
Abstract
Although climate change-related concerns have long been raised regarding the sudden dieback of Korean fir (Abies koreana), the event’s etiology and subsequent ecosystem processes must be explained. Our study aims to clarify the continuity or transience of mass mortality events within the coarse woody debris (CWD) structure and, if transient, to identify the climatic conditions (1974–2021) that could be responsible for the massive dying phenomena in Korean fir populations. On average, precipitation during the non-growing season (November–April as winter) constituted 18.5% relative to the growth period; in the winter of 1999, it was 4.8% due to an abnormal drought event. The dead stems occurred evenly across all size classes. In the CWD structure, the density and biomass of the dead fir individuals peaked in decay classes II or III. The size distribution of the retained fir was inverse-J shaped across the entire altitudinal range. The abnormal winter drought event, causing root damage by soil frost and heaving, may be one of the factors that increased Korean fir mortality across the entire stem size range. Despite transient cohort senescence, the retained Korean fir individuals transmitted drought-resistant traits into the regional pool following the drought event.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Since the early 2000s, Korean fir trees (Abies koreana) growing in the montane forest of the southern Korean Peninsula have been dying in large numbers, possibly due to climate change-related stress (abnormal drought or heat) and species vulnerability (Korea National Arboretum 2014; Koo and Kim 2020) (Plate S1). Concerns regarding climate-driven (drought and heat stress) mortality of coniferous trees in temperate forests have been increasing (Allen et al. 2010). Several canopy types and tree dieback events in Pacific forests are also regarded as natural aspects of long-term forest dynamics (Mueller-Dombois 1992). Despite the complex and ambiguous effects of climate change on the viability of Korean firs (Kim et al. 1994; Kim and Choo 2000; Koo and Kim 2020), landscape change (vegetation succession and soil development) and the evolutionary context of the species (regeneration requirements of the species) can also account for the abrupt senescence phenomena (Korea National Arboretum 2014, 2015). An empirical approach is necessary to further understand the etiology of tree death and outline future perspectives for the dynamics of Korean fir (Koo and Kim 2020).
Based on the unimodal age distribution, the regeneration of Korean fir requires a catastrophic disturbance causing overstory release (Kim and Yoon 1991; Cho 1994; Kang et al. 1997); however, the timing and nature of the disturbance remain unclear. Climate features (drought, cooling, and extreme heat) and regional disturbance regimes (typhoons in summer) are potential causes of overstory release. Summer droughts (McGregor et al. 2021) and typhoons (Suzuki et al. 2013) accompanied by strong winds disrupt the plant’s growing season and result in mortality, primarily among large-sized individuals. Climate anomalies (e.g., abnormal winter droughts) can exhibit out-of-season mortality in a range of species and stem sizes (Kullman and Högberg 1989).
Several reports used tree ring growth variation to explain the cause of the massive dieback of Korean fir (Noh et al. 2018; Seo et al. 2021). However, the dendrological approach was more suitable for the growth patterns of individual trees but insufficient for the occurrence of mortality (Koo et al. 2001; Bigler and Bugmann 2004). Although it is imprecise data, the coarse woody debris (CWD) structure leaves distinct traces of ecological events (Spies et al. 1988; Stokland 2001). For example, soil frost induced by winter drought and the consequent root damage caused by winter drought can result in dead stems across the various size classes, especially for species with shallow rooting depths (Cleavitt et al. 2008). In addition, the event will be relatively transient if there is a concentrated density distribution within the specific decay classes of CWD. The relationship between climatic anomalies and forest structure can exhibit the time-lag effect (Kullman and Högberg 1989) and the importance of subsequent weather conditions (drought or extreme heat) (Auclair et al. 2010).
Our study aims to clarify the continuity or transience of mass mortality events within the CWD structure and, if transient, to identify the climatic conditions (1974–2021) that could be responsible for the massive dying phenomena in Korean fir populations. In this study, we use a CWD structure and climate data to exhibit the temporality of the mass senescence of Korean fir. The objectives of this study are as follows: (1) to clarify the modality of CWD structure and retained individuals of Korean fir, (2) to explore the potential climate events, and (3) to discuss the expected ecosystem processes.
Material and Methods
Species and Montane Landscape
Korean fir is an endemic tree species distributed across deciduous broad-leaved and mixed forests in montane vegetation between 800 and 1900 m above sea level in the southern Korean Peninsula (Korea National Arboretum 2014, 2015). The largest inland and island habitats of the Korean fir are Mt. Jirisan and Mt. Hallasan. In other montane habitats, Korean fir is distributed in small numbers, primarily along rocky ridges. Even on Mt. Jirisan, the largest population is inland; the spatial range with a sufficient abundance of Korean fir for the vegetation experiment is from the deciduous broad-leaved forest at 1300 m or higher to the rocky ridge at the top. The distribution area of Korean fir on Mt. Jirisan decreased by approximately 18%, from 262 ha in 1981 to 216 ha in 2007 (Kim and Lee 2013). Species belonging to the section Balsamea in the genus Abies, including Korean fir, have a regenerative mechanism that utilizes the changed conditions after dieback or decline due to a mass mortality event (Iwaki and Totsuka 1959; Sprugel 1976; Kohyama 1982, 1984; Diaci et al. 2011).
In the montane vegetation of the Korean Peninsula, anthropogenic effects, such as food and fuel gathering and outdoor activities, were maintained up to the 1980s. Such human activities led to the expansion of relatively early seral vegetation with opened canopies and well-drained soil, resulting in the expansion of Korean fir (Korea National Arboretum 2014, 2015). Since the 1980s, when legal inhibition for natural resource utilization was adopted, vegetation development has been in progress.
Study Area
The average annual temperature of Mt. Jirisan (1915 m above sea level) is 6.1 °C (min. – 0.5 °C to max. 11.9 °C), the annual precipitation is 1491.2 mm (min. 1365 mm to max. 1606.7 mm), the warmth index is 57.0 (min. 23.1 to max. 97.8) and the coldness index is – 32.7 (min. – 43.9 to max. – 22.3) (internal GIS data of the Korea National Arboretum). Mt. Jirisan, a national park, comprises a mixed floristic composition with warm and cool temperate floristic zones (Jung and Cho 2020). In addition, depending on the elevation, Mt. Jirisan has two vegetation types: (1) mixed forests with coniferous and deciduous trees in montane vegetation, and (2) deciduous forests (DeFs) dominated mainly by Quercus mongolica and Q. serrata in lowland vegetation (Cho et al. 2020). The main tree species are Korean fir, Jezo spruce (Picea jezoensis), red pines (Pinus koraiensis and P. densiflora), birches (Betula ermanii and B. cordata), oaks, maples (Acer pseudosieboldianum and A. pictum), and ashes (Fraxinus rhynchophylla and F. chiisanensis).
In mixed forests at high altitudes on Mt. Jirisan, soil pH ranges from 4.2 to 4.6, organic content ranges from 11.3 to 14.4%, total nitrogen ranges from 0.4 to 0.9%, cation exchange capacity (CEC) ranges from 13.8 to 17.2 cmol+ kg−1, and moisture content ranges from 21.7 to 29.9% (Cho et al. 2016; Noh et al. 2018).
Vegetation Surveys
Summer field surveys were conducted in 2017 and 2018 on Mt. Jirisan, where Korean fir grew (Fig. 1). Considering the spatial distribution of Korean fir and vegetation types in Mt. Jirisan (750–1900 m), survey sites were selected by the coverage of conifers (Abies, Picea, and Pinus): 100–65% as coniferous forest (CoF), 65–25% as mixed forest (MiF), and less than 25% as DeF. The canopy coverage of conifer elements was visually estimated using the sum of live and dead canopy trees.

Topography and location of the sampling sites (dark red squares) on Mt. Jirisan
The quadrat method was applied in a field survey (Barbour et al. 1998). For tree measurements in quadrats, 13 plots (1.07 ha) were established in DeF (five plots with 0.33 ha), MiF (four plots with 0.41 ha), and CoF (four plots with 0.33 ha). By considering the height of the canopy trees, the size of the plots (225–400 m2) varied. The elevational distribution of quadrats was as follows: 1310–1436 m in DeF, 1427–1619 m in MiF, and 1613–1811 m in CoF. In each plot, species name, height (m), and diameter at breast height (DBH, 1.3 m from the ground) were measured for all live tree species. Height and DBH were measured for Korean fir regardless of whether the tree was dead or alive. The status of dead trees was recorded as follows: DS as standing dead, DB as broken dead, DL as leaning dead or leaning dead against a tree, and DF as dead fallen (i.e., dead by uprooting).
Soil samples were collected from the plots to analyze their chemical properties. Three randomly placed soil samples were obtained from below the litter layer within each plot. Each soil sample was air-dried, sieved through a 2-mm mesh, and then composited for chemical analyses. All analyses were conducted at the National Instrumentation Center for Environmental Management (Seoul, Korea). Analyzed soil properties comprised pH, organic matter content (%), total phosphorus (mg kg−1), and exchangeable cations (Ca, Mg, K, and Na) (mg kg−1).
Regional Climate
Precipitation and temperature data were obtained at Sancheong and Hamyang, the closest appropriate weather stations to Mt. Jirisan. The data were available from 1973 to 2021. Rainfall and temperature data from the two stations were combined, and mean values were applied to represent the study area.
To capture the climate anomaly, climate factors were divided into the non-growing season (November–April) and the growing season (May to October) of Korean fir. Because the winter dormancy of Korean fir spans 2 years, it was only calculated from 1974 to 2020, unlike the growing period of spring and summer in the study region. For the two divided phenological periods, we also calculated the standardized precipitation evapotranspiration index (SPEI) using a time series of the climatic water balance (precipitation minus potential evapotranspiration) (Vincente 2010). In the SPEI, potential evapotranspiration was followed by Thornthwaite (1948).
Cautiously, precipitation in the non-growing season in 1974 is the summed precipitation from November 1973 to April 1974. Thus, for example, if the precipitation of the non-growing season in 1990 was 1,000 mm, it means the summed precipitation is for 2 months (November and December) of 1989 and 4 months (January–April) of 1990.
Coarse Woody Debris (CWD)
The CWD (DBH > 10 cm) distribution was analyzed following the methodology of Fukasawa et al. (2014), who applied five decay classes using visual criteria according to Antos and Parish (2002) for snags and Sollins (1982) for logs. For snags, the presence or absence of needles and bark is an essential criterion to classify the decay class. For logs, the primary criterion is bark status (intact or sloughing). To measure the wood density (WD), three dead individuals were selected for dead types and decay classes (except the zero class, which is a green tree). Wood chips were collected using an increment borer at a 5–10 cm depth. After measuring the dry weight by drying the collected wood chips in a dryer at 40 °C for 7 days, the volume was calculated from the inner diameter (0.5 cm) of the increment borer and the length of the sample to calculate WD. The CWD mass was calculated by multiplying the volume by WD.
The volume of CWD (snags and logs) and retained Korean fir was calculated using the formula by Olajuyigbe et al. (2011). For snag volume, including live individuals, the diameters of the upper and lower ends were estimated by multiplying the DBH by 0.8 and 1.2, respectively.
where L is the length of the log (m), and d1 and d2 are the diameters at the large and small ends, respectively.
where h is the height of the snag (m), and R and r are the measured radii of the upper and lower ends of the dead tree, respectively.
Statistical Analysis
The structure of the declining Korean fir population was analyzed according to the abundance (size, density, volume, and mass) distribution of live and dead trees. First, the size distribution of all trees and Korean fir was calculated by summing the measured values in quadrats established in three vegetation types and expressed by converting them into a unit area (ha). Next, the measured stem density (D, stems ha−1) and breast height area (BHA, m2 ha−1) were calculated.
By considering the low sample size (four plots in a vegetation type), a Kruskal–Wallis one-way nonparametric analysis of variance was performed for environmental factors (site conditions and soil properties) and the mean abundance (volume and mass) of snag CWD among decay classes. A comparison for the means of log CWD was not conducted because of the low number of samples. Site conditions included elevation (m), litter layer depth (cm), aspect (°), slope (°), and rock coverage (%) by visual estimation. Soil properties included pH, organic matter content (%), total phosphorus (mg kg−1), and exchangeable cations (Ca, Mg, K, and Na) (mg kg−1). R version 4.1.2 (R Core Team 2021) was used for all statistical analysis: packages “ggplot2” version 3.3.3 (Wickham 2016) for data visualization and “rstatix” version 0.7.0 (Kassambara 2021) for univariate analysis.
Results
Regional Climate Anomaly
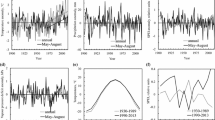
The mean annual precipitation was approximately 1437.8 mm from 1974 to 2021, with a marginally increasing pattern (Fig. 2a). On average, precipitation during the non-growing winter season constituted 18.5% of the annual rainfall (Fig. 2b, c). In 2000, it was 4.8% as the abnormal winter drought occurred. SPEI in the non-growing season also exhibited a similar pattern in 2000 (Fig. 2d). In the growing season, precipitation revealed an increasing pattern. In 1985 and 2020, abnormal precipitation events were observed (Fig. 2e). The growing season had no apparent drought events (Fig. 2f).

Precipitation with simple regression (solid lines) of a total, c non-growing season, and I growing season of the study area (1974–2021). b Precipitation ratios of the non-growing and growing seasons. Standardized precipitation evapotranspiration index (SPEI) as a drought index is also constructed for rainfall parameters of the d non-growing season and (f) growing season. The red arrows in (c, d) indicate the occurrence of the drought event in the non-growing season precipitation
The mean temperature in the study region was 12.6 ℃, with no significant high- or low-temperature years (Fig. S1). The mean highest and lowest temperatures in the non-growing season were 11.5 ℃ and – 1.1 ℃, respectively. The mean highest and lowest temperatures in the growing season were 26.8 ℃ and 15.6 ℃, respectively.
Vegetation Structure and Soil Environment
Korean fir was the dominant species in CoF and Q. mongolica in MiF and DeF (Table 1). The mean BHA and D of Korean fir increased with upward elevation: 6.0 ± 0.8 m2 ha−1 and 269.2 ± 103.7 stems ha−1 in DeF, 8.5 ± 1.9 m2 ha−1 and 409.5 ± 60.4 stems ha−1 in MiF, and 11.8 ± 5.7 m2 ha−1 and 618.5 ± 192.4 stems ha−1 in CoF. The abundance of B. ermanii and Acer komarovii, as sub-canopy layer compositors, also increased along the elevation, but the opposite was true for Q. mongolica. The co-occurring trees in the study sites were three coniferous species and seven broad-leaved deciduous species.
For the soil properties (Table 2), only total phosphorus content exhibited significant differences among vegetation types: 729.3 ± 175.8 mg kg−1 in CoF, 880.5 ± 166.2 mg kg−1 in MiF, and 721.4 ± 220.0 mg kg−1 in DeF. Organic matter content of the soil was higher in the order of MiF (23.0% ± 5.4%), CoF (21.4% ± 9.3%), and DeF (20.5% ± 4.6%), and pH was somewhat higher in CoF (4.2 ± 0.2) than in MiF and DeF.
Dead Korean Fir and CWD
The relative density of retained (10.5–31.7%) and dead (6.1–16.1%) Korean fir increased along the elevation gradient. Live trees had a higher density than dead trees in all vegetation types (Fig. 3a). DS was the predominant status of dead fir trees across the vegetation, and DL exhibited higher density with decreasing elevation (Fig. 3b).

a Relative density of alive (retained) and dead stems, and b status composition of dead individuals of Korean fir. The status classification is as follows: dead broken (DB), dead fallen (DF), dead leaning (DL), and dead standing (DS)
Most CWD occurred in snags (> 95% of CWD mass in CoF and MiF) across the vegetation. The volume (170.1 m3 ha−1) and mass (55.1 mg ha−1) of CWD in MiF were higher than those in CoF and DeF (Table 3). The abundance of CWD was concentrated in the second and third decay classes. Fresh CWD did not occur in CoF and DeF, with MiF being an exception. The mean volume and mass of CWD were significantly higher in decay class II (Fig. 4a, b) in CoF or III (Fig. 4e, f) in DeF. In MiF, no significance was observed, but the abundance of CWD was higher in decay class II (Fig. 4c, d).

Means (± standard deviation) of the abundance of coarse woody debris (CWD) along decay classes and vegetation types. a, c, and I are the volume of CWD in the coniferous forest (CoF), mixed forest (MiF), and deciduous forest (DeF), respectively, and b, d, and f are the mass of CWD in CoF, MiF, and DeF, respectively. The lowercase letters in the figure indicate the result of the post-hoc comparison for the mean, and different letters indicate significant differences to other groups. Because of the small sample size, decay class V in CoF has no letters in (a, b)
The shape of the size distribution of retained Korean fir was multi-modal (CoF and MiF) or reverse-J (DeF), and small, living trees had a higher density across the vegetation types (Fig. 5a–c). Dead Korean fir occurred in every size class and constituted a large portion of whole dead trees (Fig. 5d–f). The size distribution of dead Korean fir by applied decay classes revealed a concentration of dead stems in the second and third decay classes, similar to the concentration of CWD mass in the same classes (Fig. 6). Old and fresh dead stems were relatively rare in every vegetation type.

Size distribution of alive (a–c) and dead (d–f) stems of Korean fir by vegetation types. The gray color in the panel indicates the size distributions of entire tree species

Size distribution of dead stems (dark blue) of Korean fir by coarse woody debris (CWD) decay classes in vegetation types. The gray color in the panel indicates the summed density of dead stems of Korean fir
Discussion
The natural cause of Korean fir mortality across all size classes and elevational ranges was attributed to root damage caused by soil frost and frost heaving in conjunction with an atypical winter drought event. Roots of northern coniferous species are minimally frost-hardened, and the dieback is strongly correlated with winter freezing injury to roots by frost penetration in the absence of snow cover (Auclair et al. 2010). We discussed the consequent ecological processes following cohort senescence. For example, the persistence of trees across entire size classes after a winter drought event provides perspective on the regeneration of plant species. In particular, population turnover via the spread of winter drought-tolerant traits by retained stems and reproduction contributes to the resilience of high-altitude ecosystems to climate warming.
Anomalous Climate Feature and the Korean Fir Dieback
Our results on the Korean fir population structure in Mt. Jirisan suggest that the abnormal drought event in the non-growing season could be a factor inciting the transient dying event. In the regional climate, the abnormal winter drought occurred from November 1999 to April 2000, and drought or hot extremes in the growing period were relatively absent. Furthermore, the low rooting depth of Korean fir, even in the deep soil site (Korea National Arboretum 2014, Plate S1), makes the species more vulnerable to soil freezing. Such root morphology can contribute to dying events that do not occur in other tree species.
In three vegetation types, dead stems with mainly standing status occurred in entire stem size classes and concentrated in the middle decay classes. In Northeast Asia, tree mortality caused by typhoon disturbances with strong wind and heavy rain primarily concentrates on large individuals with broken and uprooted dead status (Suzuki et al. 2013; Choung et al. 2015; Sato et al. 2017). In South Korea, regular typhoons in summer are major disturbance events. Consistent with our results, DS predominates in the dead status of Korean fir mortality in Mt. Jirisan (Cho et al. 2016; Noh et al. 2018; Chun et al. 2021). Time-lag effects of typhoon disturbances on mortality caused by stresses from weakened soil conditions and root lifting by strong winds could have existed. In this case, smaller individuals in understory environments can be safe because trees in the upper forest layer will shield understory trees from the strong wind. Canopy trees are more susceptible to drought stress than smaller trees (O'Brien et al. 2017; McGregor et al. 2021). In the study region, neither droughts nor extreme heat events were observed during the growing period. The drought and heat stress of cliffs and talus slopes are detrimental to plant growth (Lee and Kim 1994); however, compared to terrained slopes (Plate S1), there are fewer dead trees among the Korean fir trees growing in those locations. Hong et al. (2008) also reported that the micro-environment supported by rocks could facilitate the expansion of Korean fir on Mt. Hallasan. Thus, the contribution of regular typhoons or drought events in summer to the structure of the dead Korean fir is limited.
Decreased snowpack caused by winter drought and consequent exceptional soil freezing and frost heaving can lead to size-independent mortality and the transient dieback of Korean fir. Winter (a non-growing season) drought implies little snowpack. While plants in snowpack experience relatively mild winter conditions, a lack of snowpack can be fatal to plants that have adapted to snow cover in winter (Körner and Hiltbrunner 2021). Snowpack reduction increases soil freezing and frost heaving, adversely affecting plant root and soil processes (Groffman et al. 2011). The weakened root function makes species more susceptible to various stresses (Marchand et al. 1986). Even in the released tree density, the annual ring width of Korean fir across all life stages began to decrease continuously after 2000 on Mt. Jirisan (Cho et al. 2016; Noh et al. 2018). Following the recent vegetation development initiated by prohibiting fuelwood gathering and decreasing animal grazing, improved soil water capacity can contribute to soil freezing in a montane forest (Korea National Arboretum 2014, 2015). The impairment of soil water drainage likely increases the mortality of Korean fir on Mt. Hallasan (Ahn et al. 2018).
Dieback and Ecosystem Processes
With sudden increases in CWD, consequent direct and indirect ecosystem processes are progressing. The volume and mass of CWD in CoF and MiF were much higher than those in the DeF (1.5–24.5 Mg ha−1) of South Korea (Kim et al. 2004, 2006). For example, the volume (146.8 m3 ha−1) and mass (55.1 Mg ha−1) of CWD in CoF were comparable to those of cold temperate rain coniferous forests in North America (60–1,189 m3 ha−1 and 10–511 Mg ha−1) (Harmon et al. 1986) and subalpine coniferous forests (217.9 m3 ha−1 and 42.4 Mg ha−1) in Japan (Fukasawa et al. 2014). Considering that the CWD of other tree species was extremely low at the survey site, Korean fir mortality events were a solitary source of the CWD. In the montane ecosystems in Korea, forest resources for fuel and food were actively used until the 1980s, resulting in vegetation with little CWD on the forest floor (Korea National Arboretum 2014; Cho et al. 2018). The CWD degradation proceeds relatively slowly in high-elevation ecosystems (Yin 1999), and even the CWD as downed logs does not significantly contribute to nutrient cycling after disturbance in conifer forests in the cold temperate forests (Laiho and Prescott 2004). Therefore, a sudden and massive accumulation of dead stems as standing snags is unlikely to contribute significantly to the rapid changes in soil nutrients.
According to the author's observations, the decline of small patches of Korean fir populations in the study area started around 2006 (Plate S1); therefore, the measured soil properties are at least ten years after the decline. The result of soil property was similar to reports of decline in Korean fir at other sites (Cho et al. 2016; Noh et al. 2018). As tree individuals die in the early stages of a sudden tree population decline, leaf litter and available soil nutrients increase (Kaňa et al. 2013). Improved light conditions through canopy release also benefit understory plant species. However, because of significantly reduced litterfall, the site where the Korean fir tree declines may change into a nutrient-poor soil environment, and the benefits of other co-existing tree species may be limited. The 10-year (2009–2018) monitoring report for Mt. Jirisan revealed minimal variation in the diameter class distribution of tree species other than Korean fir (Chun et al. 2021). Studies on the consequent changes in belowground properties that result from Korean fir dieback are required.
With large amounts of standing dead snags in Korean fir vegetation, species (insects and birds) that use snags as habitats or resource guilds can proliferate. By harboring various microhabitats, higher snag density contributes significantly to forest biodiversity (Paillet et al. 2017). Maser and Trappe (1984) reported that optimum symbiosis between conifer roots and mycorrhizal fungi could occur in middle decay classes. In our study, decay classes II and III were abundant.
As indirect effects, considering a decrease in litterfall as stem density release, the degradation of soil properties in Korean fir vegetation can progress. For example, a decrease in soil nutrients and an increase in water drainage will occur. Such degradation of soil environments may affect the regeneration of Korean fir, which prefers early successional soil conditions (Korea National Arboretum 2014, 2015).
Ecological Implications
Despite the sudden mortality events in various stem sizes across the elevational ranges, species resistance to anomalous climatic features might increase by facilitating population turnover (Fig. 7). Korean fir individuals in various sizes and reverse-J size distributions are retained in all vegetation types. Paradoxically, recent density reduction and canopy release by dying events may promote its regeneration environments as early seral conditions. Similar to the cohort dynamics model after the dieback tree population (see Boehmer et al. 2013), the seedling establishment and growth of advanced individuals are expected to be better in open canopy conditions caused by disturbance (Kim and Yoon 1991; Cho 1994; Lee and Cho 1993). Therefore, improved soil drainage and light conditions following the dying events may benefit the advanced understory individuals and the seedling establishment from the retained Korean fir. Regardless, the retained Korean fir in the study region may be flexible to soil freezing and frost heaving, and tolerant traits will spread into the regional population. For example, to resist freezing damage, the root system may possess relatively strong cellular vitality and physical flexibility from stretching (Cleavitt et al. 2008). Species recovery through dispersal and maturation, and the interval between abnormal events, can affect the dissemination of tolerant traits in the species. Consequently, whether the dieback originated from disturbances from a rotating or stochastic natural event or an anthropogenic climate change-related event, the phenomenon has highly significant etiological and evolutionary implications (Mueller-Dombois 1987; Kullman 1989).

Schematic representation of the cyclic regeneration of Korean fir by climate anomaly in the montane forests of South Korea
As mentioned, the pattern of CWD distribution was strongly related to the drought event in the winter of 1999, and the expected consequent changes in ecosystem processes are discussed. In-situ and controlled experiments on the effects of soil freezing on the root system of Korean fir are important in explaining the dying events. The retained Korean fir can exhibit greater susceptibility to changed disturbance regimes, such as the increased strength of typhoons. In the progression of climate change, warmer and drier or warmer and wetter conditions are expected in the future, and these changed conditions facilitate biotic (insects and pathogens) and abiotic (drought, wind, and snow) disturbance agents (Seidl et al. 2017). Thus, only long-term field observation can improve our understanding of dieback dynamics and forest persistence or evolution (Boehmer et al. 2013), including approaches to the changes in range size of Korean fir and related vegetation composition. Specifically, understanding the phenotypic plasticity of marginal populations and interactions between species from lower elevations, such as those from the genus Quercus and Prunus, is essential.
Data Availability
The datasets are available from the corresponding author upon reasonable request.
References
Ahn US, Kim DS, Yun YS, Ko SH, Kim KS, Cho IS (2018) The inference about the cause of death of Korean fir in Mt. Halla through the analysis of spatial dying pattern - Proposing the possibility of excess soil moisture by climate changes. Korean J Agric for Meteorol 21:1–28 (in Korean with English abstract)
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J-H, Allard G, Running SW, Semerci A, Cobb N (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259:660–684
Antos JA, Parish R (2002) Structure and dynamics of a nearly steady-state subalpine forest in south-central British Columbia, Canada. Oecologia 130:126–135
Auclair AND, Heilman WE, Brinkman B (2010) Predicting forest dieback in Maine, USA: a simple model based on soil frost and drought. Can J for Res 40:687–702
Barbour M, Burk J, Pitts W, Gilliam F, Schwartz M (1998) Terrestrial plant ecology Benjamin Cummings, ed. Addison Wesley Longman Inc., Menlo Park
Bigler C, Bugmann H (2004) Predicting the Time of Tree Death Using Dendrochronological Data. Ecol Appl 14(3):902–914
Boehmer HJ, Wagner HH, Jacobi JD, Gerrish GC, Mueller-Dombois D (2013) Rebuilding after collapse: evidence for long-term cohort dynamics in the native Hawaiian rain forest. J Veg Sci 24:639–650
Cho DS (1994) Community structure, and size and age distribution of conifers in subalpine Korean fir (Abies koreana) forest in Mt. Chiri. Korean J Ecol 17:415–424
Cho MG, Chung JM, Im HI, Noh I, Kim TW, Kim CY, Moon HS (2016) Ecological characteristics of sub-alpine coniferous forest in Banyabong, Mt. Jiri. J Clim Change Res 7:465–476 (in Korean with English abstract)
Cho YC, Kim NS, Koo BY (2018) Changed land management policy and the emergence of a novel forest ecosystem in South Korea: landscape dynamics in Pohang over 90 years. Ecol Res 33:351–361
Cho YC, Jung S, Lee D-H, Kim H-G, Kim J-H (2020) Forest of Korea (VI) Biogeography: flora and vegetation. Korea National Arboretum, Pochen (in Korean with English title)
Choung YS, Jeon M, Lee K (2015) Gap formation and susceptible Abies trees to windthrow in the forests of Odaesan National Park. J Ecol Env 38:175–183
Chun YM, Kim SM, Park EH, Park SH, Lee HY, Kim JW (2021) Monitoring on the vegetation structure and dynamics of Abies koreana populations in Jirisan National Park. Korean J Environ Ecol 35:408–423 (in Korean with English abstract)
Cleavitt NL, Fahey TJ, Groffman PM, Hardy JP, Henry KS, Driscoll CT (2008) Effects of soil freezing on fine roots in a northern hardwood forest. Can J for Res 38:82–91
Diaci J, Rozenbergar D, Anic I, Mikac S, Saniga M, Kucbel S, Visnjic C, Ballian D (2011) Structural dynamics and synchronous silver fir decline in mixed old-growth mountain forests in Eastern and Southeastern Europe. Forestry 84:479–491
Fukasawa Y, Katsumata S, Mori AS, Osono T, Takeda H (2014) Accumulation and decay dynamics of coarse woody debris in a Japanese old-growth subalpine coniferous forest. Ecol Res 29:257–269
Groffman PM, Hardy JP, Fashu-Kanu S, Driscoll CT, Cleavitt NL, Fahey TJ, Fisk MC (2011) Snow depth, soil freezing and nitrogen cycling in a northern hardwood forest landscape. Biogeochemistry 102:223–238
Harmon ME, Franklin JF, Swanson FJ, Sollins P, Gregory S, Lattin J, Anderson N, Cline S, Aumen N, Sedell J (1986) Ecology of coarse woody debris in temperate ecosystems. Adv Ecol Res 15:133–302
Hong SG, Kim JJ, Cho HK (2008) Studies on natural regeneration of Abies koreana. J Korean Natl Acad Sci 47:71–84 (in Korean with English abstract)
Iwaki H, Totsuka T (1959) Ecological and physiological studies on the vegetation of Mt. Shimagare II. On the crescent-shaped “dead trees strips” in the Yatsugatake and Chichibu Mountains. Bot Mag Tokyo 72:255–260
Jung S, Cho YC (2020) Redefining floristic zones in the Korean Peninsula using high-resolution georeferenced specimen data and self-organizing maps. Ecol Evol 10:11549–11564
Kaňa J, Tahovská K, Kopáček J (2013) Response of soil chemistry to forest dieback after bark beetle infestation. Biogeochemistry 113:369–383
Kang S-J, Kwak A-K, Kikuchi T (1997) A phytosociological description of the Abies koreana forest on Mt. Halla in Cheju Island, Korea. Korean J Ecol Environ 20:293–298
Kassambara A (2021) Rstatix: pipe-friendly framework for basic statistical tests (0.7.0)
Kim GT, Choo GC (2000) Comparison of growth condition of Abies koreana Wilson by districts. Korean J Environ Ecol 14(1):80–87 (in Korean with English abstract)
Kim NS, Lee HC (2013) A study on changes and distributions of Korean fir in sub-alpine zone. J Korean Soc Environ Restor Tech 16:49–57 (in Korean with English abstract)
Kim SD, Yoon WS (1991) Regeneration process of Abies koreana forest in Banya-bong, Mt. Jiri. Research Report of Environment and Technology, Chungnam University 9: 97–106. (in Korean with English abstract)
Kim, ES, Kim SD, Cha YJ (1994) Final report on the decline of tree growth and the changes of environmental factors on high altitude mountains. Korea Science and Engineering Foundation (KOSEF 921–1500–018–2) (in Korean with English abstract)
Kim R, Son Y, Hwang J (2004) Comparison of mass and nutrient dynamics of coarse woody debris between Quercus serrata and Q. variabilis stands in Yangpyeong. Korean J Ecol 27:115–120
Kim RH, Son Y, Lim JH, Lee IK, Seo KW, Koo JW, Noh NJ, Ryu SR, Hong SK, Ihm BS (2006) Coarse woody debris mass and nutrients in forest ecosystems of Korea. Ecol Res 21:819–827
Kohyama T (1982) Studies on the Abies population of Mt. Shimagare II. Reproductive and life history traits. Bot Mag Tokyo 95:167–181
Kohyama T (1984) Regeneration and coexistence of two Abies species dominating subalpine forests in central Japan. Oecologia 62:156–161
Koo KA, Kim DB (2020) Review forty-year studies of Korean fir (Abies koreana Wilson). Korean J Environ Ecol 34:358–371 (in Korean with English abstract)
Koo KA, Park WK, Kong WS (2001) Dendrochronological analysis of Abies koreana W. at Mt. Halla Korea: effects of climate change on the growths. Korean J Ecol 24:281–288 (in Korean with English abstract)
Korea National Arboretum (2014) Forest of Korea: Conservation of Korean fir (Abies koreana) in a changing environment, Ed, Vol I. Sumeungil, Seoul (in Korean with English title)
Korea National Arboretum (2015) Ecology of woody plants in South Korea: Conifers, Ed, Vol I. Sumeungil, Seoul (in Korean with English title)
Körner C, Hiltbrunner E (2021) Why is the alpine flora comparatively robust against climatic warming? Diversity 13:383
Kullman (1989) Cold-induced dieback of montane spruce forests in the Swedish Scandes—a modern analogue of paleoenvironmental processes. New Phytol 113:377–389
Kullman L, Högberg N (1989) Rapid natural decline of upper montane forests in the Swedish Scandes. Arctic 42(3):217–226
Laiho R, Prescott CE (2004) Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: a synthesis. Can J for Res 34:763–777
Lee CS, Cho HJ (1993) Structure and dynamics of Abies koreana Wilson community in Mt. Gaya, Korean. J Ecol Environ 16:75–91 (In Korean with English abstract)
Lee GS, Kim JH (1994) Primary succession on talus area at Mt. Kariwangsan, Korea. J Ecol Environ 17(2):121–130
Marchand PJ, Goulet FL, Harrington TC (1986) Death by attrition: a hypothesis for wave mortality of subalpine Abies balsamea. Can J for Res 16:591–596
Maser C, Trappe J (1984) The seen and unseen world of the fallen tree. USDA Forest Service. Pacific Northwest Forest Experiment Station, General Technical Report PNW-GTR-164
McGregor IR, Helcoski R, Kunert N, Tepley AJ, Gonzalez-Akre EB, Herrmann V, Zailaa J, Stovall AEL, Bourg NA, McShea WJ, Pederson N, Sack L, Anderson-Teixeira KJ (2021) Tree height and leaf drought tolerance traits shape growth responses across droughts in a temperate broadleaf forest. New Phytol 231:601–616
Mueller-Dombois D (1987) Natural dieback in forests. Bioscience 37:575–583
Mueller-Dombois D (1992) A natural dieback theory, cohort senescence as an alternative to the decline disease theory. Forest Decline Concepts. APS Press St. Paul, Minnesota, pp 26–37
Noh I, Chung JM, Kim TW, Tamirat S, Moon HS (2018) The ecological characteristics by slope of an Abies koreana forest in Seseok of Jurisan National Park. J Clim Change Res 9:293–302 (in Korean with English abstract)
O’Brien MJ, Engelbrecht BMJ, Joswig J, Pereyra G, Schuldt B, Jansen S, Kattge J, Landhäusser SM, Levick SR, Preisler Y, Väänänen P, Macinnis-Ng C (2017) A synthesis of tree functional traits related to drought-induced mortality in forests across climatic zones. J Appl Ecol 54:1669–1686
Olajuyigbe SO, Tobin B, Gardiner P, Nieuwenhuis M (2011) Stocks and decay dynamics of above- and belowground coarse woody debris in managed Sitka spruce forests in Ireland. For Ecol Manag 262:1109–1118
Paillet Y, Archaux F, Boulanger V, Debaive N, Fuhr M, Gilg O, Gosselin F, Guilbert E (2017) Snags and large trees drive higher tree microhabitat densities in strict forest reserves. For Ecol Manag 389:176–186
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Sato T, Yamazaki H, Yoshida T (2017) Extending effect of a wind disturbance: mortality of Abies sachalinensis following a strong typhoon in a natural mixed forest. J for Res 22:336–342
Seidl R, Thom D, Kautz M, Martin-Benito D, Peltoniemi M, Vacchiano G, Wild J, Ascoli D, Petr M, Honkaniemi J, Lexer MJ (2017) Forest disturbances under climate change. Nat Clim Change 7:395–402
Seo JW, Choi EB, Park JH, Kim YJ, Lim HI (2021) The Role of Aging and Wind in Inducing Death and/or Growth Reduction in Korean Fir (Abies Koreana Wilson) on Mt. Halla, Korea. Atmosphere 12(9):1135
Sollins P (1982) Input and decay of coarse woody debris in coniferous stands in western Oregon and Washington. Can J for Res 12:18–28
Spies TA, Franklin JF, Thomas TB (1988) Coarse woody debris in Douglas-fir forests of western Oregon and Washington. Ecology 69:1689–1702
Sprugel DG (1976) Dynamic structure of wave-regenerated Abies balsamea forests in the north- eastern United States. J Ecol 64:889–911
Stokland JN (2001) The coarse woody debris profile: an archive of recent forest history and an important biodiversity indicator. Ecol Bull 49:71–83
Suzuki SN, Kachi N, Suzuki J-I (2013) Spatial variation of local stand structure in an Abies forest, 45 years after a large disturbance by the Isewan typhoon. J for Res 18:139–148
Thornthwaite CW (1948) An approach toward a rational classification of climate. Geogr Rev 38:55–94
Wickham H (2016) Ggplot2: Elegant graphics for data analysis, 2nd edn. Springer International Publishing
Yin X (1999) The decay of forest woody debris: numerical modeling and implications based on some 300 data cases from North America. Oecologia 121:81–98
Acknowledgements
The authors would like to thank Seol J and Dr. Jung S for conducting the field survey.
Funding
This study was supported by the Korea National Arboretum (KNA-1–2-43, 23–1).
Author information
Authors and Affiliations
Contributions
HK and YCC conceived the study, designed the methodology, conducted the experiment, and analyzed the data; SL and EK conducted the experiment and analyzed the data. All authors contributed critically to the drafts of the manuscript and approved its submission for publication.
Corresponding author
Supplementary Information
Below is the link to the electronic supplementary material.
12374_2023_9413_MOESM1_ESM.tif
Plate 1. Decline and root systems of Korean fir in the study area, Mt. Jirisan. Pictures were captured by the author at the same locations in 2006 (a, c, and e) and 2015 (b, d, and f). The dieback of Korean fir started before 2006 on Mt. Jirisan. The red arrows in (a) and (b) indicate the same healthy Korean fir individual in the rocky and steep site. The rooting depth of Korean fir (e and f) is shallow, even at locations with deep soil and a gentle slope
12374_2023_9413_MOESM2_ESM.tif
Figure S1. Mean temperature and the highest and lowest temperatures in the dormancy and growing periods in the study area. DP and GP in the legend represent the dormancy and growing periods, respectively. The solid lines indicate simple linear regression for each temperature parameter
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kim, H., Kim, E., Lee, S. et al. Abnormal Winter Drought-Induced Transient Dieback of Korean Fir in the Montane Forests of Mt. Jirisan, South Korea. J. Plant Biol. 67, 123–136 (2024). https://doi.org/10.1007/s12374-023-09413-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12374-023-09413-5