 are associated with childhood atopic dermatitis. Genes Immun 2005; 6: 262–264.
McClenaghan J, Warrington NM, Jamrozik EF, Hui J, Beilby JP, Hansen J et al. The PHF11 gene is not associated with asthma or asthma phenotypes in two independent populations. Thorax 2009; 64: 620–625.
Blakey JD, Sayers I, Ring SM, Strachan DP, Hall IP . Positionally cloned asthma susceptibility gene polymorphisms and disease risk in the British 1958 Birth Cohort. Thorax 2009; 64: 381–387.
Clifford RL, John AE, Brightling CE, Knox AJ . Abnormal histone methylation is responsible for increased vascular endothelial growth factor 165a secretion from airway smooth muscle cells in asthma. J Immunol 2012; 189: 819–831.
Heinemeyer T, Wingender E, Reuter I, Hermjakob H, Kel AE, Kel OV et al. Databases on transcriptional regulation: TRANSFAC, TRRD and COMPEL. Nucleic Acids Res 1998; 26: 362–367.
Faisst S, Meyer S . Compilation of vertebrate-encoded transcription factors. Nucleic Acids Res 1992; 20: 3–26.
Quandt K, Frech K, Karas H, Wingender E, Werner T . MatInd and MatInspector: new fast and versatile tools for detection of consensus matches in nucleotide sequence data. Nucleic Acids Res 1995; 23: 4878–4884.
Knight JC, Keating BJ, Rockett KA, Kwiatkowski DP . In vivo characterization of regulatory polymorphisms by allele-specific quantification of RNA polymerase loading. Nat Genet 2003; 33: 469–475.
Yan H, Yuan W, Velculescu VE, Vogelstein B, Kinzler KW . Allelic variation in human gene expression. Science 2002; 297: 1143.
Daniels SE, Bhattacharrya S, James A, Leaves NI, Young A, Hill MR et al. A genome-wide search for quantitative trait loci underlying asthma. Nature 1996; 383: 247–250.
Anderson GG, Cookson WO . Recent advances in the genetics of allergy and asthma. Mol Med Today 1999; 5: 264–273.
Falandry C, Fourel G, Galy V, Ristriani T, Horard B, Bensimon E et al. CLLD8/KMT1F is a lysine methyltransferase that is important for chromosome segregation. J Biol Chem 2010; 285: 20234–20241.
Xu PF, Zhu KY, ** Y, Chen Y, Sun XJ, Deng M et al. Setdb2 restricts dorsal organizer territory and regulates left-right asymmetry through suppressing fgf8 activity. Proc Natl Acad Sci USA 2010; 107: 2521–2526.
Beenken A, Mohammadi M . The FGF family: biology, pathophysiology and therapy. Nat Rev Drug Discov 2009; 8: 235–253.
Shi Y, Seto E, Chang LS, Shenk T . Transcriptional repression by YY1, a human GLI-Kruppel-related protein, and relief of repression by adenovirus E1A protein. Cell 1991; 67: 377–388.
Seto E, Shi Y, Shenk T . YY1 is an initiator sequence-binding protein that directs and activates transcription in vitro. Nature 1991; 354: 241–245.
Houbaviy HB, Usheva A, Shenk T, Burley SK . Cocrystal structure of YY1 bound to the adeno-associated virus P5 initiator. Proc Natl Acad Sci USA 1996; 93: 13577–13582.
Natesan S, Gilman M . YY1 facilitates the association of serum response factor with the c-fos serum response element. Mol Cell Biol 1995; 15: 5975–5982.
Schwenger GT, Fournier R, Hall LM, Sanderson CJ, Mordvinov VA . Nuclear factor of activated T cells and YY1 combine to repress IL-5 expression in a human T-cell line. J Allergy Clin Immunol 1999; 104 (Part 1): 820–827.
Thomas MJ, Seto E . Unlocking the mechanisms of transcription factor YY1: are chromatin modifying enzymes the key? Gene 1999; 236: 197–208.
Kurisaki K, Kurisaki A, Valcourt U, Terentiev AA, Pardali K, Ten Dijke P et al. Nuclear factor YY1 inhibits transforming growth factor beta- and bone morphogenetic protein-induced cell differentiation. Mol Cell Biol 2003; 23: 4494–4510.
Sinclair AH, Berta P, Palmer MS, Hawkins JR, Griffiths BL, Smith MJ et al. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 1990; 346: 240–244.
Milsted A, Serova L, Sabban EL, Dunphy G, Turner ME, Ely DL . Regulation of tyrosine hydroxylase gene transcription by Sry. Neurosci Lett 2004; 369: 203–207.
Weiss LA, Pan L, Abney M, Ober C . The sex-specific genetic architecture of quantitative traits in humans. Nat Genet 2006; 38: 218–222.
Clarke E, Rahman N, Page N, Rolph MS, Stewart GJ, Jones GJ . Functional characterization of the atopy-associated gene PHF11. J Allergy Clin Immunol 2008; 121: 1148–1154 e3.
Zhang Y, Dean C, Chessum L, Nguyen D, Stewart M, Taylor M et al. Functional analysis of a novel ENU-induced PHD finger 11 (Phf11) mouse mutant. Mamm Genome 2014.
Ikari J, Inamine A, Yamamoto T, Watanabe-Takano H, Yoshida N, Fujimura L et al. Plant homeodomain finger protein 11 promotes class switch recombination to IgE in murine activated B cells. Allergy 2014; 69: 223–230.
Sandford AJ, Shirakawa T, Moffatt MF, Daniels SE, Ra C, Faux JA et al. Localisation of atopy and beta subunit of high-affinity IgE receptor (Fc epsilon RI) on chromosome 11q. Lancet 1993; 341: 332–334.
Schreiber E, Matthias P, Muller MM, Schaffner W . Rapid detection of octamer binding proteins with 'mini-extracts', prepared from a small number of cells. Nucleic Acids Res 1989; 17: 6419.
Bradford MM . A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976; 72: 248–254.
Hacking D, Knight JC, Rockett K, Brown H, Frampton J, Kwiatkowski DP et al. Increased in vivo transcription of an IL-8 haplotype associated with respiratory syncytial virus disease-susceptibility. Genes Immun 2004; 5: 274–282.
Dixon AL, Liang L, Moffatt MF, Chen W, Heath S, Wong KC et al. A genome-wide association study of global gene expression. Nat Genet 2007; 39: 1202–1207.
Moffatt MF, Kabesch M, Liang L, Dixon AL, Strachan D, Heath S et al. Genetic variants regulating ORMDL3 expression contribute to the risk of childhood asthma. Nature 2007; 448: 470–473.
Holt RJ, Zhang Y, Binia A, Dixon AL, Vandiedonck C, Cookson WO et al. Allele-specific transcription of the asthma-associated PHD finger protein 11 gene (PHF11) modulated by octamer-binding transcription factor 1 (Oct-1). J Allergy Clin Immunol 2011; 127: 1054–1062 e1-2.
Ding C, Cantor CR . A high-throughput gene expression analysis technique using competitive PCR and matrix-assisted laser desorption ionization time-of-flight MS. Proc Natl Acad Sci USA 2003; 100: 3059–3064.
Acknowledgements
YZ was supported by a Research Council UK Fellowship. WOCM and MFM are recipients of a Senior Investigator award from the Wellcome Trust.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Holt, R., Vandiedonck, C., Willis-Owen, S. et al. A functional AT/G polymorphism in the 5′-untranslated region of SETDB2 in the IgE locus on human chromosome 13q14. Genes Immun 16, 488–494 (2015). https://doi.org/10.1038/gene.2015.36
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/gene.2015.36
- Springer Nature Limited
This article is cited by
-
Equine vitiligo-like depigmentation in grey horses is related to genes involved in immune response and tumor metastasis
BMC Veterinary Research (2021)
-
Emerging roles of H3K9me3, SETDB1 and SETDB2 in therapy-induced cellular reprogramming
Clinical Epigenetics (2019)
-
The SETDB2 locus: evidence for a genetic link between handedness and atopic disease
Heredity (2018)